γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法
技术领域
1.本发明涉及一种提高植物抗逆性和产量的方法,尤其涉及一种γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法,属于基因工程、遗传育种、合成生物学技术领域。
背景技术:
2.干旱、土壤盐渍化等非生物胁迫已经成为世界上许多国家、地区农作物生产的主要限制因素。在中国,约一半的国土面积处于干旱和半干旱区。随着全球气温升高,我国水资源愈发短缺,干旱对农作物产量的影响越来越大。此外,土壤盐渍化也是影响作物产量的主要环境因素。目前世界上约20%耕地和近50%灌溉用地受到盐渍的严重危害,中国盐渍化土地约占全球盐渍土面积的10%。利用现代生物学技术培育抗旱、抗盐的优质高产作物新品种具有重要的战略意义,是提高作物产量、保障粮食安全和农业可持续发展的重要选择。而开发抗逆且高产的优质基因并通过转基因工程改造农作物是避免其在逆境条件下减产的有效途径。
3.植物转基因技术应用十分广泛,主要应用在抗虫、抗除草剂、抗逆和提高产量等方面。随着1995年世界第一例转基因抗虫玉米在美国获得商业化种植批准,越来越多的转基因作物在美国、巴西等国家开始大面积推广种植。
4.目前,国际上转基因抗旱作物的研究大部分来自于一些渗透调节物质合成相关的基因、转录因子、抗逆信号传导途径中的相关基因等,还有一些水通道蛋白基因、伴侣蛋白基因、aba合成途径中的相关酶基因等。例如,2007年qin等在玉米中过表达转录因子zmdreb2a显著提高了玉米的抵抗干旱和高温的能力;2010年zhang等将盐芥中的转录因子tscbf1转入玉米中从而提高了玉米的抗旱性;将来自细菌的渗透调节物质甜菜碱合成途径中的酶gsmt2、dmt2、beta转入玉米中提高了玉米的抗旱性;将来自大麦的lea蛋白家族的hav1基因导入玉米显著提高了玉米的抗旱性;zmcipk2作为响应逆境的信号途径中的一个蛋白激酶,在玉米中过表达后能明显改善玉米的抗旱性;在玉米中异源表达拟南芥los5基因,可以显著提高玉米中的aba含量和抗旱性。
5.研究报道表明,盐胁迫信号途径主要可以分为渗透和离子平衡信号途径、细胞损伤与修复以及生长调节过程。目前为止,许多参与渗透保护物质合成、离子平衡、活性氧清除以及转录因子等过程的关键耐逆基因已被克隆,并且这些基因都不同程度的提高了转基因植物的耐逆性。例如,将合成甜菜碱的cmo(胆碱单氧化酶)基因转入烟草后,可以显著提高烟草的抗盐性(wu等,2010);过表达拟南芥atsos1显著提高了转基因植物的耐盐性(qiu et al.,2002;shi et al.,2003);过表达拟南芥液泡膜na
/h
反向转运蛋白(atnhx1)基因可以显著提高转基因拟南芥、番茄的抗盐性(zhang and blumwald,2001;zhang et al.,2001);过表达拟南芥anac019、anac055和anac072/rd26能显著提高转基因植物的抗盐性;在棉花中过量表达拟南芥h
‑
ppase基因atavp1增加了转基因棉花的耐盐性和抗旱能力
(pasapula et al.,2011);转tsvp基因玉米可以显著提高玉米的抗旱和抗盐性(wei等,2008)。上述研究结果表明,虽然已经有许多植物耐盐基因被克隆和表达,但是仍主要集中在拟南芥、烟草和番茄等模式植物中,在作物中并未广泛应用。
6.已报道的一些转基因抗逆玉米大都是转单个功能基因,虽然提高了玉米的某些抗逆性,但是在正常生长环境下极少或并未有同时提高玉米产量的报道。因此,筛选既能增强作物抗逆性又能在正常条件下和胁迫条件下都能保持高产的基因成为焦点。
7.γ
‑
聚谷氨酸(γ
‑
poly glutamic acid,γ
‑
pga)是一种微生物发酵产物,在纳豆中首次被发现。γ
‑
pga由l
‑
谷氨酸和d
‑
谷氨酸通过脱水缩合而成的,其分子呈直链状,含有大量酰胺键和可游离的羧基,是一种阴离子聚合物,成品为白色无味的粉末状固体。由于γ
‑
pga具有良好的延展性、柔韧性、生物相容性、黏着性、稳定性、保湿性、吸水性、阻氧性、成膜性及生物可降解性等特点,是一种极具开发潜力的生物可降解高分子材料,广泛应用于医药、食品、化妆品、饲料、农业、环保等多个领域,是一种公认的极具发展潜力的绿色化学产品。
8.在农业领域,大量的研究发现,γ
‑
pga可以作为农药和肥料的增效剂,在低养分条件下促进作物生长,有利于提高种子活力和发芽率,促进胚芽和胚根生长发育;γ
‑
pga还可以改善土壤的理化和吸附特性,从而促进植物对养分的吸收;由于γ
‑
pga具有较强的吸附特性,可以作为吸附剂或螯合剂,对土壤中的有毒重金属离子具有明显的螯合效果,避免作物从土壤中吸收过多的有毒重金属;γ
‑
pga还可以提高植物根系的生物量,从而增强植物对氮、磷、钾的吸收,促进植物的生长。此外,γ
‑
pga作为一种保水剂可以显著提高作物的抗旱性。γ
‑
pga因其分子中含有大量的亲水基团,具有很强的吸水性与保水性,决定了其在农业节水领域的重要价值。
9.作为一种新型的绿色高分子生物材料,虽然γ
‑
