基因ghsinas在防治棉花黄萎病中的应用
技术领域
1.本发明涉及植物病害防治技术领域,具体而言,涉及基因ghsinas在防治棉花黄萎病中的应用。
背景技术:
2.大丽轮枝菌是一种能引起黄萎病的极具破坏性的土传真菌病原体,这种病菌会影响200多种植物的生长发育,其中包括重要的农业经济作物棉花。众所周知,大丽轮枝菌是一种非常难控制的病菌,其具有极强的侵染致病性以及在土壤中的存活能力强,使得大丽轮枝菌能够穿透主根部木质部导管和维管组织,严重堵塞了植物的导管,最终导致植物发育迟缓和枯萎。除此之外,这种病菌的宿主范围很广,即使没有宿主也能在土壤中存活很多年。在棉花生产中,抗病品种的选育是一种有效且实用的控制黄萎病菌威胁的策略。可是,由于现有的抗黄萎病菌的资源有限,利用传统的方法培育抗性品种是具有挑战性的。近些年来,以基因工程为基础的棉花抗性候选基因的研究已成为缓解黄萎病菌侵害的另一种策略,其中已报道的包括ghmyb1、gbsobir1、ghcrr1、gbtsa1和ghwak7a。
3.泛素化蛋白酶体系统(ups)在植物生长发育和生理及环境压力的防御反应中发挥重要的作用。泛素化进程通常通过三种酶的连续作用进行,泛素活化酶(e1),泛素结合酶(e2)和泛素连接酶(e3),通过26s蛋白酶体降解系统。将泛素与候选蛋白底物连接,实现目标蛋白降解。泛素e1和e2是相对保守的,但是e3是非常多样化的,因为e3能招募特定靶蛋白进行泛素化。以亚单元组成为基础,e3泛素连接酶被分成单亚基和多亚基。hect和ring/u
‑
box型e3连接酶作为单亚基行使功能,然而skp
‑
cullin
‑
f
‑
box复合物和细胞分裂后期产生的促进复合体是以多亚基泛素连接酶通过分离亚基来结合底物。由于这些亚单位组成不同,e3连接酶决定了泛素化候选底物的特异性。
4.sina蛋白是一种ring型e3连接酶,包含一个定位在n端的ring结构域,其次是与底物识别和聚合有关的保守sina结构域。第一个被鉴定的sina e3连接酶是在黑腹果蝇中,调节果蝇光感受器分化。之后的一系列研究发现,sina同源物在动物中发挥多种极其重要的作用,如肿瘤抑制、细胞凋亡、白血病发生、缺氧反应和自身免疫。同样地,sina e3连接酶也参与到不同的植物生长发育阶段和一些环境应激反应。在拟南芥中,通过sinat2和atrap2.2互作来介导叶胡萝卜素的合成。sinat1和sinat2蛋白与自噬相关蛋白atg6互作来调控自噬通路。在水稻中,沉默干旱诱导sina基因1(osdis1)能显著提高水稻的抗旱性。在番茄中,slsina4基因过表达可以激活防御反应有关的细胞死亡信号。香蕉中,masina1通过介导maice1稳定性负向调控冷胁迫的应激反应。转基因苹果中,超表达mdsina2基因增强了对aba的敏感性。在热胁迫条件下,tasina e3连接酶提高小麦生物量和产量。然而,与抗黄萎病相关的sina e3连接酶的功能和应用仍是未知的。
5.有鉴于此,特提出本发明。
技术实现要素:
6.本发明为了研究e3泛素连接酶基因在抗陆地棉黄萎病菌中的功能作用,鉴定了三个由黄萎病侵染所诱导的sina e3连接酶基因ghsina7、ghsina8和ghsina9。在这三个基因中,每个超表达转基因株系都提高了对黄萎病的抗性,然而每个基因的沉默都抑制了对病原体感染的防御能力。这些发现表明sina e3连接酶参与了植物对病原真菌的防御,同时通过翻译后修饰为培育抗黄萎病棉花品种提供了有效的基因资源。
7.基于上述内容,本发明提供了以下技术方案:
8.基因ghsinas在防治棉花黄萎病中的应用,所述基因ghsinas包括基因ghsina7、基因ghsina8和基因ghsina9中的任一种或多种;
9.基因ghsina7的核酸序列如seq id no:1和/或seq id no:2所示;
10.基因ghsina8的核酸序列如seq id no:3和/或seq id no:4所示;
11.基因ghsina9的核酸序列如seq id no:5和/或seq id no:6所示。
12.sina e3泛素连接酶是普遍存在的调节因子,可在翻译后水平调控植物生长和应激反应。本发明首先系统性地鉴定了24个陆地棉中含有高度保守的ring指域和sina结构域的sina成员。用qrt
