内侧神经节隆起前体细胞的体外产生
1.本技术是申请日为2014年3月14日和发明名称为“内侧神经节隆起前体细胞的体外产生”的201480013159.5号发明专利申请的分案申请。
2.交叉引用
3.本技术主张2013年3月14日提交的美国临时专利申请no.61/783,594的权益,该申请的全文以引用的方式并入文中。
4.关于联邦资助的研究的声明
5.本发明在政府支持下以美国国立卫生研究院授予的批准号mh081880进行。政府对本发明具有某些权利。本发明在支持下以加利福尼亚州再生医学研究所授予的批准号rc1
‑
00346和rb2
‑
01602进行。
6.发明背景
7.抑制性中间神经元占大脑皮层中神经元的约20%。中间神经元的不足与多种神经障碍有关。大多数皮层中间神经元源于大脑的发育中腹侧端脑区的内侧神经节隆起(mge)。
8.小鼠mge移植物已被证实改善神经障碍的多个啮齿动物模型,表明人mge细胞可代表一种独特的治疗候选。
9.但是,用于有效地生成具有mge细胞的特征的细胞的体外方法不可用。
10.如此,需要用于有效地体外生成mge前体细胞及用于在mge前体细胞中富集的细胞群体的方法。
技术实现要素:
11.提供了用于体外生成mge前体细胞及富集的mge前体细胞的组合物的方法和系统。所述方法和系统提供功能性mge前体的有效产生,所述功能性mge前体分化成功能性gaba能中间神经元。
12.提供了一种由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法。
13.在某些实施方案中,所述方法包括在含有音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞以生成mge前体细胞。可以粘附培养或悬浮培养培养pps细胞。
14.在某些实施方案中,所述方法包括在含有音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞以生成拟胚体(eb),其中eb包含mge前体细胞。
15.在某些情况中,神经诱导补充物可为b27。在某些情况中,神经诱导补充物可为ns21。
16.在某些实施方案中,pps细胞可为人多能干(hps)细胞。hps细胞可为人胚胎干(hes)细胞或诱导性多能干(ips)细胞。
17.在某些实施方案中,在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞前,可诱导pps细胞分化。例如,可通过pps细胞培养物的过生长,或在具有低粘附基质的培养容器中悬浮培养pps细胞,在饲养层不存在下培养pps,或在包含
音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞前添加分化因子例如fgf,来诱导pps细胞分化。
18.在某些实施方案中,所述方法可包括分离eb;将分离eb接种到粘附基质上以提供粘附eb;并培养粘附eb。
19.在某些实施方案中,所述方法可包括分离eb;以机械或酶的方式解离eb以产生单细胞或细胞群;将解离细胞接种在粘附基质上以提供粘附单层;以及培养粘附单层。
20.在某些实施方案中,所述方法可包括分离eb;以机械或酶的方式解离eb以产生单细胞或细胞群;将解离细胞接种在细胞饲养层上以提供粘附共培养物;以及培养粘附共培养物。
21.在某些实施方案中,所述方法可包括分离eb、粘附eb、单层、或共培养物;以机械的方式或以酶的方式解离eb、粘附eb、单层、或共培养物以产生单细胞;用mge前体细胞的细胞表面标记物的抗体孵育单细胞;以及分离前体细胞。
22.在某些实施方案中,所述方法可包括分离eb、粘附eb、单层、共培养物、解离培养物、或分离前体细胞;以及添加冷冻保护剂,例如,防冻剂化合物,例如,二醇类(丙三醇、乙二醇、丙二醇)、二甲亚砜(dmso)、或蔗糖。
23.在某些情况中,用于由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法可包括在不含血清的培养基中培养pps细胞以生成拟胚体(eb),其中eb包括mge前体细胞,其中所述不含血清的培养基包括音速刺猬路径的活化剂、rho
‑
相关激酶(rock)的抑制剂、smad的抑制剂、wnt的抑制剂和b27。
24.pps细胞为人多能干(hps),其可为人胚胎干(hes)细胞或诱导性多能干(ips)细胞。
25.在某些情况中,所述方法可进一步包括分离eb;将分离eb接种在粘附基质上以提供粘附eb;以及培养粘附eb。
26.在某些实施方案中,使粘附eb在包含音速刺猬路径的活化剂、smad的抑制剂、wnt的抑制剂、及b27的不含血清的培养基中培养。
27.在某些实施方案中,使粘附eb在不含有rock的抑制剂的不含血清的培养基中培养。
28.提供了一种产生抑制性中间神经元的方法,所述方法可包括分离eb、粘附eb、单层、共培养物、解离培养物、或如上述产生的分类的细胞;产生分离细胞的细胞悬浮液及将细胞悬浮液移植到灵长类动物神经系统中。
29.特别地,本发明涉及以下各项:
30.1.一种由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法,所述方法包括:
31.在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞以生成所述mge前体细胞。
32.2.根据第1项所述的方法,其中所述方法包括在包含所述音速刺猬路径的活化剂和所述神经诱导补充物的不含血清的培养基中培养所述pps细胞以生成拟胚体(eb),其中所述eb包含所述mge前体细胞。
33.3.根据第1
‑
2中任一项所述的方法,其中所述神经诱导补充物包含b27。
34.4.根据第1
‑
2中任一项所述的方法,其中所述神经诱导补充物包含ns21。
35.5.根据第1
‑
4中任一项所述的方法,其中所述pps细胞为人多能干(hps)细胞。
36.6.根据第5项所述的方法,其中所述hps细胞为人胚胎干(hes)细胞。
37.7.根据第5项所述的方法,其中所述hps细胞为诱导性多能干(ips)细胞。
38.8.根据第1
‑
7中任一项所述的方法,其中所述pps细胞以粘附培养进行培养。
39.9.根据第1
‑
7中任一项所述的方法,其中所述pps细胞以悬浮培养进行培养。
40.10.根据第1
‑
9中任一项所述的方法,其中使所述pps细胞诱导分化,然后在包含所述音速刺猬路径的活化剂和所述神经诱导补充物的所述不含血清的培养基中培养所述pps细胞。
41.11.根据第10项所述的方法,其中使所述pps细胞诱导分化,然后通过在饲养层不存在下培养来在包含所述音速刺猬路径的活化剂和所述神经诱导补充物的所述不含血清的培养基中培养所述pps细胞。
42.12.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
43.分离所述eb;
44.将所述分离eb接种在粘附基质上以提供粘附eb;以及
45.培养所述粘附eb。
46.13.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
47.分离所述eb;
48.以机械或酶的方式解离所述eb以产生单细胞或细胞群;
49.将所述解离细胞接种在粘附基质上以提供粘附单层;以及
50.培养所述粘附单层。
51.14.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
52.分离所述eb;
53.以机械或酶的方式解离所述eb以产生单细胞或细胞群;
54.将所述解离细胞接种在细胞饲养层上以提供粘附共培养物;以及
55.培养所述粘附共培养物。
56.15.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
57.分离所述eb、所述粘附eb、所述单层、或所述共培养物;
58.以机械或酶的方式解离所述eb、所述粘附eb、所述单层、或所述共培养物以产生单细胞;
59.将单细胞与mge前体细胞的细胞表面标记物的抗体一起孵育;以及
60.分离所述前体细胞。
61.16.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
62.分离所述eb、所述粘附eb、所述单层、所述共培养物、所述解离培养物、或所述分类细胞;以及
63.添加冷冻保护剂。
64.17.根据第2
‑
11中任一项所述的方法,其中所述方法包括:
65.分离所述eb、所述粘附eb、所述单层、所述共培养物、所述解离培养物、或所述分类细胞;以及
66.将细胞悬浮液移植到所述灵长类动物神经系统中。
67.附图简述
68.图1(图版a
‑
b)说明了mge
‑
样前体细胞的生成。
69.图2(图版a
‑
f)说明了hesc
‑
mge
‑
样先祖细胞表现出vz和svz放射状胶质干细胞
‑
样分裂。
70.图3(图版a
‑
e)说明了hesc
‑
mge
‑
样祖细胞分化成具有端脑gaba能中间神经元的性质的神经元。
71.图4(图版a
‑
h)描述了hesc
‑
mge
‑
样nkx2.1
‑
gfp 细胞群体的微阵列基因表达谱。
72.图5(图版a
‑
j)说明了hesc
‑
mge
‑
样细胞源性的gaba能中间神经元成熟和放电性质。
73.图6(图版a
‑
j)说明了hesc源性的中间神经元的gaba能突触性质。
74.图7(图版a
‑
h)表明了在小鼠脑中的hesc源性的mge
‑
样中间神经元前体细胞成熟和功能整合。
75.图8(图版a
‑
f)提供了分化方案以及分化的hesc的facs分析的示意图。
76.图9说明了hesc源性的细胞具有端脑mge
‑
样同一性和gaba能神经元命运。
77.图10(图版a
‑
f)描述了hesc源性的nkx2.1
‑
gfp 细胞的转录表达谱。
78.图11描述了hesc源性的mge
‑
样细胞成熟为gaba能中间神经元亚型。
79.图12(图版a
‑
c)显示了中间神经元亚型在源自mge的培养物中的发育。
80.图13(图版a
‑
f)显示了hesc源性的中间神经元放电性的成熟。
81.图14(图版a
‑
g)描述了在小鼠脑中hesc源性的mge
‑
样中间神经元和亚型放电性的成熟。
82.图15提供了对在自hesc分化期间标记物表达的总结。
83.图16提供了对hesc分化方案优化、动物移植、和肿瘤发病率的总结。
84.图17描述了自hesc系esi17体外分化的mge前体细胞。
85.图18描述了自hesc系esi35体外分化的mge前体细胞。
86.图19描述了自hesc系esi51体外分化的mge前体细胞。
87.图20描述了自hesc系h9体外分化的mge前体细胞。
88.图21说明了通过天然人多能干细胞的分化生成mge前体细胞。
89.图22(图版a
‑
n)说明了富集mge的增强子序列用于源自通过hpsc的分化生成的mge前体细胞的中间神经元的筛选和纯化的用途。
90.图23(行a
‑
d)描述了利用长期悬浮培养生成mge源性的中间神经元。
91.图24(图版a
‑
e)说明了bmp和wnt信号传导路径的多种小分子抑制剂有效地诱导hesc分化成mge前体细胞。
92.定义
93.如文中所用,“拟胚体”、“eb”或“eb细胞”通常指由源自经历过分化的多能干细胞(例如,灵长类动物多能干细胞(pps)、胚胎干(es)细胞、诱导性多能干(ips)细胞)的未分化和分化细胞的群体组成的形态、三维、或类器官型结构。在适于eb形成的培养条件下,es细胞增殖并形成开始分化的小块细胞。在分化的第一阶段中,通常相当于人细胞分化的大概第1
‑
4天,小块细胞在外层上形成内胚层细胞层,并被视为一种“简单的拟胚体”。在第二阶
段中,通常相当于人细胞分化后的大概第3
‑
20天,形成“复杂的拟胚体”,其特征在于外胚层和中胚层细胞及衍生组织的广泛分化。如文中所用,术语“拟胚体”或“eb”涵盖简单的和复杂的拟胚体,除非上下文另外要求。确定拟胚体何时在es/ips细胞培养物中形成通常由本领域技术人员例如通过目测形态、检测细胞标记物进行。约20个或更多细胞(例如,es/ips细胞)的浮动块被视为悬浮拟胚体(seb)(参见,例如,schmitt,r.,等(1991)genes dev.5:728
‑
740;doetschman,t.c.,等(1985)j.embryol.exp.morph.87:27
‑
45)。可将悬浮eb接种在粘附基质上以生成粘附eb(aeb)。
94.如文中所用,“内侧神经节隆起(mge)前体细胞”或“mge神经前体细胞”指表达由发育脑的mge区中细胞所表达的标记物的有丝分裂和有丝分裂后细胞的群体。一般来讲,mge前体细胞表达标记物,如同源异型盒基因nkx2.1、lim
‑
同源异型盒基因lhx6、lhx7、或lhx8。mge前体细胞可在适宜的分化条件下分化成中间神经元。
[0095]“多能干细胞”或“多能细胞”意指一种在适宜条件下具有产生身为三个所有胚芽层(外胚层、中胚层和内胚层)的衍生物的多种不同细胞类型的后代的能力的细胞。多能干细胞能够形成畸胎瘤。多能干细胞的实例为胚胎干(es)细胞、胚胎生殖干(eg)细胞、胚胎癌干(ec)细胞、和诱导性多能干(ips)细胞。ps细胞可来自于所关注的任何生物体,包括,例如,人;灵长类动物;非
‑
人灵长类动物;狗;猫;鼠;马;猪;鸟;骆驼;牛;羊等。
[0096]“胚胎干细胞”或“es细胞”意指一种a)可自我更新,b)可分化产生生物体中的多种细胞,以及c)源自发育生物体或为源自发育生物体的确立的es细胞系的细胞。es细胞可源自发育生物体的囊胚的内细胞团、或源自发育生物体的外胚层。es细胞可源自涉及从发育生物体中去除单卵裂球的由单卵裂球活检(sbb)产生的卵裂球。一般来讲,sbb提供内细胞团分离的非破坏性替代项。sbb和自活检卵裂球的hes细胞的生成在cell stem cell,2008年2月7日;2(2):113
‑
7中有描述。可使es细胞在长时间内培养同时保持在生物体内分化成所有种类的细胞的能力。在培养中,es细胞一般以大核质比率,限定的边界和突起的核仁呈平板菌落生长。此外,hes细胞表达ssea
‑
3、ssea
‑
4、tra
‑1‑
60、tra
‑1‑
81、和碱性磷酸酶,但不表达ssea
‑
1。生成及表征es细胞的方法的实例可参见,例如,美国专利号7,029,913、美国专利号5,843,780、和美国专利号6,200,806,其公开内容以引用的方式并入文中。es细胞的实例包括天然es细胞。
[0097]“胚胎生殖干细胞”、“胚胎生殖细胞”或“eg细胞”意指一种a)可自我更新,b)可分化产生生物体中的多种细胞,以及c)源自生殖细胞及生殖细胞先祖细胞,例如原始生殖细胞,即会变成精子和卵的那些细胞。胚胎生殖细胞(eg细胞)被认为具有类似于如上所述的胚胎干细胞的性质。生成及表征eg细胞的方法的实例可参见,例如,美国专利号7,153,684;matsui,y.,等,(1992)cell 70:841;shamblott,m.,等(2001)proc.natl.acad.sci.usa 98:113;shamblott,m.,等(1998)proc.natl.acad.sci.usa,95:13726;以及koshimizu,u.,等(1996)development,122:1235,其公开内容以引用的方式并入文中。
[0098]“诱导性多能干细胞”或“ips细胞”意指一种a)可自我更新,b)可分化产生生物体中的多种细胞,以及c)源自体细胞的细胞。ips细胞具有es细胞
‑
样形态,以大核质比率,限定的边界和突起的核仁呈平板菌落生长。此外,ips细胞表达本领域技术人员已知的一种或多种关键的多能性标记物,包括但不限于碱性磷酸酶、ssea3、ssea4、sox2、oct3/4、nanog、tra160、tra181、tdgf 1、dnmt3b、foxd3、gdf3、cyp26a1、tert、和zfp42。ips细胞可通过提供
细胞“重编程因子”生成,即,作用于细胞以改变转录,进而重编程细胞为多能性的生物活性因子的一种或多种,例如混合物。生成及表征ips细胞的方法的实例可参见,例如,申请号us20090047263、us20090068742、us20090191159、us20090227032、us20090246875、和us20090304646,其公开内容以引用的方式并入文中。
[0099]“体细胞”意指在试验操作不存在下,一般在生物体中不产生所有种类的细胞的生物体中的任何细胞。换句话说,体细胞为充分分化使之不能自然地生成体内三个所有生殖层(即,外胚层、中胚层和内胚层)的细胞的细胞。例如,体细胞可包括神经元和神经先祖细胞,其中后者可自我更新并自然地产生所有或一些细胞类型的中枢神经系统但不能产生中胚层或内胚层谱系的细胞。
[0100]
术语“细胞系”指一般源自单祖细胞或源自祖细胞的定义的和/或基本上相同的群体的大体上或基本上相同的细胞的群体。细胞系已被或可以保持在培养状态一段较长时间(例如,月、年、一个不限定的时间段)。
[0101]“内胚层”意指在动物胚胎形成期间形成的产生胃肠道、呼吸道、内分泌腺和器官、听觉系统的某些结构、以及泌尿系统的某些结构的生殖层。
[0102]“中胚层“意指在动物胚胎形成期间形成的产生肌肉、软骨、骨、真皮、生殖系统、脂肪组织、肠的结缔组织、腹膜、泌尿系统的某些结构、间皮、脊索、和脾的生殖层。
[0103]“外胚层”意指在动物胚胎形成期间形成的产生神经系统、牙釉质、表皮、毛发、指甲、和粘膜组织的衬里的生殖层。
[0104]“骨形态形成蛋白”或“bmp”意指身为转化生长因子β(tgfβ)超家族的亚家族的生长因子的家族。bmp(例如bmp1、bmp2、bmp3、bmp4、bmp5、bmp6、bmp7、bmp8a、bmp8b、bmp9/gdf、bmp10、bmp11/gdf11、bmp12/gdf7、bmp13/gdf6、bmp14/gdf5、bmp15/gdf9b)首先因其诱导骨和软骨形成的能力被发现。bmp与细胞表面上的特异性受体相互作用,所述受体被称作骨形态形成蛋白受体(bmpr)。通过bmpr的信号转导导致蛋白的smad家族的成员的动员,继而调控目的基因的转录。bmp信号传导的抑制剂易由本领域普通技术人员通过多种方法中的任一种识别,例如,用于结合bmp或bmp受体的竞争性结合测定,功能测定,例如,测定下游信号传导蛋白的活性的提高,例如smad,如,br
