具有遗传修饰的钠通道的啮齿动物及其使用方法
1.相关申请的交叉引用
2.本技术要求2019年2月22日提交的美国临时申请号62/808,957的优先权权益,其整个内容通过引用并入本文。
3.序列表的通过引用并入
4.于2020年2月20日创建并通过efs
‑
web递交给美国专利和商标局(united states patent and trademark office)的781kb的命名为36328pct_10403wo01_sequencelisting.txt的ascii文本文件中的序列表通过引用整体并入本文。
背景技术:
5.电压门控通道α亚基9(scn9a)是编码nav1.7蛋白的基因。nav1.7是电压门控钠通道家族的一个成员,并且对于大多数可兴奋细胞的电信号传递而言是重要的。nav1.7存在于疼痛感知神经伤害感受器中,并且辅助传递疼痛的感觉。人scn9a基因中的功能突变的获得已经与疼痛综合征相关联,而功能突变的丧失会造成对疼痛的不敏感性。
技术实现要素:
6.本文中公开了遗传修饰成表达外源性nav1蛋白(例如,nav1.2蛋白)的非人动物的实施方案。在某些实施方案中,非人动物包含外源性scn核苷酸序列(例如,scn2a基因序列,例如,人scn2a基因序列)。本文中也公开了可用于制备这样的遗传修饰的非人动物的方法和组合物的实施方案,以及使用这样的遗传修饰的非人动物产生结合nav1.7蛋白(例如,人nav1.7蛋白)的抗体或其功能部分的方法的实施方案。scn9a是编码nav1.7蛋白的基因的名称。scn2a是编码nav1.2蛋白的基因的名称。在某些实施方案中,非人动物是啮齿动物(例如,小鼠或大鼠)。
7.在实施方案的一个方面,本文中公开了遗传修饰的啮齿动物(例如,小鼠或大鼠),其基因组(例如,种系基因组)包含编码nav1.2蛋白的核酸分子。在某些实施方案中,编码nav1.2蛋白的核酸分子是在内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座处。在某些实施方案中,编码nav1.2蛋白的核酸分子是在包括在伤害感受器中表达的基因的基因座处。在某些实施方案中,编码nav1.2蛋白的核酸分子是在转录上有活性的或允许的基因座处,例如,rosa26基因座(zambrowicz等人,1997,pnas usa 94:3789
‑
3794,其通过引用并入本文)、bt
‑
5基因座(michael等人,1999,mech.dev.85:35
‑
47,其通过引用并入本文)或oct4基因座(wallace等人,2000,nucleic acids res.28:1455
‑
1464,其通过引用并入本文)。在某些实施方案中,从遗传修饰的啮齿动物(例如,大鼠或小鼠)的基因组中的编码nav1.2蛋白的核酸分子表达nav1.2蛋白。
8.在某些实施方案中,核酸分子编码人、黑猩猩、恒河猴、马来西亚飞行狐猴(sunda flying lemur)、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟或眼镜王蛇的nav1.2蛋白。在某些实施方案中,核酸分子编码人nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少95%同一性的氨基酸序列的nav1.2蛋白。在某些
实施方案中,核酸分子编码包含与seq id no:4具有至少96%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少97%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少98%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有大于99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4相同的氨基酸序列的nav1.2蛋白。
9.在某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至啮齿动物(例如,大鼠或小鼠)scn启动子。在某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至啮齿动物(例如,大鼠或小鼠)scn9a启动子。在某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至在内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座处的内源性啮齿动物(例如,大鼠或小鼠)scn9a启动子。
10.在某些实施方案中,编码nav1.2蛋白的核酸分子是dna(例如,基因组dna或cdna)。在某些实施方案中,编码nav1.2蛋白的核酸分子包含从atg起始密码子至scn2a基因的终止密码子的邻接核苷酸的核苷酸序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的5'utr的dna序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码scn2a基因的5'utr的dna序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的3'utr的dna序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码scn2a基因的3'utr的dna序列。
11.在某些实施方案中,编码nav1.2蛋白的核酸分子是在位于内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座的内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的基因组片段处。在某些实施方案中,基因组片段包含编码内源性啮齿动物(例如,大鼠或小鼠)nav1.7蛋白的核苷酸序列。在某些实施方案中,内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的编码区(例如,从atg密码子至终止密码子)已经被替换。
12.在某些实施方案中,编码nav1.2蛋白的核酸分子是scn2a基因的基因组片段。在某些实施方案中,编码nav1.2蛋白的核酸分子是cdna。在某些实施方案中,编码nav1.2蛋白的核酸分子是重组dna。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包含从野生型序列修饰的核苷酸序列。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包含从野生型序列修饰的核苷酸序列,例如,从野生型序列优化的密码子。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包含从野生型序列修饰的核苷酸序列,例如,经修饰以从野生型序列除去t
‑
细胞表位。
13.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就编码nav1.2蛋白的核酸分子而言是杂合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就编码nav1.2蛋白的核酸分子而言是纯合的。
14.在某些实施方案中,作为内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的灭活(例如,但不限于,完全或部分缺失,或完全或部分倒转)或替换(完全或部分)的结果,遗传修饰的啮齿动物(例如,大鼠或小鼠)不能表达啮齿动物(例如,大鼠或小鼠)nav1.7蛋白。
15.在某些实施方案中,当用nav1.7免疫原(例如,人nav1.7免疫原)免疫时,遗传修饰
的啮齿动物(例如,大鼠或小鼠)产生针对nav1.7蛋白(例如,人nav1.7蛋白)的抗体。在某些实施方案中,nav1.7免疫原可以是蛋白免疫原、dna免疫原或它们的组合。
16.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含如本文中所述的人源化的免疫球蛋白重链基因座、人源化的免疫球蛋白轻链基因座或它们的组合。
