1.本发明属于抗体工程领域,具体涉及一种针对冠状病毒的单抗及其应用,特别是涉及一种结合冠状病毒rbd的人源单抗、制备方法及应用。
背景技术:
2.人感染了冠状病毒后常见体征有呼吸道症状、发热、咳嗽、气促和呼吸困难等。在较严重病例中,感染可导致肺炎、严重急性呼吸综合征、肾衰竭,甚至死亡。目前对于新型冠状病毒所致疾病没有特异治疗方法,需根据患者临床情况进行治疗。《新型冠状病毒感染的肺炎诊疗方案(试行第五版)》公布,在重型、危重型病人治疗的其他治疗措施中,可采用恢复期血浆治疗。2月8 日,首期在江夏区第一人民医院开展了3名危重患者的新冠特免血浆治疗,目前连同后续医院治疗的危重病人超过了10人。经临床反映,患者接受治疗12 至24小时后,实验室检测主要炎症指标明显下降,淋巴细胞比例上升,血氧饱和度、病毒载量等重点指标全面向好,临床体征和症状明显好转。围绕康复者血浆中和病毒的治疗目的,细化了临床使用的适应证、禁忌症和不宜使用的情况。康复者血浆主要用于病情进展较快、重症、危重症新冠肺炎患者。原则上病程不超过3周;新冠病毒核酸检测阳性或临床专家判定患者存在病毒血症,在病情急性进展期应当尽早使用。尽管康复患者血浆疗法在临床上取得了一定的成效,然而由于抗原患者血浆来源有限、纯化抗体安全隐患高、特异性抗体效价不稳定。效价高、性能稳定、安全性好的单克隆抗体对于控制新冠状病毒疫情具有良好的应用前景。目前现有文献已经公开或教导了针对新冠病毒rbd 制备保护性中和单抗的报道。利用新冠病毒刺突蛋白rbd产生抗新冠病毒的保护性中和抗体(如:biorxiv,“sars-cov-2and sars-cov spike-rbd structureand receptor binding comparison and potential implications on neutralizingantibody and vaccine development”,20200220)。sars刺突蛋白rbd和新冠病毒刺突蛋白rbd存在交叉中和表位肽,抗sars的单克隆抗体cr3022能够结合新冠病毒刺突蛋白rbd(emerging microbes&infections,9(1):382-385, 20200217)。采用同源建模的方法明确了新冠病毒病毒ctd1/人ace2复合物的蛋白-蛋白相互作用界面的热点和关键残基,筛选靶向ctd1区域与ace2结合表面的候选抑制剂,阻断病毒与人体ace2蛋白的识别与结合。
3.随着抗sars-cov-2疫苗和治疗性抗体研究的持续推进,最近有报道表明部分治疗性抗体用于sars-cov-2治疗时可产生ade现象。所谓的ade,是指抗体依赖的增强作用,某些病毒在特异性抗体协助下复制或感染能力显著增强,在感染过程中会引发更严重病理损伤。通俗的解释就是,抗体不能中和病毒,反而充当了“特洛伊木马”,让病毒感染免疫细胞的能力更强,产生更多的子代病毒,造成严重症状。ade(antibody-dependent enhancement)效应,即抗体依赖的增强效应,最早是在登革热病毒感染过程中被发现的,结合非中和性抗体的登革热病毒可以通过旁路进入巨噬细胞里面进行增殖,然后造成第二次感染,尤其是第二次感染和第一次感染的病毒株不一样的时候,第二次感染的症状更重,发生ade效应。对于冠状病毒,上世纪八十年代初对猫传染性腹膜炎病毒(fipv)的疫苗研究中
发现针对s蛋白的低效价中和抗体反而会加剧症状、导致更严重的死亡。在对人造成感染的冠状病毒中,对sars病毒和 mers病毒的研究发现疫苗接种产生的针对s蛋白的低亲和力抗体,可以介导病毒进入免疫细胞。基于冠状病毒建立感染的分子机理,如果抗新冠病毒 sars-cov-2抗体也会产生ade效应,则将严重影响抗新冠病毒sars-cov-2 疫苗和治疗性抗体的临床应用。然而,对于具体的抗新冠病毒sars-cov-2s 蛋白的中和抗体是否产生ade效应、产生ade效应的分子机制、以及应对措施,目前还未见报道。
技术实现要素:
4.为解决上述问题,本发明从新冠病毒感染后康复人员pbmc中分离冠状病毒rbd特异性记忆b细胞、扩增获得抗体的轻重链可变区序列,进行了瞬时表达,经检测其中至少20个抗体能够以高亲和力特异性结合冠状病毒rbd,并且至少7个抗体能够阻断或抑制新型冠状病毒rbd与宿主受体ace2的结合。进一步研究发现所述能够阻断或抑制新型冠状病毒rbd与宿主受体ace2 结合的7个抗体分子在sars-cov-2感染thp-1细胞和k562细胞的过程中均不产生ade效应,其中两个抗体corn-01、corn-05则能够促进sars-cov-2 进入raji宿主细胞,存在ade效应。通过raji宿主细胞的膜结合型fc受体种类分析抗体产生ade的机理,并进而通过对抗体fc段突变降低或消除了抗体通过fc与宿主细胞fcγrs受体的结合,从而避免了在sars-cov-2感染宿主细胞过程中产生ade效应。具体而言:
5.一方面,本发明提供一种sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于:所述单克隆抗体是从新型冠状病毒感染患者康复期血液样本中分离单个b细胞克隆获得的特异性结合sars-cov-2s蛋白 rbd区的全人源抗体,其对sars-cov-2感染thp-1细胞和k562细胞均不产生ade效应。
6.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体对sars-cov-2感染raji细胞不产生 ade效应。
7.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体在10-10000ng的部分浓度范围内对 sars-cov-2感染raji细胞产生ade效应。
8.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体在50-3000ng的部分浓度范围内对 sars-cov-2感染raji细胞产生ade效应。
9.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体与sars-cov-2s蛋白rbd具有高亲和性,kd值为5.0
×
10-9
m以下。
10.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体阻断aceii与sars-cov-2s蛋白rbd 结合的ic50值小于50nm,优选小于30nm、25nm、20nm、15nm或10nm。
11.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体fc段的点突变改变了其与受体的结合。
12.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体fc段的点突变降低或消除了其与fc γrs的结合。
13.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体fc段的点突变包括在由第234位、第 235位组成的组中任意一个或两个位点进行氨基酸的替换、缺失或插入突变。
14.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体fc段的点突变包括l234a、l235a 突变。