pga具有广泛的应用前景,但是由于利用微生物发酵生产γ
‑
pga相对成本较高,也限制了其在农业生产中的大规模应用。同时,作为一种高分子生物材料,其长期在土壤中使用的生态、环境效应也缺乏系统、全面的研究与评估。
10.基于现有技术,申请人克隆了来自于微生物的、合成γ
‑
聚谷氨酸过程中的关键基因,利用合成生物学和基因工程技术,以玉米为实施案例转入植物中,评估异源γ
‑
聚谷氨酸合成基因在玉米中的表达以及对玉米抗旱性、耐盐性、产量的影响及对其他性状的改良效果,并建立了一种γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法,获得的产γ
‑
聚谷氨酸的转基因玉米不仅显著的提高了玉米的抗旱性,同时也显著提高了玉米的抗盐性,并且在干旱和正常生长条件下还可以显著提高玉米的生物量。本发明还对转基因玉米籽粒的品质进行了测定,发现在玉米中合成γ
‑
pga还可以提高玉米籽粒中的淀粉含量和直链淀粉含量。经检索,目前尚未有在作物体内异源合成γ
‑
聚谷氨酸并研究其对作物抗旱性、耐盐性、产量、品质影响的报道。
技术实现要素:
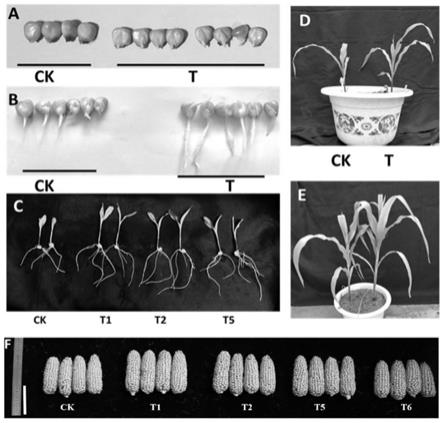
11.针对现有技术的不足,本发明要解决的问题是提供一种γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法。
12.本发明所述的γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法,步
骤是:
13.(1)克隆合成γ
‑
聚谷氨酸的基因:从产γ
‑
聚谷氨酸的菌株中克隆合成γ
‑
pga的3个关键酶基因pgsa、pgsb、pgsc;其中,基因pgsa的genbank id:aia08848.1,其氨基酸序列如seq id no.1所示;基因pgsb的genebank id:aia08846.1,其氨基酸序列如seq id no.2所示;基因pgsc的genebank id:aia08847.1,其氨基酸序列如seq id no.3所示;
14.(2)密码子优化:按照pgsa、pgsb、pgsc基因的氨基酸序列,通过玉米或待转基因的其他植物中的密码子偏好性分析并结合cpg dinucleotides content、gc content、mrna secondary structure、cryptic splicing sites、premature polya sites、internal chi sites and ribosomal binding sites、negative cpg islands、rna instability motif(are)、repeat sequences(direct repeat,reverse repeat,and dyad repeat)、restriction sites that may interfere with cloning等分析最终确定优化的序列,并根据序列人工从头合成优化后的pgsa、pgsb、pgsc基因;
15.(3)构建植物表达载体:将密码子优化后的pgsa、pgsb、pgsc基因连入含有bar基因筛选标记的植物表达载体pu130
‑
bar,获得含有目的基因的植物表达载体,命名为植物表达载体pga001;
16.(4)植物抗逆性和产量提高的转基因植物的获得;
17.其特征在于:
18.步骤(1)所述产γ
‑
聚谷氨酸的菌株是地衣芽孢杆菌(bacillus licheniformis)或解淀粉芽孢杆菌(bacillus amyloliquefaciens);
19.步骤(2)所述密码子优化后的pgsa基因核苷酸序列如seq id no.4所示;所述密码子优化后的pgsb基因核苷酸序列如seq id no.5所示;所述密码子优化后的pgsc基因核苷酸序列如seq id no.6所示;
20.步骤(3)所述植物表达载体pga001的核苷酸序列如seq id no.7所示;该表达载体各功能元件描述为:
21.[0022][0023]
步骤(4)所述植物抗逆性和产量提高的转基因植物是转基因玉米,获得该转基因玉米的方法是:
[0024]
1)含植物表达载体pga001的农杆菌菌株制备
[0025]
选择eh105农杆菌为转化菌株,在50μl该农杆菌感受态细胞中加入含pga001表达载体的质粒3μl,冰浴30分钟,液氮速冻5分钟,37℃水浴5分钟;再加入800
‑
1000μl yep液体培养基,25
‑
28℃、180
‑
250rpm振荡培养3小时;取出菌液涂布于含利福平及卡那霉素的固体yep培养基上,置25
‑
28℃黑暗倒置培养3
‑
4天,取菌落进行菌落pcr验证,并进行测序,对测序正确的农杆菌保存,用于转化;
[0026]
2)愈伤组织的准备
[0027]
选取玉米自交系kn5585作为受体自交系,取其授粉后10
‑
12d的果穗,去苞叶,在无菌工作台中,先用70%酒精处理5
‑
6min,再用无菌水冲洗4
‑
5遍,剥取1.5
‑
2mm的幼胚,盾片向上放置在幼胚诱导培养基上,28℃、黑暗培养诱导愈伤组织;2
‑