17.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
h
基因区段、一个或多个人d
h
基因区段和一个或多个人j
h
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的hoh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是杂合的。
18.在某些实施方案中,包含人源化的hoh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含重链,其中每个重链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人重链可变结构域。
19.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的loh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是杂合的。
20.在某些实施方案中,包含人源化的loh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含免疫球蛋白链,其中每个免疫球蛋白链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人轻链可变结构域。
21.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个免疫球蛋白轻链恒定区基因的上游(例如,与其可操作地连接)。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vκ基因区段和一个或多个人jκ基因区段。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vλ基因区段和一个或多个人jλ基因区段。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cκ。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cλ。
22.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人vκ
基因区段和一个或多个人jκ基因区段,所述区段是在cκ基因的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的kok基因座”。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是杂合的。
23.在某些实施方案中,包含人源化的kok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含κ轻链,其中每个κ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人κ轻链可变结构域。
24.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lol基因座”。在某些实施方案中,人源化的lol基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因和一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个小鼠cλ基因包含小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是杂合的。
25.在某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。在某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
26.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在cκ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lok基因座”。在某些实施方案中,人源化的lok基因座的cκ基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例
如,大鼠或小鼠)在人源化的lok基因座处是杂合的。
27.在某些实施方案中,包含人源化的lok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含轻链,其中每个轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人λ轻链可变结构域。
28.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lik基因座”。在某些实施方案中,人源化的lik基因座的cλ基因是啮齿动物(例如,大鼠或小鼠)cλ基因。在某些实施方案中,人源化的lik基因座的cλ基因是小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是杂合的。
29.在某些实施方案中,包含人源化的lik基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。
30.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个人cλ基因上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。在某些实施方案中,这样的人源化的免疫球蛋白κ轻链基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是杂合的。在某些实施方案中,包含这样的人源化的免疫球蛋白κ轻链基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
31.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
32.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
33.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lok基因座或它们的
组合处是纯合的。
34.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
35.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
36.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
37.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
38.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lik基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
39.在某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座缺少功能性内源性啮齿动物adam6基因。在某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)表达一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段,所述核苷酸序列被包括在与人源化的免疫球蛋白重链(例如,hoh或loh)基因座相同的染色体上。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座包含一个或多个核苷酸序列,所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物