‑
smad至细胞核的重定位以及如本领域所知的下游基因目标的转录激活。
[0105]“转化生长因子β”、“tgf
‑
β”、及“tgfb”意指属于转化生长因子β(tgfβ)超家族的亚家族的tgfb分泌蛋白。tgfb(tgfb1、tgfb2、tgfb3)为调节增殖、分化、粘附、和迁移并在多种细胞中的多功能肽。成熟肽可以同型二聚体或以与其它tgfb家族成员的异质二聚体出现。tgfb与细胞表面上的转化生长因子β受体(tgf
‑
βrs,或tgfbr)相互作用,其结合激活map激酶
‑
、akt
‑
、rho
‑
和rac/cdc42
‑
定向的信号转导途径,smad蛋白的细胞结构和核定位的重组,和目的基因转录的调控。tgfb信号传导的抑制剂易由本领域普通技术人员通过多种方法中的任一种识别,例如,用于结合tgfb或tgfb受体的竞争性结合测定,或功能测定,例如,对下游信号传导蛋白如mapk、akt、rho、rac、和smad,例如,ar
‑
smad等的活性的抑制的测量,如本领域中熟知。
[0106]“wnt”意指在胚胎形成和成熟组织中发挥关键作用的高度保守的分泌型信号传导分子的家族。人wnt基因家族具有至少19个成员(wnt
‑
1、wnt
‑
2、wnt
‑
2b/wnt
‑
13、wnt
‑
3、wnt3a、wnt
‑
4、wnt
‑
5a、wnt
‑
5b、wnt
‑
6、wnt
‑
7a、wnt
‑
7b、wnt
‑
8a、wnt
‑
8b、wnt
‑
9a/wnt
‑
14、
wnt
‑
9b/wnt
‑
15、wnt
‑
10a、wnt
‑
10b、wnt
‑
11、wnt
‑
16)。wnt蛋白通过结合wnt受体复合物调节细胞活性,所述wnt受体复合物包括来自蛋白的frizzled(fz)家族的多肽和蛋白的低密度脂蛋白受体(ldlr)
‑
相关蛋白(lrp)的多肽。一旦被wnt结合激活,wnt受体复合物将激活一种或多种细胞内信号传导级联反应。这些包括经典wnt信号传导路径;wnt/平面细胞极化(wnt/pcp)路径;和wnt
‑
钙(wnt/ca2 )路径。
[0107]
在“非粘附条件”下培养意指在抑制细胞粘附到在其内它们被培养的容器例如组织培养板或烧瓶底部的条件下培养。在一些情况中,细胞是自然地非粘附型,即它们不会粘附表面除非表面涂布了基体组合物,例如纤维连接蛋白、层粘连蛋白、多聚鸟氨酸、多聚赖氨酸、胶原蛋白iv、基质胶、和聚碳酸酯膜。在一些情况中,可通过搅拌培养物使细胞维持在非粘附状态。
[0108]
在“粘附条件”下培养意指在促进细胞粘附到在其内它们被培养的容器例如组织培养板或烧瓶底部的条件下培养。在一些情况中,只通过保持培养基静止就可使细胞粘附到容器。在一些情况中,可使需促进对其粘附性的容器壁涂布有细胞可粘附的组合物,例如纤维连接蛋白、层粘连蛋白、多聚鸟氨酸、多聚赖氨酸、胶原蛋白iv、基质胶、和聚碳酸酯膜。
[0109]
术语“处理”在文中一般用于意指获得所需的药理和/或生理效果。效果可就完全地或部分地阻止疾病或其症状而言具预防性和/或就部分地或完全地治愈疾病和/或可归因于疾病的副作用而言具有治疗性。如文中所用“处理”覆盖对哺乳动物的疾病的任何处理,且包括:(a)阻止疾病在倾向于患有该疾病但尚未诊断出患有的受试者中发生;(b)抑制疾病,即,阻止其发展;或(c)缓解疾病,即,使疾病退行。治疗剂可在疾病或损伤发作前、期间或之后施用。对进行中疾病的处理受到特别关注,其中处理稳定或减少患者所不期望的临床症状。此类处理需在感染组织的功能完全损失前进行。受试者治疗需在疾病的症状期间施用,在一些情况中在疾病的症状期后施用。
[0110]
术语“个体”、“受试者”、“宿主”、和“患者”在文中可交换使用并指需要诊断、处理、或治疗的任何哺乳动物受试者,尤其指人。
[0111]
在细胞培养的上下文中的术语“培养基”或短语“细胞培养基(cell culture medium)”或“细胞培养基(cell medium)”指适于培养所关注的细胞群体的细胞生长培养基。细胞培养基的实例包括最低必需培养基(mem)、伊格尔氏培养基、达尔伯克氏改良伊格尔培养基(dmem)、达尔伯克氏改良伊格尔培养基:营养混合物f
‑
12(dmem/f12)、f10营养混合物、ham f10营养混合物、ham f12营养混合物、培养基199、rpmi、rpmi1640、低血清培养基、基础培养基(bme)、dmem/f12(1:1)、neurobasal培养基等,以及其组合。可通过添加一种或多种因子,如,补充物、分化因子、抗凋亡剂修饰培养基或细胞培养基。
[0112]
在细胞或细胞群体的上下文中的术语“分离的”指在不同于其天然环境的环境中的细胞,如,除了有机体的组织。
[0113]
如文中所用,短语“分化因子”指包括在用于培养本发明细胞的培养基中的制剂,该制剂促进细胞从第一细胞类型分化到第二细胞类型,其中相比于第一细胞类型,第二细胞类型被分化。
[0114]
在细胞个体发生的上下文中,形容词“分化的”是相对术语。“分化细胞”是比所比较的细胞沿发育途径进一步进展的细胞。因此,多能胚胎干细胞可分化成谱系限制性前体细胞。这些可继而进一步沿途径进一步分化成细胞,或末期分化细胞,如gaba能中间神经
元。
[0115]“饲养细胞”或“饲养物”为用于描述与另一种细胞共培养的一种细胞的术语,以提供第二种细胞可生长的环境。如果细胞在分裂后已经生长了至少一轮,其中没有添加新鲜饲养细胞以支持pps细胞的生长,则pps细胞群体被视为“基本上不含”饲养细胞。
[0116]
如文中所用,在标记物的上下文中,“表达”以及其语法等效项指标记物的产生以及标记物水平或量。例如,标记物的表达或标记物在细胞中的存在或细胞对标记物而言呈阳性,指标记物的表达水平类似于阳性对照水平。阳性对照水平可通过由已知具有标记物相关的细胞命运的细胞表达的标记物的水平确定。类似地,标记物表达的不存在或细胞对于标记物而言呈阴性,指标记物表达水平类似于阴性对照水平。阴性对照水平可通过由已知不具有标记物相关的细胞命运的细胞表达的标记物的水平确定。如此,标记物的不存在不只是简单地暗示标记物表达的不可检测水平,在某些情况中,细胞可表达标记物但表达低于阳性对照或细胞水平可类似于阴性对照。
[0117]
如文中所用,“标记物”指可测定或检测的任何分子。例如,标记物可包括,不限于,核酸,如,基因的转录物、基因的多肽产物、糖蛋白、糖类、糖脂类、脂质、脂蛋白、糖类、或小分子(例如,具有小于10,000amu的分子量的分子)。
[0118]“变体”多肽意指如下定义的与天然序列多肽具有至少70%、75%、80%、85%、90%、95%、98%、或99%序列同一性的生物活性多肽。该变体包含这样的多肽,其中将一个或多个氨基酸残基添加到天然序列n
‑
或c
‑
端或内部;约一个至40个氨基酸残基被删除,以及任选地被一个或多个氨基酸残基替代;以及以上多肽的衍生物,其中一个氨基酸残基已经被共价修饰以使所得产物具有非天然存在的氨基酸。一般,生物活性变体将具有与天然序列多肽具有至少约90%,至少约95%,或至少约99%氨基酸序列同一性的氨基酸序列。变体多肽可为天然或非天然糖基化,即,多肽具有不同于对应的天然存在的蛋白中所见的糖基化模式的糖基化模式。变体多肽可具有在天然多肽中见不到的翻译后修饰。
[0119]
术语“富集有”或“富集的”在文中可交换使用并意指一种类型的细胞的得率(分率)比起始培养物或制剂中的那种类型的细胞的分率增加至少10%。
[0120]“生长环境”指所关注细胞将体外增殖、分化、或成熟的环境。该环境的特征包括培养细胞的培养基,可存在的任何生长因子或分化
‑
诱导因子,以及支撑结构(如固体表面上的基质)(如果存在)。
具体实施方式
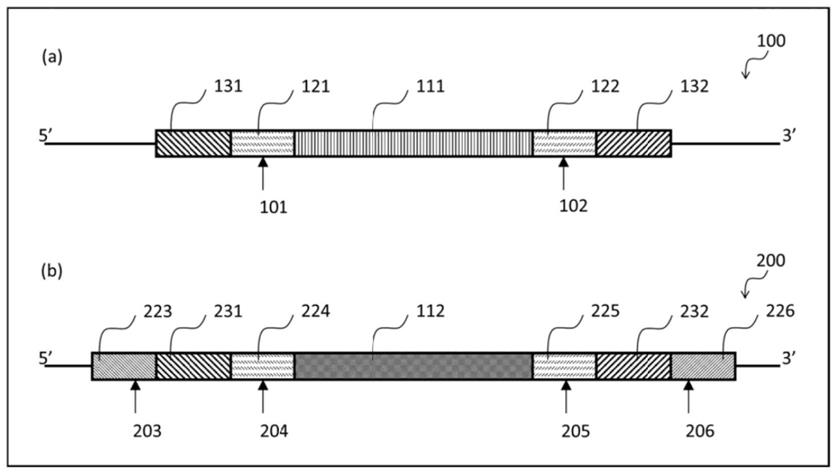
[0121]
如上所述,提供了用于体外生成mge前体细胞的方法和系统以及富集mge前体细胞的组合物。所述方法和系统提供功能性mge前体的有效产生,该功能性mge前体分化成功能性gaba能中间神经元。
[0122]
在对本发明进一步描述前,应了解本发明不限于所述的具体实施方案,因此理所当然可变化。也应了解文中所用的术语只是用于描述具体实施方案,而无意限制,因为本发明范围只受限于随附的权利要求。
[0123]
提供数值范围时,应了解,在该范围的上限和下限之间的每个居间值(除非上下文明确另外指出,否则达到下限的单位的十分之一),与所述范围中任何其它所述值或居间值均包含在本发明内。这些较小范围的上限和下限可独立地包含在这些较小范围内,并也包
含在本发明内,遵守所述范围中的任何特定排除的限值。当所述范围包括一个或两个限值时,排除那些所包括的限制的范围也包括在本发明中。
[0124]
除非另外定义,否则文中所用的所有技术和科学术语具有与本发明所属领域的普通技术人员通常理解的相同含义。虽然类似于或等效于文中所述的那些内容的任意方法和材料也可用于实施或测试本发明,但现在描述优选的方法和材料。文中所述的所有公布以引用的方式并入文中以公开及描述与公布引述相关的方法和/或材料。
[0125]
必须注意,如文中所用及在随附权利要求中,单数形式“一”及“所述”包括复数个指示物,除非上下文明确另外指出。因此,例如,关于“smad抑制剂”的引述包括复数个此类抑制剂并且关于“rock抑制剂”的引述包括对一种或多种rock抑制剂以及本领域技术人员已知的其等效物的引述,以此类推。应进一步注意到,可草拟权利要求以排除任何任选的元素。如此,此陈述意图用作用于与权利要求元素陈述内容相关的如“仅”、“只”等的排他性术语,或用于“否定式”限制的前述基础。
[0126]
文中所讨论的公布只在本技术的提交日期前提供其公开内容。文中内容没有被视为允许本发明无权通过先前发明先于该公布。此外,所提供公布的日期可不同于需独立地确认的实际公布日期。
[0127]
生成mge前体细胞的方法
[0128]
在某些实施方案中,提供了一种由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)的方法。
[0129]
一般来讲,pps保持在未分化状态直到开始启用用于产生mge前体细胞的方法。
[0130]
所述方法可包括在含有音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞以生成mge前体细胞。pps细胞可以粘附培养或悬浮培养进行培养。
[0131]
在某些实施方案中,在用于产生mge前体细胞的方法开始时,将pps细胞接种到具有利于pps细胞附着的粘附基质的细胞培养容器中并使细胞与含有音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基接触以生成mge前体细胞。
[0132]
在某些实施方案中,所述方法可包括使pps细胞在含有音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养以生成拟胚体(eb),其中eb包含mge前体细胞。
[0133]
在某些实施方案中,在用于产生mge前体细胞的方法开始时,可将pps细胞以悬浮方式接种在具有低粘附性基质的培养容器中以允许悬浮拟胚体形成。在示例性方法中,收集pps细胞的汇合单层培养物,然后接种在非粘附细胞培养板上,保持细胞悬浮。
[0134]
在某些情况中,胶原酶iv/分散酶可用于优先地筛选pps菌落。可使菌落胰蛋白酶化成单细胞并接种到低附着圆底平板上以形成悬浮eb。
[0135]
在某些情况中,分化过程可通过使pps细胞分化,例如以形成拟胚体或聚集物:例如,通过供体pps细胞培养物的过生长,或通过在具有低粘附性基质的培养容器中悬浮培养pps细胞以允许拟胚体形成,或在饲养层不存在下培养pps而引起。在示例性方法中,收集pps细胞的汇合单层培养物,然后接种在非粘附细胞培养板上,保持细胞悬浮,并利用营养培养基提供正常饲养。
[0136]
替换地或另外地,可通过与阻止细胞维持未分化显型的某些因子一起培养来开始分化过程。初始分化因子不需限制向mge前体细胞谱系的分化,但应包含在分化群体中的细胞类型范围内的mge前体细胞或它们的前体。
[0137]
在某个阶段,培养可更具体地针对于mge前体细胞谱系。这可通过在培养基中包含更具体地促进mge前体细胞的生成和增殖的因子而实现。促进mge前体细胞的形成和/或生长的示例性因子包括如文中提供的神经诱导补充物,shh信号传导的活化剂、bmp
‑
信号传导的抑制剂、tgf
‑
β信号传导的抑制剂、wnt抑制剂、和抗
‑
凋亡剂,并且在一些情况中,可包括fgf信号传导的活化剂。
[0138]
生成mge前体细胞的示例性方法如下所述。
[0139]
在某些情况中,所述方法可包括生成seb的步骤,随后的生成aeb的步骤。在其它情况中,生成seb的步骤可被粘附培养替代。
[0140]
悬浮拟胚体(seb)的生成
[0141]
在示例性方法中,在具有低粘附性基质的培养容器中悬浮培养pps细胞以允许形成悬浮拟胚体可在shh的活化剂和神经诱导补充物例如b27或ns21存在下进行。pps细胞可在饲料层不存在下悬浮培养0天
‑
9天,然后将shh的活化剂和/或神经诱导补充物加入培养基中,例如,pps细胞可悬浮培养至少0小时、1小时、3小时、6小时、12小时、18小时、24小时、36小时、48小时、2天、3天、4天、5天、6天、7天、8天、或9天,然后将shh的活化剂和/或神经诱导补充物加入培养基中。因此,在文中所述的神经诱导补充物和shh信号传导的活化剂存在下,使pps诱导形成seb。
[0142]
可使pps细胞在文中所述的神经诱导补充物和shh信号传导的活化剂存在下悬浮培养至少一天,例如,1
‑
100天、1
‑
60天、1
‑
50天、2
‑
100天、2
‑
50天、3
‑
100天、4
‑
100天、5
‑
10天、或7
‑
10天、或25
‑
100天以形成seb。在一些情况中,其中可使pps细胞悬浮培养少于9天的时间以形成seb,可将shh的活化剂和/或神经诱导补充物在自开始培养pps细胞以形成seb的0
‑
8天内加入培养基中。
[0143]
在某些实施方案中,将pps悬浮接种在具有低粘附性基质的培养容器中,在包含如文中提供的神经诱导补充物和shh信号传导的活化剂的细胞培养基中。
[0144]
除了文中所提供的神经诱导补充物和shh信号传导的活化剂,用于悬浮培养pps细胞以形成seb的培养基可包含抗凋亡剂、smad抑制剂(例如,tgf
‑
β抑制剂、bmp抑制剂、activin抑制剂、nodal抑制剂、或生长分化因子(gdf)信号传导路径抑制剂)、和wnt抑制剂中的一种或多种。
[0145]
在某些情况中,用于由pps细胞产生mge前体细胞的方法可包括在包括抗凋亡剂例如rock抑制剂的培养基中培养pps细胞约1小时
‑
35天,例如,至少1小时、至少3小时、至少10小时、至少24小时、至少36小时、至少48小时、至少2天、至少3天、如,4天、5天、6天、7天、8天、9天、15天、20天、25天、或35天。示例性方法可包括将pps细胞悬浮接种在包含抗凋亡剂的培养基中,在抗凋亡剂存在下培养pps细胞1小时
‑
35天。在某些情况中,抗凋亡剂可在开始悬浮培养pps细胞时存在并可在1小时
‑
35天,例如,1天至7天,例如,1天、2天、3天、4天、5天、6天、或7天后去除。
[0146]
在某些情况中,抗凋亡剂可在pps分化成mge细胞期间暂时地存在,例如,抗凋亡剂可在pps细胞暴露于文中所提供的神经诱导补充物和shh信号传导的活化剂时第一天存在于培养基中。pps细胞的分化可在文中所提供的神经诱导补充物、shh信号传导的活化剂、以及抗凋亡剂存在下进行1小时至35天,如上所述,然后可在抗凋亡剂不存在下继续培养。
[0147]
在某些情况中,用于由pps细胞产生mge前体细胞的方法可包括在包含wnt的一种
或多种抑制剂的培养基中培养pps细胞。虽然在开始培养pps细胞时可将wnt信号抑制剂加入培养基中,但在培养多天(例如,培养10天内的某一时间)后可将它加入培养基中。在某些情况中,在开始悬浮培养pps细胞的5天内的某一时间将wnt信号抑制剂加入培养基中,例如,在0天、1天、或3天内。wnt抑制剂可在整个seb生成步骤中存在或可存在5天
‑
10天,例如,5天、6天、7天、8天、9天、或10天,然后在wnt抑制剂不存在下可继续培养。
[0148]
在某些情况中,用于由pps细胞产生mge前体细胞的方法可包括在包括smad的一种或多种抑制剂的培养基中培养pps细胞。虽然在开始培养pps细胞时可将smad信号抑制剂加入培养基中,但在培养多天(例如,培养10天内的某一时间)后可将它加入培养基中。在某些情况中,在开始悬浮培养pps细胞的5天内的某一时间将一种或多种smad信号抑制剂加入培养基中,例如,0天、1天、或3天内。wnt抑制剂可在整个seb生成步骤中存在或可存在5天
‑
10天。
[0149]
在某些情况中,pps在shh活化剂、文中所提供的神经诱导补充物、和smad抑制剂存在下分化5至15天(例如,5
‑
10天,例如,5天、6、天、7天、8天、9天、10天、12天),然后在smad抑制剂不存在下可继续分化。
[0150]
在某些实施方案中,由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法可包括在不含血清的培养基中培养pps细胞以生成seb,其中seb包括mge前体细胞,其中不含血清的培养基包括音速刺猬路径的活化剂、抗凋亡剂、smad的抑制剂、wnt的抑制剂和b27。由文中所述的方法产生的seb包括mge前体细胞的群体。
[0151]
在某些实施方案中,由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法可包括在不含血清的培养基中以悬浮培养来培养pps细胞以生成mge前体细胞,其中不含血清的培养基包括音速刺猬路径的活化剂、抗凋亡剂、smad的抑制剂、wnt的抑制剂和b27。在某些情况中,可使seb解离并以单层接种以生成包括mge前体细胞的单层。