3周后从诱导出的愈伤组织中,选取生长速度较快、质地松软、松散易碎、颜色鲜艳的胚性愈伤组织转移到继代培养基上继代培养,每2周继代一次,用于农杆菌侵染;
[0028]
3)农杆菌侵染
[0029]
挑取含有pga001表达载体的农杆菌单菌落,加入5
‑
6ml的yep(kan)培养基中,培养过夜;室温下,5000
‑
6000rpm,离心5
‑
10min,收集菌体;将菌体悬浮于含有乙酰丁香酮(as)的侵染液中,并使od
600
=0.6
‑
0.8,混匀,待用;将制得的侵染液在25
‑
28℃,摇床180rpm活化
1
‑
2小时,用于侵染;将玉米的胚性愈伤组织收集在一起放入一个无菌的三角瓶中,然后倒入侵染液,将大的愈伤组织块打散摇匀,使愈伤组织充分接触农杆菌,侵染15
‑
20min;将愈伤组织取出,在滤纸上吸干多余的菌液,然后接种到共培养培养基上,19
‑
22℃黑暗培养3天;
[0030]
4)恢复培养
[0031]
使用含有抗生素的无菌水冲洗愈伤组织表面3
‑
5次,待加入的水不再浑浊时倒掉液体,将愈伤组织转入铺有滤纸的平皿中,在超净台中吹干愈伤表面水分,转入恢复培养基,28℃暗培养7
‑
10天;
[0032]
5)筛选培养
[0033]
恢复培养后将转化愈伤组织转入添加有草铵膦的筛选培养基上,筛选培养两周,然后更换新的筛选培养基,25
‑
28℃暗培养15
‑
20天,将愈伤组织打散再转入新的筛选培养基继续筛选,总共筛选2轮,培养30
‑
40天;
[0034]
6)分化生根
[0035]
将抗性愈伤组织转入分化培养基,25
‑
28℃暗培养7
‑
10天,然后转入光照培养箱25
‑
28℃培养,直至再生芽长至3
‑
5cm时转入生根培养基;
[0036]
7)炼苗移栽后自交收种
[0037]
待长出大量健壮根后,炼苗2
‑
3天,洗净根部培养基后移栽在灭菌的营养土中,室内炼苗7天后移栽到大田;期间取其幼叶进行pat/bar蛋白快速检测试纸条检测,转基因阳性玉米自交收获种子;获得t1代转基因玉米植株后,经过两代严格自交,最终获得转基因玉米纯合株系。
[0038]
上述转基因玉米方法涉及的培养基如下:
[0039][0040][0041]
本发明还提供了一种能使γ
‑
聚谷氨酸在植物中异源合成并提高植物抗逆性和产量的植物表达载体,其特征在于:所述植物表达载体命名为植物表达载体pga001,其核苷酸序列如seq id no.7所示。
[0042]
本发明还公开了利用γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法及获得的植物转基因品系。
[0043]
其中:所述植物转基因品系优选是转基因玉米纯合株系;所述抗逆性是指抗旱、抗盐性。
[0044]
(5)转基因玉米的检测:
[0045]
检测方法包括pat/bar蛋白快速检测试纸条(artron,详细步骤见说明书)检测、pcr检测、rt
‑
pcr检测,产物γ
‑
pga含量的检测。其中,γ
‑
pga含量的检测参考ny/t 3039
‑
2016方法进行。
[0046]
将玉米植株在60℃真空烘干后,研碎,称取10g混合均匀的试样,分别测定试样中经盐酸水解后的和未经水解的游离谷氨酸的含量,两者之差即为聚谷氨酸的含量。
[0047]
结果显示pgsa、pgsb、pgsc三个基因已经成功转入到玉米中(图1),并成功在玉米中检测到了γ
‑
pga(表1)。
[0048]
(6)转基因玉米表型鉴定:
[0049]
选择转基因阳性玉米株系对其整个发育期的表型进行鉴定,并进行产量测定,并对其种子品质(淀粉含量和直链淀粉含量)进行测定。
[0050]
结果显示,转基因阳性玉米株系具有更好的萌发率,更高的株高,更发达的根系,说明在玉米中合成γ
‑
pga可以促进玉米的生长,同时可以提高正常条件下玉米的产量,主要是提高了穗粒数、穗粒重、百粒重(表3),还可以提高玉米籽粒的品质,主要是淀粉含量和直链淀粉含量(表4)。
[0051]
(7)转基因玉米萌发期、苗期抗旱性和抗盐性的检测:
[0052]
转基因玉米的抗旱性检测:对转基因阳性玉米和阴性对照玉米分别进行干旱胁迫处理,并对其表型进行研究分析,干旱处理分peg模拟干旱处理和断水干旱胁迫处理。
[0053]
peg模拟干旱处理主要选萌发期和三叶期这两个时期,玉米培养液选择hogland营养液(海博生物),萌发期干旱胁迫处理选择的peg6000的浓度分别为14%、18%。苗期模拟干旱处理选择的peg6000的浓度为18%;苗期干旱处理采用盆栽断水处理,并测定游离脯氨酸、可溶性糖、叶绿素和aba含量等生理生化指标。
[0054]
在苗期抗性检测中,首先将玉米种子表面消毒后萌发,再将萌芽小苗插入到营养液(hogland玉米营养液)中培养,待植株长至三叶期时分别转移到含有200mmol/l nacl或18%peg的营养液中进行处理,观察表型变化,并进行生物量测定。
[0055]
