1.本发明属于兽医学领域,具体涉及一种鸭坦布苏病毒抑制剂。
背景技术:
2.鸭坦布苏病毒(duck tembusu virus, dtmuv)是引起蛋鸭产蛋率骤降、肉鸭和育成鸭死亡的主要病原之一,对养鸭业造成的经济损失巨大。dtmuv所引起的鸭坦布苏病毒病在我国开始于2010年的东南沿海,近年来已成为水禽养殖业常见且频繁流行的疫病之一,也是必须监测和防控的疫病之一。目前对于该病的预防主要依靠商品化疫苗,但疫病仍频繁出现,且呈地方性流行趋势,表明dtmuv已在不同区域定植并在不同选择压力下突变产生地方流行株。鉴于病毒的突变加速和相应疫苗研制的滞后,开发能够有效抑制所有地方流行株且长期有效的生物制剂成为当务之急,但近年来相关研究罕见报导。
3.dtmuv属黄病毒科(flaviviridae)、黄病毒属(flavivirus),为有囊膜不分节段单股正链rna病毒,基因结构与其他黄病毒类似:全长约10.9kb,5’和3’端各有一非编码区,其余基因为一个开放阅读框,从5
’‑3’
依次编码c蛋白、prm/m蛋白、e蛋白、非结构蛋白ns1
‑
ns2a
‑ꢀ
ns2b
‑
ns3
‑
ns4a
‑
2k
‑
ns4b
‑
ns5。其中e蛋白是病毒表面主要结构蛋白,以反向平行的方式形成二聚体和三聚体存在于病毒颗粒表面,包含许多与保护性免疫、病毒毒力、组织嗜性、宿主细胞膜融合及宿主细胞表面受体结合相关的抗原决定簇。依据其结构,e蛋白可分为三个结构域(domain i
‑
iii或d i
‑
iii):位于中心位置的β桶状结构域i、包含融合环结构的结构域ii和包含潜在受体结合区的免疫球蛋白样(ig
‑
like)结构域iii。d iii被认为是介导黄病毒与宿主受体相结合的主要蛋白:首先,ig
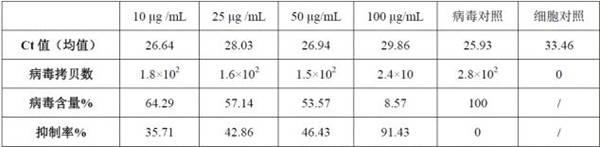
‑
like结构使其具有黏附功能;其次,该结构域独立于e蛋白的其它结构域,并垂直伸出病毒表面形成突起;再次,多项研究表明,重组的e d iii或者针对e d iii的抗体能够有效抑制病毒进入靶细胞,这也是在结构基础上判定某蛋白为病毒吸附蛋白的必要条件。
4.预防病毒感染最有效的方法就是阻止病毒颗粒进入细胞。病毒与细胞表面相应受体的结合是其生命周期中的始动环节,也是影响宿主范围、组织嗜性和致病性的决定因素之一,所以阻断病毒与细胞受体的结合是防止感染最直接有效的方法。所以,研发能够与病毒受体相结合的制剂,抑制病毒的入胞与感染,对于控制dtmuv传播与流行具有重要意义。
技术实现要素:
5.针对现有技术中缺乏对不同区域dtmuv流行株均有效果的生物制剂的问题,本发明提供了一种高效且具备长使用时效dtmuv抑制剂,通过鸭坦布苏病毒e蛋白第三结构域抑制dtmuv进入细胞。
6.为实现上述目的,本发明采用如下技术方案。
7.一种鸭坦布苏病毒e d iii蛋白的制备方法,包括以下步骤:(1)构建重组大肠杆菌,诱导表达获得e d iii蛋白包涵体;(2)将包涵体复性、纯化后获得e d iii蛋白的溶液。
b(de3),获得重组大肠杆菌,然后进行ediii的诱导表达。表达产物sds
‑
page结果显示,当温度为37℃,iptg浓度为1mm诱导培养时,ediii以包涵体形式表达,蛋白表达量可占到菌体总蛋白的35%。与cn104198736a获得的蛋白相比,本实验表达的蛋白不含有额外的标签蛋白,不会干扰蛋白的复性与活性。
19.2.ediii蛋白的高效表达将获得的重组大肠杆菌按照1%的接种量转接到含有2llb(氨苄青霉素抗性)培养基的摇瓶中,37℃200rpm培养至od
600
=0.4
‑
0.6,加入iptg至终浓度为1mm,37℃,160rpm继续培养5
‑
6h;将培养好的菌液移入300ml离心管中,4℃5000rpm离心10min,弃上清,然后用约60ml1
×
pbs悬浮菌体,超声裂解,12000rpm,离心15min,收集离心管下部呈白色致密块的包涵体;用移液器或玻棒小心的将包涵体上面一层细胞碎片去除,使用washingbuffer(含0.5%triton
‑
100,50mmtris(ph8.0),300mmnacl,10mmedta,10mmdtt)洗涤后,包涵体用resuspensionbuffer(含50mmtris(ph8.0),100mmnacl,10mmedta,10mmdtt)悬浮,12000rpm,离心15min;称取包涵体重量,按30mg/ml的比例用盐酸胍溶解,4℃搅拌溶解,12000rpm,15min,取上清分装成1ml/管保存于
‑
80℃备用。
20.实施例2ediii蛋白的复性与纯化配制refoldingbuffer(含100mmtris(ph8.0),400mml
‑
arghcl,2mmedta,5mmgsh,0.5mmgssg)于4℃预冷;将盛有500mlrefoldingbuffer的烧杯置于磁力搅拌器中,加入转子,设置合适的搅拌速度;取实施例1中1管ediii蛋白液逐滴缓慢滴入refoldingbuffer内,慢慢搅拌8小时复性;然后进行浓缩,直至体积到5ml以内,4℃,12000rpm离心10分钟,去除沉淀,取上清吸入aktafplc(快速蛋白液相色谱),在层析柱上以1ml/min运行,根据分子筛层析的结果检测复性效果(图1),收集相应分子量(11kda)的蛋白峰,进行sds