[0152]
在某些情况中,自pps细胞开始分化起的约5天后,可使seb解离并以单层接种。例如,在seb形成后的1
‑
100天内,例如,1
‑
75天、1
‑
50天、1
‑
30天、1
‑
10天,可使seb解离并以单层接种。在示例性情况中,在seb形成的约10天后,例如,10
‑
50天、10
‑
40天、10
‑
30天、10
‑
20天、10天、12天等,可使seb解离并以单层接种。所用的分化因子和添加剂、补充物、或因子以及其添加/去除的发生时间可如上公开。
[0153]
在某些实施方案中,由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法可包括在不含血清的培养基中以粘附培养来培养pps细胞以生成mge前体细胞,其中不含血清的培养基包括音速刺猬路径的活化剂、抗凋亡剂、smad的抑制剂、wnt的抑制剂和b27。
[0154]
一般来讲,由文中所述的方法产生的mge前体细胞表达mge前体细胞的标记物,如,nkx2.1。
[0155]
在某些情况中,在开始培养生成mge前体细胞时的ps细胞是以103至107个细胞/ml的细胞密度存在。
[0156]
悬浮培养中所用的培养基可利用任何基础培养基制得。培养基可为bme培养基、bgjb培养基、cmrl 1066培养基、glasgow mem培养基、改良的mem锌选项培养基、imdm培养基、medium 199培养基、伊格尔氏mem培养基、dmem培养基、ham培养基、rpmi 1640培养基、fischer培养基、neurobasal培养基、以及其混合培养基等。可通过添加如文中所公开的添
加剂、补充物、或因子来修饰培养基。
[0157]
具有粘附基质的细胞培养容器可用于以粘附培养来培养pps的方法中。所用的分化因子及添加剂、补充物、或因子以及其添加/去除的发生时间可如上所公开。
[0158]
粘附拟胚体(aeb)的生成
[0159]
在某些情况中,可将由上述方法生成的seb接种到具有利于seb附着以形成粘附eb的粘附基质的细胞培养容器中。一般来讲,可将seb接种到在包含如文中提供的神经诱导补充物和shh信号传导的活化剂的培养基中具有粘附基质的细胞培养容器中。
[0160]
在以粘附培养来培养pps细胞的实施方案中,如上所述,所述方法可在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中进一步培养pps细胞以生成aeb,该aeb包括mge前体细胞。
[0161]
可将重新接种并以粘附培养进行培养以形成aeb的seb培养1
‑
100天。在示例性方法中,seb的重新接种可涉及以下步骤:以机械或酶的方式解离eb以产生单细胞或细胞群,将解离细胞接种到粘附基质以提供粘附单层;并培养粘附单层以生成aeb。在某些方法中,在粘附条件下进一步培养前,不使seb解离。
[0162]
在某些实施方案中,用于由pps细胞产生mge前体细胞的方法可包括在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞以生成seb并将seb接种到具有粘附基质的细胞培养容器并在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中的粘附基质上培养接种的seb以生成aeb,其中aeb包括mge前体细胞。
[0163]
在某些情况中,可使aeb解离并以单层重新接种,使该单层在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养以生成mge前体细胞。
[0164]
用于粘附培养的培养以由seb生成aeb的细胞培养基也可包括一种或多种因子,例如抗凋亡剂、smad的抑制剂、及wnt的抑制剂。因子可在粘附培养开始时存在或在粘附培养开始的5天内添加,如,自粘附培养开始起的0小时、1小时、3小时、1 0小时、1天、2天、或3天。可在培养1天至20天后,将因子从粘附培养基中移除。
[0165]
在某些实施方案中,用于由pps细胞生成mge前体细胞的方法可包括在包含音速刺猬路径的活化剂、神经诱导补充物、wnt和smad的抑制剂的培养基中以粘附培养来培养通过文中所述方法得到的seb的细胞4
‑
20天,然后在包含音速刺猬路径的活化剂和神经诱导补充物但不包含wnt和smad的抑制剂的培养基中以粘附培养来培养8
‑
20天。
[0166]
通过文中所述的方法生成的aeb包含mge前体细胞的群体。一般来讲,通过文中所述的方法产生的aeb中存在的mge前体细胞表达nkx2.1和foxg1。在某些情况中,通过文中所公开的方法产生的mge前体细胞可表达mge前体细胞的一种或多种标记物,如,nkx2.1、lhx6、lhx7/8、foxg1、olig2、dlx1/2、和ascl1。
[0167]
在某些实施方案中,通过文中所述的方法产生的mge前体细胞也可包含由mge前体细胞分化的细胞的群体,如,中间神经元,例如,gaba能中间神经元。
[0168]
一般来讲,文中所述的方法导致在高效率下生成mge前体细胞,得到细胞培养物中至少50%(例如,65%、70%、75%、80%、85%、90%、或95%)的细胞为mge前体细胞的细胞培养物。
[0169]
如此,所述方法可包括在包含音速刺猬路径的活化剂和神经诱导补充物的不含血
清的培养基中培养pps细胞10
‑
100天(例如,5
‑
50天)以生成mge前体细胞,其中pps细胞以粘附培养或悬浮培养进行培养,其中使pps细胞诱导分化,然后在音速刺猬路径的活化剂和神经诱导补充物存在下培养。如此,pps细胞可包含分化细胞,如eb,然后在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养pps细胞。在某些情况中,不含血清的培养基可另外包括抗凋亡剂,smad的抑制剂,和wnt的抑制剂。
[0170]
在某些情况中,可将由seb获得的aeb以悬浮培养重新接种以形成seb或解离或以粘附培养呈单层重新接种。
[0171]
以下进一步描述在用于生成mge前体细胞的方法中以粘附培养来培养pps细胞。
[0172]
mge前体细胞的生成的粘附培养
[0173]
如上所述,在某些实施方案中,在用于产生mge前体细胞的方法开始时,将pps细胞接种到具有利于pps细胞附着的粘附基质的细胞培养容器中并使细胞与包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基接触以生成mge前体细胞。
[0174]
在某些情况中,以粘附培养生成的mge前体细胞可以aeb存在。
[0175]
在一些情况中,可使通过所述培养方法产生的aeb解离并以单层重新接种并在包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基中培养以生成mge前体细胞。可使aeb保持在文中所述的补充物和因子中1
‑
100天,然后重新接种在粘附培养基中并进一步培养成seb或然后解离并以单层重新接种在粘附培养基中。在某些情况中,时间长度可为1
‑
75天、1
‑
50天、1
‑
30天、1
‑
10天,例如,10
‑
50天、10
‑
40天、10
‑
30天、10
‑
20天、5天、10天、20天、或30天。
[0176]
本领域已知的粘附基质以及文中所述的那些可用于在生成mge前体细胞的方法中以粘附培养来培养pps。
[0177]
pps细胞可以粘附培养生长一段时间,然后与包含音速刺猬路径的活化剂和神经诱导补充物的不含血清的培养基接触。在某些情况中,通过供体pps细胞培养物的过生长,或在饲养层不存在下培养pps,或在fgf存在下培养pps细胞等可诱导pps细胞分化。替换地或另外地,可通过与阻止细胞保持未分化显型的某些因子一起培养而开始分化过程。起始分化因子不需要限制向mge前体细胞谱系的分化,但应包含在分化群体中的细胞类型范围内的mge前体细胞或它们的前体。
[0178]
在某个阶段,培养可更具体地针对于mge前体细胞谱系。这可通过在培养基中包含更具体地促进mge前体细胞的生成和增殖的因子而实现。促进mge前体细胞的形成和/或生长的示例性因子包括如文中提供的神经诱导补充物、shh信号传导的活化剂、bmp
‑
信号传导的抑制剂、tgf
‑
β信号传导的抑制剂、wnt抑制剂、和抗
‑
凋亡剂,在一些情况中,可包括fgf信号传导的活化剂。
[0179]
生成mge前体细胞的示例性方法如下所述。
[0180]
可使pps细胞在粘附条件下培养0天
‑
9天,然后将shh的活化剂和/或神经诱导补充物加入培养基中,例如,可使pps细胞在粘附条件下培养至少0小时、1小时、3小时、6小时、12小时、18小时、24小时、36小时、48小时、2天、3天、4天、5天、6天、7天、8天、或9天,然后将shh的活化剂和/或神经诱导补充物加入培养基中。在某些实施方案中,使pps在饲养细胞层不存在下分化。
[0181]
在一些情况中,其中可使pps细胞在粘附条件下在含有shh的活化剂和/或神经诱
导补充物的培养基中培养自pps细胞培养开始起的1
‑
100天以形成mge前体细胞。
[0182]
除了文中所提供的神经诱导补充物和shh信号传导的活化剂,用于在粘附条件下培养pps细胞的培养基可含有抗凋亡剂、smad抑制剂(例如,tgf
‑
β抑制剂、bmp抑制剂、activin抑制剂、nodal抑制剂、或gdf信号传导路径抑制剂)、和wnt抑制剂中的一种或多种。
[0183]
分化因子的添加和去除的发生时间可如上针对aeb形成所述。
[0184]
在某些情况中,用于由pps细胞产生mge前体细胞的方法可包括在包含抗凋亡剂例如rock抑制剂的培养基中培养pps细胞约1小时
‑
35天,例如,至少1小时、至少3小时、至少10小时、至少24小时、至少36小时、至少48小时、至少2天、至少3天,如,4天、5天、6天、7天、8天、9天、15天、20天、25天、或35天。示例性方法可包括将pps细胞以粘附培养接种在含有抗凋亡剂的培养基中,在抗凋亡剂存在下培养pps细胞1小时
‑
35天。在某些情况中,抗凋亡剂可自pps细胞培养生成mge前体细胞开始时存在并在1小时
‑
35天,如,1天至7天后移除。
[0185]
在某些情况中,由pps细胞产生mge前体细胞的方法可包括在含有wnt的一种或多种抑制剂的培养基中培养pps细胞。虽然可在pps细胞培养已经开始时将wnt信号抑制剂加入培养基中,但在培养了数天后(例如,培养10天内的某个时间)可将它加入培养基中。在某些情况中,在粘附条件下开始培养pps细胞的5天内的某个时间,如,0天、1天、或3天内,将wnt信号抑制剂加入培养基中。wnt抑制剂可在整个培养中存在或可存在5天
‑
10天。
[0186]
在某些情况中,用于由pps细胞产生mge前体细胞的方法可包括在包含smad的一种或多种抑制剂的培养基中培养pps细胞。虽然可在pps细胞培养已经开始时将smad信号抑制剂加入培养基中,但在培养数天后(例如,培养10天内的某个时间)可将它加入培养基中以生成mge前体细胞。在某些情况中,在粘附条件下开始培养pps细胞的5天内的某个时间,如,0天、1天、或3天内,将一种或多种smad信号抑制剂加入培养基中。wnt抑制剂可在整个培养中存在以生成mge前体细胞或可存在5天
‑
10天。
[0187]
在某些实施方案中,由灵长类动物多能干(pps)细胞产生内侧神经节隆起(mge)前体细胞的方法可包括在粘附条件下培养pps细胞以生成mge前体细胞,其中培养基包括音速刺猬路径的活化剂、抗凋亡剂、smad的抑制剂、wnt的抑制剂和b27。
[0188]
在某些情况中,培养生成mge前体细胞开始时的ps细胞以103至107个细胞/ml的细胞密度存在。
[0189]
如上所述,不含血清的培养基可用于由pps细胞生成mge前体细胞的方法。不含血清的培养基意指不含未调节或未纯化血清,如,胎牛血清、小牛血清的培养基。不含血清的培养基可包括血清替代,如,文中所述的那些,例如,b27或ns21。
[0190]
mge前体细胞的培养
[0191]
通过文中所述方法生成的seb和aeb可以机械或酶的方式解离,并以单层在具有粘附基质的细胞培养容器上培养。因此,seb和aeb中存在的mge前体细胞可以单层培养。
[0192]
在某些情况中,以单层培养mge前体细胞可进行1
‑
100天,如,10天
‑
15天。
[0193]
以单层培养mge前体细胞可在含有文中提供的神经诱导补充物和shh信号传导的活化剂的培养基中进行。
[0194]
在某些情况中,通过文中所述的方法生成的mge前体细胞可在促进神经元(如,抑制性中间神经元,例如,gaba能中间神经元)生成的培养基中培养。因此,由通过文中所述方法生成的seb和aeb的解离所产生并包含mge前体细胞的seb、aeb、和单层可与促进mge前体
细胞分化成有丝分裂后神经元的培养基接触。在某些情况中,此培养基可不包含smad抑制剂。此外,在某些情况中,此培养基可包含smad活化剂。如此,培养基可包括smad活化剂以增加通过文中所述方案生成的mge前体细胞中存在的中间神经元的群体。示例性smad活化剂包括tgfs(例如,tgfβ3)、bmp(例如,bmp2、bmp4、bmp8)、activin、nodal、gdf、和ide1。
[0195]
在某些情况中,促进mge前体细胞分化成中间神经元的培养基可包括notch抑制剂、bdnf、gdnf、nt3、nt4、camp、维生素c、血清、基质胶、胰岛素、igf、sdf1a、神经调节蛋白1、tgfβ。
[0196]
以单层培养mge前体细胞可导致mge前体细胞的增殖和/或mge前体细胞向具有神经元细胞命运的细胞的分化。在某些情况中,分化成具有神经元细胞命运的细胞的mge前体细胞表达dlx1/2、tuj、map2、gad1/2、和gaba,以及可表达nkx2.1、ascl1、lhx6、lhx7/8、dcx、neun、和vgat中的一个或多个,并可表达亚型标记物钙结合蛋白、钙网膜蛋白、生长激素抑制素、和小清蛋白。
[0197]
在某些情况中,通过文中所述的方法生成的mge前体细胞可与支持细胞群体共培养以诱导mge前体细胞到中间神经元(如,gaba能中间神经元)的分化。
[0198]
未分化态的pps细胞的繁殖
[0199]
利用促进增殖但不促进分化的培养条件,pps细胞可持续地培养繁殖。示例性es培养基利用80%dmem(如knockout dmem“ko dmem”)、20%定义的胎牛血清(fbs,hyclone)或血清替代品(例如,敲除血清替代品(ksr))、1%非必需氨基酸(neaa)、1%青霉素
‑
链霉素
‑
谷氨酰胺(1mm l
‑
谷氨酰胺)、0.0008%β
‑
巯基乙醇、和10ng/ml fgf
‑
碱性(bfgf)制得。
[0200]
pps细胞可通过在抑制分化的环境中培养以未分化态扩增。传统上,pps细胞可在源自小鼠的胚胎或胎儿组织的饲养细胞层上培养。培养板每孔(经放射以抑制增殖但允许支持pps细胞的因子的合成)接种有375,000个放射性小鼠胚胎成纤维细胞(mef),并在接种后使用5小时至10天。在某些实施方案中,也可使用人饲养细胞。
[0201]
pps细胞可保持在未分化状态,即使在饲养细胞不存在下。用于不含饲养物的培养物的环境包含适宜的培养基质,尤其细胞外基体如或层粘连蛋白。pps细胞以>15,000个细胞cm
‑2(优先90,000cm
‑2至170,000cm
‑2)接种。不含饲养物的培养物通过含有在不分化下支持细胞增殖的因子的营养培养基支持。通过利用分泌此类因子的细胞培养培养基,如,放射性(~4,000rad)原代小鼠胚胎成纤维细胞、热调聚的小鼠成纤维细胞、或源自pps细胞的人饲养细胞,可将此类因子引入培养基中。通过将饲养物以~5
‑6×
104cm
‑2的密度接种在不含血清的培养基如补充了20%血清替代品和4至8ng/ml bfgf的ko dmem中来条件化培养基。已条件化1
‑
2天的培养基补充了其它bfgf,并用于支持pps细胞培养1
‑
2天。在以引用的方式并入文中的国际专利公布wo99/20741&wo01/51616;和xu等,nat.biotechnol.19:971,2001中进一步讨论了不含饲养物的培养方法的特征。
[0202]
因子
[0203]
本公开的方法和组合物涉及多种因子的使用,如,神经诱导补充物、抗凋亡剂、分化因子等。以下描述了用于本公开的方法和组合物的神经诱导补充物、抗凋亡剂、分化因子的实例。
[0204]
神经诱导补充物
[0205]
示例性神经诱导补充物包括b27、ns21、或等效补充物。
[0206]
在某些实施方案中,神经诱导补充物可为b27。不含血清的补充物可购自life technologies。b27补充物含有牛血清白蛋白、转铁蛋白、胰岛素、孕酮、皮质酮、三碘
‑1‑
甲腺原氨酸、视黄醇乙酸酯、dl生育酚、dl生育酚乙酸酯、生物素、亚油酸、亚麻酸、乙醇胺、亚硒酸钠、l
‑
肉毒碱、还原型谷胱甘肽、接触酵素、超氧化物歧化酶、d
‑
半乳糖和腐胺。在某些情况中,可使用b27
‑
维生素a。
[0207]
在某些情况中,神经诱导补充物可为ns21。在y.chen等,j.neurosci.methods.,171:239,2008中描述了ns21。y.chen等表明ns21在神经元培养中等效于b27补充物。ns21的配制在y.chen等中有所描述并在下表1中再现。
[0208]
表1.ns1配制
[0209][0210][0211]
在某些情况中,神经诱导补充物可以0.5%至10%范围的浓度,例如,0.5%
‑
5%,例如,0.5%、1%、2%、或3%存在于用于培养pps细胞的不含血清的培养基中。
[0212]
在某些实施方案中,用于培养pps以生成eb的包含shh活化剂和神经诱导补充物的不含血清的培养基不包括ksr或n2补充物。在某些实施方案中,用于生成mge前体细胞的方法不包括在包含bfgf或fgf
‑