‑
pcr方法分析了棉花在接种黄萎病菌24h后,与未感染的对照植株相比,ghsina7、ghsina8和ghsina9基因的转录水平显著上调。
13.sina基因编码c3hc4型ring e3连接酶,它们通常作为二聚体活跃,以维持自身在体内的稳定性,并发挥不同的生物学功能。事实上,ring结构域在空间上招募e2,是c3hc4型ring e3连接酶形成二聚物所必需的。c3hc4型ring e3连接酶,如slsina、mtsina和mdsina,与自身或与其它sina蛋白形成同源或异源二聚体。二聚体的形成需要依赖体外ring e3连接酶的活性。酵母双杂实验,我们发现ghsina7、ghsina8和ghsina9蛋白会自身互作,或分别与其它两种ghsinas相互作用形成同源或异源二聚体。因此,ghsina e3泛素连接酶通过形成同源或异二聚体复合物来完成泛素化功能。
14.泛素化是植物中控制一系列生理进程的翻译后调控机制。e3连接酶能特异性识别靶蛋白进行泛素化。本发明中,发明人鉴定了ring型e3连接酶家族的成员sina,探究了它在陆地棉响应黄萎病过程中的作用。实验中选取24个ghsinas,其中ghsina7、ghsina8和ghsina9在侵染黄萎病菌24小时后表达量明显上调。这3个ghsinas都具有e3泛素连接酶活性并定位于细胞核。除此之外,这些ghsinas可以相互作用形成同源二聚体和异源二聚体。ghsina7、ghsina8和ghsina9基因分别
‑
异位超表达拟南芥提高了对黄萎病菌的抗性,然而单独敲除掉ghsina7、ghsina8和ghsina9降低棉花对病原体的抗性。因此,ghsina7、ghsina8和ghsina9在棉花防御黄萎病菌中起到正向调控作用。
15.如在一些实施方式中,基因ghsinas为基因ghsina7;在一些实施方式中,基因ghsinas为基因ghsina8;在一些实施方式中,基因ghsinas为基因ghsina9;在一些实施方式中,基因ghsinas为基因ghsina7和基因ghsina8;在一些实施方式中,基因ghsinas为基因ghsina7和基因ghsina9;在一些实施方式中,基因ghsinas为基因ghsina8和基因ghsina9;在一些实施方式中,基因ghsinas为基因ghsina7、基因ghsina8和基因ghsina9。
16.另外,需要说明的是,本发明中的基因ghsina7、ghsina8和ghsina9均分别存在于a亚组染色体和d亚组染色体,不同染色体上存在的同源基因对的序列高度相似,具体见上述基因ghsina7、ghsina8和ghsina9的两个核酸序列。通过实时荧光pcr(qrt
‑
pcr)是很难区分
这些同源基因对,因此,在后续研究其过表达和基因敲除时,均对每个同源基因设计一对引物来进行扩增。
17.进一步地,所述基因ghsinas包括基因ghsina8和基因ghsina9中的任一种或多种。
18.如在一些实施方式中,基因ghsinas为基因ghsina8;在一些实施方式中,基因ghsinas为基因ghsina9;在一些实施方式中,基因ghsinas包括基因ghsina8和基因ghsina9。
19.进一步地,所述基因ghsinas在棉花抗黄萎病遗传改良或分子育种方面的应用。
20.即本发明中的基因ghsinas能够用于棉花抗黄萎病遗传改良或分子育种方面。
21.现有的证据表明,增强棉花材料中基因ghsinas表达,获得基因ghsinas过表达的棉花种苗,该棉花种苗对黄萎病抗性增加。
22.具体地,所述棉花材料中,过表达基因ghsina7所用的扩增引物序列如seq id no:7和seq id no:8所示;
23.过表达基因ghsina8所用的扩增引物序列如seq id no:9和seq id no:10所示;
24.过表达基因ghsina9所用的扩增引物序列如seq id no:11和seq id no:12所示。
25.本发明通过对基因ghsinas分析,设计了用于过表达基因ghsinas所用的引物,扩增得到的目的基因片段经载体转入植物体中,能有效的稳定过表达基因ghsinas,且抗黄萎病性状表现明显。