15.优选的,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其特征在于所述单克隆抗体的重链恒定区具有seq id no:44所示的序列。
16.在一个具体的实施例中,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其具有:
17.选自seq id no:1、7、13、19、25、31、37的hcdr1;
18.选自seq id no:2、8、14、20、26、32、38的hcdr2;
19.选自seq id no:3、9、15、21、27、33、39的hcdr3;
20.选自seq id no:4、10、16、22、28、34、40的lcdr1;
21.选自seq id no:5、11、17、23、29、35、41的lcdr2;
22.选自seq id no:6、12、18、24、30、36、42的lcdr3。
23.更进一步,本发明所述sars-cov-2s蛋白-aceii受体结合阻断性单克隆抗体或其片段,其具有:
24.seq id no:1所示hcdr1、seq id no:2所示hcdr2、seq id no:3所示 hcdr3,seq id no:4所示lcdr1、seq id no:5所示lcdr2、seq id no:6 所示lcdr3;
25.seq id no:7所示hcdr1、seq id no:8所示hcdr2、seq id no:9所示 hcdr3,seq id no:10所示lcdr1、seq id no:11所示lcdr2、seq id no:12 所示lcdr3;
26.seq id no:13所示hcdr1、seq id no:14所示hcdr2、seq id no:15 所示hcdr3,seq id no:16所示lcdr1、seq id no:17所示lcdr2、seq id
ꢀꢀ
no:18所示lcdr3;
27.seq id no:19所示hcdr1、seq id no:20所示hcdr2、seq id no:21 所示hcdr3,seq id no:22所示lcdr1、seq id no:23所示lcdr2、seq id
ꢀꢀ
no:24所示lcdr3;
28.seq id no:25所示hcdr1、seq id no:26所示hcdr2、seq id no:27 所示hcdr3,seq id no:28所示lcdr1、seq id no:29所示lcdr2、seq id
ꢀꢀ
no:30所示lcdr3;
29.seq id no:31所示hcdr1、seq id no:32所示hcdr2、seq id no:33 所示hcdr3,seq id no:34所示lcdr1、seq id no:35所示lcdr2、seq id
ꢀꢀ
no:36所示lcdr3;或
30.seq id no:37所示hcdr1、seq id no:38所示hcdr2、seq id no:39 所示hcdr3,seq id no:40所示lcdr1、seq id no:41所示lcdr2、seq idno:42所示lcdr3。
31.第二方面,本发明提供一种多核苷酸,其编码本发明前述的单克隆抗体或其片段。
32.第三方面,本发明提供一种核酸构建体,其包含本发明前述的多核苷酸。
33.优选的,本发明所述核酸构建体用于表达本发明前述单克隆抗体或其片段。
34.第四方面,本发明提供一种宿主细胞,其包含本发明前述的多核苷酸或本发明前述的核酸构建体。
35.第五方面,本发明提供一种组合物,其包含一种或多种选自由本发明前述单克隆抗体或其片段组成的组中的单克隆抗体或其片段,以及任选的药学上可接受的载体。
36.优选的,本发明所述的组合物,其包含选自由权利要求1至13中任一项所述单克隆
抗体或其片段组成的组中的任意两种、三种、四种、五种、六种、或七种单克隆抗体或其片段。
37.第五方面,本发明提供本发明前述任一项所述单克隆抗体或其片段、本发明前述的多核苷酸、本发明前述的核酸构建体、本发明前述的宿主细胞、本发明前述的组合物在制备预防或治疗sars-cov-2感染药物中的应用。
38.优选的,本发明所述的应用,其特征在于所述预防或治疗sars-cov-2感染包括减少或降低sars-cov-2感染的风险、缓解sars-cov-2感染相关疾病(例如covid-19)的症状、缩短sars-cov-2感染相关疾病(例如covid-19) 的病程、促进sars-cov-2感染相关疾病(例如covid-19)的康复、减少 sars-cov-2感染导致的死亡。
39.第六方面,本发明提供一种消除抗体在sars-cov-2感染宿主细胞时ade 效应的方法,其中,所述的抗体为抗sars-cov-2s蛋白的中和性抗体,通过对抗体fc段进行点突变改变了抗sars-cov-2s蛋白抗体与其受体的结合。
40.优选的,本发明所述消除抗体在sars-cov-2感染宿主细胞时ade效应的方法,其中所述fc段的点突变降低或消除了抗sars-cov-2s蛋白抗体与fc γrs的结合。
41.优选的,本发明所述消除抗体在sars-cov-2感染宿主细胞时ade效应的方法,其中所述fc段点突变包括在由第234位、第235位组成的组中任意一个或两个位点进行氨基酸的替换、缺失或插入突变。
42.优选的,本发明所述消除抗体在sars-cov-2感染宿主细胞时ade效应的方法,其特征在于所述抗sars-cov-2s蛋白的中和性抗体选自本发明前述任一项所述单克隆抗体或其片段。
43.在一个具体的实施例中,本发明提供一种特异性结合sars-cov-2s蛋白的抗体,其中,所述抗体是在天然人源抗sars-cov-2s蛋白抗体的基础上进行fc段点突变,避免了天然人源抗sars-cov-2s蛋白抗体用于阻止 sars-cov-2感染宿主细胞时的ade效应(抗体依赖的增强效应);
44.所述特异性结合sars-cov-2s蛋白的抗体在10-10000ng/ml的浓度范围内对sars-cov-2感染thp-1细胞、raji细胞、k562细胞都不产生ade效应。。
45.进一步,本发明所述的抗体,其中,所述天然人源抗sars-cov-2s蛋白抗体能够中和sars-cov-2对vero e宿主细胞的感染。
46.进一步,本发明所述的抗体,其中,所述天然人源抗sars-cov-2s蛋白抗体至少在10-10000ng/ml的部分浓度范围内对sars-cov-2感染raji细胞具有ade效应。
47.进一步,本发明所述的抗体,其中,所述人源抗sars-cov-2s蛋白抗体至少在50-3000ng/ml的部分浓度范围内对sars-cov-2感染raji细胞具有 ade效应。
48.进一步,本发明所述的抗体,其中,所述天然人源抗sars-cov-2s蛋白抗体具有:
49.选自seq id no:1、7、13、19、25、31、37的hcdr1;
50.选自seq id no:2、8、14、20、26、32、38的hcdr2;
51.选自seq id no:3、9、15、21、27、33、39的hcdr3;
52.选自seq id no:4、10、16、22、28、34、40的lcdr1;
53.选自seq id no:5、11、17、23、29、35、41的lcdr2;
54.选自seq id no:6、12、18、24、30、36、42的lcdr3。
55.进一步,本发明所述的抗体,其中,所述天然人源抗sars-cov-2s蛋白抗体具有