‑
page鉴定;用阴离子交换柱resourceq进一步纯化,收集洗脱峰进行sds
‑
page检测(图2)。最终获得纯度在90%以上的复性ediii蛋白。
21.实施例3ediii蛋白的结晶与结构解析将实施例2中纯化后的蛋白液经超滤浓缩管浓缩至约20mg/ml,使用hamptonresearch公司的crystalscreeni和ii、index、saltrx、peg/ion等试剂盒,利用悬滴气相扩散法于4℃
‑
18℃筛选结晶条件,3
‑
7天后,置于显微镜下观察晶体生长情况。如果观察到有晶体生长,记下相应的条件并将相应的晶体送到x光机上衍射鉴定。获得的初始结晶条件为:4℃下以hamptonresearch公司的index
tm
hr2
‑
144的59号溶液(0.02mmgcl2,0.1mhepesph7.5,22%w/vpoly(聚丙烯酸钠)5100)为结晶溶液,结晶3天即可观察到明显的晶体。对蛋白质浓度、结晶温度、结晶溶液组成和ph进行优化后,确定结晶条件为:以蛋白质溶液浓度10mg/ml在4℃下在组成为23%w/vpoly5100、0.015mmgcl2、0.1mhepesph8.0的结晶溶液中生长7天,获得针状蛋白质晶体(图3)。将蛋白质晶体进行x光衍射分析(图4a),收集数据后利用hkl2000、ccp4、coot等软件进行数据的处理和修正,获取蛋白质精细结构,如图4b所示:结构为含6个β片层的桶状ig结构。
22.实施例4ediii蛋白抑制dtmuv感染1.ediii蛋白与受体互作结构分析在黄病毒属病毒中,dtmuvediii与西尼罗河病毒(westnilevirus,wnv)、日
本乙型脑炎(japaneseencephalitisvirus,jev)的同源性最高,故结构的相似性是最高的。同时,已有研究表明,jev、wnv等e蛋白的388
‑
390位点(rgd/e)为病毒与受体相互作用的位点,此位点前后的序列相对保守。而对应于dtmuvediii,在389
‑
391区域的gkg/e与jev、wnv等e蛋白的388
‑
390区域相对应,且其前后区域的序列十分保守。故推测,gkg/e是dtmuvediii与受体相互作用的关键区域。
23.为了验证这一推测,接下来对ediii的结构进行了分析。dtmuvediii的整体结构包含6个β片层和1个α螺旋,与wnv含10个β片层和1个α螺旋的结构以及jev只含6个β片层的结构有所区别,这跟蛋白氨基酸序列组成密切相关。进一步对jev、wnv等e蛋白的388
‑
390区域(rgd/e)的构象进行了分析,该部位(红色表示)突出于蛋白表面(图5a
‑
b),利于与其他蛋白的相互作用。而在dtmuve蛋白的389
‑
391位置,位点gkg/e形成1个α
‑
helix(红色表示),且突出于蛋白表面(图5c),特征与wnvediiirgd/e区域特征一致,符合与受体结合的结构特征,故ediii包含与受体结合的潜在位点与构象。通过图5中jev、wnv两种病毒和dtmuvediii的结构对比也可以知道,三者结构存在构象以及位置上的不同,即通过wnv和jev是不能直接推定dtmuv该蛋白的功能的。
24.随后有对所有dtmuvediii的氨基酸序列进行了比对(图6),以确定受体结合潜在区域的保守性。结果表明,在已报道的dtmuvediii中,gkg/e及其临近区域十分保守,依此为基础研制的产品能够适用于不同分离毒株。
25.2.ediii蛋白对dtmuv感染的抑制vero细胞传代后铺于96孔板,待长成单层后与稀释成不同浓度的实施例2中的复性ediii蛋白液和cn104198736a中的ediii蛋白液(蛋白纯度≥90%)共孵育,浓度梯度分别为10、25、50、100μg/ml,孵育时间为4度条件下1.5小时,同时使用bsa代替ediii蛋白作为阴性对照;随后用无血清无双抗的mem冲洗细胞2次,洗去未结合的蛋白;将测定滴度的dtmuv接种各孔细胞,接种浓度为200tcid
50
,吸附1小时后用无血清无双抗的mem冲洗细胞2次,洗去未吸附的病毒;加入细胞维持液将细胞至于37度、5%co2培养箱中进行培养,于24小时(此时间点位于dtmuv在vero细胞上增长的对数期)后收集细胞,一部分提取rna进行荧光定量的测定,另一部分测定病毒的tcid
50
,明确各组病毒含量的差异和增长趋势。
26.表1ediii对dtmuv入胞的抑制作用(荧光定量法)表2cn104198736a中表达的ediii对dtmuv入胞的抑制作用(荧光定量法)
表1和图7结果表明,孵育了e d iii蛋白的接毒组,无论是荧光定量核酸检测还是细胞测定病毒滴度的结果,病毒含量较对照组均显著降低,显著抑制了dtmuv入胞的数量,所以dtmuv e d iii能够有效抑制病毒的入胞和感染。
27.通过表1和表2对比可看出,专利cn104198736a中利用pet32a/e d iii表达的e d iii虽然是活性蛋白,但其浓度越大对病毒抑制率越低,可能是由于携带的trx标签遮蔽了e d iii中与受体相互作用的位点(e蛋白的389
‑
391区域)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。