(例如,大鼠或小鼠)具有替代人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有替换人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。
40.在某些实施方案中,所提供的遗传修饰的啮齿动物具有基因组(例如,种系基因组),其含有包含第一和第二人v
h
基因区段的一个或多个人v
h
基因区段、以及在第一人v
h
基因区段和第二人v
h
基因区段之间的编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列。在某些实施方案中,第一人v
h
基因区段是v
h1‑
2且第二人v
h
基因区段是v
h6‑
1。
41.在某些实施方案中,编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列是在人v
h
基因区段和人d
h
基因区段之间。
42.在某些实施方案中,编码一种或多种啮齿动物adam6多肽的一个或多个核苷酸序列恢复或增强雄性啮齿动物的能育性。
43.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)包含外源性末端脱氧核苷酸转移酶(tdt)基因。在某些实施方案中,与没有外源性tdt基因的啮齿动物相比,包含外源性末端脱氧核苷酸转移酶(tdt)基因的啮齿动物(例如,大鼠或小鼠)可以具有增加的抗原受体多样性。
44.在某些实施方案中,如本文中所述的啮齿动物具有包含可操作地连接至转录控制元件的外源性末端脱氧核苷酸基转移酶(tdt)基因的基因组。
45.在某些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任意组合。
46.在某些实施方案中,外源性tdt位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
47.在某些实施方案中,tdt是人tdt。在某些实施方案中,tdt是tdt的短异形体(tdts)。
48.在实施方案的另一个方面,本文中公开了制备遗传修饰的啮齿动物(例如,小鼠或大鼠)的方法,所述方法包括修饰啮齿动物基因组(例如,种系基因组),使得经修饰的啮齿动物基因组包含编码nav1.2蛋白的核酸分子。在某些实施方案中,编码nav1.2蛋白的核酸分子是在内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座处。在某些实施方案中,编码nav1.2蛋白的核酸分子是在包括在伤害感受器中表达的基因的基因座处。在某些实施方案中,编码nav1.2蛋白的核酸分子是在转录上有活性的或允许的基因座处,例如,rosa26基因座(zambrowicz等人,1997,pnas usa 94:3789
‑
3794,其通过引用并入本文)、bt
‑
5基因座(michael等人,1999,mech.dev.85:35
‑
47,其通过引用并入本文)或oct4基因座(wallace等人,2000,nucleic acids res.28:1455
‑
1464,其通过引用并入本文)。在某些实施方案中,从在遗传修饰的啮齿动物(例如,大鼠或小鼠)的基因组中编码nav1.2蛋白的核酸分子表达
nav1.2蛋白,并制备包含经修饰的基因组的啮齿动物。在某些实施方案中,所述啮齿动物不表达内源性nav1.7。
49.在所述方法的某些实施方案中,通过包括以下步骤的方法来修饰啮齿动物基因组:(i)将编码nav1.2蛋白的核酸分子引入啮齿动物胚胎干(es)细胞,使得所述核酸分子整合进内源性啮齿动物scn9a基因座;(ii)得到包含经修饰的基因组的啮齿动物es细胞,其中所述核酸分子已经整合进内源性啮齿动物scn9a基因座;和(iii)从得到的包含经修饰的基因组的啮齿动物es细胞制备啮齿动物。
50.在所述方法的某些实施方案中,核酸分子编码人、黑猩猩、恒河猴、马来西亚飞行狐猴、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟或眼镜王蛇的nav1.2蛋白。在某些实施方案中,核酸分子编码人nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少95%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少96%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少97%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少98%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有至少99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4具有大于99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,核酸分子编码包含与seq id no:4相同的氨基酸序列的nav1.2蛋白。
51.在所述方法的某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至啮齿动物(例如,大鼠或小鼠)scn启动子。在所述方法的某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至啮齿动物(例如,大鼠或小鼠)scn9a启动子。在某些实施方案中,编码nav1.2蛋白的核酸分子可操作地连接至在内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座处的内源性啮齿动物(例如,大鼠或小鼠)scn9a启动子。
52.在所述方法的某些实施方案中,编码nav1.2蛋白的核酸分子是dna(例如,基因组dna或cdna)。在某些实施方案中,编码nav1.2蛋白的核酸分子包含从atg起始密码子至scn2a基因的终止密码子的邻接核苷酸的核苷酸序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的5'utr的dna序列。在某些实施方案中,所述核苷酸序列可操作地连接至scn2a基因的5'utr。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的3'utr的dna序列。在某些实施方案中,编码nav1.2蛋白的核酸分子包括编码scn2a基因的3'utr的dna序列。
53.在所述方法的某些实施方案中,编码nav1.2蛋白的核酸分子是在内源性啮齿动物(例如,大鼠或小鼠)scn9a基因座的内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的基因组片段处。在某些实施方案中,基因组片段包含编码内源性啮齿动物(例如,大鼠或小鼠)nav1.7蛋白的核苷酸序列。在某些实施方案中,内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的编码区(例如,从atg密码子至终止密码子)已经被替换。
54.在所述方法的某些实施方案中,编码nav1.2蛋白的核酸分子是scn2a基因的基因组片段。在某些实施方案中,编码nav1.2蛋白的核酸分子是cdna。在某些实施方案中,编码nav1.2蛋白的核酸分子是重组dna。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包
含从野生型序列修饰的核苷酸序列。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包含从野生型序列修饰的核苷酸序列,例如,从野生型序列优化的密码子。在某些实施方案中,编码nav1.2蛋白的核酸分子可以包含从野生型序列修饰的核苷酸序列,例如,从野生型序列修饰以除去t
‑
细胞表位。
55.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就编码nav1.2蛋白的核酸分子而言是杂合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就编码nav1.2蛋白的核酸分子而言是纯合的。