2的不含血清的培养基中培养pps细胞。在某些情况中,使pps细胞在包含shh活化剂和神经诱导补充物并不含有ksr或n2补充物或bfgf或fgf
‑
2的不含血清的培养基中培养足以生成seb或aeb的一段时间。
[0213]
在某些情况中,可使seb在包含shh活化剂和神经诱导补充物以及进一步含有ksr补充物、n2补充物、bfgf、和fgf
‑
2中一种或多种的不含血清的培养基中进一步培养足以生成aeb的一段时间。
[0214]
在某些情况中,可使pps细胞在包含shh活化剂和神经诱导补充物以及进一步含有ksr补充物、n2补充物、bfgf、和fgf
‑
2中一种或多种的不含血清的培养基中培养足以生成seb的一段时间。可在与文中所述的shh活化剂和神经诱导补充物的相同时间或稍晚的时间点,如,在pps细胞暴露于文中所述的shh活化剂和神经诱导补充物后的5天
‑
2周后,如,1周
‑
2周后,添加额外补充物。在某些情况中,ksr补充物和/或n2补充物可在pps细胞接触文中所述的shh活化剂和神经诱导补充物后,在分化的第0天,或稍晚些,如,第5天、第7天、第10天、第14天、第21天添加存在。
[0215]
在某些实施方案中,文中所公开的方法中所用的细胞培养基不包含血清替代品,如,ksr或n2。
[0216]
抗凋亡剂
[0217]
在文中所述的方法和组合物的某些实施方案中,用于ps培养细胞的培养基中可包括抗凋亡剂。
[0218]
在某些情况中,抗凋亡剂可为rho
‑
相关的蛋白激酶(rock)的抑制剂。在某些情况中,rock抑制剂可为y27632、ha
‑
100、h
‑
1152、( )
‑
反式
‑4‑
(1
‑
氨乙基)
‑1‑
(吡啶
‑4‑
基氨基羰基)环己烷二氢
‑
氯单水合物(在wo00078351、wo00057913中有所描述)、咪唑并吡啶衍生物(在美国专利no.7,348,339中有所描述)、取代的嘧啶和吡啶衍生物(在美国专利no.6,943,172中有所描述)和取代的异喹啉
‑
磺酰基化合物(在ep00187371中有所描述)、或gsk429286a、rockii抑制剂、或thiazovivin、或其类似物或衍生物。
[0219]
抗凋亡剂可以0.1μm、0.3μm、0.5μm、1μm、至少约1.3μm、至少约1.5μm、至少约2μm、至少约2.3μm、至少约2.5μm、至少约2.8μm、至少约3μm、至少约3.5μm、至少约4μm、至少约4.5μm、至少约5μm、至少约10μm、至少约20μm、至少约30μm、至少约40μm或至少约50μm、如,0.5μm
‑
50μm、1μm
‑
25μm、或2.5μm
‑
20μm的浓度存在。
[0220]
smad抑制剂
[0221]
在文中所述的方法和组合物的某些实施方案中,smad抑制剂可存在于培养细胞的培养基中。在一些实施方案中,smad抑制剂可以10ng/ml、200ng/ml、300ng/ml、400ng/ml、500ng/ml、1μg/ml、1.5μg/ml、2μg/ml、2.5μg/ml、或5μg/ml的浓度,例如,以500ng/ml
‑
3μg/ml,例如,1μg/ml
‑
3μg/ml的浓度存在于用于培养细胞的培养基中。
[0222]
smad抑制剂可以至少约0.01μm、至少约0.03μm、至少约0.1μm、至少约0.2μm、至少约0.25μm、至少约0.3μm、至少约1μm、至少约1.3μm、至少约1.5μm、至少约2μm、至少约2.3μm、至少约2.5μm、至少约2.8μm、至少约3μm、至少约3.5μm、至少约4μm、至少约4.5μm、至少约5μm、至少约10μm、至少约20μm、至少约30μm、至少约40μm或至少约50μm,如,0.5μm
‑
50μm、1μm
‑
25μm、或5μm
‑
20μm的浓度存在。
[0223]
在某些实施方案中,smad抑制剂可为tgf
‑
β信号传导的抑制剂。例如,smad抑制剂可为结合tgf
‑
β1、tgf
‑
β2、tgf
‑
β3、tgf
‑
β受体i和/或ii的alk抑制剂,或其抗体或片段。在某些实施方案中,tgf
‑
β信号传导的抑制剂可为小分子抑制剂。在某些情况中,tgf
‑
β信号传导的抑制剂可为ly364947(sd208)、sm16、sb
‑
505124、alk5抑制剂ii、sb
‑
431542、ly2157299、
ldn
‑
193189、a83
‑
01、( )
‑
itd
‑
1、itd
‑
1(4
‑
([1,1'
‑
联苯]
‑4‑
基)
‑
2,7,7
‑
三甲基
‑5‑
氧代
‑
1,4,5,6,7,8
‑
六氢喹啉
‑3‑
羧酸乙酯)、或itdt。
[0224]
在某些实施方案中,smad抑制剂可为bmpria
‑
fc、noggin、或其衍生物。
[0225]
在某些实施方案中,smad抑制剂可为bmp路径抑制剂,如,dorsomorphin。
[0226]
在某些实施方案中,smad抑制剂可为activin抑制剂、nodal抑制剂、或gdf信号传导路径抑制剂。示例性activin抑制剂包含sb431542、卵泡抑素、a8301、dmh1、dorsomorphin、k02288、和sb505124。在某些情况中,可使用nodal抑制剂,如,sb431542、lefty、或cerebrus。在某些情况中,sb431542、d4476、gw788388、ly364947、repsox、sb525334、sd208可用于抑制gdf信号传导路径。
[0227]
在某些实施方案中,文中所述的方法中使用的细胞培养基可包括两种或更多种smad抑制剂。
[0228]
音速刺猬信号传导的活化剂
[0229]
在文中所述的方法和组合物的某些实施方案中,音速刺猬信号传导的活化剂可存在于培养细胞的培养基中。音速刺猬信号传导的活化剂可以至少约0.01μm、至少约0.03μm、至少约0.1μm、至少约0.2μm、至少约0.25μm、至少约0.3μm、至少约1μm、至少约1.3μm、至少约1.5μm、至少约2μm、至少约2.3μm、至少约2.5μm、至少约2.8μm、至少约3μm、至少约3.5μm、至少约4μm、至少约4.5μm、至少约5μm、至少约10μm、至少约20μm、至少约30μm、至少约40μm或至少约50μm,如,0.05μm
‑
5μm、0.01μm
‑
2.5μm、0.05μm
‑
2μm、或0.1μm
‑
2μm的浓度存在。
[0230]
在某些情况中,音速刺猬信号传导的活化剂可为shh或其衍生物。在某些情况中,音速刺猬信号传导的活化剂可为小分子,如,purmorphamine、sag平滑激动剂、hh
‑
ag1.5、或其衍生物及类似物。
[0231]
wnt抑制剂
[0232]
在文中所述的方法和组合物的某些实施方案中,wnt信号传导的抑制剂可存在于培养细胞的培养基中。
[0233]
wnt抑制剂为下调wnt表达或活性的制剂。所关注制剂可直接与wnt相互作用,例如,药物,即小分子、阻断抗体等,或可与wnt相关蛋白相互作用,例如,wnt共受体lrp5/6和跨膜蛋白kremen。大量wnt抑制剂已被描述并在本领域已知。
[0234]
所关注的wnt抑制剂干扰可溶性细胞外wnt蛋白与存在于正常细胞表面上的卷曲蛋白受体之间的相互作用。所述制剂包括,但不限于,包含wnt结合域的可溶性卷曲蛋白多肽;可溶性卷曲蛋白相关多肽;wnt特异性抗体;卷曲蛋白特异性抗体;以及能够阻断细胞外wnt信号传导的其它分子。
[0235]
在已知的wnt抑制剂之中的是dickkopf(dkk)基因家族的成员(参见krupnik等(1999)gene 238(2):301
‑
13)。人dkk基因家族的成员包括dkk
‑
1、dkk
‑
2、dkk
‑
3、和dkk
‑
4、以及dkk
‑
3相关蛋白soggy(sgy)。
[0236]
其它wnt抑制剂包括wise(itasaki等(2003)development 130(18):4295
‑
30),其为一种分泌蛋白。wise蛋白以物理方式与wnt共受体脂蛋白受体
‑
相关蛋白6(lrp6)相互作用,并可与wnt8竞争lrp6结合。
[0237]
抑制剂也可包括天然抑制剂的衍生物、变体、和生物活性片段。
[0238]
在某些情况中,wnt抑制剂可为小分子,如,cki
‑
7、iwp类似物、iwr类似物、xav939、
53ah、wnt
‑
c59。
[0239]
在某些情况中,wnt抑制剂可以10ng/ml、200ng/ml、300ng/ml、400ng/ml、500ng/ml、1μg/ml、1.5μg/ml、2μg/ml、2.5μg/ml、或5μg/ml的浓度,例如,以500ng/ml
‑
3μg/ml,例如,1μg/ml
‑
3μg/ml的浓度存在于培养基中。
[0240]
评价细胞群体的生成
[0241]
在某些情况中,可监测根据文中所公开的方法培养的细胞群体以评价通过培养(例如,在文中所公开的培养方法的一个或多个时间点期间)所赋予的细胞的变化以表征所产生的细胞群体。在某些实施方案中,可通过确定这些细胞群体所特有的标记物的表达来评价mge前体细胞(有丝分裂mge前体细胞和/或有丝分裂后中间神经元)的产生。
[0242]
在某些情况中,通过检测标记物转录或蛋白表达的存在或不存在来确定某些标记物的表达。或者,可通过测量标记物在细胞培养物或细胞群体的细胞中存在的水平确定某些标记物的表达。在此类方法中,标记物表达的测量可为定性的或定量的。定量由标记物基因产生的标记物的表达的一种方法是通过使用定量pcr(q
‑
pcr)。进行q
‑
pcr的方法在本领域是公知的。本领域已知的其它方法也可用于定量标记物基因表达。例如,可通过使用对所关注的标记物基因产物特异的抗体检测标记物基因产物的表达。在某些方法中,可确定所关注的细胞群体的标记物基因特征的表达以及ps细胞和其它细胞类型的标记物基因特征的显著表达的缺乏。
[0243]
可通过确定nkx2.1基因的表达监测mge前体细胞的生成。如此,通过文中所述的方法产生的mge前体细胞表达nkx2.1标记物基因,进而产生nkx2.1基因产物。由文中所述的方法产生的mge前体细胞也表达foxg1,并可表达lhx6、lhx7/8、olig2、ascl1、和dlx2。并且,由文中所述方法产生的mge前体细胞不表达pax6。
[0244]
在文中所述的一些实施方案中,mge前体细胞中的nkx2.1标记物和/或foxg1标记物的表达比非
‑
mge前体细胞(例如,多能干细胞)中的nkx2.1标记物和/或foxg1标记物的表达高出至少约2
‑
倍至至少约10,000
‑
倍。在其它实施方案中,在mge前体细胞中的nkx2.1标记物和/或foxg1标记物的表达比非
‑
mge前体细胞(例如,多能干细胞)中的nkx2.1标记物和/或foxg1标记物的表达高出至少约4
‑
倍、至少约6
‑
倍、至少约8
‑
倍、至少约10
‑
倍、至少约15
‑
倍、至少约20
‑
倍、至少约40
‑
倍、至少约80
‑
倍、至少约100
‑
倍、至少约150
‑
倍、至少约200
‑
倍、至少约500
‑
倍、至少约750
‑
倍、至少约1000
‑
倍、至少约2500
‑
倍、至少约5000
‑
倍、至少约7500
‑
倍或至少约10,000
‑
倍。
[0245]
在某些情况中,可通过对所关注的细胞进行功能分析来监测mge前体细胞(有丝分裂mge前体细胞和/或有丝分裂后中间神经元)的生成。例如,通过文中所述的方法生成的mge前体细胞可体内或体外生成中间神经元。在某些情况中,通过文中所公开的方法产生的mge前体细胞可生成中间神经元,中间神经元分化成抑制性gaba能中间神经元,抑制性gaba能中间神经元可迁移并在功能上与神经元体内整合。
[0246]
在某些情况中,所述方法不包括监测mge前体细胞的生成。
[0247]
细胞群体的富集、分离和/或纯化
[0248]
所关注的细胞群体,如,通过上述方法中任一种产生的mge前体细胞(有丝分裂mge前体细胞和/或有丝分裂后中间神经元)可通过利用对此类细胞特异的亲和标签富集、纯化和/或纯化。对所关注细胞或细胞群体特异的亲和标签的实例包括抗体、配体、或对标记物
分子特异的其它结合剂,如多肽,其存在于所关注的细胞的细胞表面上但基本上不存在于通过文中所述的方法产生的细胞培养物中所发现的其它细胞类型上。
[0249]
制得抗体并使用它们进行细胞分离的方法在本领域是已知的并且可实施这些方法与文中所述的抗体和细胞一起使用。在一个方法中,使结合所关注的细胞群体所表达的标记物的抗体附接到磁珠并随后允许结合到细胞培养物中所关注的细胞,该细胞培养物以酶的方式经处理以减少细胞间与基质粘附。然后,使细胞/抗体/珠复合物暴露于磁场以将珠
‑
结合的定型内胚层细胞与未结合细胞分开。一旦所关注细胞以物理的方式与培养的其它细胞分开,则抗体结合被破坏并将细胞重新接种在适当的组织培养基中。
[0250]
也可使用用于得到富集的、分离的、或纯化的所关注的细胞群体的额外方法。例如,在一些实施方案中,用于所关注细胞表达的标记物的抗体为包含所关注细胞的已经处理以减少细胞间和基质粘附的孵育细胞培养物。然后,对细胞进行冲洗、离心和再悬浮。然后,使细胞悬浮液与二抗,如可结合一抗的fitc
‑
共轭抗体一起孵育。然后,对细胞冲洗、离心并再悬浮在缓冲液中。然后,分析细胞悬浮液并利用荧光活化细胞分类仪(facs)分类。单独地从未结合标记物特异性抗体的细胞中收集抗体
‑
结合细胞,进而分离出所关注的细胞。如果需要,可通过利用另一种基于亲和力的方法或通过利用对所关注的细胞特异的相同或不同标记物的另一轮分类对分离的细胞组合物进行进一步纯化。在某些情况中,mge前体细胞通过基于尺寸分类细胞来富集。
[0251]
在某些情况中,在使ps细胞培养物诱导分化成mge前体细胞谱系后,从其它类型的细胞中富集、分离和/或纯化所关注的细胞,如,mge前体细胞。应了解,上述富集、分离和纯化步骤可利用在分化的任何阶段的此类培养物。
[0252]
除了上述步骤,所关注的细胞,如,mge前体细胞也可通过用于细胞分离的其它技术分离。此外,也可通过在促进所关注的细胞的选择性存活或选择性扩增的生长条件中的连续传代培养的方法富集或分离所关注的细胞,如,mge前体细胞。
[0253]
利用文中所述的方法,产生富集了未富集的细胞群体至少约2
‑
至约1000倍的所关注的细胞(如,mge前体细胞)的细胞群体或细胞培养物。在一些实施方案中,相比未处理细胞群体或细胞培养物,mge前体细胞可富集至少约5
‑
至约500
‑
倍。在其它实施方案中,相比未分化细胞群体或细胞培养物,mge前体细胞可富集至少约10
‑
至约200
‑
倍,至少约20
‑
至约100
‑
倍,至少约40
‑
至约80
‑
倍,或至少约2
‑
至约20
‑
倍。
[0254]
本公开的细胞群体的遗传型特点
[0255]
当源自分离的ps细胞,或ps细胞的确立的谱系时,本公开的细胞群体可表征为来源细胞或细胞谱系的后代。因此,细胞群体将具有与所源自的细胞相同的基因组。此意指除了任何核型变化,ps细胞与其生成的细胞群体之间的染色体dna同一性超过90%。因为所有非操作的基因成分被保留,所以经过重组方法处理以引入转基因或敲除内源基因的本公开的细胞群体仍被认为具有与所源自的谱系相同的基因组。本公开的细胞群体和ps细胞通过标准基因技术确定具有相同基因组。具有相同基因组也可推断出细胞群体是否通过正常有丝分裂的过程获自未分化谱系。
[0256]
在某些工业应用中,此特征为本公开的细胞群体的有价值的特点。具体地讲,因为可根据需求使ps细胞增殖并分化成更多的本公开细胞群体,所以来源ps细胞的可利用性进一步供应基因上匹配的分化细胞群体。并且,ps细胞可分化成其它治疗上重要的谱系。
[0257]
本技术中所述的技术允许在分化前或后扩增细胞来制造共享相同基因组的大细胞群体。108、10
10
、或10
12
个细胞群体在理论上是可能的。通常将此类大群体分开加入适于进一步培养、药物筛选、或治疗施用的单独容器中。
[0258]
本公开的某些实施方案包括来源细胞(如未分化的ps细胞谱系),或中间群体,与具有mge前体细胞特征的分化细胞的一种或多种群体的组合。所述群体可在同一个容器中,在同一个设备的分开的容器中,或在两个不同地点。未分化和分化细胞可同时或在不同时间点存在,如,当使未分化细胞的培养物分化成mge前体细胞时,如文中所述。
[0259]
本公开的包含细胞群体的组合物
[0260]
通过上述方法产生的细胞组合物包括含有分离的mge前体细胞和富集分离的mge前体细胞的细胞群体的细胞培养物。
[0261]
在一些实施方案中,可产生包含本公开的细胞的细胞组合物,其中培养物中至少约50%
‑
80%细胞为所关注的细胞。文中所述的分化方法可导致多能细胞到所关注的细胞的约5%、约10%、约15%、约20%、约25%、约30%、约35%、约40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%、或大于约95%转化。
[0262]
在一些实施方案中,其中例如通过使用结合所关注的细胞的亲和试剂分离所关注的细胞,可恢复所关注的基本上纯的细胞群体。
[0263]
文中所述的一些实施方案涉及细胞组合物,其包含至少约5%的所关注的细胞至至少约95%的所关注的细胞。在一些实施方案中,细胞培养物或细胞群体包括哺乳动物细胞。在优选实施方案中,细胞培养物或细胞群体包括人细胞。例如,某些特定实施方案涉及包含人细胞的细胞组合物,其中至少约5%至至少约95%的人细胞为mge前体细胞。其它实施方案涉及包含人细胞的细胞组合物,其中至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%或大于90%的人细胞为mge前体细胞。
[0264]
在一个特定实施方案中,提供了包含人细胞的组合物,其中人细胞中至少70%为人mge前体细胞。
[0265]
在另一个实施方案中,提供了包含人细胞的组合物,其中人细胞中至少80%为人mge前体细胞。
[0266]
在某些实施方案中,所述组合物可包含多能干细胞和/或抑制性中间神经元。
[0267]