干旱试验:选取大小均匀的转基因玉米t3代种子和野生型kn5585种子播种于大小一致塑料小花盆中,盘中装有等量的质地均匀的肥沃土壤。每盘一半播种转基因材料,一半播种野生型材料,各2粒,每株系种3盘。正常浇水至小苗长至3叶期,然后进行干旱胁迫处理,即一次性浇足水分后停止浇水,一直到野生型植株死亡后再恢复浇水,观察转基因株系在干旱条件下的生长状况及恢复浇水后的成活率和恢复速率。田间干旱处理(实施地点为三亚南繁基地)选择开花前断水处理的方法,具体实施办法为:断水15天,复水一次,再断水15天,一直到收获,对抗逆表型和产量进行测定。
[0056]
结果表明,转基因阳性玉米在peg处理下仍可以较好的萌发,而阴性对照玉米的萌发却受到了明显的抑制(图3)。大田干旱断水试验结果表明,在大田干旱条件下,转基因阳性玉米也表现出良好的抗旱性。
[0057]
转基因玉米抗盐性研究包括萌发期和苗期:对萌发期玉米选择不同浓度的nacl溶液进行盐胁迫处理。选取转基因阳性材料及阴性对照材料的种子,表面消毒后分别在含150mm nacl、200mm nacl溶液的滤纸上萌发,并观察萌发情况;苗期抗盐试验采取溶液胁迫处理的方法:将玉米种子表面消毒使其萌发后转移至营养液(hogland玉米营养液)中培养,将培养至三叶期的玉米分别移至含200mm nacl的营养液中培养,并统计表型变化。
[0058]
结果表明,无论在较低浓度还是高浓度nacl胁迫条件下,转基因玉米种子的萌发情况都比对照阴性玉米好(图7
‑
图8),说明转基因玉米在萌发期具有更好的抗盐性,苗期水培抗盐试验结果也显示转基因阳性玉米具有明显的抗盐性。
[0059]
(8)转基因玉米在正常条件下和干旱条件下产量分析
[0060]
在田间条件下进行了产量分析,分正常灌溉条件和干旱断水条件。正常灌溉条件是保持转基因玉米和对照自交系在正常自然生长条件下生长;干旱试验主要采取开花前断水处理的方法,具体实施办法为:断水15天,复水一次,再断水15天,一直到收获,对产量和产量相关性状进行测定(果穗形态、穗长、穗重、穗粒数、穗粒重、百粒重等)。
[0061]
结果显示,在干旱条件下,转基因阳性玉米的产量显著提高。与正常灌溉条件下产量相比,转基因玉米受干旱的影响较小,而阴性对照玉米在干旱条件下产量显著下降。
[0062]
本发明公开了一种γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法,同时公开了利用γ
‑
聚谷氨酸在植物中异源合成提高植物抗逆性和产量的方法所获得的植物转基因品系,优选是转基因玉米纯合株系。实验证实:获得的产γ
‑
聚谷氨酸的转基因玉米不仅显著的提高了玉米的抗旱性,同时也显著提高了玉米的抗盐性,并且在正常生长条件下还显著提高了玉米的生物量。本发明的有益效果是:(1)创造性的实现了在植物中合成γ
‑
聚谷氨酸,并证明在获得的转基因玉米中可以产生γ
‑
聚谷氨酸。(2)评估异源γ
‑
聚谷氨酸合成基因在玉米中的表达以及对玉米抗旱性、耐盐性、产量的影响及对其他性状的改良效果显示,通过本发明方法得到的转基因植物表现出明显的抗旱性、耐盐性和对产量的提升作用,证实本发明方法是一种绿色、高效、可持续的解决作物在干旱、缺水、盐碱胁迫下稳产或少减产的有效途径,可广泛应用于植物、作物的抗旱、耐盐分子育种与新品种培育,具有广阔的应用前景。
附图说明
[0063]
图1示植物表达载体图谱示意图和转基因玉米的检测。
[0064]
其中,a:植物表达载体pga001图谱示意图;b:转基因玉米的pat/bar蛋白快速检测,ck:阴性对照自交系kn5585;1
‑
9:转基因阳性玉米株系;c:转基因玉米bar基因的pcr检测结果;m:dna marker;ck:阴性对照自交系kn5585;1
‑
9、12、13、15:转基因阳性玉米株系;d:转基因玉米中pgsa、pgsb、pgsc基因的rt
‑
pcr检测结果;t1、t2、t5:转基因阳性玉米株系。
[0065]
图2示转基因阳性玉米与阴性对照玉米自交系在正常生长条件下的表型。
[0066]
其中,a:转基因阳性玉米与阴性对照玉米自交系的在正常条件下萌发30h后的情况;其中ck:阴性对照自交系kn5585,t:转基因阳性玉米株系;b:转基因阳性玉米与阴性对照玉米自交系在正常条件下萌发48h后的情况;其中ck:阴性对照自交系kn5585,t:转基因阳性玉米株系;c:转基因阳性玉米和阴性对照玉米的根系发育情况;其中ck:阴性对照自交系kn5585,t1、t2、t5:转基因阳性玉米株系;d:转基因阳性玉米和阴性对照玉米盆栽条件下
苗期的长势情况;其中ck:阴性对照自交系kn5585,t:转基因阳性玉米株系;e:转基因阳性玉米和阴性对照玉米盆栽条件下苗期的长势情况;f:转基因阳性玉米与阴性对照玉米的果实表型;其中ck:阴性对照自交系kn5585,t1、t2、t5、t6:转基因阳性玉米株系。
[0067]
图3示转基因玉米和野生型对照玉米自交系萌发期抗旱性检测。
[0068]
图4示转基因阳性植株和阴性对照玉米peg模拟干旱试验。
[0069]
其中,a:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的及未处理的阴性对照玉米peg模拟干旱对比试验;b:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的及未处理的阴性对照玉米在正常条件下和peg处理条件下地上部分的鲜重对比;c:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的及未处理的阴性对照玉米在正常条件下和peg处理条件下地下部分的鲜重对比。