56.在所述方法的某些实施方案中,由于内源性啮齿动物(例如,大鼠或小鼠)scn9a基因的灭活(例如,但不限于,完全或部分缺失,或完全或部分倒转)或替换(完全或部分),遗传修饰的啮齿动物(例如,大鼠或小鼠)不能表达啮齿动物(例如,大鼠或小鼠)nav1.7蛋白。
57.在所述方法的某些实施方案中,当用nav1.7免疫原(例如,人nav1.7免疫原)免疫时,遗传修饰的啮齿动物(例如,大鼠或小鼠)产生针对nav1.7蛋白(例如,人nav1.7蛋白)的抗体。在某些实施方案中,nav1.7免疫原可以是蛋白免疫原、dna免疫原或它们的组合。
58.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含如本文中所述的人源化的免疫球蛋白重链基因座、人源化的免疫球蛋白轻链基因座或它们的组合。
59.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
h
基因区段、一个或多个人d
h
基因区段和一个或多个人j
h
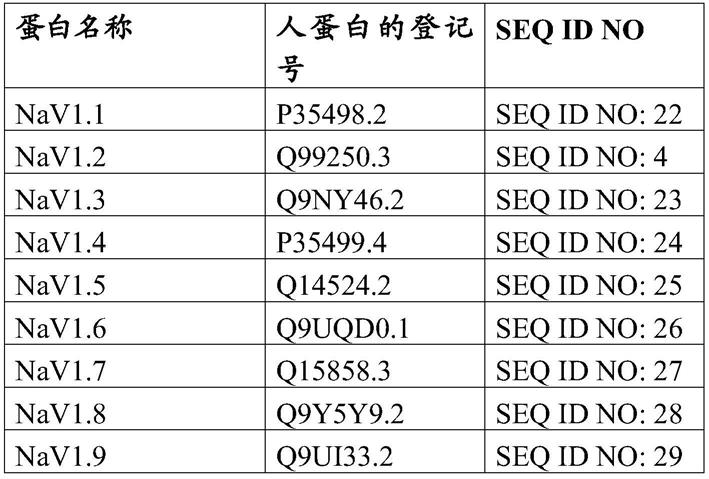
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的hoh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是杂合的。
60.在所述方法的某些实施方案中,包含人源化的hoh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含重链,其中每个重链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人重链可变结构域。
61.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的loh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是杂合的。
62.在所述方法的某些实施方案中,包含人源化的loh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含免疫球蛋白链,其
中每个免疫球蛋白链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人轻链可变结构域。
63.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含含有一个或多个人v
l
基因区段和一个或多个人j
l
基因区段的人源化的免疫球蛋白轻链基因座,所述区段是在一个或多个免疫球蛋白轻链恒定区基因的上游(例如,与其可操作地连接)。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vκ基因区段和一个或多个人jκ基因区段。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vλ基因区段和一个或多个人jλ基因区段。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cκ。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cλ。
64.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人vκ基因区段和一个或多个人jκ基因区段,所述区段是在cκ基因的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的kok基因座”。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是杂合的。
65.在所述方法的某些实施方案中,包含人源化的kok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含κ轻链,其中每个κ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人κ轻链可变结构域。
66.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lol基因座”。在某些实施方案中,人源化的lol基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因和一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个小鼠cλ基因包含小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是杂合的。
67.在所述方法的某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ
轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。在所述方法的某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
68.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在cκ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lok基因座”。在某些实施方案中,人源化的lok基因座的cκ基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lok基因座处是杂合的。
69.在所述方法的某些实施方案中,包含人源化的lok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含轻链,其中每个轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人λ轻链可变结构域。
70.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lik基因座”。在某些实施方案中,人源化的lik基因座的cλ基因是啮齿动物(例如,大鼠或小鼠)cλ基因。在某些实施方案中,人源化的lik基因座的cλ基因是小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是杂合的。
71.在所述方法的某些实施方案中,包含人源化的lik基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。
72.在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个人cλ基因上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。在某些实施方案中,这样的人源化的免疫球蛋白κ轻链基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是杂合的。在某些实施方案中,包含这样的人源化的免疫球蛋白κ轻链基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻
链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
73.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
74.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
75.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lok基因座或它们的组合处是纯合的。
76.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
77.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
78.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