本公开的细胞群体,如,包含mge前体细胞的seb、包含mge前体细胞的aeb、mge前体细胞、或自mge前体细胞生成的神经元可以包含这些细胞群体中的一种或多种的组合物存在。
[0268]
本公开的细胞群体可即产即用或采用本领域可接受的方法储存,如,在解冻和使用前,冷冻保存1天至10年。
[0269]
可利用文中所述的标记物和方法以及本领域已知的那些评价上述方法产生的细胞组合物和其组合物。
[0270]
可利用文中所述的方法和本领域已知的那些富集、分离或纯化文中所述方法产生的细胞组合物和其组合物。
[0271]
本公开的细胞群体的用途
[0272]
用于筛选的细胞群体
[0273]
本公开的细胞可用于筛选影响mge前体细胞和/或其生成的细胞(如,中间神经元)的特征的试剂(如,小分子、肽、多核苷酸)或环境条件(如,培养条件或操作)。
[0274]
在一个实例中,mge前体细胞用于筛选促进向中间神经元的成熟,或促进mge前体细胞在长期培养中的增殖和保持的因子。例如,候选分化因子或生长因子通过加入不同孔的细胞中,然后根据用于细胞的进一步培养和使用的理想标准确定产生的任何表型变化来进行测试。此可得到用于产生和/或保持mge前体细胞和/或其产生的细胞的改良的衍生和培养方法。
[0275]
本公开的其它筛选方法涉及药物化合物对mge前体细胞和/或其生成的细胞的潜在副作用的测试。此类筛选不仅适于化合物何时被设计以对mge前体细胞自身具有药理作用,而且也用于测试针对在别处的主要药理作用设计的化合物的与mge前体细胞相关的副作用。
[0276]
其它筛选方法涉及mge前体细胞用于测定具有影响mge前体细胞的潜能的小分子药物的效果的用途。对此,可使细胞与测试化合物体外组合,并确定化合物对mge前体细胞的影响。
[0277]
在美国专利号5,030,015中,以及在教材in vitro methods in pharmaceutical research,academic press 1997中描述了药物筛选的一般原理。候选药物化合物的活性的评价一般涉及将本发明的分化细胞单独地或与其它药物组合的方式与候选化合物组合。研究者确定归因于化合物的细胞的形态学、标记物、或功能活性的任何变化(与未处理细胞或经阴性对照化合物处理的细胞比较),然后使化合物效果与观察到的变化相关联。
[0278]
临床治疗中的mge前体细胞
[0279]
包含mge前体细胞的细胞群体,如,富集了mge前体细胞以及通过文中方法产生的纯化的mge前体细胞的细胞群体可用于多种临床应用。
[0280]
在某些实施方案中,利用文中提供的方法产生的mge前体细胞可用于治疗需要利用mge前体细胞的治疗的受试者。
[0281]
在某些情况中,需要利用mge前体细胞的治疗的受试者可为患有由降低的抑制性中间神经元活性表征的神经障碍或具有患有该神经障碍风险的患者。在某些情况中,患者可具有降低的抑制性神经元功能和/或提高的刺激性神经元功能。
[0282]
在某些情况中,可将mge前体细胞移植到受试者的目标部位,该受试者为mge前体细胞分化成中间神经元,如,gaba能抑制性中间神经元提供适宜的分化条件。可通过将细胞递送到组织中的本领域的多种标准方法中的任一种移植细胞,例如,将它们以悬浮液注入携带体中,如,可溶性溶液或固体或半固体支持体。适宜溶液包括盐水、pbs、l15、dmem、iscove培养基等。适宜的固体载体包括珠、例如网格过滤器的过滤器、膜、等。
[0283]
在某些情况中,可将mge前体细胞施用到受试者的神经系统中。在某些情况中,可通过将mge前体细胞移植到受试者神经系统的一处或多处位置来进行施用。
[0284]
在某些实施方案中,可将mge前体细胞施用到受试者的神经系统的一处或多处位置,如,中枢神经系统,如,脑,例如,小脑、大脑皮层、海马体、纹状体(例如,基底核)、丘脑、下丘脑、丘脑下核、和脊髓。
[0285]
在某些实施方案中,mge前体细胞的施用可导致抑制性神经元功能恢复。在某些情况中,施用可包括将mge前体细胞移植到受试者大脑的第一部分中并恢复远离第一部分的大脑的第二部分的抑制性神经元功能。
[0286]
在某些实施方案中,可将mge前体细胞施用到患有神经障碍或具有患有该神经障碍风险的受试者,该神经障碍例如,癫痫症,例如,癫痫、亨廷顿氏病(huntington’s disease)、帕金森氏病(parkinson’s disease)、als、精神分裂症、阿尔茨海默病(alzheimer’s disease)、自闭症、运动障碍、慢性疼痛、僵直、神经性疼痛、多发性硬化、外伤性脑损伤、脱髓鞘疾病、双向情感障碍、抑郁症、和癌症。
[0287]
实施例
[0288]
给出以下实施例以向本领域普通技术人员提供如何制作及使用本发明的完整公开内容和具体内容,并且无意限制被发明者视为本发明的内容的范围,也无意表示以下试验为进行的所有或仅有的试验。已付出努力以确保就所用数值(例如,量、温度等)而言的准确度,但应考虑一些试验误差及偏差。除非另外指出,份为重量份,分子量为重量平均分子量,温度为摄氏度,以及压力为大气压或近大气压。可使用标准缩写,例如,bp,碱基对;kb,千碱基对;pl,皮升;s或sec,秒;min,分钟;h或hr,小时;aa,氨基酸;kb,千碱基对;bp,碱基对;nt,核苷酸;i.m.,肌内;i.p.,腹膜内;s.c.,皮下;等。
[0289]
虽然本发明已参照其特定实施方案加以描述,但本领域技术人员应了解可进行多种变化并且可在不脱离本发明的实质主旨和范围内可替代有效项。此外,可进行多处修饰以使特定情况、材料、物质组合物、方法、一个或多个方法步骤适合本发明目的、主旨和范围。所有这些修饰在随附的权利要求的范围内。
[0290]
材料和方法
[0291]
细胞培养和facs
‑
分类
[0292]
使hes
‑
3hesc保持在具有20%敲除血清替代品、1%非必需氨基酸(neaa)、1%青霉素
‑
链霉素
‑
谷氨酰胺、0.0008%2
‑
巯基乙醇、和10ng/ml fgf
‑
碱性(invitrogen)的敲除dmem中的经辐射的小鼠胚胎成纤维细胞(millipore)上。通过胶原酶iv/分散酶(每种1mg/ml;invitrogen)对hesc菌落的优选选择开始分化。使菌落胰蛋白酶化成单细胞,并如所述(eiraku,m.,等(2008).cell stem cell 3,519
‑
532),将~10,000个细胞/孔接种在低粘附性圆底96
‑
孔板(corning或nof)中以在优化的b27 5f分化培养基#1中形成一个seb/孔,该培养基#1由neurobasal
‑
a、2%b27
‑
维生素a(invitrogen)、和hesc培养基中相同的补充物但不含fgf组成。同样,添加y27632(10μm)、sb431542(10μm)、purmorphamine(1
‑
2μm)(stemgent)、bmpria
‑
fc(1.5μg/ml)、和dkk1(1μg/ml)(invitrogen)。
[0293]
在第7天,移除y27632,并将seb整块地粘附接种在含培养基#1的基质胶(bd biosciences)涂覆的板上。在第14天,移除培养基#1中的因子,除了purmorphamine。在第25天,使aeb胰蛋白酶化并以解离重新单层接种在基质胶或多聚鸟氨酸/层粘连蛋白涂覆的板上。从第27天
‑
第30天添加dapt(10μm)(tocris)。在第35天进行facs分类后,将gfp 细胞重新接种(10
‑
25,000个细胞/cm2)在培养基#1中的皮质胶质细胞上。胶质细胞由新生的小鼠皮层制得,利用血清传代至少3x以去除小鼠神经元,并在汇合下经ara
‑
c(5μm)(sigma)预处理。每隔3
‑
4天,一半的培养基被分化取代基#2替换:neurobasal
‑
a、2%b27 维生素a(invitrogen),以及与培养基#1中相同的补充物,不含因子,除了移除neaa和
purmorphamine,并添加bdnf(25ng/ml)(r&d systems)。每隔3
‑
4天替换培养基。
[0294]
对facs aria ii(bd biosciences)进行facs分析和分类。针对活细胞dapi排除、小分散尺寸、单细胞、nkx2.1
‑
gfp信号、和psa
‑
ncam
‑
apc或
‑
pe(miltenyi biotech)信号强度分类细胞。使用gfp
‑
阴性细胞和同型抗体(santa cruz)对照。利用flowjo(treestar)软件分析数据。对于活细胞成像,分化与以上类似,但将5,000个细胞/孔接种在aggrewell
‑
800板(stem cell technologies)上,并在第4
‑
7天,整块地接种seb。利用温度(37℃)和气体(5%o2,5%co2,90%n2)对照(leica sp5)通过延时共聚焦显微镜进行成像。
[0295]
将培养一周的mge细胞,或在第35天分类的hesc重新接种在皮层培养基上(~10
‑
20,000个细胞/cm2)。
[0296]
对hesc进行的所有试验遵守ucsf批准的干细胞研究监管委员会和人研究方案委员会。
[0297]
移植和移植物增殖
[0298]
从分类的细胞颗粒中移除过量培养基以产生1000个细胞/nl的浓缩细胞悬浮液。将细胞负载到固定在立体定位的液压注射器的斜切的玻璃微量吸液管(wiretrol 5ul,drummond scientific company)中。通过低体温使p2cb.17
‑
scid幼崽(charles river)麻醉并定位在注射器平台上的粘土头模具中。利用以下坐标将每个注射点10
‑
100,000个细胞以经颅的方式递送到右大脑皮层中:距中线0.9mm(矢状窦)、及距λ2.6mm。注射深度距离皮肤表面0.45mm。所有移植试验遵守ucsf批准的机构动物护理和使用委员会方案。
[0299]
首先,针对nkx2.1
‑
gfp 分类第35天的单层培养物,不采用psancam筛选,并注入新生的scid小鼠皮层中。但是,在4个月内,经注射的小鼠大脑(12/12只小鼠)在注射点和/或近注射道的软膜表面处含有肿瘤样过生长(未显示数据)。这些不是多能细胞源性的畸胎瘤;没有发现oct4 细胞或极化神经上皮玫瑰花状结构。生长主要含有nkx2.1
‑
gfp 神经细胞。肿瘤被定义为在注射后坚持四个月以上的人
‑
特异性核抗原(hna)阳性以及ki67 人细胞(mpi)的核心。首先,因为第35天的培养物似乎含有很少表达ki67、gfap、或olig2的神经祖细胞(图3e),所以肿瘤发病率是个惊喜。但是,在扩展的共培养物中偶尔检测到局灶生长(~1个病灶/2500个接种细胞,n=3),与体内肿瘤形成一致。因此,我们进行方案优化以阻止这些生长。低密度单层培养促进神经元分化并降低肿瘤发病率到50%(6/12只小鼠)。通过在注射前简单地添加dapt、notch信号传导的γ分泌酶抑制剂以降低神经元分化使肿瘤发病率进一步降到33%(3/9只小鼠)。当在共培养或移植前,使第35天的培养物经dapt预处理并经facs
‑
分类出nkx2.1
‑
gfp 和psa
‑
ncam 细胞,没出现体外局灶生长(n=3)和体内肿瘤发病率(0/6只小鼠),但是gfp 和psa
‑
ncam
‑
阴性细胞继续形成病灶和肿瘤(3/4只小鼠)。提供了关于注射动物的总结(图16)。
[0300]
免疫染色
[0301]
将培养细胞在4%多聚甲醛中固定10
‑
20min。在穿心灌注和利用振动切片机或滑动式切片机切片(50μm)后,使小鼠大脑在
‑
4℃下固定过夜。利用低温恒温器切片(25μm)eb和人组织。对细胞和切片染色:5
‑
10min的利用煮沸的0.01m柠檬酸盐缓冲液ph=6的抗原修复,1小时的利用5%血清和0.1%tritonx100的pbs的封闭,
‑
4℃下与在封闭缓冲液(除了用于gaba抗体的不含triton的缓冲液)中的一抗过夜,在pbs tritonx100中冲洗3x,2小时二抗(invitrogen),在pbs tritonx100中冲洗4x。表2列出一抗。
[0302]
表2.抗体
[0303]
抗体公司编号稀释ascl1cosmo biosk
‑
t01
‑
0031
‑
500calbswantcb 381
‑
2000calrswant76991
‑
2000chatmilliporeab144p1
‑
300couptfiiperseus proteomicspp
‑
h7147
‑
001
‑
1000darpp32santa cruzsc
‑
113651
‑
1000dcxcell signaling4604s1
‑
500dlx2来自k.yoshikawa的赠品 1
‑
1000er81covanceprb
‑
362c1
‑
1000foxg1来自y.sasai的赠品 1
‑
500gabasigmaa20521
‑
2000gfapmilliporemab34021
‑
500gfpaves labsgfp
‑
10201
‑
1000hnamilliporemab12811
‑
500islet1dshb39.4d51
‑
200ki67abcamab155801
‑
500ncadherinbd biosciences5615541
‑
50neunmilliporemab3771
‑
150neunmilliporemab377b1
‑
150nkx2.1novacastrattf
‑1‑
l
‑
ce1
‑
150nkx2.1santa cruzsc
‑
130401
‑
250nkx2.2dshb74.5a51
‑
100olig2milliporeab96101
‑
500pax6milliporeab54091
‑
250pvsigmap30881
‑
4000pvswantpv
‑
251
‑
2000rax来自y.sasai的赠品 1
‑
500rfpclontech6324961
‑
500rfpchromotek5f81
‑
1000sstbachemt
‑
4103.00501
‑
500tbr1abcamab319401
‑
500thpelfreezep40101
‑
01
‑
1000tujicovancemms
‑
435p1
‑
1000vgatsynaptic systems131 0031
‑
500
[0304]
转录表达
[0305]
利用rneasy试剂盒(qiagen)由细胞颗粒制得rna。利用superscript iii
‑
第一链试剂盒(invitrogen)制得cdna。通过实时pcr系统(applied biosystems)利用sybr绿色主
instruments)制得斑状电极。使吸液管尖端填装有包含以下的内部溶液(mm):125k
‑
葡萄糖酸盐、15kcl、10hepes、4mgcl2、4na2atp、0.3na3gtp、10tris
‑
磷酸肌酸、0.2egta。
[0311]
对于培养的神经元,使浴槽不断地灌注包含以下的新鲜记录培养基(mm):145nacl、3kcl、3cacl2、2mgcl2、10hepes、8葡萄糖。利用组织切片机(leica vt1200s)切割横向切片(300μm)并保持在具有含有以下的acsf的孵育室中(mm):110胆碱cl、2.5kcl、0.5cacl2、7mgcl2、1.3nah2po4、25nahco3、10葡萄糖。切片记录培养基包含(mm):125nacl、2.5kcl、2cacl2、1.3mgcl2、1.3nah2po4、25nahco3、10葡萄糖。利用axon 700b膜片钳放大器和1320a界面(axon instruments)进行记录。利用放大器电路在2khz下过滤信号,在10khz下采样,并利用clampex 10.2(axon instruments)分析。
[0312]
利用具有gfp激发带通滤波器和光脉冲的汞灯(75mw)递送光刺激并利用maste
–
8(a.m.p.i.)通过高速快门(uniblitz)产生光脉冲,蓝光的功率密度(boyden,e.s.,等(2005).nat neurosci 8,1263
‑
1268)(nagel,g.,等(2003).proc natl acad sci usa 100,13940
‑
13945)为8
‑
12mw
·
mm
‑
2,其利用功率表(coherent instruments)测量。
[0313]
统计分析
[0314]
数据呈现为平均值
±
s.e.m。图3e数据表示成共表达/gfp 细胞的平均值%。图4c
‑
h数据表示成平均珠信号强度。图5c数据表示成ubc
‑
rfp 、chr2
‑
yfp 、或tuj 神经元的平均值%。图7d
‑
e数据表示成hna 或ubc
‑
rfp 人细胞的平均值%。统计对比利用单因素anova,以及使用邦弗朗尼事后检验(post hoc bonferroni test)用于电生理数据,并使用双侧、双样品不等方差学生t检验用于免疫染色数据。利用使用matlab插件的imaris(bitplane)软件计算细胞数。表4列出关于每个试验和标记物的样品大小和细胞数的总结。
[0315]
表4.对每个图、标记物、及分化期的样品大小和细胞数的总结
[0316][0317]
b27 5f方法的发展
[0318]
我们对比了三个公布方案(方法#1、#2、和#3)的诱导来自hesc的nkx2.1 mge前体细胞的能力。第一个方法(#1)以~13%效率报道了hesc源性的nkx2.1 和foxg1 mge前体细胞(watanabe,k.,等(2007).nat biotechnol 25,681
‑
686)。此35天方案采用了不含血清(敲除血清替代品(ksr)
‑
早期/b27
‑
晚期)的补充培养基以及bmp(通过bmpria
‑
fc)和activin/nodal(结点)(通过sb431542)信号传导路径的双
‑
smad抑制以指导神经外胚层样分化,以及wnt路径抑制(通过dkk1)以详述拟胚体(eb)的前脑样特性。从第24
‑