56.seq id no:1所示hcdr1、seq id no:2所示hcdr2、seq id no:3所示hcdr3,seq id no:4所示lcdr1、seq id no:5所示lcdr2、seq id no:6 所示lcdr3;
57.seq id no:7所示hcdr1、seq id no:8所示hcdr2、seq id no:9所示hcdr3,seq id no:10所示lcdr1、seq id no:11所示lcdr2、seq id
ꢀꢀ
no:12所示lcdr3;
58.seq id no:13所示hcdr1、seq id no:14所示hcdr2、seq id no:15 所示hcdr3,seq id no:16所示lcdr1、seq id no:17所示lcdr2、seq id
ꢀꢀ
no:18所示lcdr3;
59.seq id no:19所示hcdr1、seq id no:20所示hcdr2、seq id no:21 所示hcdr3,seq id no:22所示lcdr1、seq id no:23所示lcdr2、seq id
ꢀꢀ
no:24所示lcdr3;
60.seq id no:25所示hcdr1、seq id no:26所示hcdr2、seq id no:27 所示hcdr3,seq id no:28所示lcdr1、seq id no:29所示lcdr2、seq id
ꢀꢀ
no:30所示lcdr3;
61.seq id no:31所示hcdr1、seq id no:32所示hcdr2、seq id no:33 所示hcdr3,seq id no:34所示lcdr1、seq id no:35所示lcdr2、seq id
ꢀꢀ
no:36所示lcdr3;或
62.seq id no:37所示hcdr1、seq id no:38所示hcdr2、seq id no:39 所示hcdr3,seq id no:40所示lcdr1、seq id no:41所示lcdr2、seq idno:42所示lcdr3。
63.进一步,本发明所述的抗体,其中所述fc段的点突变改变了抗sars-cov-2 s蛋白抗体与其受体的结合。
64.进一步,本发明所述的抗体,其中,所述fc段的点突变降低或消除了抗 sars-cov-2s蛋白抗体与fcγrs的结合。
65.进一步,本发明所述的抗体,其中,所述抗体在10-10000ng/ml的浓度范围内对sars-cov-2感染thp-1细胞、raji细胞、k562细胞都不产生ade效应。
66.进一步,本发明所述的抗体,其中,fc段点突变包括在由第234位、第235 位组成的组中任意一个或两个位点进行氨基酸的替换、缺失或插入突变。
67.进一步,本发明所述的抗体,其中,fc段点突变包括在由第234位、第235 位组成的组中任意一个或两个位点进行单个氨基酸的替换、缺失或插入突变。
68.进一步,本发明所述的抗体,其中,所述fc段点突变包括l234a、l235a 突变。
69.在另一个具体的实施例中,本发明提供抗体在制备治疗sars-cov-2感染药物中的应用,其中所述抗体如本发明第一方面所述。
70.在另一个具体的实施例中,本发明提供抗体在制备治疗sars-cov-2感染导致的疾病药物中的应用,其中所述抗体如本发明第一方面所述,所述疾病包括covid-19。
71.在另一个具体的实施例中,本发明提供一种多核苷酸,其编码本发明第一方面所述的抗体。
72.在另一个具体的实施例中,本发明提供一种载体,其包含本发明第四方面所述的多核苷酸。
73.在另一个具体的实施例中,本发明提供一种宿主细胞,其包含本发明第四方面所述的多核苷酸或本发明第四方面所述的载体。
74.在另一个具体的实施例中,本发明提供一种药物组合物,其包含一种或多种选自由本发明第一方面所述抗体组成的组中的抗体,以及任选的药学上可接受的载体。
75.在另一个具体的实施例中,本发明提供一种消除抗体在sars-cov-2感染宿主细胞
时ade效应的方法,其中,所述的抗体为抗sars-cov-2s蛋白的中和性抗体,通过对抗体fc段进行点突变改变了抗sars-cov-2s蛋白抗体与其受体的结合。
76.进一步,本发明所述消除抗体在sars-cov-2感染宿主细胞时ade效应的方法,其中,所述fc段的点突变降低或消除了抗sars-cov-2s蛋白抗体与 fcγrrs的结合。
77.进一步,本发明所述消除抗体在sars-cov-2感染宿主细胞时ade效应的方法,所述fc段点突变包括在由第234位、第235位组成的组中任意一个或两个位点进行氨基酸的替换、缺失或插入突变。
78.为更好理解本发明,首先定义一些术语。其他定义则贯穿具体实施方式部分而列出。
79.术语“冠状病毒”是指套式病毒目(nidovirales)、冠状病毒科(coronaviridae)、冠状病毒属(coronavirus)的成员。本发明所述冠状病毒主要涉及感染人的冠状病毒,包括hcov-229e、hcov-oc43、hcov-nl63、 hcov-hku1、sars-cov、mers-cov、sars-cov-2(2019-ncov),本发明所述冠状病毒特别涉及sars-cov、mers-cov、sars-cov-2(2019-ncov)。
80.术语“特异性”是指在蛋白和/或其他生物异质群体中确定是否存在所述蛋白,例如本发明所述单抗与sars-cov-2rbd蛋白的结合反应。因此,在所指定的条件下,特定的配体/抗原与特定的受体/抗体结合,并且并不以显著的量与样本中存在的其它蛋白结合。
81.本文中的术语“抗体”意在包括全长抗体及其任何抗原结合片段(简称抗体片段)或单链。全长抗体是包含至少两条重(h)链和两条轻(l)链的糖蛋白,重链和轻链由二硫键连接。各重链由重链可变区(简称vh)和重链恒定区构成。重链恒定区由三个结构域构成,即ch1、ch2和ch3。各轻链由轻链可变区(简称vl) 和轻链恒定区构成。轻链恒定区由一个结构域cl构成。vh和vl区还可以划分为称作互补决定区(cdr)的高变区,其由较为保守的框架区(fr)区分隔开。各 vh和vl由三个cdr以及四个fr构成,从氨基端到羧基端以fr1、cdr1、 fr2、cdr2、fr3、cdr3、fr4的顺序排布。重链和轻链的可变区包含与抗原相互作用的结合域。抗体的恒定区可以介导免疫球蛋白与宿主组织或因子的结合,包括多种免疫系统细胞(例如,效应细胞)和传统补体系统的第一组分(c1q)。
82.术语“单克隆抗体”或“单抗”或“单克隆抗体组成”是指单一分子组成的抗体分子制品。单克隆抗体组成呈现出对于特定表位的单一结合特异性和亲和力。
83.本文中的术语,抗体的“抗原结合片段”(或简称为抗体片段),是指抗体的保持有特异结合抗原能力的一个或多个片段。已证实,抗体的抗原结合功能可以通过全长抗体的片段来实施。包含在抗体的“抗原结合部分”中的结合片段的例子包括(i)fab片段,由vl、vh、cl和ch1构成的单价片段;(ii)f(ab
′
)2片段,包含铰链区二硫桥连接的两个fab片段的二价片段;(iii)由vh和ch1构成的fd片段;(iv)由抗体单臂vl和vh构成的fv片段;(v)由vh构成的dab片段 (ward et al.,(1989)nature 341:544-546);(vi)分离的互补决定区(cdr);以及(vii) 纳米抗体,一种包含单可变结构域和两个恒定结构域的重链可变区。此外,尽管fv片段的两个结构域vl和vh由不同的基因编码,它们可以通过重组法经由使两者成为单蛋白链的合成接头而连接,其中vl和vh区配对形成单价分子(称为单链fc(scfv);参见例如bird et al.,(1988)science 242:423-426;andhuston et al.,(1988)proc.natl.acad.sci.usa 85:5879-5883)。这些单链抗体也意在包括在术语涵义中。这些抗体片段可以通过本领域技术人员已知的常用技术而得到,且片段可以通过与完整抗体相