26.基因过表达的基本原理是通过人工构建的方式在目的基因上游加入调控元件,使基因可以在人为控制的条件下实现大量转录和翻译,从而实现基因产物的过表达。
27.基因过表达的一般步骤如下:目的基因克隆,并选择特定的载体(含增强的启动子),将目的基因连接在特定的载体上;然后将载体导入生物材料中。
28.其中,载体介导转化方法,即将目的基因插入到农杆菌的质粒或病毒的dna等载体分子上,随着载体dna的转移而将目的基因导入到植物基因组中。另外,还可以通过其他途径,如种质系统法,包括花粉管通道法、生殖细胞侵染法、胚囊和子房注射法;又如基因直接导入法,通过物理或化学的方法直接将外源目的基因导入植物的基因组中,物理方法包括基因枪转化法、电激转化法、超声波法、显微注射法和激光微束法等,化学方法有peg介导转化方法和脂质体法等。
29.本发明中,涉及的基因过表达的具体内容按照常规步骤进行。
30.进一步地,所述棉花材料包括适宜于有性繁殖、无性繁殖或可再生的细胞的组织培养的材料。
31.具体地,所述棉花材料包括以下中的任一种:叶、根、茎、胚根、胚芽、种子。
32.上述内容中,棉花材料中的基因ghsinas的过表达,该处的棉花材料可以是种植或生长中的棉花组织,也可以是离体的用于棉花繁殖的材料。
33.本发明还提供了棉花耐黄萎病性能的检测方法,检测待测样品中基因ghsinas的表达情况,若基因ghsinas过表达,则判断为棉花耐黄萎病;
34.其中,所述基因ghsinas包括基因ghsina7、基因ghsina8和基因ghsina9中的任一种或多种;
35.基因ghsina7的核酸序列如seq id no:1和/或seq id no:2所示;
36.基因ghsina8的核酸序列如seq id no:3和/或seq id no:4所示;
hirsutum)、海岛棉(gossypium barbadense)、亚洲棉(gossypium arboreum)和草棉(gossypium herbaceum)以及这些棉花品种的变种。也就是说,存在与本发明基因ghsinas相同或类似基因的棉花品种均在本发明的保护范围内。类似基因如可以与本发明基因ghsinas序列相似度达80%以上,也可以是相似度达85%以上或90%以上或95%以上或96%以上或97%以上或98%以上或99%以上等等。
57.与现有技术相比,本发明的有益效果至少包括如下方面:
58.(1)本发明首次明确基因ghsinas在棉花抗黄萎病菌中的功能作用,为防治黄萎病侵染棉花提供了靶基因。
59.(2)本发明通过多方面试验得到ghsina7、ghsina8、ghsina9基因的过表达提高了转基因植物对黄萎病菌的抗性;敲除ghsina7、ghsina8、ghsina9基因降低植株对黄萎病菌的抗性,表明sinas在响应大黄萎病侵染中丧失和获得的功能与ghsinas在植物中对黄萎病菌的抗性是一致的。
60.(3)本发明发现的基因ghsinas可应用于防治棉花黄萎病,具体可用于棉花抗黄萎病遗传改良或分子育种方面,如培育抗黄萎病棉花品种。
附图说明
61.为了更清楚地说明本发明实施例或现有技术中的技术方案,以下将对实施例或现有技术描述中所需要使用的附图作简单地介绍。
62.图1为本发明实施例1中棉花sina泛素连接酶的保守结构域与发育系统分析图;
63.图2为本发明实施例2中抗性品种陆地棉中植棉2号在根部接种大丽轮枝菌24h后sina基因的表达水平柱形图;
64.图3为本发明实施例3中酵母双杂交实验用来检测3个ghsina蛋白之间的互作(ghsina7、ghsina8和ghsina9)结果图;
65.图4为本发明实施例3中ghsina7、ghsina8和ghsina9蛋白的体外泛素化分析结果图;
66.图5为本发明实施例3中通过egfp融合检测烟草表皮细胞中ghsina7、ghsina8和ghsina9蛋白的亚细胞定位结果图;
67.图6为本发明实施例4中陆地棉cla1基因作为沉默阳性对照的表型图;
68.图7为本发明实施例4中qrt
‑
pcr检测的转录水平证实ghsina7、ghsina8和ghsina9在不同vigs植株中的沉默效率结果柱形图;
69.图8为本发明实施例4中基因ghsina7、ghsina8和ghsina9抑制的特异性检测柱形图;