35天,移走抑制剂,并加入音速刺猬(shh)以详述腹前前脑样细胞。第二种方法(#2)报道了在不含血清(ksr/n2)的补充培养基中28天后~84%效率下的nkx2.1 和foxg1 mge前体细胞(li,x.j.,等(2009).development 136,4055
‑
4063)。没有使用双
‑
smad抑制,但在分化期间(第10
‑
28天)早期开始shh处理,同时进行或没有进行wnt抑制。虽然这些方案被报道生成foxg1 和nkx2.1 细胞,但是没有研究腹侧端脑mge
‑
样对poa/隔膜
‑
样特性。
[0319]
在我们看来,方法#1(图8b)、方法#2(未显示)、和方法#1的优化版本(图9a)不能生成nkx2.1
‑
gfp mge前体
‑
样细胞。当使用shh来替代purmorphamine时,同样如此。但是,我们发现混合法可以如荧光活化细胞分类(facs)量化的12%效率生成nkx2.1
‑
gfp mge前体
‑
样细胞(图8c)。此25天混合法涉及在整个方案中双
‑
smad(通过sb431542和bmpria
‑
fc)和wnt(经由dkk1)抑制以及同时从第10
‑
25天在不含血清(ksr/b27)的补充培养基中进行平滑激动剂purmorphamine处理。在b27/b27补充培养基中达到类似效率(未显示)。我们认为gfp 细胞诱导由早期(第10天)shh路径活化引起,与方法#1的晚期(第24天)shh不同,我们假设甚至更早地添加purmorphamine(从第0天
‑
第25天)将增加nkx2.1 mge前体细胞的百分比。但是,此修饰导致在ksr/b27培养基中的效率降低(1.9%)(图8d)。
[0320]
但是,在25天方案中,利用b27补充培养基替换ksr,以及早期添加五种因子[rho
‑
相关蛋白激酶(rock)抑制剂(y27632)、双
‑
smad抑制剂(sb431542和bmpria
‑
fc)、wnt抑制剂(dkk1)、和平滑激动剂(purmorphamine)]出人意料地导致大多数细胞(70.2%)变成nkx2.1
‑
gfp mge前体细胞(图8e)。并且,当分化两周后移走抑制剂时,方案延长到第35天,我们获得通过facs分析高达90.8%的nkx2.1
‑
gfp 分化效率(图8f)。因此,shh路径的早期活化与b27或缺少ksr的培养基(b27 5f方法)的组合导致有效的自hesc的腹侧前脑
‑
样分化。
[0321]
在此研究期间,第三种方法(方法#3)被报道以引导在fgf2和视黄酸存在下,以12
‑
14%效率的自hes
‑
3nkx2.1gfp/w hesc的下丘脑脑前
‑
样分化(goulburn,a.l.,等(2011).stem cells 29,462
‑
473)。没有使用shh,但是培养基看起来诱导shh、和wnt抑制剂在培养中表达并促进腹前神经命运,但是使用了视黄酸,其可用作尾侧激活剂。50天后,一些细胞表达foxg1,但是没确定这些细胞的效率和命运。在我们看来,方法#3以~7%效率生成nkx2.1
‑
gfp 细胞。符合下丘脑
‑
样特性,与b27 5f方法相比,方法#3在第24天生成较少的nkx2.1 表达和较多的nkx2.2 细胞(数据未显示)。
[0322]
nkx2.1
‑
gfp 细胞衍生需要shh路径活化。在此研究中,我们使用1
‑
2μm purmorphamine或500ng/ml shh。虽然2
‑
6μm purmorphamine略微增加在eb培养基中的gfp 分化效率,但是这些更高的浓度降低后期单层培养物的生存力(未显示)。我们也研究了对于hesc
‑
端脑
‑
样特性是否可省去wnt抑制,如先前所表明(li,x.j.,等(2009).development 136,4055
‑
4063)。虽然gfp 细胞分化效率类似,但是不存在dkk1 wnt抑制剂会导致表达端
脑foxg1的细胞数从70%降到45%(未显示)。除了wnt,fgf在神经发育期间用作重要的喙尾模式因子(borello,u.,等(2008).neural dev 3,17)(mason,i.(2007).nat rev neurosci 8,583
‑
596)(ye,w.,等(1998).cell 93,755
‑
766),fgf8关联到自小鼠esc的mge端脑
‑
样发育(danjo,t.,等(2011).j neurosci31,1919
‑
1933)。我们没有将fgf8加入我们的培养物中,但是我们确实检测到fgf8转录表达,与其在shh下游的作用一致(gutin,g.,等(2006).development 133,2937
‑
2946)(ohkubo,y.,等(2002).neuroscience 111,1
‑
17)。在开始分化时(第0
‑
25天),添加fgf抑制剂(pd173074)使表达foxg1的细胞减少(55%),当在第14天(第14
‑
25天)添加相同抑制剂对foxg1水平没有影响(未显示)。因此,类似于胎儿前脑发育,wnt的早期抑制和fgf信号传导路径的活化,连同shh活化,在模式化hesc源性的培养物的腹侧
‑
端脑
‑
样特性中发挥作用。
[0323]
与冗长的方案以及以昂贵的重组蛋白为主的试剂相比,将优选实惠的细胞产生法,并且细胞的冷冻保存将利于日后加工。这里,我们采用了3个小分子(y27632、sb431542、和purmorphamine)和2个重组蛋白(bmpria
‑
fc和dkk1)。对于bmp和wnt路径的抑制,目前存在多个小分子替代品。我们发现dorsomorphin(bmp路径抑制剂)和cki
‑
7(wnt路径抑制剂)在b27 5f方法中可替代蛋白(kim,d.s.,等(2010).stem cell rev 6,270
‑
2810)(osakada,f.,等(2009).j cell sci 122,3169
‑
3179)。虽然细胞生存力减少~50%,但是nkx2.1
‑
gfp 效率是相当的(未显示)。此外,在冷冻保存和解冻后~25%的hesc源性的mge
‑
样细胞是活的,并且成熟为神经元的解冻细胞具有通过电生理证实的功能性质。
[0324]
对于临床应用,需要gmp
‑
级hpsc谱系。因此,我们检查了cgmp
‑
匹配的hesc谱系(esi17、esi35、esi51、和esi53(biotime))的生成nkx2.1 mge
‑
样细胞的能力。所有谱系在利用b27 5f方法的nkx2.1 分化效率方面类似于hes
‑
3(参见实施例8)。因为临床级谱系不具有nkx2.1敲入报道子,所以日后工作需要确定可接受的杂质阈值。hesc源性的nkx2.1 细胞的75%纯度可能足矣,尤其如果其余杂质表示非
‑
mge
‑
型gaba能中间神经元。建议此可能性,我们观察到大多数hes3 nkx2.1
‑
gfp
‑
阴性神经元表达gaba,并且一些表达th,但没有表达tbr1或chat刺激性神经元标记物。为得到mge
‑
特异性细胞标记物的nkx2.1 mge
‑
样细胞抗体、dna质粒、rna、或病毒的进一步纯化的组合物以递送选择mge细胞或中间神经元的转基因(可选择的标记物、荧光、抗生素抗性、基因过表达/抑制)(potter,g.b.,等(2009).mol cell neurosci 40,167
‑
186),可使用以facs
‑
/磁性macs
‑
为基础的纯化,或抗有丝分裂化合物(例如arac或mitoc)。
[0325]
慢病毒制备
[0326]
自失活的慢病毒质粒fugw
‑
ubc
‑
rfp(rfp二聚体2)或plenti
‑
突触蛋白
‑
hchr2(h134r)
‑
eyfp
‑
wpre(来自karl deisseroth的慷慨赠品)与δ8.9和vsvg质粒共转染进入239t细胞(atcc)中。收集慢病毒颗粒上清液,利用超速离心进行浓缩,并用于利用8μg/ml聚凝胺(millipore)转导细胞过夜。
[0327]
实施例1:hesc源性的端脑mge
‑
样特性
[0328]
为利于识别hpsc源性的nkx2.1 mge前体细胞,我们使用具有插入nkx2.1的第二外显子中的gfp敲入构建体的hes
‑
3hesc谱系(hes
‑
3 nkx2.1
gfp/w
)(goulburn,a.l.,等(2011).stem cells 29,462
‑
473)。nkx2.1表达标记发育的神经系统中的腹侧前脑
‑
特异性特性,包括端脑mge、视前区(poa)、隔膜、和间脑下丘脑。与双
‑
smad(sb431542和bmpria
‑
fc)和wnt
抑制(dkk1)组合,我们发现早期shh路径活化(purmorphamine)和b
‑
27补充允许自hesc的高效及可重现的腹侧前脑
‑
样分化。分化后的第20
‑
30天的平均nkx2.1
‑
gfp 效率为74.9
±
2.1%(n=25个独立分化试验)。我们也发现额外的hpsc谱系,研究级cgmp
‑
匹配的esi
‑
17、35、51、和53(biotime)、和h9(wicell)、hesc谱系、和成人黑素细胞源性的hipsc谱系,在类似效率下分化成nkx2.1 mge前体细胞。优化的b27 五个因子(b27 5f)方法在图1a,图8中给出。mge前体细胞也称为mge
‑
样祖细胞。
[0329]
为确定nkx2.1
‑
gfp 细胞的区域特性,我们进行50天的悬浮拟胚体(seb)分化过程并对前脑发育的标记物进行免疫染色分析(图1b和9)。我们通过10天的分化检测到稳健的nkx2.1
‑
gfp表达,其与nkx2.1蛋白共存并在整个过程期间表达。相比之下,没有检测到pax6(背部端脑神经祖细胞的标记物)。到第20天在大多数细胞中发现端脑标记物foxg1并保持高表达。与此趋势相反,nkx2.2表达(下丘脑的标记物和更多腹尾区域),主要只在第10
‑
20天之间表达。到第20
‑
30天,额外的腹侧端脑祖细胞标记物olig2和ascl1(mash1)被诱导(图9)。islet1在侧ge(lge)和poa中强烈表达,并在mge内的分散细胞中表达;其在后期(d30
‑
40)时间点被诱导。到第30天,细胞表达迁移的神经元标记物、双皮质素(dcx)、和神经传递素gaba。我们在整个过程期间没有检测到下丘脑标记物rax或胆碱能神经元标记物chat。因此,hesc源性的nkx2.1
‑
gfp 细胞看起来表示端脑mge
‑
样祖细胞和gaba能神经元谱系。提供了关于在seb培养期间标记物表达的总结(图15)。
[0330]
图1.hesc源性的端脑mge
‑
样中间神经元前体细胞(也称为mge前体细胞)。(a)概述b27 5f方法用于产生mge前体细胞和从其源自的gaba能中间神经元,以及相应图。缩写:seb=悬浮拟胚体;aeb=粘附拟胚体;ml=单层;facs=荧光活化细胞分类;y27632=rho
‑
相关激酶(rock)抑制剂;sb431542=tgfβ1激活素受体
‑
样激酶的抑制剂;bmpria=骨形态发生蛋白受体1a fc嵌合体;dkk1=dickkopf同源物1;pm=purmorphamine;bdnf=脑源性神经营养因子;dapt=γ
‑
分泌酶的抑制剂。也参见图8。(b)seb剖面和典型的免疫荧光分析显示了nkx2.1
‑
gfp、pax6、foxg1、和nkx2.2表达。蓝色:dapi。比例尺:100μm。也参见图9和图15。
[0331]
图8.分化的hesc的facs分析显示对先前公布的方案的修改以及通过早期shh路径活化和b27培养基补充的组合来诱导nkx2.1
‑
gfp 细胞,这与图1相关。(a)方法#1a:ksr
‑
n2
‑
b27补充的培养基连同后期(第25天)pm。(b)方法#1b:ksr
‑
b27培养基连同后期(第25天)pm处理。(c)方法#1/2混合:ksr
‑
b27培养基连同中期(第10天)pm处理。(d)类似于(c)但连同早期(第0天)pm处理。(e)类似于(d)但连同b27
‑
b27培养基以得到70%gfp 效率。(f)类似于(e):b27
‑
b27培养基以及早期(第0天)pm添加,但在第14天移除抑制剂并进行pm处理到第35天以得到高达90%gfp 效率。着色柱状图=分化培养物。空白柱状图=未分化培养物对照。
[0332]
图9.hesc源性的端脑mge
‑
样特性和gaba能神经元命运,与图1和图15相关。对seb切片的额外免疫染色分析表明腹侧端脑标记物olig2和ascl1(早期)、islet1(后期)、和有丝分裂标记物ki67(整个期间)的诱导。迁移的神经元标记物dcx和gaba随时间诱导。相比之下,在分化50天后,没有检测到下丘脑标记物rax和胆碱能神经元chat。蓝色=dapi。
[0333]
图15.在自hesc的悬浮拟胚体分化期间标记物表达的总结,与图1和图9相关。
[0334]
实施例2:hesc源性的mge前体细胞表现出vz和svz放射状胶质
‑
样干细胞行为
[0335]
胚胎神经发育的定义性特点为顶面
‑
底侧极性的获得以及放射状胶质神经干细胞的发育(kriegstein和gotz,2003)。有证据表明hesc源性的神经玫瑰花状结构中的神经上
皮祖细胞表示顶面
‑
底侧
‑
样极性(elkabetz,y.,等(2008).genes dev 22,152
‑
165)。当在第4
‑
7天将seb整块接种时,粘附eb(aeb)变平并揭示了nkx2.1
‑
gfp 细胞以玫瑰花状结构的组织(图2a和2b)。n
‑
钙黏蛋白表达受限于玫瑰花状结构腔面,证实了极性,并与定位于胚胎脑中心尖室表面上放射性胶质脚板一致。ki67在多种细胞中表达,尤其玫瑰花状结构腔附近的那些。相比而言,远离腔检测到神经元标记物ascl1和dcx(图2b)。
[0336]
玫瑰花状结构标记有rfp[泛素c启动子
‑
rfp(ubc
‑
rfp)病毒]并利用活体时移成像来监测。我们检测到玫瑰花状结构中的nkx2.1
‑
gfp 和ubc
‑
rfp 细胞,其在分裂前表现出脑室区(vz)放射性胶质(vrg)
‑
样动力间胞核迁移(inm)行为(图2c和2e)。vrg
‑
样细胞体移向玫瑰花状结构腔并利用垂直卵裂面(平行于纤维)分开。有意思的是,子细胞看起来扩展放射性纤维,类似归因于胚胎vrg的对称分裂,该胚胎vrg利用垂直卵裂面分开。
[0337]
我们随后研究了最近描述的(fietz,s.a.,等(2010).nat neurosci 13,690
‑
699)(hansen,d.v.,等(2010).nature 464,554
‑
561)人
‑
富集的外亚
‑
vz(svz)放射性胶质(org)
‑
样细胞是否存在于我们的培养物中。我们集中在单极纤维远离玫瑰花状结构簇的nkx2.1
‑
gfp 细胞(图2d和2f)。我们发现gfp org
‑
样细胞在分裂前表现出有丝分裂胞体位移(mst)。这些细胞体移向单极纤维并利用水平卵裂面(垂直于纤维)分裂。总之,hesc源性的mge
‑
样祖细胞(也称为mge前体细胞)可概括vz和人
‑
富集的svz放射性胶质
‑
样干细胞行为。
[0338]
图2.hesc
‑
mge
‑
样祖细胞(也称为mge前体细胞)表现出vz和svz放射性胶质干细胞
‑
样分裂。(a)在第4
‑
7天整块接种seb,在第14天固定aeb以分析;或利用ubc
‑
rfp病毒和时移成像的活培养物感染aeb。(b)第14天nkx2.1
‑
gfp表达并以整合并分开的方式用红色显示标记物的平面:n
‑
钙黏蛋白、ki67、ascl1、和dcx。蓝色:dapi。比例尺:50μm。(c)仅具有rfp荧光或与nkx2.1
‑
gfp融合的rfp荧光的玫瑰花状结构簇。(d)表达簇外细胞的nkx2.1
‑
gfp。c,d比例尺:100μm。(e)显示了三个rfp 细胞(蓝色,橙色,和绿色箭头)的盒状区域(c)的时移成像系列表明vrg
‑
样inm行为:移向玫瑰花状结构腔并利用垂直卵裂面分裂(星状)。时间:小时。比例尺:20μm。(f):盒状区域(d)的时移成像系列。具有特征性单极形态(白色箭头和更小箭头用于标记纤维)的gfp 细胞展现了org
‑
样mst行为:移向纤维(46μm)并通过水平卵裂面分裂(星状)。时间:小时。比例尺:50μm。
[0339]
实施例3:hesc源性的mge
‑
样祖细胞生成的gaba能中间神经元
[0340]
我们进一步探索hesc源性的培养物的特性和命运。对d25 aeb进行免疫染色分析(图3a和3b)。也使第25天的aeb解离,以单层(ml)培养,并进行免疫染色分析并在第35天定量(图3c
‑
3e)。在25天和35天阶段,nkx2.1
‑
gfp表达对表达内源性nkx2.1蛋白的细胞具特异性(91.3
±
1.7%)(图3e)。在第25天,foxg1和olig在大多数nkx2.1
‑
gfp 细胞中表达,提供了腹侧端脑
‑
样特性的另一个证据(图3b)。到第35天,大多数gfp 细胞继续表达foxg1(81.5
±
3.6%)但下调了olig2(6.8
±
3%)。大多数gfp 细胞到第35天也表达ascl1(79.9
±
6.5%)和dlx2(79.8
±
3.7%)。因为olig2标记ge祖细胞而dlx2标记gaba能神经元谱系,所以这些结果表明第25天的hesc源性的mge
‑
样祖细胞(文中也称为mge前体细胞)到第35天开始分化成gaba能神经元。的确,通过tuj1(92
±
2.4%)染色证实神经元特性(图3d和3e)。大多数gfp /tuj1 细胞共表达gaba(75.8
±
2.3%)。中间神经元亚型标记物钙结合蛋白(calbindin)(calb1或calb)到第35天也表达(31.1
±
5.4%),但是其它中间神经元亚型标
记物钙网膜蛋白(calretinin)(calb2或calr)、生长抑素(somatostatin)(sst)、和小清蛋白(parvalbumin)(pvalb或pv)在此阶段没有检测到,除了对于sst或calr的极少情况。
[0341]
少数gfp 细胞表达间脑/少突细胞标记物nkx2.2(13.6
±
4.7%)、神经祖细胞/神经胶质细胞标记物gfap(3.9
±
3.9%)和ki67(2.8
±
1.5%)、lge/poa
‑
富集的标记物islet1(7.6