79.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
80.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lik基因座。在某些实施
方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
81.在所述方法的某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座缺少功能性内源性啮齿动物adam6基因。在某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)表达一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段,所述核苷酸序列被包括在与人源化的免疫球蛋白重链(例如,hoh或loh)基因座相同的染色体上。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座包含一个或多个核苷酸序列,所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有替代人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有替换人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。
82.在所述方法的某些实施方案中,所提供的遗传修饰的啮齿动物具有基因组(例如,种系基因组),其含有包含第一和第二人v
h
基因区段的一个或多个人v
h
基因区段、以及在第一人v
h
基因区段和第二人v
h
基因区段之间的编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列。在某些实施方案中,第一人v
h
基因区段是v
h1‑
2且第二人v
h
基因区段是v
h6‑
1。
83.在所述方法的某些实施方案中,编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列是在人v
h
基因区段和人d
h
基因区段之间。
84.在所述方法的某些实施方案中,编码一种或多种啮齿动物adam6多肽的一个或多个核苷酸序列恢复或增强雄性啮齿动物的能育性。
85.在所述方法的某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)包含外源性末端脱氧核苷酸转移酶(tdt)基因。在某些实施方案中,与没有外源性tdt基因的啮齿动物相比,包含外源性末端脱氧核苷酸转移酶(tdt)基因的啮齿动物(例如,大鼠或小鼠)可以具有增加的抗原受体多样性。
86.在所述方法的某些实施方案中,如本文中所述的啮齿动物具有包含可操作地连接至转录控制元件的外源性末端脱氧核苷酸基转移酶(tdt)基因的基因组。
87.在所述方法的某些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任意组合。
88.在所述方法的某些实施方案中,外源性tdt位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
89.在所述方法的某些实施方案中,tdt是人tdt。在某些实施方案中,tdt是tdt的短异形体(tdts)。
90.在实施方案的另一个方面,本文中公开了分离的啮齿动物细胞或啮齿动物组织,其基因组包含在内源性啮齿动物scn9a基因座处的编码nav1.2蛋白的核酸分子。在某些实施方案中,所述分离的啮齿动物细胞或啮齿动物组织是小鼠细胞或小鼠组织或大鼠细胞或大鼠组织。在某些实施方案中,所述分离的啮齿动物细胞或啮齿动物组织是小鼠细胞或小鼠组织。在某些实施方案中,所述分离的啮齿动物细胞或啮齿动物组织是大鼠细胞或大鼠组织。
91.在某些实施方案中,分离的啮齿动物细胞是啮齿动物es细胞。在某些实施方案中,分离的啮齿动物细胞是b细胞。
92.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
h
基因区段、一个或多个人d
h
基因区段和一个或多个人j
h
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的hoh基因座”。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座处是杂合的。
93.在某些实施方案中,分离的啮齿动物细胞是包含人源化的hoh基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含重链,其中每个重链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人重链可变结构域。
94.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个免疫球蛋白轻链恒定区基因的上游(例如,与其可操作地连接)。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vκ基因区段和一个或多个人jκ基因区段。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vλ基因区段和一个或多个人jλ基因区段。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cκ。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cλ。
95.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人vκ基因区段和一个或多个人jκ基因区段,所述区段是在cκ基因的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的kok基因座”。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施
方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的kok基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的kok基因座处是杂合的。
96.在某些实施方案中,分离的啮齿动物细胞是包含人源化的kok基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含κ轻链,其中每个κ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人κ轻链可变结构域。
97.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lol基因座”。在某些实施方案中,人源化的lol基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因和一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个小鼠cλ基因包含小鼠cλ1基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lol基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lol基因座处是杂合的。
98.在某些实施方案中,分离的啮齿动物细胞是包含人源化的lol基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。在某些实施方案中,分离的啮齿动物细胞是包含人源化的lol基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
99.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在cκ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lok基因座”。在某些实施方案中,人源化的lok基因座的cκ基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lok基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lok基因座处是杂合的。
