调控拟南芥种子含油量和千粒重的基因atoil3
技术领域
1.本发明属于植物基因工程领域,具体涉及一种拟南芥含油量调节基因 atoil3,通过基因工程技术提高atoil3基因的表达量能够提高拟南芥种子含油 量及千粒重,达到增加种子含油量的目的。
背景技术:
2.随着人民生活水平的不断提高,中国油脂需求快速增长(刘成,冯中朝,肖 唐华,等.我国油菜产业发展现状,潜力及对策[j].中国油料作物学报,2019, 41(4):485.),人均食用植物油消费量从1996年的每年不足8kg,上升至2016年 的每年24.8kg(王佳友,何秀荣,王茵.中国油脂油料进口替代关系的计量经济 研究[j].统计与信息论坛,2017,32(5):69-75.)。在国内油料产量有限的 情况下,增加油料进口成为满足国内需求的主要途径(何杰夫,张博.中国食用 植物油的供应量和消费量究竟是多少?[j].中国农村经济,2011(4):87-92.)。 2017年全国植物油消费总量为3565万吨,其中,我国自产植物油为1100万吨, 自给率仅为30.8%,国产油料难以满足消费需求,供需缺口与日俱增(王汉中.我 国油菜产业发展的历史回顾与展望[j].中国油料作物学报,2010,32(2):300-302.)。
[0003]
我国油菜常年种植面积约1.1亿亩(张蓓蓓,马颖,耿维,等.中国油菜秸 秆资源的生物质能源利用潜力评价[j].可再生能源,2017,35(1):126-134.), 菜籽油占国产油料作物产油量的57%以上,是目前国产食用植物油的第一大来 源,在维护国家食用油供给安全战略中居于核心地位(刘成,冯中朝,肖唐华,等. 我国油菜产业发展现状,潜力及对策[j].中国油料作物学报,2019,41(4):485.)。 然而,与加拿大、澳大利亚等国家比,目前我国的菜籽含油量低、品质差、产量 低、生产成本高导致农民种植积极性不高。现阶段急需高产油量的新品种来提高 油菜籽的市场竞争力。油菜籽含油量每增加1个百分点,相当于提高产量2.5个 百分点(王汉中,殷艳.我国油料产业形势分析与发展对策建议[j].中国油料作 物学报,2014,36(3):414r421.)。大力发展油菜生产,是有效应对我国大豆进口不 确定性、维护国家食用油和饲用蛋白供给安全的战略举措。因此,大力提高油菜 含油量,培育高产高含油量品种具有重要的意义。
[0004]
脂肪酸和油脂的生物合成途径在植物体内研究的已较为透彻,从不同物种中 已经有相当多的相关基因得到分离和鉴定,研究也表明不同物种脂肪酸和油脂合 成的化学途径基本上是相同的(lung s c,weselake r j.diacylglycerol acyltransferase:a key mediator of plant triacylglycerol synthesis[j].lipids,2006, 41(12):1073-1088.)。种子油脂合成的主要前提包括乙酰辅酶a,nad(p)h和atp, 这些合成前提的来源和调控直接影响油脂积累的速率和数量(voelker t,kinney a j.variations in the biosynthesis of seed-storage lipids[j].annual review of plant biology,2001,52(1):335-361.)。植物油脂代谢是一个由多基因参与的复杂生物学 过程,存在着多个调控点与调控因子如关键转录因子、脂肪酸合成与转运、tag 的积累、糖酵解、乙酰辅酶a、磷酸戊糖途径、淀粉的合成与降解等(夏晗,王 兴军,李孟军,等.利用基因工程改良植物脂肪酸