±
3.3%)、或多巴胺能神经元标记物th(4.4
±
1.3%)(图3)。基本上没有gfp 细胞表达新皮层标记物pax6、尾部ge(cge)/背部mge标记物couptfii、纹状体中度棘神经元标记物darpp32、苍白球投射神经元标记物er81/etv1、胆碱能神经元标记物chat、或谷氨酸能新皮层投射神经元亚型标记物tbr1(图3e)。基于这些结果,hesc源性的mge
‑
样祖细胞看起来已主要地分化成有丝分裂后gaba能中间神经元。
[0342]
图3.hesc
‑
mge
‑
样祖细胞分化成具有端脑gaba能中间神经元的性质的神经元。(a)aeb固定用于第25天的免疫荧光染色。(b)第25天mge
‑
样祖细胞表达nkx2.1
‑
gfp、nkx2.1、foxg1、olig2、ascl1、和dlx2。蓝色:dapi。比例尺:50μm。(c)aeb解离,以ml重新接种,并固定用于第35天的免疫荧光。(d)第35天的解离细胞表达nkx2.1
‑
gfp、tuj1、ascl1、gaba、和calb。蓝色:dapi。比例尺:50μm。(e)第35天免疫染色的定量。大多数nkx2.1
‑
gfp 细胞表达nkx2.1、foxg1、ascl1、tuj、gaba、和dlx2。数据表示成平均值
±
sem。
[0343]
实施例4:hesc源性的nkx2.1
‑
gfp 微阵列分析
[0344]
我们进行微阵列分析以得到未分化hesc和facs
‑
分类的nkx2.1
‑
gfp 群体在55天分化期间的整体转录组对比:自20天aeb(蓝色柱条)或35天粘附单层(橙色柱条)、或自标记有ubc
‑
rfp病毒、在第35天分类gfp,并与神经胶质细胞共培养的55天培养物(绿色柱条)(图4a)。nkx2.1
‑
gfp 细胞的百分比在解离的单层培养物中(在第35天~81%)或在共培养中(在第55天~94%rfp 细胞)保持在高水平(图4b)。树状图聚类分析显示了分化的gfp 群体比未分化的hesc彼此更密切相关(图10a)。为了更详细地检查hesc源性的gfp 群体的特性,我们选择了谱系特异性标记物组合并随时间评价转录杂交强度(图4c
‑
4h及10c
‑
10f)。对于标记物的子集,进行定量rtpcr并证实阵列数据(图10b)。只在未分化hesc中检测到多能标记物,而诱导神经谱系标记物(hes5、dcx、syp、syn1)分化gfp 细胞。没有检测到神经胶质细胞、神经嵴、或小神经胶质的标记物。相比而言,gfp 细胞表达神经元标记物(tubb3、dcx、syp、syn1),且转录水平随时间增加。然后我们测试了前后中枢神经系统(cns)模式并检测了前cns标记物(foxg1、six3、otx2)的表达。与先前结果一致,间脑(以及近尾部)nkx2.2短暂表达并随后下调。意外地,cns底板标记物foxa2在第20天祖细胞中表达但在后期时间点没有被检测到。
[0345]
我们随后研究了识别前脑的亚区的标记物。除了nkx2.2的暂时表达,间脑下丘脑的标记物没有稳健表达(图10c)。同样,没有检测到背部刺激性神经元谱系标记物(图10d)。反而,表达腹侧端脑gaba能神经元谱系标记物(ascl1、dlx1、dlx5)和mge标记物(nkx2.1、lhx6、lhx8、cxcr7),并且它们的表达强度一般随时间增加(图4f和4h)。没有检测到非
‑
mge腹侧端脑的标记物,也没有检测到nkx2.1 poa/隔膜或苍白球的标记物(图10e)。发现gaba能标记物(gad1、slc32a1、slc6a1),但谷氨酸能或胆碱能神经元标记物没有表达(图4g)。我们检测到多巴胺能神经元转录(th)但没有识别出表达th蛋白的多个神经元(图3e)。这些结果表明gfp 细胞属于主要的mge
‑
样gaba能神经元谱系。
[0346]
除了皮层gaba能中间神经元,mge产生纹状体gaba能中间神经元和苍白球的gaba
能投射神经元。因为gfp 细胞不表达苍白球标记物转录物或蛋白,所以我们进一步评价皮层/纹状体中间神经元谱系的标记物。
[0347]
纹状体中间神经元保持nkx2.1和lhx8表达,但迁移的皮层中间神经元消除这些标记物并表达zeb2、maf、arx、cxcr7、和cxcr4。支持皮层
‑
样中间神经元谱系,在gfp 细胞中检测到稳健的cxcr7表达。虽然在后期发现增强的zeb2、arx、和cxcr4转录信号,但是整体水平是适中的,nkx2.1和lhx8继续表达,表明不成熟阶段的纹状体中间神经元
‑
样谱系和/或皮层
‑
样谱系(图4f和4h)。最后,在标记中间神经元的亚型的神经肽和钙结合蛋白中,只有sst转录物在第55天被稳健地检测到(图4h)。总之,hesc源性的nkx2.1
‑
gfp 群体表示mge
‑
样神经祖细胞和gaba能皮层
‑
和/或纹状体
‑
样中间神经元。
[0348]
图4.hesc
‑
mge
‑
样nkx2.1
‑
gfp 细胞群体的微阵列基因表达图谱。(a)微阵列数据的示意图和图注。未分化的hesc(黑色);以及自第20天aeb的经facs
‑
分类的gfp 细胞(蓝色),第35天ml培养物(橙色),以及自第35天共培养到第55天的gfp 细胞(绿色)。(b)每个分化阶段和未分化hesc对照(黑色)的典型的facs柱状图分析。(c
‑
h)标记物组合的平均转录杂交信号强度。in=中间神经元,da=多巴胺能,ach=胆碱能,glu=谷氨酸能。数据表示成平均值
±
sem。也参见图10。
[0349]
图10.hesc源性的nkx2.1
‑
gfp 细胞的转录表达图谱,与图4相关。
[0350]
(a)微阵列数据的树状图聚类分析确定第20天、第35天、和第55天gfp 群体比未分化hesc彼此更密切相关。
[0351]
(b)未分化hesc(黑色柱条)和3周gfp 细胞(蓝色柱条)相对于gapdh表达的定量rtpcr分析。数据表示成平均值
±
sd。
[0352]
(c
‑
f)来自微阵列分析的额外标记物。图注:未分化的hesc(黑色)、第20天gfp (蓝色)、第35天gfp (橙色)、和第55天gfp (绿色)样品。组合显示:下丘脑(c)、皮层刺激性神经元谱系(d)、腹侧端脑(e)、和一般胎儿发育标记物(f)。gp=苍白球、poa=视前区、sep=隔膜、pn=投射神经元。没有检测到下丘脑和皮层刺激性神经元标记物。也没有检测到poa/sep、gp、和mge源性的pn标记物、etv1和gbx2。以低水平确定背部mge和cge标记物nr2f2(couptfii),与表达couptfii蛋白的稀有gfp 细胞(图3e)一致,并在第20天也微弱地检测到nkx6
‑
2。早期胚胎标记物dppa4、lin28、和lin28b用于评价源自人多能干细胞的神经细胞的发育阶段(patterson,m.,等.(2012).cell res 22,178
‑
193)。在人胚胎脊髓中,lin28表达下调了7gw,而dppa4和lin28b直到13gw才下降。在我们的培养基中,到第35天检测不到dppa4和lin28,lin28b表达持续到第55天。这些结果表明,第35
‑
55天的gfp 细胞可能类似于7
‑
13gw胚胎发育阶段。数据表示成平均值
±
sem。
[0353]
实施例5:hesc源性的mge
‑
样细胞向gaba能中间神经元的亚型的延长的成熟
[0354]
我们随后更有力地试图确定由nkx2.1
‑
gfp mge
‑
前体样细胞所生成的神经元亚型和亚型成熟的发育时间表。为研究它们的成熟,使第35天的经facs
‑
分类的gfp 细胞与皮层胶质细胞共培养(图5a),在共培养前,使一些细胞标记有ubc
‑
rfp病毒。在分化后的5、10、20、和30周(wpd)后,使培养物固定以用于分析,并定量神经元亚型标记物表达(图5c)。在30wpd后,rfp hesc源性的神经元在神经元体细胞尺寸上明显更大并表达gaba能神经元
‑
特异性标记物vgat和中间神经元亚型标记物sst、calb、和calr(图5b)。10
‑
20
‑
wpd培养物的图像显示在图11中。在5个wpd下几乎所有神经元表达nkx2.1
‑
gfp,但是,类似于皮层中间神经
元,nkx2.1 神经元的百分比到30wpd明显下降(66.7
±
6.1%;p=0.03)。大多数神经元自5
‑
30wpd表达gaba(75
‑
86%)和vgat(53
‑
78%)。除了稀有细胞(3,110个神经元中的11个),刺激性神经元标记物tbr1没有表达。calb在整个时程中在神经元中表达(24
‑
36%)。相比而言,sst 和calr 神经元的百分比随时间增加并从10到20wpd显著下降(sst:2.8
±
1%至12.8
±
9%;p=0.03,以及calr:8.8
±
4.9%至52.6
±
6.2%;p=0.004)。到30wpd,sst 神经元的百分比增至40.6
±
8.6%,calr 神经元增至77.7
±
14.9%。相反,到30wpd,检测不到pv 神经元(1,146个神经元中0个),并且npy 神经元罕见(819个神经元中6个,未显示)。因此,在10wpd与20wpd之间,nkx2.1
‑
gfp mge
‑
样细胞成熟为表达calb、calr、和/或sst亚型标记物的nkx2.1 和nkx2.1
‑
阴性gaba能中间神经元,以及sst和calr亚型明显成熟。
[0355]
令人惊讶的是,我们的hesc源性的gaba能中间神经元需要20
‑
30周来显示sst和calr的实质表达。但是,此延长的分化时间表类似于人胎儿和婴儿中间神经元亚型发育(fertuzinhos,s.,等(2009).cereb cortex 19,2196
‑
2207)。我们利用我们自己的关于发育的人皮层和mge的组织学分析证实这些发现(图12a和12b)。因此,hesc源性的mge
‑
样细胞成熟平行于内源性和培养的mge发育:相当的中间神经元亚型以类似序列和时帧生成。
[0356]
图5.hesc
‑
mge
‑
前体样细胞源性的gaba能中间神经元成熟和放电性质。
[0357]
(a)解离的ml培养物感染有ubc
‑
rfp慢病毒,在gfp 细胞第35天经facs
‑
分类,并共培养。
[0358]
(b)30周培养物的免疫染色显示表达vgat、sst、calb、和calr的高分支rfp 人神经元。比例尺:50μm。
[0359]
(c)在5、10、20、及30周内的免疫染色分析的定量。数据表示成平均值
±
sem。也参见图11和12。
[0360]
(d)12和30wpd的hesc源性的神经元的dic图像,插图显示记录的神经元的rfp表达。比例尺:20μm。
[0361]
(e)统计结果显示膜电阻(rm)、膜静电位(rmp)、膜电容(cm)、和动作电位(ap)1/2
‑
宽度。
[0362]
(f)近阈值(上限)和超阈值(下限)电流注入后,每个阶段的典型的ap放电情况。比例尺:50mv和100ms。也参见图13。
[0363]
(g)阈值电流注入后,平均第一ap痕量。比例尺:25mv和25ms。
[0364]
(h)统计结果显示每个阶段的ahp(虚线=基线)。
[0365]
(i
‑
j)在每个阶段的na
(i)和k
电流(j)的i
‑
v曲线,在阶梯式电压下测量(持续500ms)。e,h,i,j:数据表示成平均值
±
sem。***表示p<0.001。
[0366]
图11.hesc源性的mge前体
‑
样细胞向gaba能中间神经元亚型的成熟,与图5相关。
[0367]
预先标记有ubc
‑
rfp病毒,在第35天经facs
‑
分类gfp,并分化后共培养10及20周(wpd)的nkx2.1
‑
gfp 细胞的免疫染色分析。到10wpd,hesc源性的神经元表达vgat和钙结合蛋白(calb1),并且罕见细胞表达钙网膜蛋白(calb2)或sst。到20wpd,人神经元表达vgat、calb1、calb2、和sst。在任何时间点都检测不到小清蛋白(pvalb)。蓝色=dapi。
[0368]
图12.中间神经元亚型在源自mge的培养物中的发育,与图5相关。
[0369]
(a)calb和calr在所有样品中表达,但sst和pv分别在24gw和8mo中,以及只在8mo样品中表达。蓝色=dapi。
[0370]
(b)calb和calr表达并且与nkx2.1共存。蓝色=dapi。
[0371]
(c)rfp 细胞表达calb、calr、sst、和gaba,但不表达tbr1。蓝色=dapi。
[0372]
实施例6:hesc源性的mge
‑
样细胞的功能成熟:中间神经元
‑
样放电性质,突触形成,以及gaba能输出
[0373]
为检测hesc源性的细胞是否为功能神经元,我们进行全细胞膜片记录以检查它们在不同wpd下的电生理性质(8周,n=21;12周,n=35;15周,n=31;30周,n=18)。我们发现hesc源性的神经元的动作电位(ap)放电模式在8wpd下非常不成熟,这通过第一ap的宽ap 1/2
‑
宽度、小后超极化(ahp)、以及不能在高电流注入后重复发电进行判断(图5e
‑
5h)。峰值电压
‑
门控的na 和k 通道电流从8到12wpd明显增加(图5i和5j),伴随发生膜电阻(rm)的显著下降(图5e)。多个神经元表明在12和15wpd时近阈值电流注入时更成熟的重复的ap放电(图5f、13b和13c)。到30wpd,hesc源性的神经元表现出在超阈值电流注入时高频的重复ap放电(图5f和13d),以及膜电容(cm)的相应增加以及更超极化膜静电位(rmp)(图5e)。此外,30
‑
wpd神经元表现出更小的ap 1/2
‑
宽度(图5e)和更大的ahp(图5g和5h)。与这些更成熟的生物物理性质一致,我们也注意到相比早期的12wpd阶段,在30wpd的利用多次长处理的hesc源性的神经元的更成熟的形态(图5d)。
[0374]
然后,我们通过研究它们与小鼠神经胶质细胞共培养时的突触性质研究了mge
‑
样细胞是否为gaba能神经元。hesc源性的nkx2.1
‑
gfp 神经元过程与点状突触前vgat表达共区域,表明gaba能突触的形成(图6a)。自发突触后电流(spsc)通过分化后八周来检测并通过gabaa受体抑制剂甲碘荷包牡丹碱(bmi,20μm)完全阻断,表明功能性gaba能
‑
特异性突触形成(图6b)。接收spsc的神经元的百分比从8wpd的33.3%(n=12)增到12wpd的82.1%(n=28,图6c)。为了证实gaba能神经元能发送产出到相邻神经元,我们利用突触蛋白启动子
‑
光敏感通道2
‑
eyfp(chr2
‑
eyfp)通过慢病毒感染转染一半的神经元。蓝光刺激在eyfp
‑
阳性神经元中稳妥地引起工作电位放电(图13f)(weick,j.p.,等(2011).proc natl acad sci usa 108,20189
‑
20194),并在相邻神经元中诱发产生稳健的突触后电流(psc)(图6d和6e)。此外,psc显示出长衰变时间(31.4
±
1.9ms,n=26),此为gaba能psc的特征。通过bmi的光
‑
诱发的psc的可逆性阻断进一步对此证实(图6d和6e)。光诱发psc的逆转电位为
‑
32.7mv(图6f和6g),接近于在我们记录的条件下的预期的cl
‑
逆转电位[
‑
37.3mv=
‑
53.4mv(通过能斯特方程) 16.1mv(接头电位)]。这些结果表明hesc源性的mge
‑
样中间神经元只产生gaba能突触输出。
[0375]
得到rfp
‑
阴性原发性皮层神经元的全细胞记录(图6h)。hesc源性的神经元的蓝光刺激在被bmi完全阻断的记录的原发性神经元中诱导gaba能
‑
特异性psc(图6i和6j)。并且,我们发现光刺激后的多突触反应(图6i),这些也被bmi阻断,表明hesc源性的神经元在培养的神经元回路中的稳健的突触整合。因此,hesc源性的神经元表明功能性神经元性质,gaba能
‑
排他性突触输出,以及中间神经元放电性质的缓慢30
‑
周成熟,这与亚型标记物表达的缓慢节奏一致。
[0376]
图6.hesc源性的中间神经元的gaba能突触性质。
[0377]
(a)图像显示12wpd的hesc源性的nkx2.1
‑
gfp 神经元中的vgat表达。右:虚线矩形的放大。比例尺:左,50μm;右,10μm。
[0378]
(b)踪迹显示hesc源性的神经元中的自发突触后电流(psc),底端:psc被bmi完全
阻断。比例尺:100pa,中间踪迹的5s和0.25s(虚线)。
[0379]
(c)神经元的百分比显示不同阶段的自发psc。
[0380]
(d)利用chr2
‑
eyfp转染hesc源性的神经元。踪迹表明蓝光脉冲(蓝色柱条)诱发被bmi可逆阻断的相邻细胞中的psc。比例尺:50pa和50ms。也参见图13。
[0381]
(e)光
‑
诱发的gaba能psc的平均振幅及bmi的应用。
[0382]
(f
‑
g)踪迹显示在不同保持电压下的光
‑
诱发的(蓝色柱条)psc。综述性结果(n=7)显示光
‑
诱发的gaba能psc的i
‑
v曲线(g)。
[0383]
(h)合并图像显示与分类的ubc
‑
rfp 和chr2转染的hesc源性的神经元共培养的皮层细胞的dic。比例尺:20μm。
[0384]
(i)踪迹显示在rfp
‑
阴性记录的皮层神经元中hesc源性的神经元
‑
诱发的psc的蓝光(蓝色柱条)刺激。上图版显示psc单
‑
突触反应,下图版显示psc和多
‑
突触反应
‑
都完全被bmi阻断。比例尺:50pa和50ms。
[0385]
(j)光诱发的psc的平均振幅和bmi的应用。e,j:数据表示成平均值
±
sem。
[0386]
图13.hesc源性的中间神经元放电性质的成熟,与图5和6相关。
[0387]
(a
‑
d)分化后不同阶段的动作电位(ap)的实例踪迹。在
‑
60mv至
‑
70mv的v
保持
下在不同阶段将阶梯式电流注入记录的神经元中:(a)8wpd(b)12wpd(c)15wpd(d)30wpd。红色踪迹指示了阈值电流注入时的ap。黑色踪迹指示了在两或三倍阈值电流注入时的ap。比例尺:50mv和200ms.