[0070]
图5示转基因玉米的盆栽干旱断水试验。
[0071]
其中,a:转基因阳性植株与阴性对照自交系bar基因试纸条检测结果;b:转基因阳性玉米盆栽干旱断水试验结果。
[0072]
图6示转基因阳性玉米在田间条件下的干旱断水试验及产量分析。
[0073]
其中,a:转基因阳性玉米及阴性对照玉米苗期断水8d后的表型;b:转基因阳性玉米及阴性对照玉米在开花前干旱断水8d后的表型;c:干旱条件下的转基因阳性玉米和阴性对照玉米的产量分析;其中ck:阴性对照自交系kn5585;t1、t2、t5、t6、t8、t9、t12、t13:转基因阳性玉米株系。
[0074]
图7示转基因玉米和野生型对照玉米自交系萌发期抗盐性检测。
[0075]
图8示转基因阳性株系和阴性对照植株的苗期抗盐试验。
[0076]
其中,ck:阴性对照自交系kn5585;t1、t2、t5:转基因阳性自交系kn5585。
[0077]
图9示转基因阳性(内源合成γ
‑
pga)玉米与γ
‑
pga外源处理的野生型对照玉米抗旱和抗盐性对比试验。
[0078]
其中,a:转基因阳性(内源合成γ
‑
pga)玉米与γ
‑
pga外源处理的野生型对照玉米抗盐性对比试验;b:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的野生型对照玉米以及野生型对照玉米在正常条件下和盐胁迫处理条件下地上部分的鲜重对比;c:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的野生型对照玉米以及野生型对照玉米在正常条件下和盐胁迫处理条件下地下部分的鲜重对比;其中ck:阴性对照自交系kn5585;t1、t2:转基因阳性自交系。
[0079]
图10为植物表达载体pu130
‑
bar图谱示意图。
具体实施方式
[0080]
下面结合具体附图和实施例对本发明内容进行详细说明。如下所述例子仅是本发明的较佳实施方式,应该说明的是,下述说明仅仅是为了解释本发明,并非对本发明作任何形式上的限制,凡是依据本发明的技术实质对实施方式所做的任何简单修改、等同变化与修饰,均属于本发明技术方案的范围内。
[0081]
下述实施例中,所使用的材料、试剂等,如无特殊说明,均从商业途径得到。
[0082]
实施例1:pgsa、pgsb、pgsc氨基酸序列的获得
[0083]
从产γ
‑
聚谷氨酸的菌株地衣芽孢杆菌(bacillus licheniformis)中以公知的方
法克隆合成γ
‑
pga的3个关键酶基因pgsa、pgsb、pgsc;其中,基因pgsa的genbank id:aia08848.1,其氨基酸序列如seq id no.1所示;基因pgsb的genebank id:aia08846.1,其氨基酸序列如seq id no.2所示;基因pgsc的genebank id:aia08847.1,其氨基酸序列如seq id no.3所示。
[0084]
实施例2:密码子优化
[0085]
按照已获得的pgsa、pgsb、pgsc基因的氨基酸序列,通过玉米或待转基因的其他植物中的密码子偏好性分析并结合cpg dinucleotides content、gc content、mrna secondary structure、cryptic splicing sites、premature polya sites、internal chi sites and ribosomal binding sites、negative cpg islands、rna instability motif(are)、repeat sequences(direct repeat,reverse repeat,and dyad repeat)、restriction sites that may interfere with cloning等分析最终确定优化的序列,并根据序列人工从头合成优化后的pgsa、pgsb、pgsc基因;其中,所述密码子优化后的pgsa基因核苷酸序列如seq id no.4所示;所述密码子优化后的pgsb基因核苷酸序列如seq id no.5所示;所述密码子优化后的pgsc基因核苷酸序列如seq id no.6所示。
[0086]
实施例3:植物表达载体构建
[0087]
将密码子优化后的pgsa、pgsb、pgsc基因以公知的方法连入含有bar基因筛选标记的植物表达载体pu130
‑
bar(载体图谱如图10所示),获得含有目的基因的植物表达载体,命名为植物表达载体pga001,载体图谱如图1所示;其中所述植物表达载体pga001的核苷酸序列如seq id no.7所示;该表达载体各功能元件描述为:
[0088][0089][0090]
上述植物表达载体pga001构建采用多片段重组酶方法构建,具体构建步骤是:
[0091]
用限制性内切酶hindlll和spel酶切1μg载体pu130
‑
bar,37℃下反应1