100.在某些实施方案中,分离的啮齿动物细胞是包含人源化的lok基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含轻链,其中每个轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人λ轻链可变结构域。
101.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lik基因座”。在某些实施方案中,人源化的lik基因座的cλ基因是啮齿动物(例如,大鼠或小鼠)cλ基因。在某些实施方案中,人源化的lik基因座的cλ基因是小鼠cλ1基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lik基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lik基因座处是杂合的。
102.在某些实施方案中,分离的啮齿动物细胞是包含人源化的lik基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。
103.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个人cλ基因上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。在某些实施方案中,这样的人源化的免疫球蛋白κ轻链基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织就这样的人源化的免疫球蛋白κ轻链基因座而言是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织就这样的人源化的免疫球蛋白κ轻链基因座而言是杂合的。在某些实施方案中,分离的啮齿动物细胞是b细胞或脾细胞,其包含这样的人源化的免疫球蛋白κ轻链基因座,且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
104.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
105.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
106.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的lok基因座或它们的组合处是纯合的。
107.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
108.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动
物组织在其基因组中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
109.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
110.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
111.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的lik基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
112.在某些实施方案中,本文提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含缺乏内源性啮齿动物adam6基因的人源化的免疫球蛋白重链(例如,hoh或loh)基因座。在某些实施方案中,本文提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含编码一种或多种啮齿动物adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织表达一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列,所述核苷酸序列被包括在与人源化的免疫球蛋白重链(例如,hoh或loh)基因座相同的染色体上。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座,所述基因座包含编码一种或多种啮齿动物adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有替代人adam6假基因的基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有替换人adam6假基因的基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。
113.在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含含有第一和第二人v
h
基因区段的一个或多个人v
h
基因区段、以及在第一人v
h
基因区段和第二人v
h
基因区段之间的编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,第一人v
h
基因区段是v
h1‑
2且第二人v
h
基因区段是v
h6‑
1。
114.在某些实施方案中,编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列是在人v
h
基因区段和人d
h
基因区段之
间。
115.在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织包含外源性末端脱氧核苷酸转移酶(tdt)基因。在某些实施方案中,分离的啮齿动物细胞是包含外源性末端脱氧核苷酸转移酶(tdt)基因的b细胞或脾细胞,且当与不具有外源性tdt基因的分离的啮齿动物细胞(例如,b细胞或脾细胞)相比时可以具有增加的抗原受体多样性。
116.在某些实施方案中,如本文中所述的分离的啮齿动物细胞或啮齿动物组织具有包含可操作地连接至转录控制元件的外源性末端脱氧核苷酸基转移酶(tdt)基因的基因组。
117.在某些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任意组合。
118.在某些实施方案中,外源性tdt位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
119.在某些实施方案中,tdt是人tdt。在某些实施方案中,tdt是tdt的短异形体(tdts)。
120.在实施方案的另一个方面,本文中公开了包含本文描述的啮齿动物es细胞的啮齿动物胚胎。
121.在实施方案的一个方面,本文中公开了一种靶向核酸构建体,其包含编码nav1.2蛋白的核酸分子,所述核酸分子侧接能够介导所述核酸分子向内源性啮齿动物scn9a基因座中的同源重组和整合的5'和3'啮齿动物核苷酸序列。
122.在实施方案的另一个方面,本文中公开了生产抗
‑
nav1.7抗体的方法,所述方法包括用nav1.7免疫原(例如,人nav1.7免疫原)免疫本文描述的遗传修饰的啮齿动物(例如,大鼠或小鼠)。在某些实施方案中,生产抗
‑
nav1.7抗体的方法包括从经免疫的啮齿动物分离抗
‑
nav1.7抗体。在某些实施方案中,所述抗体是单克隆抗体。在某些实施方案中,生产抗
‑
nav1.7抗体的方法包括从经免疫的啮齿动物分离表达抗
‑
nav1.7抗体的b细胞。在某些实施方案中,还提供了生产抗
‑
人nav1.7抗体的杂交瘤。在某些实施方案中,生产抗
‑
人nav1.7抗体的杂交瘤源自从经免疫的啮齿动物分离的b细胞。