70.图9为本发明实施例4中侵染黄萎病菌v991菌株的ghsina7、ghsina8和ghsina9沉默植株进行病害症状分析;
71.图10为本发明实施例4中不同植株中疾病等级的分布;
72.图11为本发明实施例4中侵染黄萎病菌后,ghsina7、ghsina8和ghsina9基因沉默棉株和野生型棉株纵切茎的维管颜色变化情况;
73.图12为本发明实施例4中对照和沉默植株表面消毒的茎段进行真菌恢复实验结果图;
74.图13为本发明实施例4中沉默植株和对照植株在接种黄萎病菌后的相对真菌生物量柱形图;
75.图14为本发明实施例4中独立过表达ghsina7、ghsina8和ghsina9的拟南芥植株的情况相关图。
具体实施方式
76.下面将结合实施例对本发明的实施方案进行详细描述,但是本领域技术人员将会理解,下列实施例仅用于说明本发明,而不应视为限制本发明的范围。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
77.部分不同实施例中共同涉及的材料和方法的说明
78.1、植株材料和生长条件
79.以陆地棉的中植棉2号为实验材料,在温室营养土中播种,生长条件为28℃、16h光照/8h黑暗条件。
80.将拟南芥野生型col
‑
0种子种在ms培养基上,之后将幼苗移栽至含有营养土和蛭石的花盆中,放置在23℃培养室内,光周期条件为16h光照/8h黑暗。
81.烟草幼苗在温室25℃、16h光照/8h黑暗条件下培养大约6周。
82.2、病原菌制备及接种处理
83.将大丽轮枝菌的菌株v991培养在25℃马铃薯葡萄糖琼脂(pda)培养基活化4天,之后在25℃,180rpm条件下,将活化后的黄萎病菌v991接种于察氏培养基中培养5天,用四层纱布过滤菌液。收集过滤后的菌液,通过血球计数板计算,将孢子浓度稀释至1*106个/毫升,备用。采用蘸根法,将在水培条件下培养两周的棉花幼苗根部接种孢子悬液(10ml),24h后收集用于rna提取。对于sina沉默的棉花植株,使用1ml注射器注射到棉花子叶下1厘米处的下胚轴处(3μl)。拟南芥的接种是将4周大的幼苗轻轻从土壤中拔出,用无菌水小心冲洗根部并浸泡在孢子悬浮液中(4
×
105个分生孢子/毫升)2分钟。之后将植株转移到灭过菌的新鲜土壤中,继续在23℃、16h光照/8h黑暗的温室下培养观察。
84.3、疾病指数、病原菌恢复
85.病害指数di的计算公式为:di=[(σ病害等级
×
染病株数)/(总株数
×
4)]
×
100。在黄萎病菌侵染后,根据疾病严重程度,幼苗被分为0、1、2、3、4和5五个等级。
[0086]
将对照组和sina沉默棉花植株经表面杀菌处理后,取第一节间切片分别切成3~5mm的薄片并在25℃pda培养基上培养3天。为了检测大丽轮枝菌的繁殖程度,在体式显微镜下解剖观察子叶节的纵切面(leica,wetzlar,germany)。
[0087]
4、真菌生物量的测量
[0088]
棉花和拟南芥接种黄萎病菌后,采集棉花植株的茎和拟南芥的根进行dna提取。利用真菌特异的its1
‑
f引物及大丽轮枝菌特异性反向引物stve1
‑
r检测棉花及拟南芥侵染黄萎病菌后组织中的菌含量。利用棉花histone3和拟南芥actin2基因作为内参。对基因组dna进行实时荧光定量pcr分析。
[0089]
5、rt
‑
pcr和qrt
‑
pcr分析
[0090]
使用trizol试剂(tiangen,北京,中国)从棉花或拟南芥的不同组织中提取总rna。
用1μg总rna逆转录成cdna(r323版本vazyme,南京,中国)a。rt
‑
pcr分析如前所述。qrt
‑
pcr检测采用sybr green real
‑
time pcr master mix(vazyme),以棉花histone3或拟南芥actin2基因为内参。采用德国罗氏公司的lightcycler480体系检测目标基因表达量。
[0091]
实施例1
[0092]
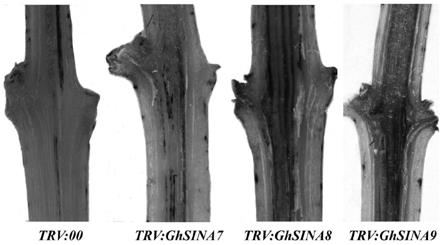
生物信息学分析
[0093]