±
0.2小时,使用dc301试剂盒(市售pcr凝胶回收试剂盒)回收酶切载体产物。
[0092]
实施例2中密码子优化后的pgsa、pgsb、pgsc的pcr产物35s pgsa tnos,35s pgsb tnos,35s pgsc tnos,直接由生物公司合成。
[0093]
然后进行重组。重组连接体系如下:合成pcr产物pgsa pgsb pgsc分别为3μl;酶切载体产物,3μl;重组酶,2μl;重组buffer,4μl;h2o,2μl。37℃30min后冰上或者4℃保存;
[0094]
然后转化大肠杆菌dh5α:10μl以上连接产物 100μl大肠杆菌感受态。
[0095]
感受态dh5α冰箱取出后,迅速放入冰上,5分钟后待菌块溶解加入连接产物,冰上静置25min,42℃热击45s,冰上放置2min(勿晃动),加入无抗生素的lb 100μl,37℃,200rpm,摇菌1h,涂板,lb k
,37℃培养一天。
[0096]
然后进行菌落pcr,检测引物是:
[0097]
qc
‑
f:gcggccgagcccctcatcgagaa
[0098]
qc
‑
r:gatcaggcccggcacgatgatg
[0099]
取阳性菌落进行测序。测序结果正确的质粒用于转化农杆菌。
[0100]
实施例4:产γ
‑
pga转基因玉米的获得
[0101]
(1)玉米胚性愈伤组织的获得
[0102]
选取玉米自交系kn5585作为受体自交系,取其授粉后10
‑
12d的果穗,去苞叶,在无菌工作台中,先用70%酒精处理5min,再用无菌水冲洗4
‑
5遍,剥取1.5
‑
2mm的幼胚,盾片向上放置在诱导培养基(ms 1mg/l2,4
‑
d 1.38g/l l
‑
脯氨酸 0.5g/l水解酪蛋白 30g/l蔗糖 7g/l琼脂,ph5.8)上,28℃、黑暗培养诱导愈伤组织。2
‑
3周后从诱导出的愈伤组织中,选取生长速度较快、质地松软、松散易碎、颜色鲜艳的胚性愈伤组织转移到继代培养基上(ms 1mg/l2,4
‑
d 0.69g/l l
‑
脯氨酸 0.5g/l水解酪蛋白 30g/l蔗糖 7g/l琼脂,ph5.8)继代培养,每2周继代一次,用于农杆菌侵染。
[0103]
(2)含植物表达载体的农杆菌的制备
[0104]
选择eh105农杆菌为转化菌株,在50μl eha105商业化农杆菌感受态细胞(博迈德)中加入含pga001表达载体的质粒3μl,冰浴30分钟,液氮速冻5分钟,37℃水浴5分钟;再加入800μlyep液体培养基,28℃、250rpm振荡培养3小时;取出菌液涂布于含利福平及卡那霉素的固体yep培养基上,置28℃黑暗倒置培养3
‑
4天,取菌落进行菌落pcr验证,并进行测序,对测序正确的农杆菌保存,用于转化;
[0105]
(3)农杆菌侵染与共培养
[0106]
挑取含有pga001表达载体的农杆菌单菌落,加入5ml的yep(kan)培养基中,培养过夜;室温下,6000rpm,离心5min,收集菌体;将菌体悬浮于含有100mm乙酰丁香酮(as)的侵染液(ms 1mg/l2,4
‑
d 68.5g/l蔗糖 36g/l葡萄糖 100mm as,ph5.2)中,并使od
600
=0.6
‑
0.8,混匀,待用;将制得的侵染液在25℃摇床,180rpm活化1小时,用于侵染;将玉米的胚性愈伤组织收集在一起放入一个无菌的三角瓶中,将侵染液倒入,将大的愈伤组织块打散,摇匀使得愈伤组织充分接触农杆菌,侵染15min;将愈伤组织取出,在滤纸上吸干多余的菌液,然后接种到共培养培养基(ms 1mg/l 2,4
‑
d 100mm as 0.5g/l mes 20g/l蔗糖 7.5g/l琼脂,ph5.2)上,22℃黑暗培养3天。
[0107]
(4)恢复培养与抗性愈伤组织的筛选
[0108]
使用含有抗生素的无菌水冲洗愈伤组织表面3
‑
5次,待加入的水不再浑浊时倒掉液体,将愈伤组织转入铺有滤纸的平皿中,在超净台中吹干愈伤表面水分。转入恢复培养基(ms 1mg/l2,4
‑
d 0.69g/l l
‑
脯氨酸 0.5g/l水解酪蛋白 20g/l蔗糖 250mg/l头孢霉素 7.5g/l琼脂,ph5.8),28℃暗培养7
‑
10天。恢复培养后将转化愈伤组织转入添加有草铵膦(ppt)的筛选培养基(ms 1mg/l2,4
‑
d 0.69g/l l
‑
脯氨酸 0.5g/l水解酪蛋白 20g/l蔗糖 250mg/l头孢霉素 10mg/l ppt 7.5g/l琼脂,ph5.8)上,开始筛选培养两周,然后更换新的
筛选培养基,25
‑
28℃暗培养15天,将愈伤组织打散转入新的筛选培养基继续筛选,总共筛选2轮。
[0109]
(5)抗性愈伤组织的分化和生根
[0110]
将抗性愈伤组织转入分化培养基(ms 0.5mg/l6
‑
ba 0.5g/lmes 10mg/lppt 250
‑
300mg/l的头孢霉素 20g/l蔗糖 7g/l琼脂,ph5.8),25
‑
28℃暗培养7
‑
10天,然后转入光照培养箱25
‑
28℃培养,直至再生芽长至3
‑
5cm时转入生根培养基(ms 20g/l蔗糖 7g/l琼脂,ph5.8)。待长出大量健壮根后,炼苗2