同的方式进行功能筛选。
84.本发明的抗原结合片段包括能够特异性结合冠状病毒rbd的那些。抗体结合片段的实例包括例如但不限于fab、fab'、f(ab')2、fv片段、单链fv(scfv) 片段和单结构域片段。
85.fab片段含有轻链的恒定结构域和重链的第一恒定结构域(ch1)。fab'片段与fab片段的不同之处在于在重链ch1结构域的羧基末端处的少数残基的添加,包括来自抗体铰链区的一个或多个半胱氨酸。通过切割在f(ab')2胃蛋白酶消化产物的铰链半胱氨酸处的二硫键产生fab'片段。抗体片段的另外化学偶联是本领域普通技术人员已知的。fab和f(ab')2片段缺乏完整抗体的片段可结晶(fc)区,从动物的循环中更快速地清除,并且可能具有比完整抗体更少的非特异性组织结合(参见例如,wahl等人,1983,j.nucl.med.24:316)。
86.如本领域通常理解的,“fc”区是不包含抗原特异性结合区的抗体的片段可结晶恒定区。在igg、iga和igd抗体同种型中,fc区由两个相同的蛋白质片段组成,衍生自抗体的两条重链的第二和第三恒定结构域(分别为ch2和ch3 结构域)。igm和ige fc区在每条多肽链中含有三个重链恒定结构域(ch2、 ch3和ch4结构域)。
[0087]“fv”片段是含有完整靶识别和结合位点的抗体的最小片段。该区域由以紧密的非共价结合的一个重链和一个轻链可变结构域的二聚体(vh-vl二聚体) 组成。在该构型中,每个可变结构域的三个cdr相互作用,以限定在vh-vl二聚体的表面上的靶结合位点。通常,六个cdr对抗体赋予靶结合特异性。然而,在一些情况下,甚至单个可变结构域(或仅包含对于靶特异性的三个cdr的fv 的一半)可以具有识别且结合靶的能力,尽管其亲和力低于整个结合位点。
[0088]“单链fv”或“scfv”抗体结合片段包含抗体的vh和vl结构域,其中这些结构域存在于单条多肽链中。一般地,fv多肽进一步包含在vh和vl结构域之间的多肽接头,其致使scfv能够形成有利于靶结合的结构。
[0089]“单结构域片段”由对冠状病毒rbd显示出足够亲和力的单个vh或vl结构域组成。在一个具体实施方案中,单结构域片段是骆驼化的(参见例如, riechmann,1999,journal ofimmunological methods 231:25
–
38)。
[0090]
本发明的抗冠状病毒rbd的抗体包括衍生化抗体。例如,衍生化抗体通常通过糖基化、乙酰化、聚乙二醇化、磷酸化、酰胺化、通过已知保护/封闭基团的衍生化、蛋白酶解切割、与细胞配体或其它蛋白质的连接来修饰。可以通过已知技术进行众多化学修饰中的任一种,所述技术包括但不限于特定的化学切割、乙酰化、甲酰化、衣霉素的代谢合成等。另外,衍生物可以含有一种或多种非天然氨基酸,例如,使用ambrx技术(参见例如,wolfson,2006,chem.biol. 13(10):1011-2)。
[0091]“人源抗体”包括具有人免疫球蛋白的氨基酸序列的抗体,并且包括从人免疫球蛋白文库或动物中分离的抗体,所述动物对于一种或多种人免疫球蛋白是转基因的,并且不表达内源免疫球蛋白。人抗体可以通过本领域已知的各种方法制备,所述方法包括使用衍生自人免疫球蛋白序列的抗体文库的噬菌体展示方法。参见美国专利号4,444,887和4,716,111;以及pct公开wo 98/46645;wo 98/50433;wo 98/24893;wo 98/16654;wo 96/34096;wo96/33735;和 wo 91/10741。还可以使用不能表达功能性内源免疫球蛋白,但可以
表达人免疫球蛋白基因的转基因小鼠来产生人抗体。参见例如,pct公开wo 98/24893; wo 92/01047;wo 96/34096;wo 96/33735;美国专利号5,413,923;5,625,126; 5,633,425;5,569,825;5,661,016;5,545,806;5,814,318;5,885,793;5,916,771;和5,939,598。另外,使用与上述类似的技术,公司例如lakepharma,inc. (belmont,ca)或creative biolabs(shirley,ny)可以从事于提供针对所选抗原的人抗体。可以使用被称为“引导选择”的技术生成识别所选表位的全人抗体。在该方法中,选择的非人单克隆抗体,例如小鼠抗体,用于引导识别相同表位的完全人抗体的选择(参见,jespers等人,1988,biotechnology12:899-903)。
[0092]
术语“识别抗原的抗体”以及“对抗原特异的抗体”在本文中与术语“特异结合抗原的抗体”交替使用。
[0093]
术语“高亲和性”对于igg抗体而言,是指对于抗原的kd为1.0
×
10-6
m以下,优选5.0
×
10-8
m以下,更优选1.0
×
10-8
m以下、5.0
×
10-9
m以下,更优选 1.0
×
10-9
m以下。对于其他抗体亚型,“高亲和性”结合可能会变化。例如,igm 亚型的“高亲和性”结合是指kd为10-6
m以下,优选10-7
m以下,更优选10-8
m 以下。
[0094]
术语“kassoc”或“ka”是指特定抗体-抗原相互作用的结合速率,而术语“kdis”或“kd”是指特定抗体-抗原相互作用的离解速率。术语“kd”是指解离常数,由kd与ka比(kd/ka)得到,并以摩尔浓度(m)表示。抗体的kd值可以通过领域内已知的方法确定。优选的确定抗体kd的方式是使用表面等离子共振仪(spr)测得的,优选使用生物传感系统例如biacoretm系统测得。
[0095]
术语“ec50”,又叫半最大效应浓度,是指引起50%最大效应的抗体浓度。
[0096]
术语“ic50”,是指被测量的拮抗剂的半抑制浓度。它能指示某一药物或者物质(抑制剂)在抑制某些生物程序(或者是包含在此程序中的某些物质,比如酶,细胞受体或是微生物)的半量。
[0097]
术语“表位”是指抗原上b和/或t细胞作出反应的位点。b细胞表位可均由连续氨基酸或因蛋白质的三级折叠而并列的不连续氨基酸形成。由连续氨基酸形成的表位在暴露于变性溶剂时典型地被保留,而通过三级折叠而形成的表位在用变性溶剂处理时典型地丢失。表位典型地包括呈独特空间构象的至少3 个,且更通常是至少5或8-10个氨基酸。
[0098]
术语“组合物”,本发明提供包含如本文所述抗体或其抗原结合片段的组合物。根据本发明的组合物可以与适合的载剂、赋形剂和并入调配物中以提供改良的转移、递送、耐受性和类似性质的其它药剂一起给予。在所有医药化学工作者已知的配方书中可以发现许多适当的调配物:remington
′
spharmaceutical sciences,mackpublishing company,easton,pa。这些调配物包括例如粉剂、糊剂、膏剂、果冻、蜡、油、脂质、含脂质(阳离子或阴离子)的囊泡(如lipofectintm)、dna结合物、无水吸收糊剂、水包油和油包水乳液、乳液碳蜡(各种分子量的聚乙二醇)、半固体凝胶和含有碳蜡的半固体混合物。还参见powell等人“compendium of excipients for parenteral formulations