为了研究棉花sina泛素连接酶的作用,在棉花基因组数据库中共鉴定12个亚洲棉、12个雷蒙德氏棉和24个陆地棉sina基因。其中,3个棉花品种陆地棉、亚洲棉和雷蒙德氏棉从cottonfgd数据库(http://www.cottonfgd.org/)中获得。之后,克隆了陆地棉sina基因,在其初始注释的基础上对测序获得的ghsina8编码序列进行了修改。这些sinas编码两个典型的保守结构域,一个ring指域(位于氨基端))和一个sina结构域(位于羧基端)(图1a))。为了研究陆地棉sina同源结构域序列及每个残基的保守频率,进行了序列比对并获得了sina蛋白序列特征,如图1b所示,序列标识图表明陆地棉sina泛素连接酶含有高度保守的ring指域和sina结构域。通常情况下,ring指域含有39个保守的基础氨基酸残基,而sina结构域含有200个保守残基,这些残基负责与特定目标底物相互作用。
[0094]
为了进一步研究棉花与拟南芥sina蛋白的进化关系,构建了进化树。其中,5个拟南芥sinas氨基酸序列(sinat1
‑
at2g41980、sinat2
‑
at3g58040、sinat3
‑
at3g61790、sinat4
‑
at4g27880和sinat5
‑
a t5g53360)采用blastp程序在3个棉花蛋白数据库查询,筛选候选的棉花sina蛋白。随后pfam(https://www.pfam.xfam.org)和interpro数据库(https://www.ebi.ac.uk/interpro/)对候选ghsina蛋白筛选,以确定是否存在ring finger和sina结构域。系统发育分析采用mega 7.0软件,将棉花和拟南芥的所有sina蛋白序列,通过1000个bootstrap重复的邻域连接法(nj)构建系统发育树,树杈上的数字表示引导值。使用clustalx 2.0软件比对陆地棉sinas的ring finger和sina保守结构域蛋白序列。并将比对结果上传至在线工具weblogo(http://weblogo.berkeley.edu/logo.cgi)分析其具体氨基酸特征。
[0095]
所有被鉴定的sina蛋白被分成两个亚组(图1c)。较大的亚组中含有32个棉花sina基因(其中来自雷蒙德氏棉、亚洲棉和陆地棉的分别是8、8和16个)。陆地棉sina基因数量是二倍体棉亚洲棉(aa)和雷蒙德氏棉(dd)的两倍。这种现象与异源四倍体陆地棉(aadd)产生多倍体事件的结果一致,该多倍体来源于二倍体前体亚洲棉和雷蒙德氏棉的自然杂交。第二组是由2个atsinats和16个sinas(其中雷蒙德氏棉、亚洲棉和陆地棉分别是4、4和8个)组成,这表明sina基因家族在棉花基因组进化过程中明显地扩张。
[0096]
实施例2
[0097]
ghsina基因响应黄萎病的表达分析
[0098]
gfp标记的黄萎病菌侵染陆地棉24h后,检测到棉苗根上分生孢子已经萌发和黄萎病菌丝的生长延伸。为了确定ghsina基因在侵染黄萎病24h后表达水平是否变化,研究了它们在陆地棉抗性品种(中植棉2号)根系中的转录水平,涉及的引物如表1所示。其中,ghsina7、ghsina8和ghsina9均分别存在于a亚组染色体和d亚组染色体,但是它们同源基因对的序列高度相似,通过实时荧光pcr(qrt
‑
pcr)是很难区分这些同源基因对,因此,它们一起被扩增。图2显示了抗性品种中植棉2号在根部接种大丽轮枝菌24h后sina基因的表达水平,其在14日龄的幼苗接种大丽轮枝菌24h后从根部提取总rna,误差条代表三次生物学重
复,棉花histone3作为内参基因,星号表示差异有统计学意义,显著性差异用student’s t
‑
test分析(**p<0.01)。从图2可以看出,与对照组相比,ghsina7、ghsina8和ghsina9基因的表达量在接种后24h显著上调,大约分别被诱导3.5、3.2和2倍,总的来说,ghsinas基因转录水平是高于对照组的。因此,之后重点研究陆地棉ghsina7、ghsina8和ghsina9基因对黄萎病菌的调控。
[0099]
表1不同ghsina基因的qrt
‑
pcr涉及的引物
[0100][0101][0102]
实施例3
[0103]
1、酵母双杂交实验
[0104]