123.在实施方案的另一个方面,本文中公开了生产抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的方法,所述方法包括用nav1.7免疫原(例如,人nav1.7免疫原)免疫本文描述的遗传修饰的啮齿动物(例如,大鼠或小鼠)。在某些实施方案中,生产抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的方法包括使遗传修饰的小鼠产生对nav1.7免疫原的免疫应答。在某些实施方案中,生产抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的方法包括从表达抗
‑
nav1.7抗体的遗传修饰的小鼠分离b细胞。在某些实施方案中,生产抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的方法包括确定由遗传修饰的小鼠产生的抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的氨基酸序列。在某些实施方案中,生产抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的方法包括表达多肽,所述多肽包含鉴定的人重链和/或轻链可变结构域。在某些实施方案中,确定人重链和/或轻链可变结构域的氨基酸序列包括确定分别编码人重链和/或轻链可变结构域的核苷酸序列。
124.在实施方案的另一个方面,本文中公开了生产分别编码抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的人重链和/或轻链可变区的方法,所述方法包括用nav1.7免疫原
(例如,人nav1.7免疫原)免疫本文描述的遗传修饰的啮齿动物(例如,大鼠或小鼠)。在某些实施方案中,生产分别编码抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的人重链和/或轻链可变区的方法包括使遗传修饰的啮齿动物产生对nav1.7免疫原的免疫应答。在某些实施方案中,生产分别编码抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的人重链和/或轻链可变区的方法包括从表达抗
‑
nav1.7抗体的遗传修饰的啮齿动物分离b细胞。在某些实施方案中,生产分别编码抗
‑
nav1.7抗体的人重链和/或轻链可变结构域的人重链和/或轻链可变区的方法包括确定由遗传修饰的啮齿动物产生的抗
‑
nav1.7抗体的人重链和/或轻链可变区的核酸序列。
125.在实施方案的另一个方面,本文中公开了一种核酸,其编码与本文描述的啮齿动物(例如,大鼠或小鼠)相同或从其得到的抗
‑
nav1.7抗体的人重链和/或轻链可变结构域。
126.在实施方案的另一个方面,本文中公开了一种核酸,其编码与本文描述的啮齿动物(例如,大鼠或小鼠)相同或从其得到的包含抗
‑
nav1.7抗体的人重链可变结构域的免疫球蛋白重链。在某些实施方案中,编码免疫球蛋白重链的核酸进一步包含人重链恒定结构域。在某些实施方案中,编码免疫球蛋白重链的核酸进一步包含啮齿动物(例如,大鼠或小鼠)重链恒定结构域。
127.在实施方案的另一个方面,本文中公开了一种核酸,其编码与本文描述的啮齿动物(例如,大鼠或小鼠)相同或从其得到的包含抗
‑
nav1.7抗体的人轻链可变结构域的免疫球蛋白轻链。在某些实施方案中,编码免疫球蛋白轻链的核酸进一步包含人轻链恒定结构域。在某些实施方案中,编码免疫球蛋白轻链的核酸进一步包含啮齿动物(例如,大鼠或小鼠)轻链恒定结构域。
128.在实施方案的另一个方面,本文中公开了与本文描述的啮齿动物(例如,大鼠或小鼠)相同、从其得到或从其衍生出的抗
‑
nav1.7抗体的人重链和/或轻链可变结构域。
129.在实施方案的另一个方面,本文中公开了表达抗
‑
nav1.7抗体的哺乳动物细胞,所述抗体包含与本文描述的啮齿动物(例如,大鼠或小鼠)相同、从其得到或从其衍生出的抗
‑
nav1.7抗体的重链和轻链可变结构域。在某些实施方案中,哺乳动物细胞是cho细胞(例如,cho k1、dxb
‑
11cho、veggie
‑
cho)、cos(例如,cos
‑
7)、视网膜细胞、vero细胞、cv1细胞、肾细胞(例如,hek293、293ebna、msr 293、mdck、hak、bhk)、hela细胞、hepg2细胞、wi38细胞、mrc 5细胞、colo205细胞、hb8065细胞、hl
‑
60细胞、(例如,bhk21)、jurkat细胞、daudi细胞、a431细胞(表皮)、u937细胞、3t3细胞、l细胞、c127细胞、sp2/0细胞、ns
‑
0细胞、mmt 060562细胞、塞尔托利细胞、brl 3a细胞、ht1080细胞、骨髓瘤细胞、肿瘤细胞和从前述细胞衍生出的细胞系的细胞。
附图说明
130.本专利或申请的文件含有至少一幅彩色绘制的图。在请求并支付必要的费用后,专利和商标局将提供具有彩图的本专利的副本。
131.图1a
‑
1d.用于将人scn2a敲入小鼠scn9a基因座中的示例性策略。图1a显示了人scn2a和小鼠scn9a基因的基因组组构的非按比例的简图。用置于基因组序列上方的细条代表外显子。指示了要删除的约84,847bp的小鼠基因组片段和要插入的约96,735bp的人基因组片段。用星号指示在实施例1的表1中描述的测定所使用的探针的位置。图1b非按比例解
释了用于将人scn2a敲入小鼠scn9a基因座的示例性的修饰的bac载体以及连接部序列(seq id no:17,18和19)。图1c非按比例解释了在新霉素盒已经删除后具有敲入的人scn2a的人源化的小鼠scn9a基因座,以及连接部序列(seq id no:17和21)。图1d描述了小鼠scn9a(nav1.7)蛋白的氨基酸2
‑
1984(seq id no:2)和人scn2a(nav1.2)蛋白的氨基酸4
‑
2005(seq id no:4)的序列比对。
132.图2.是显示使用蛋白和dna免疫原在mnav1.7 ko中的hnav1.2ki/vi
‑
3小鼠的免疫应答的示例性分析的一个实施方案。最初将小鼠用dna免疫原免疫,并转换至蛋白免疫原作为强化。本文中使用的术语“vi
‑
3”表示包括hoh基因座和kok基因座的本文中公开的小鼠的一个实施方案。具体地,如该术语在本文中使用的,vi
‑
3小鼠就包含80个人v
h
基因区段、27个人d
h
基因区段和六个人j
h
基因区段的hoh基因座而言是纯合的;vi
‑
3小鼠就包含40个人vκ基因区段和至少一个人jκ基因区段的kok基因座而言也是纯合的。
133.图3a.人nav蛋白的示例性序列的比对,从上至下:分别是人nav1.1(seq id no:22)、人nav1.2(seq id no:4)、人nav1.3(seq id no:23)、人nav1.4(seq id no:24)、人nav1.5(seq id no:25)、人nav1.6(seq id no:26)、人nav1.7(seq id no:27)、人nav1.8(seq id no:28)和人nav1.9(seq id no:29)。基于人nav1.7来标记结构域。“cyto”:细胞质的(绿色);“tm”:跨膜(蓝色);“ec”:细胞外的(粉色);“孔形成”:参与形成孔(离子在其中穿过)的细胞外结构域的部分(棕色)。
134.图3b.九种人nav蛋白的系统树。
135.图4a.来自15个动物物种的nav1.7蛋白的示例性序列的比对,从上至下:分别是人(seq id no:27)、黑猩猩(异形体x1)(seq id no:30)、恒河猴(seq id no:31)、马来西亚飞行狐猴(异形体x1)(seq id no:32)、牛(seq id no:33)、绵羊(异形体x1)(seq id no:34)、阿拉伯骆驼(seq id no:35)、杀人鲸(异形体x1)(seq id no:36)、马(seq id no:37)、狗(异形体x1)(seq id no:38)、小鼠(seq id no:2)、大鼠(seq id no:39)、兔(seq id no:40)、鸡(seq id no:41和眼镜王蛇(部分)(seq id no:42)。“cyto”:细胞质的(绿色);“tm”:跨膜(蓝色);“ec”:细胞外的(粉色);“孔形成”:参与形成孔(离子在其中穿过)的细胞外结构域的部分(棕色)。
136.图4b.来自15个动物物种的nav1.7蛋白的系统树。