[0388]
(e
‑
f)在chr2
‑
eyfp阳性hesc源性的神经元中的蓝光引起的ap。10wpd的eyfp荧光和dic的合并图像(e)。比例尺:20μm。蓝光脉冲(蓝色踪迹)在chr2
‑
表达的神经元中稳妥地引起ap(黑色踪迹)(f)。
[0389]
实施例7:在小鼠大脑中hesc源性的mge
‑
样中间神经元成熟和功能整合
[0390]
为了严格评价细胞命运和功能,将hesc源性的mge
‑
样细胞移植到小鼠大脑中,我们修改了我们的方案以避免注入未分化的nkx2.1 神经干细胞(图16)。经dapt的处理用于引起神经元前体的神经元分化组合的psa
‑
ncam纯化,dapt为一种notch信号传导路径的γ分泌酶抑制剂(schmandt,t.,等(2005).stem cells dev 14,55
‑
64)。nkx2.1
‑
gfp 细胞的75.7
±
5.2%(n=12)的平均值对于facs的高psa
‑
ncam表达是阳性(图14a)。将富集gaba能神经元前体(图3e)的来自第35天的nkx2.1
‑
gfp 和psa
‑
ncam 细胞注入严重联合免疫缺陷(scid)的新生小鼠皮层中(图7a)。人
‑
特异性核抗原(hna)阳性人细胞在注射后存活7个月(mpi)(最长时间点),一些人细胞自注射点迁移了1mm以上(图7b、14b和14c)。在两个、四个、及七个mpi后的人细胞存活率(注入细胞的%)分别为5.6
±
2.6%、3.1
±
1.5%、和8.6
±
3.1%。两个mpi后,表达hna和nkx2.1
‑
gfp(67.8
±
1.6%)、ki67(25.5
±
1.7%)、或dcx(79.8
±
3.8%)的人细胞大多数仍位于注射点(图7b和14b)。但是到4mpi,ki67表达明显下降(1.7
±
0.27%;p=0.04),并且dcx表达随时间类似地下降(7mpi时5.9
±
4.9%;p=0.008)。同样,在7mpi后,仅在35.6
±
14%的人细胞中检测到nkx2.1
‑
gfp
‑‑
比在30wpd的共培养物中发现的百分比低。对于有丝分裂后神经元标记物neun发现了反向趋势,其到7mpi时增到68.4
±
8.3%人细胞。相比而言,神经胶质细胞标记物gfap和olig2在7mpi时由较低百分比的人细胞表达(分别11.2
±
4.3%和10.7
±
4.4%)。在注射前,使一些hesc源性的培养物标记有ubc
‑
rfp病毒。在小鼠大脑中7mpi后,发现具有神经元形态的rfp 人细胞表达gaba、sst、
calb、和calr(图7c和7e)。没有检测到pv 人细胞,除了具有微弱信号的稀有细胞外(1,829个细胞中4个)。总而言之,注入小鼠皮层的mge
‑
样gaba能神经元前体主要成熟为表达sst、calr、和calb中间神经元亚型标记物的神经元。
[0391]
为检查hesc源性的mge
‑
样细胞是否能发育成体内突触整合的功能中间神经元,我们进行了小鼠脑片7mpi的rfp 人细胞的全细胞记录。细胞内填充神经生物素和后染色揭示出记录的rfp 神经元的广泛处理分支(图7f)。在来自三只动物的总共17个人细胞中,16个神经元表现出以
‑
64.8
±
4.0mv的平均rmp放电动作电位的能力。此外,识别具有不同膜性质和放电模式的两组中间神经元,i型和ii型。i型中间神经元具有
‑
67.3
±
2.9mv的平均rmp、257
±
78mω的rm、和69.4
±
0.6pf的cm。i型中间神经元的放电模式显示了在阈值下激发明显延迟,以及超阈值电流注入时的小适应(图7g和14d)。ii型中间神经元具有更超极化的rmp(
‑
80.1
±
3.4mv)、更小的rm(91
±
28mω)、和更小的cm(27.75
±
4.6pf)。ii型中间神经元的放电模式表现出在超阈值电流注入时对初始激发的快速适应(图7g和14e)。并且,移植的hesc源性的中间神经元接收含有bmi敏感性gaba能和6
‑
氰基
‑
2,3
‑
二羟基
‑7‑
硝基
‑
喹喔啉(cnqx)敏感性谷氨酸能组分的突触输入(16个中16个)(图7h),表明功能整合到宿主皮层中。
[0392]
图7.在小鼠大脑中hesc源性的mge
‑
样中间神经元前体细胞成熟和功能整合
[0393]
(a)第35天ml培养物经facs
‑
分类nkx2.1
‑
gfp和psa
‑
ncam,并注入新生小鼠皮层中。也参见图16。
[0394]
(b)针对人
‑
特异性hna、gfp、和ki67的在2和7mpi染色的小鼠大脑组织切片。蓝色:dapi。比例尺:200μm。也参见图14。
[0395]
(c)标记有在7mpi时共表达(箭头)neun、gaba、sst、calb、和calr的ubc
‑
rfp的人细胞的组织学分析。蓝色:dapi。比例尺:50μm。
[0396]
(d
‑
e)sst、calb、和calr在2(黑色)、4(橙色)、和7(蓝色)mpi的组织学的定量(e)。数据表示成平均值
±
sem。
[0397]
(f)通过神经生物素的细胞内填充标记的hesc源性的神经元(nb,绿色)。插页:填充的神经元7mpi的rfp荧光。比例尺:20μm;插页5μm。
[0398]
(g)在7mpi时,近阈值(顶)和超阈值(底)电流注入时i型(左)和ii型(右)hesc源性的神经元的ap放电模式的踪迹。比例尺:50mv和100ms。
[0399]
(h)左图:注射后自hesc源性的神经元记录的自发psc的踪迹;右上方:采用缓慢衰减时间的bmi阻断的psc(箭头)以及采用快速衰减时间的其余psc(箭头)通过随后应用cnqx来阻断(右下图)。比例尺:50pa,放大踪迹为2.5s和0.2s(虚线)。也参见图14。
[0400]
图14.在小鼠脑中hesc源性的mge
‑
样中间神经元和亚型放电性的成熟,与图7和图16相关。
[0401]
(a)第35天的facs分析柱状图显示与同型抗体对照(灰色)相比,psa
‑
ncam的大多数gfp 细胞的高表达(红色)。
[0402]
(b)注射后2、4、和7个月(mpi)的nkx2.1
‑
gfp 和psancam hesc源性的mge
‑
样细胞的迁移和成熟的免疫染色分析。在7mpi时,人
‑
特异性核抗原(hna) 人细胞可迁移,下调gfp和dcx,以及上调neun(一种神经元成熟的标记物)。蓝色=dapi。
[0403]
(c)6只小鼠中的hesc源性的mge
‑
样细胞迁移。在2、4、和7mpi的单注射点侧的嘴和
尾皮质区中计数人细胞。在7mpi检测到一些迁移。以注射细胞的百分比制图。
[0404]
(d
‑
g)在7mpi时的i型和ii型hesc源性的中间神经元的放电性质。i型(d)和ii型(e)神经元的近阈值(顶)和超阈值(400
‑
500pa,底)电流注入时的ap放电模式。每个柱(上部踪迹和底部踪迹)表示一个神经元的ap放电模式。顶图:红色踪迹表示阈值ap放电模式;黑色踪迹为2倍阈值ap放电模式。比例尺:50mv和200ms。(f)统计结果显示i型与ii型神经元之间的ap特征的差异。数据表示为平均值
±
sem,学生t检验用于统计比较。*表示p<0.05。(g)超阈值电流注入时ap放电频率的分析。ii型神经元表现出快速适应放电性质。
[0405]
图16.对hesc分化方案优化、动物移植、和肿瘤发病率的总结,与图7和图14相关。
[0406]
实施例8:hesc源性的mge
‑
前体样细胞
[0407]
临床级gmp
‑
匹配的hesc谱系esi17(图17)、esi35(图18)、esi51(图19)、和h9(图20)分化成mge前体样细胞(顶行)以及进一步分化成中间神经元(底行)。
[0408]
图17
‑
20,顶行:利用b27 5f方法使hesc谱系分化,以悬浮拟胚体(seb)到第7天,然后以粘附eb(aeb)培养到第28天。固定培养物用于免疫荧光染色。粘附eb中大多数细胞表现出mge(nkx2.1)、端脑(foxg1)、和神经元特化(ascl1)的标记物的高表达。少数细胞表达腹侧下丘脑(nkx2.2)和少突细胞祖细胞(olig2)的标记物。
[0409]
图17
‑
20,底行:使第28天aeb培养物解离成单细胞,以单层重接种,在具有b27补充物以及具有或不具有bdnf、dapt、shh的神经基础培养基中再培养2周并固定用于类似分析。这些第42天的单层培养物表达nkx2.1,神经元标记物(tuj),并开始表达抑制性神经元标记物(gaba)。没有检测到olig2表达,并只在稀有细胞中发现nkx2.2。
[0410]
实施例9:自天然人多能干细胞的mge前体细胞的衍生
[0411]
典型的引发的hes3(nkx2.1
‑
gfp)hesc表达同质oct4但不在核中同质表达天然干细胞标记物tfe3(图21,顶行)。利用公布方法(gafni和hanna等,nature 2013)将引发的hes3(nkx2.1
‑
gfp)hesc转化成天然hes3干细胞。天然hesc表达几乎每个细胞核中的tfe3(图21,中间行)。利用b27 5f方法的天然干细胞的分化导致hes3 hesc分化成在粘附eb培养的2周至6周表达mge标记物nkx2.1
‑
gfp、nkx2.1、和lhx6的细胞(图21,底行)。
[0412]
天然es细胞比在只具有bfgf的典型培养基中生长的常见人es/ips细胞更加不分化。传统hpsc更等效于比小鼠mpsc晚期的植入后胚胎外胚层,小鼠mpsc更类似于早期植入前内细胞团。天然人es和ips细胞因此更类似于小鼠es细胞。它们的性质(基因表达和试验胚胎学)等效于植入前胚胎。它们可通过转录因子tfe3的表达和核定位,以及通过菌落形态识别。这些性质将天然细胞与传统引发的hpsc区分开。
[0413]
实施例10:mge
‑
富集的增强子序列用于源自体外由hpsc分化的mge前体细胞的中间神经元的筛选和纯化的用途
[0414]
可得到包含多种增强子
‑
报告子转基因的转基因小鼠。在这些小鼠中,报告基因以在前脑中的不同模式和谱系特异性表达,这取决于增强子的dna序列(图22a
‑
d)。
[0415]
基于它们在转基因小鼠前脑中的表达模式,将mge
‑
富集的增强子序列克隆到病毒载体中以驱动荧光受体基因和/或抗生素抗性基因以mge
‑
选择性方式的表达。构建体也包含rex1
‑
抗生素抗性盒以选择和扩展稳定的转基因hpsc(图22,e)。
[0416]
利用慢病毒将驱动mcherry rfp报告基因(i12b
‑
rfp)的基因间dlx1/2 i12b(422)增强子递送到hes3 nkx2.1
‑
gfp hesc系中,并且生成两个稳定的细胞系(#5和#10),如通过
用于mcherry的基因组dna pcr证实(图22,f)。
[0417]
利用b27 5f方法分化修饰谱系,分化3周后,检测到i12b
‑
rfp表达,以及nkx2.1(图22,g)。eb中i12b
‑
rfp 细胞共表达gaba能神经元标记物dlx2和神经元标记物tuj1(图22,h和i)。
[0418]
流式细胞仪分析证实利用b27 5f方法源性的多个细胞共表达nkx2.1
‑
gfp和i12b
‑
rfp(图22,j)。这些培养物利用notch路径抑制剂dapt的处理导致mge源性的中间神经元的双阳性群体明显增加(图22,k)。
[0419]
双阳性(nkx2.1
‑
gfp 和i12b
‑
rfp )mge前体细胞通过facs纯化并移植到scid小鼠皮层中。注射后数月,发现表达nkx2.1
‑
gfp和i12b
‑
rfp的人细胞从注射点分散并整合到周围的啮齿目动物灰白质中,与分化成mge源性的中间神经元的标志性质一致(图22,l)。
[0420]
也利用电生理学分析表达i12b
‑
rfp(图22,m)的培养的hpsc源性的mge源性的中间神经元。记录动作电位的rfp 中间神经元放电重复训练,证实它们的神经元命运(图22,n)。
[0421]
实施例11:利用长期悬浮培养生成mge源性的中间神经元
[0422]
利用b27 5f条件以seb在常氧气(20%氧含量)中分化的hes3 hesc产生表达nkx2.1
‑
gfp和lhx6的mge中间神经元。在培养的第35天分析细胞(图2,a行)。
[0423]
利用b27 5f条件以seb在常氧气中分化的hes3 hesc产生表达nkx2.1
‑
gfp和lhx6的mge中间神经元。在此分化方案中,shh激动剂(purmorphamine)在第21天被移除(与以上的图23,a行相比,其中shh激动剂(purmorphamine)在整个培养过程中存在)。在培养的第35天分析细胞(图23,b行)。
[0424]
使hes3 hesc在具有ksr、n2、和b27补充物(按顺序添加)以及rock、wnt、和smad抑制剂和shh激动剂的gmem和dmem/f12培养基中在高氧气体(40%氧含量)下培养。在培养的第35天分析细胞(图23,c行)。
[0425]
使hes3 hesc在具有ksr、n2、和b27补充物(按顺序添加)以及rock、wnt、和smad抑制剂和shh激动剂以及加入培养基的基质胶(1
‑
2%)的gmem和dmem/f12培养基中培养。使seb在高氧气下保持培养50天以上。在培养的第60天分析细胞(图23,d行)。
[0426]
实施例12:利用bmp和wnt信号传导路径的小分子抑制剂生成mge前体分子
[0427]
以下为用于生成mge前体细胞的bmp和wnt信号传导路径的小分子抑制剂的其它实例。分化方案如实施例1中所述。
[0428]
图24,a:具有分别作为bmp和wnt信号传导路径的抑制剂的bmpr1a和dkk1的b27 5f分化方案,诱导hesc分化成共表达nkx2.1gfp(顶)和foxg1(底)的mge前体细胞。(具体也参见实施例1)。
[0429]
图24,b:具有分别作为bmp和wnt信号传导路径的抑制剂的ldn193189(0.1μm,登记号04
‑
0019(stemgent))和xav939(2μm,登记号3748(tocris))的b27 5f分化方案,诱导hesc分化成共表达nkx2.1gfp(顶)和foxg1(底)的mge前体细胞。
[0430]
图24,c:具有分别作为bmp和wnt信号传导路径的抑制剂的ldn193189(0.1μm,登记号04
‑
0019(stemgent))和iwr1e(3μm,cayman chemical登记号13659)的b27 5f分化方案,诱导hesc分化成共表达nkx2.1gfp(顶)和foxg1(底)的mge前体细胞。
[0431]
图24,d:具有分别作为bmp和wnt信号传导路径的抑制剂的ldn193189(0.1μm,登记号04
‑
0019(stemgent))和iwp2(5μm,stemgent登记号04
‑
0034)的b27 5f分化方案,诱导
hesc分化成共表达nkx2.1gfp(顶)和lhx6(底)的mge前体细胞。
[0432]
图24,e:具有分别作为bmp和wnt信号传导路径的抑制剂的dorsamorphin(1μm,sigma登记号p5499)和(cki)
‑
7(1μm,sigma登记号c0742)的b27 5f分化方案,诱导hesc分化成共表达nkx2.1gfp和nkx2.1的mge前体细胞。
[0433]
bmp和wnt信号传导路径的抑制剂的添加时间如图1a中所述。
[0434]
图24,底图:使用小分子抑制剂,流式细胞仪用于确定nkx2.1gfp mge前体细胞的生成效率。nkx2.1gfp mge前体细胞的生成效率被确定为所分析细胞中多少百分比细胞为nkx2.1gfp mge前体细胞。
[0435]
对于具有分别作为bmp和wnt信号传导路径的抑制剂的bmpr1a和dkk1的b27 5f分化方案,生成效率为81.6%,当用ldn193189和xav939替代bmpr1a和dkk1时,nkx2.1gfp mge的生成效率为86.9%。以ldn193189和iwr替代bmpr1a和dkk1以89.3%的效率生成nkx2.1gfp mge。(图24,底图)。分析30,000至100,000个细胞。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。