”ꢀ
pda(1998)《医药科学技术杂志(j pharm sci technol)》52:238-311。
[0099]
本发明所述组合物优选为可注射制剂,可注射制剂可以包括用于静脉内、皮下、皮内和肌肉内注射、点滴输注等的剂型。这些可注射制剂可以通过公众已知的方法来制备。举例来说,可以例如通过将上文所描述的抗体或其盐溶解、悬浮或乳化在常规用于注射的无
菌水性介质或油性介质中来制备注射制剂。作为用于注射的水性介质,存在例如生理盐水、含有葡萄糖和其它助剂的等张溶液等,其可以与如以下的适当的增溶剂组合使用:醇(例如乙醇)、多元醇(例如丙二醇、聚乙二醇)、非离子表面活性剂[例如聚山梨醇酯80、hco-50(氢化蓖麻油的聚氧乙烯(50mol)加合物)]等。作为油性介质,存在已采用的例如芝麻油、大豆油等,其可与如苯甲酸苯甲酯、苯甲醇等增溶剂组合使用。因此制备的注射优选填充于适当的安瓿中。
[0100]
术语“氨基酸突变”在本文中意思指多肽序列中的氨基酸取代、插入和/或缺失,或针对以化学方式连接至蛋白质的部分的改变。举例来说,突变可以是附接至蛋白质的碳水化合物或peg结构的改变。“氨基酸突变”在本文中意思指多肽序列中的氨基酸取代、插入和/或缺失。为清楚起见,除非另外说明,否则氨基酸突变通常是针对dna编码的氨基酸,例如在dna和rna中具有密码子的20种氨基酸。
[0101]“氨基酸取代”或“取代”在本文中意思指亲本多肽序列中特定位置处的氨基酸被不同氨基酸置换。确切地说,在一些实施方案中,取代是针对特定位置处非天然存在的氨基酸,这些氨基酸不是生物体内天然存在的或是任何生物体中的。举例来说,取代e272y是指272位的谷氨酸被酪氨酸置换的变体多肽,在此情形中是fc变体。为清楚起见,蛋白质被工程改造成改变核酸编码序列但不改变起始氨基酸(例如cgg(编码精氨酸)变为cga(仍编码精氨酸),用以增加宿主生物体表达水平)不是“氨基酸取代”;也就是说,尽管产生了编码同一蛋白质的新基因,但如果该蛋白质在其起始的特定位置处具有相同氨基酸,就不是氨基酸取代。
[0102]
如本文所使用,“氨基酸插入”或“插入”意思指在亲本多肽序列中的特定位置处添加氨基酸序列。举例来说,-233e或233e指示在233位后并且在234 位前插入谷氨酸。另外,-233ade或a233ade指示在233位后并且在234位前插入alaaspglu。
[0103]
如本文所使用,“氨基酸缺失”或“缺失”意思指去除亲本多肽序列中特定位置处的氨基酸序列。举例来说,g236-或g236#或g236del指示在236位处的甘氨酸缺失。另外,eda233-或eda233#表示自233位开始缺失序列 gluaspala。
[0104]
如本文所使用,“残基”意思指蛋白质中的位置及其相关氨基酸的身份。举例来说,天冬酰胺297(又称为asn297或n297)是在人类抗体igg1中297 位的残基。
[0105]
与现有技术相比,本发明的技术方案具有以下优点:
[0106]
第一,本发明采用单个淋巴细胞克隆技术,从sars-cov-2感染康复期患者的大量单个b细胞中扩增获得了多个特异性抗体,经过重组表达、特异性结合能力筛选、表位竞争分析、sars-cov-2s蛋白-aceii结合阻断活性分析获得了能以高亲和力、特异性结合sars-cov-2s蛋白rbd,且能够阻断 sars-cov-2结合宿主细胞受体aceii的7株单克隆抗体,提供了其轻重链 cdrs区的氨基酸序列。筛选过程中发现,抗体阻断sars-cov-2结合宿主细胞受体aceii的活性与其特异性结合sars-cov-2s蛋白rbd的亲和力并无正相关性。本发明的单抗在假病毒中和试验、sars-cov-2病毒颗粒中和试验中都取得了良好的中和封闭的技术效果。
[0107]
第二,本发明对筛选获得的具有sars-cov-2s蛋白-aceii结合阻断活性的7株单克隆抗体进行ade分析,其在sars-cov-2感染thp-1细胞和k562 细胞的过程均不产生ade效应;提示本发明筛选的7株单抗在临床应用中具有较低的ade效应。另外,本发明的7株单克
隆抗体不仅是康复患者血清中天然存在的全人源抗体,而且针对sars-cov-2s蛋白rbd多个不同的天然表位,因此特别适合“鸡尾酒抗体疗法”。
[0108]
第三,本发明研究发现特定浓度范围的抗体corn-01、corn-05在 sars-cov-2病毒感染raji宿主细胞时会产生ade效应。结合现有技术的研究报道,探索了抗sars-cov-2的中和性抗体产生ade效应的机理,根据raji 宿主细胞与其它不产生ade效应的宿主细胞上fc受体种类的不同,分析抗 sars-cov-2中和性抗体产生ade效应的机理可能是由于抗体与宿主细胞膜上 fcγri受体的结合导致的;从而使得本领域技术人员能够通过阻断抗 sars-cov-2中和性抗体与宿主细胞fcγriib受体的结合而降低或消除ade 效应,即保留了抗体的病毒阻断活性,又消除了可能的ade效应,提高了人体临床应用的安全性。
附图说明
[0109]
通过阅读下文优选实施方式的详细描述,各种其他的优点和益处对于本领域普通技术人员将变得清楚明了。附图仅用于示出优选实施方式的目的,而并不认为是对本发明的限制。而且在整个附图中,用相同的参考符号表示相同的部件。在附图中:
[0110]
图1:sars-cov-2s1蛋白与his的融合表达。
[0111]
图2:sars-cov-2s1蛋白与mfc的融合表达。
[0112]
图3:sars-cov-2s1蛋白rbd与his的融合表达。
[0113]
图4:sars-cov-2s1蛋白rbd与mfc的融合表达。
[0114]
图5:人ace2与人fc的融合表达。
[0115]
图6:重组抗体corn-02、corn-10、corn-01阻断s1rbd-mfc与ace2 结合的能力。cr3022是us2010172917a1公开的一株抗sars-cov rbd的单抗;corn-10是本发明实施例4中制备的与s1rbd-mfc有特异性结合的单抗。
[0116]
图7:重组抗体corn-02、corn-10、corn-01阻断s1-mfc与ace2结合的能力。
[0117]
图8:重组抗体对s1rbd-mfc与aceii-hfc结合的阻断作用曲线图。
[0118]
图9:corn-01、corn-07对s1rbd-mfc与aceii-hfc结合阻断作用曲线图(竞争elisa法)。
[0119]
图10:corn-01、corn-07对s1rbd-his与aceii-his结合的阻断作用曲线图(假病毒感染荧光检测法)。
[0120]
图11:corn-01在sars-cov-2病毒颗粒感染不同宿主细胞过程中的ade 效应
[0121]
图12:corn-01、corn-04、corn-05、corn-06对sars-cov-2病毒颗粒感染raji细胞的ade效应
[0122]
图13:corn-01、corn-05、corn-07对sars-cov-2病毒颗粒感染raji细胞的ade效应
[0123]
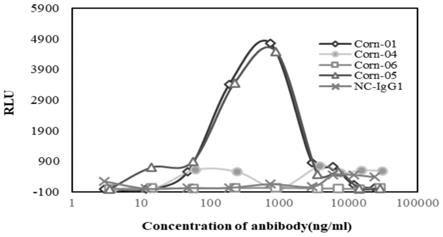