根据matchmaker gold酵母双杂交系统(clontech,palo alto,ca,usa)的说明构建酵母双杂筛选。ghsinas全长cdnas(具体涉及的引物见表2)分别克隆到诱饵载体pgbdk7和猎物载体pgadt7中。将构建的重组质粒共转化到酵母菌y2hgold中,于30℃sd/
‑
leu/
‑
trp do(ddo)培养基上培养3天。挑取大小相同的独立菌落转移到含有x
‑
α
‑
gal(clontech)的sd/
‑
leu/trp/
‑
ade/
‑
his do(qdo)培养基中,通过菌落是否变蓝,以评估ghsina蛋白之间的相互作用。
[0105]
表2用于不同载体的ghsinas引物
[0106][0107]
sina蛋白通过形成同源二聚体或异源二聚体来行使生物学功能。为了确定这三种候选ghsinas蛋白是否有形成同源或异源二聚体的能力,酵母双杂交实验对ghsina7、ghsina8和ghsina9之间的相互作用进行了鉴定。ghsina7、ghsina8和ghsina9基因的全长cdnas被独立融合到含dna结合结构域(bd)的诱饵载体pgbkt7或含激活结构域(ad)的猎物载体pgadt7,然后共转化到y2hgold中,通过在缺陷型培养基上生长情况,初步鉴定它们之间的互作关系。正如图3所示,转化酵母细胞在sd/
‑
leu/
‑
trp do(ddo)培养基和sd/
‑
leu/
‑
trp/
‑
ade/
‑
his do(qdo)培养基(含x
‑
α
‑
gal)上生长,即每个ghsina都分别与自身以及其它两种ghsinas互作形成同源或异源复合物。以ad
‑
t和bd
‑
53的相互作用作为阳性对照,验证y2h实验的有效性。
[0108]
2、体外泛素化分析
[0109]
将ghsina7、ghsina8和ghsina9基因的全长开放阅读框(orfs)独立克隆(涉及的引物如表3所示)到pmal
‑
c2x载体中,生成带有麦芽糖结合蛋白(mbp)融合表达载体。重组质粒在大肠杆菌bl21菌株中表达,使用直链淀粉树脂(neb,ipswich,ma,usa)亲和层析纯化,并如之前所述用于体外泛素化分析。纯化融合蛋白mbp
‑
sinas(3μg)在30μl泛素化反应缓冲液(50mmtris
‑
hcl,ph值7.5,5mm mgcl2,2mm dtt和2mm atp),5μg生物素标签的泛素(enzo,#bml
‑
uw9920
‑
0001),100ng e1(enzo,170#bml
‑
uw9920
‑
0001)和40ng human e2(ubch5b)在30℃下独立孵化3h(enzo,171#bml
‑
uw9920
‑
0001)。加入5
×
样品缓冲液终止反应,用7.5%的sds
‑
page凝胶分离一半的混合物。使用抗生物素抗体(细胞信号1:3000稀释)进行western blotting鉴定。之后在tanon
‑
5200化学发光成像系统(ai600 uv,美国)显影。
[0110]
表3 ghsina7、ghsina8和ghsina9基因的orf克隆引物
[0111][0112]
e3连接酶与e2泛素结合酶结合,并具有自身泛素化的功能酶活性。为了确定ghsina7、ghsina8和ghsina9蛋白是否具有e3连接酶活性。在大肠杆菌bl21菌株中构建了含有mbps标签的sinas表达载体,对其表达纯化,获得融合蛋白mbp
‑
sinas。随后进行泛素化活性分析,当存在human e1(uba1)、e2(ubch5b)、生物素标记泛素(bt
‑
ub)和mbp
‑
sinas的情况下,使用生物素抗体(anti
‑
biotin)能检测到高分子量的自泛素多聚条带(图4a
‑
c,第1列),这就表明了mbp
‑
sinas被泛素化。当mbp取代mbp
‑
sinas,或者当e1、e2或是生物素标记的泛素缺失时,并没有多聚泛素化信号(图4a
‑
c,2
‑
6列)。很明显地,ghsina8比ghsina7或ghsina9具有更强的e3连接酶活性。这些结果表明这三个sina蛋白都含有e3泛素连接酶活性。ghsina蛋白是功能性e3泛素连接酶。
[0113]
图4中,a
‑
c是在e1、e2和生物素化泛素(bt
‑
ub)(1列)存在下,检测mbp
‑
sina7(a)mbp
‑
sina8(b)和mbp
‑