和提高植物含油量的研究进展 [j].生物工程学报,2010,26(6):735r743.)。影响油脂合成的关键转录因子包括 wri1、lec1、lec2。lec1过量表达油菜转基因植株中,参与糖酵解、质体脂 肪酸合成以及油脂积累的关键基因上调,脂肪酸含量大幅增加(tan h,yang x, zhang f,et al.enhanced seed oil production in canola by conditional expression of brassica napus leafy cotyledon1 and lec1-like in developing seeds[j].plant physiology,2011,156(3):1577-1588.)。lec2是植物特有的b3转录因子家族,其 转录水平与油脂的积累成正相关(kim h u,jung s j,lee k r,et al.ectopic overexpression of castor bean leafy cotyledon2(lec2)in arabidopsis triggers the expression of genes that encode regulators of seed maturation and oil body proteins in vegetative tissues[j].febs open bio,2014,4(1):25-32.)。玉米中过表达wri1和 lec1同样导致含油量的提高(shen b,allen w b,zheng p,et al.expression of zmlec1 and zmwri1 increases seed oil production in maize[j].plant physiology, 2010,153(3):980-987.)。然而,由于多数转录因子的功能丰余性增加了研究调控 油脂含量的复杂性,目前仍有一些转录因子的作用机制未知。
[0005]
在碳源分配比例研究中,同是芸薹属的油菜种皮和模式植物拟南芥种皮都会 积累大量粘性果胶。拟南芥降低粘性多糖的合成会使碳源直接流向tag的合成, 导致种子中含油量的增加。在油菜和拟南芥中的研究结果显示,种子内的光合作 用对含油量的贡献率在40%左右(fernando 2005,setsuko 2008)。而对磷酸戊糖 途径的抑制反而增加了种子的含油量,表明对于其他提供用于脂肪酸合成的还原 力nadph,抑制磷酸戊糖途径来增加碳源可以更有效的增加油脂含量(setsuko 2008)。拟南芥中异源表达油菜wri1,种子含油量最高可以提高40%(vanhercket,el tahchy a,shrestha p,et al.synergistic effect of wri1 and dgat1 coexpressionon triacylglycerol biosynthesis in plants[j].febs letters,2013,587(4):364-369.)。
[0006]
本发明中,申请人通过分析油菜含油量差异显著的两个油菜品系各组织转录 组的基因表达差异,筛选出一类在高油材料中高表达,在低油材料中低表达的基 因。通过拟南芥同源基因的序列比对,基因全长的克隆,表达载体的构建,模式 作物拟南芥的遗传转化,最终获得一个可调控种子含油量变化的基因。同时,该 基因在拟南芥中的过表达还提高了种子的千粒重。将其应用于作物育种中,可以 大幅增加油料作物的产油量。
技术实现要素:
[0007]
本发明的目的在于提供了一种拟南芥含油量调节基因atoil3及其应用,其 核苷酸序列如seq id no.1所示,其编码的蛋白质氨基酸序列如seq id no.2 所示,在拟南芥中过表达atoil3基因不但提高了种子含油量,同时也增加了种 子的大小,千粒重变大。
[0008]
为了实现上述目的,本发明采用以下技术方案:
[0009]
一、基因的来源
[0010]
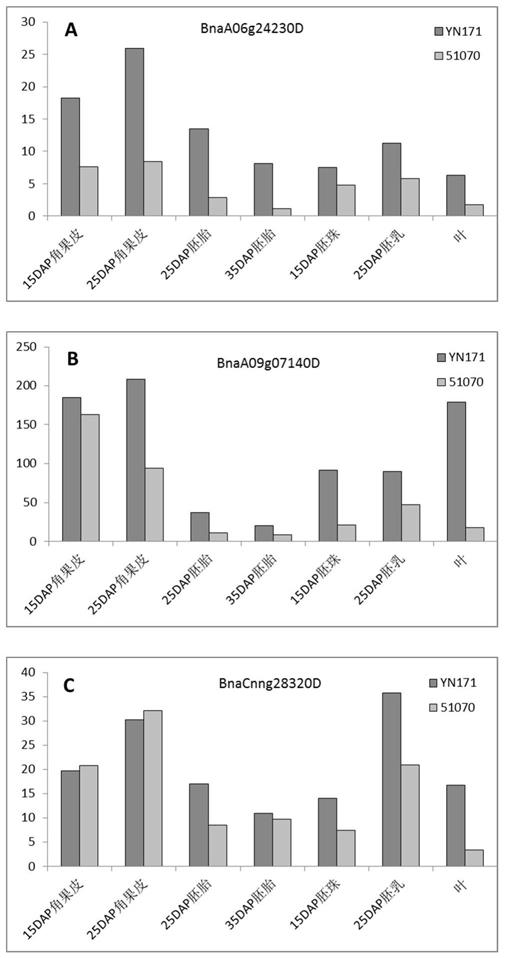
在两个种子含油量存在差异的油菜品系yn171(63.8
±
2.4)和51070(35.9
ꢀ±
1.6)的不同组织转录组分析中得到差异表达基因bnaa06g24230d、 bnaa09g07140d和bnacnng28320d,都表现为在高油品种yn171中的表达水平 要显著性高于低油品种51070(具体见图1)。序列比对发现这3个基因序列相似 性较高,为同源基因。经拟南芥数据库序