图14:corn-01、corn-01-lala对sars-cov-2病毒颗粒感染raji细胞的 ade效应
[0124]
图15:corn-05、corn-05-lala对sars-cov-2病毒颗粒感染raji细胞的 ade效应
[0125]
图16:corn-01、corn-05对sars-cov-2病毒颗粒的中和活性
[0126]
图17:corn-01、corn-01-lala对sars-cov-2病毒颗粒的中和活性分析
[0127]
图18:corn-05、corn-05-lala对sars-cov-2病毒颗粒的中和活性分析。
具体实施方式
[0128]
下面将参照附图更详细地描述本公开的示例性实施方式。虽然附图中显示了本公开的示例性实施方式,然而应当理解,可以以各种形式实现本公开而不应被这里阐述的实施方式所限制。相反,提供这些实施方式是为了能够更透彻地理解本公开,并且能够将本公开的范围完整的传达给本领域的技术人员。
[0129]
实施例1:sars-cov-2抗原及宿主受体的重组表达
[0130]
将全合成的基因s1rbd(accession:qhd43416.1,319-541aa),通过酶切方法分别克隆入c端带有his标签或mfc标签的真核瞬时表达载体中,将获得的表达质粒,转入大肠杆菌扩增,分离获得s1rbd-his和s1rbd-mfc表达质粒,并根据转染试剂293fectin(cat:12347019,gibco)的操作说明,将质粒转入hek293细胞中重组表达。细胞转染后5-6天,取培养上清,s1rbd-mfc利用proa亲和层析柱对表达上清进行纯化,获得s1rbd-mfc蛋白(氨基酸序列如 seq id no:46所示)。s1rbd-his(氨基酸序列如seq id no:45所示)利用histraphp柱亲和层析柱对表达上清进行纯化,获得s1rbd-his蛋白。并将获得的重组蛋白通过sds-page检测纯度(图3-4)。
[0131]
通过pcr的方法从购买的全长sars-cov-2表达载体(cat:vg40589-ut,北京义翘神州)中克隆出s1基因(accession:qhd43416.1,1-685aa),并通过酶切方法分别克隆入c端带有mfc标签或his标签的真核瞬时表达载体中,将获得的表达质粒,转入大肠杆菌扩增,分离获得s1-mfc、s1-his表达质粒,并根据转染试剂293fectin(cat:12347019,gibco)的操作说明,将质粒转入 hek293细胞中重组表达。细胞转染后5-6天,取培养上清,纯化,获得s1
–
mfc (氨基酸序列如seq id no:48所示)、s1-his蛋白(氨基酸序列如seq id no:47 所示)。并将获得的重组蛋白通过sds-page检测纯度(图1-2)。
[0132]
通过pcr的方法从购买的全长人aceii表达载体(cat:hg10108-acr,北京义翘神州)中克隆出aceii胞外区基因aceii(1-615)(accession:np_068576.1,1-615aa),并通过酶切方法分别克隆入c端带有hfc标签的真核瞬时表达载体中,将获得的表达质粒,转入大肠杆菌扩增,分离获得,aceii (1-615)-hfc表达质粒,并根据转染试剂293fectin(cat:12347019,gibco)的操作说明,将质粒转入hek293细胞中重组表达。细胞转染后5-6天,取培养上清,利用proa亲和层析柱对表达上清进行纯化,获得aceii(1-615)-hfc蛋白 (氨基酸序列如seq id no:49所示),并将获得的重组蛋白通过sds-page 检测纯度(图5)。
[0133]
实施例2:sars-cov-2s1蛋白rbd特异性记忆b细胞的分离
[0134]
对新型冠状病毒感染康复患者的特异性记忆b细胞进行检测和分选,采用新冠抗体检测试剂盒对患者血清中igg及igm抗体进行了检测,选择新冠抗体 igg阳性的血清样本。利用rosettesep试剂盒(cat:15064,stemcell)对b 细胞进行了富集,在此基础上采用fitc标记的s1-rbd-his去捕获新冠rbd 特异性结合的记忆性b细胞并进行流式单细胞分选。
[0135]
实施例3:人源抗sars-cov-2rbd抗体序列的扩增
[0136]
采用rna磁珠(南京诺唯赞)提取单b细胞的rna,反转录成cdna,具体方法如下:
[0137]
1.每孔分装5μl catch buffer b(tcl 1%β-me),分选单个记忆性b 细胞。
[0138]
2.贴膜,2000rpm离心1min。
[0139]
3.每孔加入10μl h2o和33μl beads,吹吸混匀,室温作用10min。
[0140]
4.置磁力架,室温5min,弃上清。
[0141]
5.用200μl无核酸酶水新鲜配置的80%乙醇漂洗磁珠,室温30s,弃上清。
[0142]
6.重复漂洗一次,弃上清,风干3min。
[0143]
7.移下磁力架,每孔加12μl mix 1,吹吸5次,室温作用5min。
[0144]
8.置磁力架,室温2min,转移10μl至新板,300g离心30s,运行程序 1。
[0145]
9.每孔加10μl mix 2,混匀、离心,运行程序2。
[0146]
10.合成好的cdna尽快进行pcr。
[0147]
mix 1:310μl h2o 50μl dntp 20μl random 6 20μl oligo_dt
[0148]
mix 2:170μl h2o 160μl buffer 40μl dtt 20μl rnase i 10μl rtase iv(cat:en0601 and 18090010,thermofisher)
[0149]
程序1:65℃5min
→
4℃∞
[0150]
程序2:23℃10min
→
50℃30min
→
80℃10min
→
4℃∞
[0151]
采用两步pcr扩增抗体重链以及轻链(kappa)可变区基因。引物序列源自于human monoclonal antibodies书中第114至117页。具体方法如下:
[0152]
第一轮pcr(ig-vh1、ig-vk1),反应体系(20μl):
[0153][0154]
运行程序:
[0155]
94℃5min
→
(94℃30s
→
51℃30s
→
72℃55s)
×
15cycles
[0156]
→
(94℃30s
→
56℃30s
→
72℃55s)
×
30 cycles
[0157]
→
72℃8min
[0158]
→
4℃∞
[0159]
第二轮pcr(ig-vh2、ig-vk2),反应体系(20μl):
[0160][0161]
运行程序:
[0162]
94℃5min
→
(94℃30s
→
57℃30s
→
72℃45s)
×
50 cycles
[0163]
→
72℃10min
[0164]
→
4℃∞
[0165]
琼脂糖凝胶电泳分离纯化pcr产物并进行抗体轻、重链可变区测序。
[0166]
实施例4:人源抗sars-cov-2rbd抗体的表达及特异性结合初筛鉴定
[0167]
对测序后的122对序列进行分析、并进一步合成其中的49对抗体轻重链可变区基
因,将其克隆入全抗体瞬时表达载体中,进行重组表达和特异性鉴定。将全合成的抗体重链可变区,通过酶切克隆入真核瞬时表达载体pkn041的人 igg1的重链恒定区编码基因的上游,全合成的抗体轻链可变区通过酶切克隆入真核瞬时表达载体pkn019的人轻链cκ的编码基因的上游,构建轻、重链表达载体,获得轻链和重链表达质粒,转入大肠杆菌扩增,分离获得抗体轻链和重链质粒,并根据转染试剂293fectin(cat:12347019,gibco)的操作说明,将抗体的轻、重链质粒转入hek293细胞中重组表达。