sna9(c)融合蛋白的e3泛素连接酶活性,样品经7.5%sds
‑
page分离,用抗生物素抗体western blotting(wb)检测,mbp作为阴性对照,考马斯亮蓝染色反应中存在等量的e3酶。
[0114]
3、烟草叶片的亚细胞定位
[0115]
在修饰的pcambia2300
‑
egfp表达载体中,ghsina7、ghsina8和ghsina9基因的开放框分别与egfp融合,ghsina7、ghsina8和ghsina9涉及的引物如表4所示。含重组质粒的农杆菌(菌株gv3101)菌液注射渗透到烟草叶片中。将其在25℃暗条件下培养48h,用激光共聚焦显微镜(olympus,德国)观察荧光信号,比例尺=20μm。
[0116]
表4亚细胞定位涉及的ghsina7、ghsina8和ghsina9扩增引物
[0117][0118]
泛素化通常发生在细胞核及细胞质中,分别控制核蛋白和细胞质蛋白来发挥生物学功能。为了确定ghsina7、ghsina8和ghsina9蛋白的亚细胞定位,在camv 35s组成型启动
子诱导的ghsina7/8/9
‑
egfp构建融合体分别在烟草叶片中转瞬表达。如图5所展示,在细胞核中观察到了游离的egfp绿色荧光。显然,这三个sina蛋白存在于细胞核中,ghsina蛋白定位在细胞核,这与它们在泛素化通路中的功能一致。
[0119]
实施例4
[0120]
1、沉默ghsinas基因降低了棉花对黄萎病的抗性
[0121]
(1)棉花病毒诱导的基因沉默(vigs)
[0122]
ptrv1和ptrv2载体被用于vigs实验。扩增ghsinas的特异片段(涉及的引物如表5所示),并将其连接ptrv2载体。将重组质粒转化农杆菌gv3101。随后将含有ptrv1和ptrv2
‑
ghsina质粒的农杆菌菌液(od600=1)按1:1的比例混合,使用注射器将上述菌液注射到7日龄棉花幼苗的两个完整子叶中。
[0123]
表5沉默基因ghsina7、ghsina8和ghsina9的扩增引物
[0124][0125][0126]
(2)结果
[0127]
在棉花研究中广泛应用基因沉默的有效方法是vigs。本文采用vigs技术研究ghsinas基因在调控棉花黄萎病抗性的作用。因此,用vigs特异性沉默这三个ghsina基因(trv2:ghsina7、trv2:ghsina8和trv2:ghsina9)来研究它们在响应黄萎病中的功能。以沉默时产生明显白化表型的trv:ghcla1和空白trv:00分别作为阳性对照和阴性对照。农杆菌菌液渗透棉花叶片7天后,陆地棉cla1基因被沉默后,棉花叶片表现出预期的白化表型(图6),说明vigs实验进行得很好。分别对不同的沉默株系中沉默基因表达量进行检测(涉及的引物如表1中的ghsina7、ghsina8和ghsina9所示)。与trv:00处理的植株相比,vigs植株中,ghsina7、ghsina8和ghsina9基因的表达水平分别显著降低(图7)。为了研究vigs介导的三种sinas对基因ghsina7、ghsina8和ghsina9抑制的特异性,检测了每种基因沉默植物中另外两个与沉默ghsina编码序列具有高相似性的非靶向sina基因的转录水平。两个非靶向性的sina基因的表达水平在每个特异性沉默的植物中都没有受到影响(图8)。
[0128]
随后,对照植株和沉默植株受到黄萎病菌侵染。大约两周之后,基因沉默的棉花幼苗,特别是含有trv2:ghsina8和trv2:ghsina9的幼苗,表现出更严重的叶片枯萎、黄化和落叶症状,甚至死亡(trv:00)(图9)。统计dis,大部分trv2:ghsina8和trv2:ghsina9植株出现了更严重的病害病变(图10),图10中,0级:健康叶片;1级:叶片0%
‑
25%枯萎;2级:叶子25%
‑
50%枯萎或褪绿;3级:叶片50%
‑
75%褪绿或坏死;4级:叶子死亡或脱落,实验进行3次重复,每次重复至少40株幼苗,星号表示秩检验判定差异有统计意义(*p<0.05,**p<0.01)。此外,与trv:00相比,ghsina7、ghsina8和ghsina9基因沉默棉株比野生型棉株茎的维管颜色变深,特别是ghsina8和ghsina9沉默的棉花植株中有更多坏死的维管组织(图11)。真菌恢复试验证实,ghsina8和ghsina9沉默的植株比对照组有更多的真菌繁殖(图