列比对,得到拟南芥同源基因at5g65300 核酸序列,命名此类基因为atoil3,该类基因为功能未知基因,且不含有任何已 公布的蛋白保守区域。
[0011]
二、基因的全长克隆
[0012]
以拟南芥参考序列设计基因全长引物,atoil3-f:5'-atggaatgcagaaa acacaa-3'(seq id no.3),atoil3-r:5'-ttaataaactcgttgccgtg-3' (seq id no.4),以哥伦比亚野生型拟南芥cdna为模板进行pcr扩增,经 测序验证,得到该基因的全长产物,核苷酸序列如seq id no.1所示,其编码 的蛋白质氨基酸序列如seq id no.2所示。
[0013]
三、植物过表达载体构建
[0014]
带有与所用骨架载体酶切位点两端重组的核苷酸序列,设计重组引物 35s::atoil3-f:5'-ggacagcccagatcaactagtatggaatgcagaaaacacaa-3', 35s::atoil3-r:5'-gcccttgctcaccatggatccttaataaactcgttgccgtg-3',扩增 得到的pcr产物经纯化回收后保存备用。用spei和bamh i双酶切植物表达载 体pcambia1305.1-gfp,并经1%琼脂糖凝胶电泳检测回收;将atoil3的基因 pcr纯化回收产物与回收的酶切质粒按一步克隆试剂盒说明书进行操作,37℃ 水浴30min后转化大肠杆菌感受态dh5α;长出的单克隆以载体上游引物35s: 5'-gacgcacaatcccactatcc-3'和下游引物nos:5'-gataatcatcgcaa gaccgg-3'进行阳性克隆测序检测,提取阳性克隆的表达载体质粒,该质粒即 为重组载体35s::atoil3。
[0015]
四、拟南芥的遗传转化及表型分析
[0016]
通过农杆菌介导的浸花法转化野生型拟南芥,经三代筛选,得到纯合转基因 株系。利用核磁共振法对收获的纯合体转基因种子进行含油量检测,同时比较种 粒大小的变化发现,通过基因工程技术提高atoil3基因的表达量能够提高拟南 芥种子的含油量和种粒大小,达到增加产油量和种粒大小的目的。经实验证实, 含atoil3的转基因拟南芥株系的含油量与受体对照(非转基因植株)相比均有 所增高,平均增幅最高为6.73%左右,千粒重平均增幅为13.56%左右。
[0017]
与现有技术相比,本发明具有以下优点及有益效果:
[0018]
本发明是国内首次公开atoil3基因在提高拟南芥含油量和千粒重中的作用。 利用两个油菜材料的不同组织转录组数据及种子含油量数据进行关联分析,获得 影响油菜种子含油量的候选基因,经拟南芥数据库序列比对和设计引物,进一步 获得拟南芥含油量调节基因atoil3。本发明实验结果表明转基因拟南芥种子含 油量与受体对照(非转基因植株)相比均有所增高,平均增幅最高为6.73%左右, 千粒重平均增幅为13.56%左右。该基因为油菜高油育种提供了新的基因源,同 时将其扩展到其他油料作物如大豆、花生、芝麻等育种的应用中,提高油料作物 的产油量。
附图说明
[0019]
图1.候选基因在不同含油量油菜中的转录组数据分析图
[0020]
高含油量油菜品种yn171(含油量63.8
±
2.4);低含油量油菜品种51070 (35.9
±
1.6);dap:day after pollination授粉后天数;a:bnaa06g24230d;b: bnaa09g07140d;c:bnacnng28320d。
[0021]
图2.油菜候选基因与拟南芥同源基因的序列比对图示
[0022]
a:bnaa06g24230d、bnaa09g07140d、bnacnng28320d和at5g65300 (atoil3)的核苷
基酸序列如seq id no.2。提取测序正确的阳性单菌落的表达载体质粒,该质 粒即为重组载体35s::atoil3(t-dna示意图如图3所示)。
[0030]
实施例3:拟南芥遗传转化及纯合转化株系筛选
[0031]