[0168]
细胞转染后24小时,取上清利用fortebio公司的octet qke system仪器,采用抗人抗体fc段的捕获抗体(ahc)生物探针捕获抗体fc段的方法测定抗体与s1rbd的结合。测定时将抗体上清及抗体,流经ahc探针(cat:18-0015, pall)表面,时间为240s。s1rbd-mfc(kn表达,lot:20200217a)作为流动相, s1rbd-mfc重组蛋白浓度为100nm。结合时间为300s,解离时间为300s。实验完毕,用软件进行1:1langmuir结合模式拟合,计算抗原抗体结合的动力学常数。共检测了39个抗体,其中与s1rbd-mfc有特异性结合的抗体为20 个(表1)。
[0169]
表1候选抗体与s1rbd-mfc的特异性结合动力学参数。
[0170]
[0171]
[0172][0173]
根据表1可知,39株成功表达的抗体分子中经过初筛对s1rbd-mfc具有特异性结合能力的20株抗体分别为克隆编号1、2、29、5、6、31、15、22、 16、17、18、36、35、33、27、28、38、32、24、11。从初筛亲和力高的抗体克隆选择1、17、22、5、24、32、38、11进一步研究。所述抗体克隆5、17、 11、24、32、38、1在下文中也被称为corn-01、corn-02、corn-03、corn-04、corn-05、 corn-06、corn-07。
[0174]
所述抗体corn-01的hcdr1-hcdr3分别为seq id no:1-3,lcdr1-lcdr3 分别为seq id no:4-6。
[0175]
所述抗体corn-02的hcdr1-hcdr3分别为seq id no:7-9,lcdr1-lcdr3 分别为seq id no:10-12。
[0176]
所述抗体corn-03的hcdr1-hcdr3分别为seq id no:13-15, lcdr1-lcdr3分别为seq id no:16-18。
[0177]
所述抗体corn-04的hcdr1-hcdr3分别为seq id no:19-21, lcdr1-lcdr3分别为seq id no:22-24。
[0178]
所述抗体corn-05的hcdr1-hcdr3分别为seq id no:25-27, lcdr1-lcdr3分别为seq id no:28-30。
[0179]
所述抗体corn-06的hcdr1-hcdr3分别为seq id no:31-33, lcdr1-lcdr3分别为seq id no:34-36。
[0180]
所述抗体corn-07的hcdr1-hcdr3分别为seq id no:37-39, lcdr1-lcdr3分别为seq id no:40-42。
[0181]
实施例5:抗体的elisa阻断活性
[0182]
将上清fortebio检测亲和力高的克隆进行进一步的elisa阻断活性检测。具体方法如下:
[0183]
1.包板:包被human ace2-hfc(1-615)(kn表达,lot:20200213c),浓度为 0.75ug/ml;每孔100ul;4℃o/n;
[0184]
2.封闭:5%bsa in pbs,37℃,120min,pbst洗板4次;
[0185]
3.加一抗:120ul 30ng/ml s1-rbd-mfc(kn表达,lot:20200217a),加入 10ug/ml浓度的corn-09、cr3022、corn-08、corn-02、corn-10、corn-01各 120ul和5 17(1:1混合,5ug/ml),略震荡混匀放置50min,每孔取2个100ul 平行加入human ace2-hfc(1-615)包被
孔;
[0186]
4.加二抗:hrp-anti-mouse igg(cat:115-035-071,jackson immuno research) (1:5000)37℃,45min,pbst洗板4次;
[0187]
5.显色:tmb(cat:me142,北京泰天河生物)显色,37℃,10min;
[0188]
6.终止:2m hcl终止反应;
[0189]
7.读数:读取并记录波长450nm下孔板的吸光度值。
[0190]
结果如图6~7所示,对于s1rbd与aceii,s1与aceii的结合,重组抗体corn-01和corn-02有明显的阻断活性,且具有协同效应。重组抗体corn-10 不能阻断s1rbd与ace2的结合,但能够抑制s1与ace2的结合。上述结果表明抗体corn-01和corn-02二者与ace2,在sars-cov-2s1蛋白上具有相同的结合位点,因而能够直接阻断s1rbd与ace2的结合。
[0191]
进一步对于抗体corn-01、corn-02、corn-03、corn-04、corn-05、corn-06、 corn-07,采用梯度稀释法进行elisa。简言之,根据上述方法包被、封闭之后,在添加一抗的步骤中,将上述待测抗体(起始浓度为40μg/ml,1.5倍连续稀释,12个梯度)和与s1-rbd-mfc 70ng/ml,各取100ul等体积混匀,37℃放置 50min后,每个样本取两个100μl平行加入到ace2-hfc包被孔;然后加二抗、显色、终止、读数。结果如图8~9所示。
[0192]
6株抗体均能明显抑制aceii-hfc与s1rbd-mfc的结合,其半数抑制浓度 ic50如表2所示:
[0193]
表2:抗体对aceii-hfc与s1rbd-mfc的结合的半数抑制浓度
[0194][0195][0196]
实施例6.抗体对sars-cov-2的病毒中和试验方法
[0197]
6.1假病毒感染试验——荧光法
[0198]
对于corn-01和corn-07中和sars-cov-2的活性进行预估,采用中检院的新冠假病毒感染huh7细胞中和活性实验观察。将不同浓度的待评估抗体与 750tcid50/孔的假病毒颗粒(转染荧光素酶报告基因)在37℃进行中和反应后,分别接种2
×
104/孔huh7细胞,置于37℃的co2培养箱内培养20-28h。20-28h 后取出细胞板每孔加入100μl荧光素酶检测试剂,避光反应2min,荧光检测仪读数,计算中和抑制率,并根据中和抑制率结果,利用reed-muench法计算ic50。
[0199]
结果如图10所示,corn-07能够剂量依赖地抑制假病毒颗粒进入宿主细胞,经过reed-muench法计算,ic50值为62ng/ml,高于corn-01(ic50,281ng/ml) 的中和活性。
[0200]
6.2 sars-cov-2病毒感染试验——细胞病变中和滴定法
[0201]
1)取生长状态良好的vero-e6细胞消化,调整细胞密度为1
×
105/ml接种于96孔板,100μl/孔(即每孔104个细胞),放置于37℃、5%co2培养箱培养12-16h;
[0202]
2)12-16h后,弃去孔中培养基,首先分别加入终浓度为不同稀释度(最高浓度100μg/ml,3倍稀释8次)的样品50μl,然后每孔加入102ccid50 的sars-cov-2病毒50μl。同时设置细胞对照,病毒对照。
[0203]
3)于加样后72h观察并测定细胞病变情况,实验结果如表3所示。