‑
3天,洗净根部培养基后移栽在灭菌的营养土中,室内炼苗7天后移栽到大田(炼苗期间用薄膜覆盖花盆以提高湿度)。
[0111]
(6)取移栽成活的转基因幼苗的幼叶进行pat/bar蛋白快速检测试纸条(artron)检测,转基因阳性玉米自交收获种子。获得t1代转基因玉米植株后,经过两代严格自交,最终获得转基因玉米纯合株系。
[0112]
实施例5:转基因玉米的检测
[0113]
转基因检测方法包括pat/bar蛋白快速检测试纸条检测、pcr检测、rt
‑
pcr检测、产物γ
‑
pga含量的检测。
[0114]
pat/bar蛋白快速检测试纸条(artron)检测参照试剂盒说明书进行;pcr、rt
‑
pcr检测参照(解光宁.玉米泛素受体zmda1,zmdar1对籽粒发育的影响[d].山东大学,2017。)所述方法。产物γ
‑
pga含量的检测参考ny/t 3039
‑
2016方法进行。
[0115]
将玉米植株在60℃真空烘干后,研碎,称取10g混合均匀的试样,分别测定试样中经盐酸水解后的和未经水解的游离谷氨酸的含量,两者之差即为聚谷氨酸的含量。
[0116]
试验结果见图1和表1
[0117]
表1、转基因玉米材料中γ
‑
pga的含量(mg/100g干重)
[0118][0119]
由上述结果可以看出pgsa、pgsb、pgsc三个基因已经成功转入到玉米中(图1),并在转基因玉米中成功检测到了γ
‑
pga。
[0120]
实施例6:转基因玉米表型鉴定
[0121]
选择转基因阳性玉米株系对其整个发育期(萌发期、苗期、拔节期等)以及种子的表型进行鉴定。
[0122]
试验结果见图2、表3,对正常生长条件下的转基因阳性玉米和阴性对照玉米的表型进行观察,确定在玉米中异源合成γ
‑
pga对玉米生长发育的影响,结果如图2所示。
[0123]
结果显示,转基因阳性玉米株系具有更好的萌发率,更高的株高,更发达的根系,说明在玉米中合成γ
‑
pga可以促进玉米的生长;发明人还对其在正常灌溉条件下的产量进行评估,发现在玉米中合成γ
‑
pga可以提高正常生长条件下玉米的产量,主要是提高了穗粒数、穗粒重、百粒重(表3)。
[0124]
实施例7转基因玉米抗逆性鉴定
[0125]
转基因玉米的抗旱性检测
[0126]
对转基因阳性玉米和阴性对照玉米分别进行干旱胁迫处理,并对其表型进行研究。干旱处理分peg模拟干旱处理和断水干旱胁迫处理。
[0127]
peg模拟干旱处理主要选萌发期和三叶期这两个时期,玉米培养液选择hogland营养液(海博生物),萌发期干旱胁迫处理选择的peg6000的浓度分别为14%、18%。选取转基因材料及对照材料的种子表面消毒,分别在含0mmol/l、12%peg、14%peg溶液的滤纸上萌发,统计萌发率,观察生长势。苗期模拟干旱处理选择的peg6000的浓度为18%;苗期干旱处理采用盆栽断水处理,并测定游离脯氨酸、可溶性糖、叶绿素含量和aba 含量等生理生化指标。田间干旱处理实施地点为三亚南繁基地,选择开花前断水处理的方法,具体实施办法为:断水15天,复水一次,再断水15天,一直到收获,对抗逆表型和产量进行测定。
[0128]
游离脯氨酸、可溶性糖、叶绿素、aba含量的测定方法参考(wang b,li z,ran q,et al.zmnf
‑
yb16 overexpression improves drought resistance and yield by enhancing photosynthesis and the antioxidant capacity of maize plants[j].frontiers in plant science,2018,9:709;li z,liu c,zhang y,et al.the bhlh family member zmptf1 regulates drought tolerance in maize by promoting root development and aba synthesis[j].journal of experimental botany,2019(19):19.)进行。
[0129]
抗旱性分析结果见图3
‑
图5。
[0130]
结果表明,转基因阳性玉米在peg处理下仍可以较好的萌发,而阴性对照玉米的萌发却受到了明显的抑制(图3)。在苗期的peg模拟干旱处理试验中,发明人将转基因阴性对照玉米一部分外源施加50mg/l的γ
‑
pga,另一部分作为对照,与转基因阳性玉米同时在18%peg溶液处理下进行培养,结果显示,在18%peg溶液处理条件下,阴性对照玉米的生长明显受到了抑制,而转基因阳性玉米和外源施加γ
‑
pga的处理组的生长状况明显优于对照材料(图4a)。发明人对18%peg处理5d后的转基因阳性玉米株系、外源施加γ
‑
pga的阴性玉米植株以及阴性对照株系进行了地上部分和地下部分的鲜重统计,发现转基因阳性玉米株系在peg处理条件下的生物量明显高于阴性对照玉米,甚至略高于外源施加γ
‑
pga的玉米组(图4b
‑
4c)。说明γ
‑
pga在玉米中的异源合成可以达到外源施加γ
‑
pga提高玉米抗旱性的效果。这表明,转基因玉米的抗逆性提高是因为其产生了γ