利用冻融法将上述构建完毕的植物过表达载体35s::atoil3转化农杆菌感受 态细胞gv3101,步骤如下:1)将保存于-80℃的农杆菌感受态细胞gv3101置 于冰上融化;2)用移液枪取3μl(100ng)表达载体质粒,浸没枪头加入到感受 态细胞中,置于冰上静止30min,液氮中急冻1min,然后在37℃恒温水浴锅中 水浴5min;3)加入600μl lb液体培养基(胰蛋白胨10g;酵母提取物5g; nacl 10g),28℃,200rpm,振荡培养4h;4)涂布于附加50μg/ml卡那霉素、 50μg/ml庆大霉素和50μg/ml利福平的固体lb培养基((胰蛋白胨10g;酵母 提取物5g;nacl 10g;agar 1.5%))上,28℃倒置培养36-48h;5)长出的单 克隆以植物表达载体上游引物35s:5'-gacgcacaatcccactatcc-3'和下游 引物nos:5'-gataatcatcgcaagaccgg-3'进行阳性克隆检测,经pcr检 测为阳性的单克隆摇菌至od600=1.8-2.0(紫外分光光度计检测),用等体积的50% 甘油保菌于-80℃超低温冰箱,以备后续研究使用。
[0032]
利用浸花法转化拟南芥:1)拟南芥抽薹后剪去主花絮顶端,待侧枝生长一 致且处于花蕾期时,准备材料进行转化;2)用200ml含有50μg/ml卡那霉素、 50μg/ml庆大霉素和100μg/ml利福平的lb液体培养基,接种携带目的基因的 农杆菌,28℃,200rpm,培养12-18h;3)菌液装于离心瓶中,5000rpm,离心 15min,用移液枪将上清吸走并弃之;4)用100ml重悬液(5%的蔗糖,0.02% 的表面活性剂),重悬农杆菌;将拟南芥花絮浸入菌液中30s,并轻柔搅动;5) 浸染完后,用塑料薄膜覆盖过夜,提高转化效率,可在5-7d后再转化一次;6) 大约一个月后,种子成熟,收获后置于37℃烘箱7d,脱粒后于4℃春化3d,标 注为t0代种子。
[0033]
筛选阳性转基因植株:1)取适量(不超过ep管的1/5)脱粒干净的拟南 芥种子于1.5ml ep管中;2)加1ml 75%酒精,震荡洗涤1min,8000rpm离 心30s,用移液枪将上清吸走并弃之;3)加1ml10%的次氯酸钠,震荡洗涤5 min,8 000rpm离心30s,用移液枪将上清吸走并弃之;4)加1ml dd h2o, 震荡洗涤1min,8 000rpm离心30s,用移液枪将上清吸走并弃之;5)重复步 骤4三次,加1ml ddh2o,于黑暗条件下,4℃,春化3d;6)铺板种植于 1/2ms培养基(ms粉2.15g;蔗糖10g;琼脂0.8%;ph 5.8)上,含25mg/l的潮霉素,培养箱暗培养5d左右,转换为正常光照培养;7)等长出两 片真叶时,将下胚轴伸长、真叶生长正常的移苗于温室培养钵中(营养土:蛭石: 珍珠岩=3:1:1);8)活下来正常生长的植株待抽薹前取叶片抽提基因组dna, 以植物表达载体上游引物35s:5'-gacgcacaatcccactatcc-3'和下游引物 nos:5'-gataatcatcgcaagaccgg-3'筛选阳性转基因植株,经pcr检测 为阳性的转基因材料为t1代,单株收获,并依次编号保存。同样将t1代转基 因材料经抗潮霉素筛选与pcr鉴定为t2代,单株收获,并依次编号保存。之 后再次将t2代筛选,得到纯合的转基因株系。
[0034]
实施例4:转atoil3基因拟南芥种子含油量与种粒大小鉴定
[0035]
利用核磁共振法对收获的纯合体转基因种子进行含油量检测,同时比较种粒 大小的变化发现,通过基因工程技术提高atoil3基因的表达量能够提高拟南芥 种子的含油量和千粒重,达到增加产油量和千粒重的目的。经实验证实,含 atoil3的转基因拟南芥株系的含油量与受体对照(非转基因植株)相比均有所 增高,平均增幅最高为6.73%左右(如图4所示),种粒大小明显变大,千粒重 平均增幅为13.56%左右(如图4所示)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。