[0204]
表3抗体中和sars-cov-2病毒感染宿主细胞的活性检测结果-ec50
[0205][0206][0207]
实施例7:corn-01抗体对sars-cov-2感染细胞的ade作用
[0208]
采用中检院的新冠病毒颗粒感染淋巴细胞细胞(thp-1、raji和k562)进行抗体的ade检测。将不同浓度的待评估新冠抗体与750tcid50/孔的假病毒颗粒(转染荧光素酶报告基因)在37℃进行中和反应后,分别接种1x105/孔宿主细胞,置于37℃,5%co2培养箱内培养20-28h。28-28h后取出细胞板每孔加入100ul荧光素酶检测试剂(g7940,promega),避光反应2min,荧光检测仪读数,根据荧光信号强度评价ade的强弱。
[0209]
结果表明(图11),igg1形式的corn-01对raji细胞在一定浓度范围内 (50ng/ml-3000ng/ml)表现出了明显的ade作用;而对于thp-1和k562在 1-1000ng/ml的浓度范围内均未产生ade效应。
[0210]
实施例8:中和性抗体在病毒感染敏感宿主细胞过程中ade效应的检测
[0211]
利用实施例7的方法,采用raji细胞对前期筛选鉴定的5种抗sars-cov-2 s1蛋白rbd区的全人源中和性抗体检测ade效应,结果如图12、图13所示。
[0212]
图12、图13表明,不同的抗sars-cov-2s1蛋白rbd区中和性抗体在病毒感染敏感宿主细胞过程中引起ade效应的能力存在差异。igg1形式的 corn-04、corn-06和corn-07三个抗体在1-1000ng/ml的浓度范围内并未观察到明显的ade效应,而corn-01和corn-05在一定浓度范围 (50ng/ml-3000ng/ml)内产生了ade现象。
[0213]
采用中检院的新冠假病毒颗粒感染raji细胞进行抗体的ade检测。将不同浓度的待评估新冠抗体与750tcid50/孔的假病毒颗粒(转染荧光素酶报告基因)在37℃进行中和反应后,分别接种1x105/孔raji细胞,置于37℃的co2 培养箱内培养20-28h。20-28h后取出细胞板每孔加入100μl荧光素酶检测试剂,避光反应2min,荧光检测仪读数,根据荧光信号强度评价ade的强弱。
[0214]
实施例9:抗体fc点突变改变了对宿主细胞受体亲和力的影响
[0215]
根据raji宿主细胞与其它不产生ade效应的宿主细胞上fc受体种类,确定fc点突变的位置,并检测突变后抗体对宿主细胞上fc受体的亲和力。将 corn-01和corn-05的fc进行lala突变(fc-l234a,l235a),构建 corn-01-lala和corn-05-lala。突变前的野生型重链恒定区的氨基酸序列如 seq id no:43所示,突变后的突变型重链恒定区的氨基酸序列如seq id no:44 所示。
[0216]
应用fortebio蛋白相互作用系统octet qke system测定测定corn-05 (wtigg1)
和corn-05-lala与fcγri(cd64)重组蛋白(10256-h08h,义翘神州)和fcγriia(cd32a)重组蛋白(10374-h08h1,义翘神州)的亲和力。采用抗人抗体fc段的捕获抗体(ahc)生物探针捕获抗体corn-05和 corn-05-lala。15μg/ml抗体流经ahc探针(cat:18-5060,pall)表面,时间为120s。浓度100nm重组抗原作为流动相。结合时间为300s,解离时间为 300s。实验完毕,扣除空白对照响应值,用软件进行1:1langmuir结合模式拟合,计算抗原抗体结合的动力学常数,结果如表4所示。
[0217]
表4.corn-05和corn-05-lala对cd64和cd32a亲和力常数
[0218]
sample idloading sample idresponsekd(m)kon(1/ms)kdis(1/s)cd64-hiscorn-050.11741.45e-081.20e 051.73e-03cd64-hiscorn-05-lala0.03443.66e-057.89e 022.89e-02cd32a-hiscorn-050.23025.60e-081.89e 061.06e-01cd32a-hiscorn-05-lala0.22546.62e-081.94e 061.29e-01
[0219]
表4的结果表明:与wtigg1型的corn-05相比,lala突变后其与各fc γrs的亲和力发生了改变。
[0220]
实施例10:抗体fc的点突变消除了抗体对新冠病毒感染宿主细胞的ade效应
[0221]
为了验证通过点突变降低中和性抗体fc与宿主细胞受体fcγri的亲和力是否能够降低或消除其对新冠病毒感染宿主细胞的ade效应,将突变后的 corn-01-lala、corn-05-lala分别与突变前的corn-01、corn-05进行对比,采用实施例7的方法检测对新冠病毒感染宿主细胞的ade效应。结果如图14、图15所示。
[0222]
图14、图15表明:corn-01-lala、corn-05-lala由于在fc段引入了 l234a、l235a突变,消除了ade现象。结合实施例9的实验结果可以推断抗sars-cov-2s1蛋白rbd区的全人源中和性抗体在sars-cov-2感染宿主过程中引起ade效应可能是通过抗体fc段与宿主细胞上fcrri受体介导的,因此通过在fc段引入l234a、l235a突变后极大降低了抗体与fcrri受体的结合,从而避免了ade的发生。
[0223]
实施例11:抗体fc的点突变不影响抗体对新冠病毒的中和活性
[0224]
采用中检院的新冠病毒颗粒感染huh-7细胞进行抗体的中和活性检测。将不同浓度的待评估新冠抗体与750tcid50/孔的假病毒颗粒(转染荧光素酶报告基因)在37℃进行中和反应后,分别接种2x104/孔huh-7细胞,置于37℃,5%co2培养箱内培养20-28h。28-28h后取出细胞板每孔加入100ul荧光素酶检测试剂 (g7940,promega),避光反应2min,荧光检测仪读数,根据荧光信号强度评价抗体对病毒的中和活性,结果如图16所示。图16的结果表明,wtigg1形式的corn-01、corn-05呈现了良好的剂量依赖的病毒中和活性。
[0225]
将消除了ade效应的corn-01和corn-05的lala突变体(fc-l234a, l235a)的中和活性进行检测,评估wtigg1与lala突变体(l234a,l235a) 的中和活性的差异,结果如图17和图18所示。
[0226]
图17和图18的结果表明与corn-01、corn-05相比,fc段lala突变体 corn-01-lala、corn-05-lala的中和活性并未发生明显改变。由此可知,对抗sars-cov-2中和性抗体的fc段进行上述点突变改造能够保留其病毒中和活性。
[0227]
以上所述,仅为本发明较佳的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到的变化或替换,
都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应以所述权利要求的保护范围为准。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。