‑
pga。
[0131]
发明人还对三叶期的转基因阳性玉米和阴性对照玉米进行了盆栽断水处理试验。发现在干旱胁迫早期条件下,转基因阳性玉米相对转基因阴性对照玉米表现出较好的生长状态。经历8天的断水处理后,阴性对照玉米几乎萎蔫致死,恢复浇水,转基因阳性玉米仍可以迅速恢复生长,而对照玉米几乎全部死亡(图5)。对断水处理6天的叶片的aba、可溶性糖、脯氨酸和叶绿素含量进行测定,结果显示,在干旱胁迫条件下,在玉米中异源合成γ
‑
pga可以显著提高玉米叶片中aba、可溶性糖、脯氨酸和叶绿素的含量,从而提高玉米的抗旱性(表2)。
[0132]
表2、干旱处理对转基因阳性玉米和阴性对照玉米中aba、可溶性糖(soluble sugar)、脯氨酸(proline)、叶绿素(chlorophyll)含量的影响
[0133][0134]
大田干旱断水试验结果见图6。结果表明,在大田干旱条件下,转基因阳性玉米也表现出良好的抗旱性。
[0135]
转基因玉米抗盐性研究包括萌发期和苗期:对萌发期玉米选择不同浓度的nacl溶液进行盐胁迫处理。选取转基因阳性材料及阴性对照材料的种子表面消毒后,分别在含150mm nacl、200mm nacl溶液的滤纸上萌发,并观察萌发情况;苗期抗盐试验采取溶液胁迫处理:将玉米种子表面消毒使其萌发后转移至营养液(hogland玉米营养液)中培养,将培养至三叶期的玉米分别移至含200mm nacl的营养液中培养,并统计表型变化。
[0136]
抗盐性分析结果见图7
‑
图8。结果表明,无论在较低浓度还是高浓度nacl条件下,转基因玉米种子的萌发情况都比对照阴性玉米好,说明转基因玉米在萌发期具有更好的抗盐性,苗期水培抗盐试验结果也显示转基因阳性玉米具有明显的抗盐性。
[0137]
转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的野生型对照玉米抗盐和抗旱性对比:转基因阳性(内源合成γ
‑
pga)玉米、γ
‑
pga外源处理的野生型自交系抗旱和抗盐性对比试验采取溶液胁迫处理的方法。首先将玉米种子表面消毒后萌发,再将萌芽小苗插入到营养液(hogland玉米营养液)中培养,待植株长至三叶期时,开始进行处理,转基因玉米株系分别转移到含有18%peg或200mmol/lnacl的营养液中进行处理,对照玉米自交系分别转移至含有20mg/lγ
‑
pga、18%peg、18%peg 20mg/lγ
‑
pga、200mmol/l nacl、200mmol/lnacl 20mg/lγ
‑
pga的营养液中进行处理,未经任何处理的野生型玉米和转基因阳性玉米株系作为对照,期间观察表型变化,最后进行生物量测定。
[0138]
结果见图9。结果表明,转基因玉米株系与外源γ
‑
pga处理的野生型玉米的抗旱性和抗盐性相当,甚至比外源添加γ
‑
pga的野生型玉米的抗性更好一些,说明转pgsa、pgsb、pgsc基因使其在玉米中内源合成γ
‑
pga在提高玉米抗旱性和抗盐性上具有更好的效果。
[0139]
实施例8:转基因玉米在正常条件下和干旱条件下产量分析
[0140]
对大田中种植的转基因阳性玉米植株和转基因阴性对照玉米植株在开花前进行干旱处理,具体实施办法为断水15天后复水,然后再断水15天,然后复水。最后对产量进行统计。
[0141]
结果见图2f、图6c、表3。
[0142]
结果显示,在干旱条件下,转基因阳性玉米的产量显著提高。与正常灌溉条件下产量相比,转基因玉米受干旱的影响较小,而阴性对照玉米在干旱条件下产量显著下降。
[0143]
表3、转基因阳性玉米与阴性对照玉米在正常灌溉条件下和干旱条件下的产量统计
[0144][0145]
实施例9:转基因阳性玉米与阴性对照玉米种子中淀粉和直链淀粉的含量检测
[0146]
对转基因阳性玉米与阴性对照玉米种子中的淀粉和直链淀粉的含量进行测定,方法参考(解光宁.玉米泛素受体zmda1,zmdar1对籽粒发育的影响[d].山东大学,2017。)。
[0147]
转基因阳性玉米与阴性对照玉米种子中淀粉和直链淀粉的含量检测对比发现转基因玉米可以显著提高玉米种子中淀粉和直链淀粉的含量。
[0148]
结果见表4。
[0149]
表4、转基因阳性玉米与阴性对照玉米种子中淀粉和直链淀粉含量
[0150][0151]
综上所述,本发明首次利用合成生物学原来和基因工程技术提供了一种在植物中合成γ
‑
pga的方法,且成功的在转基因玉米中检测到γ
‑
pga的合成,证明了γ
‑
pga可以在植物中异源合成;且本发明选择主要的粮食作物玉米作为供试材料,获得的转基因玉米证实可以明显提高其抗旱、抗盐性,在干旱条件下可以明显提高其产量;在非胁迫的正常灌溉条件下的产量也没有下降,甚至还略微提高;本发明还对转基因玉米籽粒的品质进行了测定,发现在玉米中合成γ
‑
pga还可以提高玉米籽粒中的淀粉含量和直链淀粉含量。上述研究结果提示,本发明方法可以广泛应用于作物抗逆分子育种、农作物生物量提高等方面,具有重要的理论意义和巨大的经济价值。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。