12)。相应地,ghsina8和ghsina9沉默植株的真菌生物量显著高于对照植株(图13)。图13通过qrt
‑
pcr分析,比较了黄萎病菌内部转录间隔区(its)与棉花histone3之间的dna水平,v值代表三个生物学重复的平均值
±
sds(**p<0.01,student’s t检验)。综上所述,沉默ghsina7、ghsina8和ghsina9可以抑制植物免疫系统,增强植物对黄萎病的敏感性。
[0129]
2、ghsinas基因过表达提高了转基因拟南芥对黄萎病的抗性
[0130]
(1)、质粒构建与拟南芥转化
[0131]
克隆ghsinas的开放框(涉及的引物如表6所示)并将其独立插入含有camv 35s组成启动子的载体pcambia2300(cambia)中。所有构建体均经测序确认后,分别导入根癌农杆菌gv3101。利用根癌农杆菌介导的花序浸染法对哥伦比亚野生型拟南芥进行了转化。在含有卡那霉素的ms培养基上筛选转基因拟南芥,利用基因组pcr技术对所选转基因幼苗进行进一步筛选。获得纯合子t3转基因株系进行功能分析。
[0132]
表6过表达涉及的基因ghsina7、ghsina8和ghsina9扩增引物
[0133][0134]
(2)结果
[0135]
采用基因过表达策略来评估ghsinas基因在防御响应中的功能。由于棉花转化周期较长,本试验中以模式植物拟南芥作为试验材料。期间我们获得了超过18个独立过表达的ghsina7、ghsina8和ghsina9转基因拟南芥株系。选择了sina表达量(涉及的引物如表7所示)最高的两个独立纯合子t3系进行表型分析(图14a),图14a通过rt
‑
pcr鉴定独立过表达ghsina7、ghsina8和ghsina9的转基因拟南芥系,以拟南芥actin2为内参对照。种植4周后,sinas过表达的转基因植株接种黄萎病进行抗性测定。接种两周后植株开始出现病害症状,总体上,比起野生型植株,转基因植株叶片表现出更强的抗性(图14b),图14b为野生型和ghsina7、ghsina8、ghsina9转基因植株接种黄萎病后的症状图。ghsina7转基因株系的抗病性较野生型有所提高,但远远低于ghsina8和ghsina9转基因株系。图14c为野生型和转基因拟南芥植物坏死叶片的病害率统计,误差线代表3次生物学重复(n≥32),星号表示差异有统计学意义,显著性差异用student’s t
‑
test分析(*p<0.05,**p<0.01),可以看出,ghsinas转基因株系的坏死率明显低于wt(图14c)。此外,图14d为真菌生物量的定量测定结果柱形图,采用qrt
‑
pcr分析比较黄萎病菌内部转录间隔区(its)和拟南芥actin2基因之间的转录水平,其中,星号表示差异有统计学意义,显著性差异用student’s t
‑
test分析(*p<0.05,**p<0.01)。真菌生物量试验证明转基因植株根系中积累的真菌dna较少,尤其是在ghsina8和ghsina9过表达株系中(图14d)。因此,与野生型相比,ghsinas的异位过表达大大增强了拟南芥对黄萎病菌的抗性。
[0136]
表7过表达检测涉及的引物
[0137][0138][0139]
其中,沉默ghsinas降低了棉花对黄萎病的抗性和ghsinas过表达提高了转基因拟南芥对黄萎病的抗性的研究中,真菌生物量检测所用的引物如表8所示。
[0140]
表8真菌生物量检测所用的引物
[0141]
引物名称引物its1
‑
faaagttttaatggttcgctaagastve1
‑
rcttggtcatttagaggaagtaa
[0142]
另外,独立过表达ghsina7、ghsina8和ghsina9的转基因棉花株系,初步显示具有与拟南芥一致的抗黄萎病菌性状。
[0143]
尽管已用具体实施例来说明和描述了本发明,然而应意识到,在不背离本发明的精神和范围的情况下可以作出许多其它的更改和修改。因此,这意味着在所附权利要求中包括属于本发明范围内的所有这些变化和修改。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。