137.图5a.来自15个动物物种的nav1.2蛋白的示例性序列的比对,从上至下:分别是人(seq id no:4)、黑猩猩(异形体x1)(seq id no:43)、恒河猴(异形体x1)(seq id no:44)、马来西亚飞行狐猴(异形体x1)(seq id no:45)、牛(seq id no:46)、绵羊(异形体x1)(seq id no:47)、阿拉伯骆驼(seq id no:48)、杀人鲸(异形体1)(seq id no:49)、马(seq id no:50)、小鼠(异形体1)(seq id no:51)、大鼠(seq id no:52)、兔(异形体x1)(seq id no:53)、鸡(seq id no:54)、眼镜王蛇(部分)(seq id no:55)和绿海龟(seq id no:56)。“cyto”:细胞质的(绿色);“tm”:跨膜(蓝色);“ec”:细胞外的(粉色);“孔形成”:参与形成孔(离子在其中穿过)的细胞外结构域的部分(棕色)。
138.图5b.来自15个动物物种的nav1.2蛋白的系统树。
139.图6.显示了7506等位基因的核苷酸序列(seq id no:20),即,在具有neo自删除盒的小鼠scn9a基因座中的人scn2a,其包括小鼠核苷酸(小写字母)、sgrdi位点(粗体,下划线)、人核苷酸(粗体,大写字体)、xhoi位点(粗体,下划线)、loxp(斜体字)、鱼精蛋白启动子
(粗体,下划线)、crei(斜体字)、sv40聚腺苷酸(小写字母)、hubi prm(粗体)
‑
em7 prm(粗体,下划线)、neo(斜体字)、pgk聚腺苷酸(下划线)、loxp(斜体字)、iceui(下划线)、nhei(粗体,下划线)、小鼠核苷酸(小写字母)。
140.图7a
‑
7c.在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠具有对热刺激的受损应答和对组胺的显著降低的痒应答。7a,在mnav1.7 ko中的hnav1.2ki/vi
‑
3小鼠表现出对热刺激的应答的显著延长的潜伏期(hargreaves,在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠为22.9
±
0.9s,n=15,与此相比,wt小鼠为12.3
±
0.5s,n=19,未配对student氏t检验,p<0.0001)。7b,在热板试验中的爪缩回潜伏期。在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠没有对52.5或55℃热刺激做出应答,并且在30秒的预定截止时间从热板移开以避免组织损伤。另一方面,wt小鼠快速地表现出响应于热刺激的防伤害行为(55℃:wt为6.5
±
0.5s,n=9,和在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠为30s,n=7;52.5℃:wt为10.4
±
0.6s,和在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠为30s,n=7;p<0.0001)。7c,在颈背中真皮内注射150μg组胺以后搔抓发作的总数。在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠表现出比wt小鼠少3.7倍的搔抓发作(在mnav1.7 ko中的hnav1.2ki/vi
‑
3小鼠为24
±
11次发作,与此相比,wt为81
±
20次发作,未配对的student氏t检验p=0.047)。
具体实施方式
141.本文中公开了遗传修饰成表达外源性nav1蛋白(例如,nav1.2蛋白)的非人动物的实施方案。在某些实施方案中,非人动物包含外源性scn核苷酸序列(例如,scn2a基因序列,例如,人scn2a基因序列)。本文中也公开了可用于制备这样的遗传修饰的非人动物的方法和组合物的实施方案,以及使用这样的遗传修饰的非人动物产生结合nav1.7蛋白(例如,人nav1.7蛋白)的抗体或其功能部分的方法的实施方案。scn9a是编码nav1.7蛋白的基因的名称。scn2a是编码nav1.2蛋白的基因的名称。在某些实施方案中,非人动物是啮齿动物(例如,小鼠或大鼠)。
142.nav家族
143.电压门控钠通道的家族具有九个已知的成员,在跨膜区段和细胞外环区域中具有>50%的氨基酸同一性。这些通道的蛋白被命名为nav1.1至nav1.9,且基因名被称作scn1a至scn11a。参见下面表1。
144.表1
145.蛋白名称基因名称nav1.1scn1anav1.2scn2anav1.3scn3anav1.4scn4anav1.5scn5anav1.6scn8anav1.7scn9anav1.8scn10anav1.9scn11a
146.示例性人nav蛋白序列的比对提供在图3a中,在表2中阐述了登录号和序列标识符。在图3b中描绘了人nav蛋白的相关性。
147.表2
[0148][0149]
钠通道家族的这些成员(参见表1)具有四个重复结构域,每个含有六个跨膜区段。参见图3a。第四个区段是高度保守的并作为通道的电压传感器起作用。该通道的电压敏感性是由于位于第四个区段的每第三位置的正氨基酸(nicholls等人,(2012)“from neuron to brain”,第5版第86页,其通过引用整体并入本文)。当受到跨膜电压的变化刺激时,该区段向细胞膜的细胞外侧移动,使通道变成离子可渗透的。离子被引导穿过孔,所述孔可以分成两个区域。孔的更外部(即,更细胞外的)部分由四个结构域各自的第五和第六跨膜区段之间的区域(也被称作“p
‑
环”)形成。该区域是孔的更狭窄部分并且负责其离子选择性。孔的更内部(即,更细胞质的)部分由四个结构域的组合的第五和第六跨膜区段形成。
[0150]
nav1.7
[0151]
nav1.7在背根神经节、交感神经元、许旺细胞和神经内分泌细胞处的伤害性(疼痛)神经元中表达。nav1.7是膜兴奋性的关键组分,并且对于疼痛的感觉而言是重要的。人scn9a基因中的功能突变的获得已经与疼痛综合征相关联,而功能突变的丧失与对疼痛的极度不敏感性有关。合乎需要的是,开发选择性的nav1.7通道阻滞剂作为镇痛药。
[0152]
nav1.7是在物种之间高度保守的,如图4a中来自15个动物物种的nav1.7蛋白的示例性序列的比对和图4b中的关系树所证实的。在比对中包括的示例性序列的登录号和序列标识符如下面表3中所示。
[0153]
表3
[0154]
nav1.7的物种登记号seq id no人q15858.3seq id no:27黑猩猩xp_016804947.1seq id no:30恒河猴xp_014965766.1seq id no:31马来西亚飞行狐猴xp_008588371.1seq id no:32牛np_001104257.2seq id no:33
绵羊xp_004004679.1seq id no:34阿拉伯骆驼xp_010980767.1seq id no:35杀人鲸xp_004267302.1seq id no:36马xp_001496473.1seq id no:37狗xp_022270547.1seq id no:38小鼠q62205.2seq id no:2大鼠o08562.1seq id no:39兔q28644.1seq id no:40鸡np_001280211.1seq id no:41眼镜王蛇(部分序列)daa65084.1seq id no:42
[0155]
nav1.2
[0156]
nav1.2在中枢神经元和周围神经元中表达。人scn2a基因(编码nav1.2)中的突变已经与几种癫痫发作障碍和孤独症谱群障碍相关联。
[0157]
nav1.2是在物种之间高度保守的,如图5a中提供的来自15个动物物种的nav1.2蛋白的示例性序列的比对所证实的。在比对中包括的示例性序列的登录号和序列标识符如下面表4中所示。
[0158]
表4
[0159]
nav1.2的物种登记号seq id no人q99250.3seq id no:4黑猩猩xp_003820970.1seq id no:43恒河猴xp_001100368.1seq id no:44马来西亚飞行狐猴xp_008582720.1seq id no:45牛np_001137581.1seq id no:46绵羊xp_014948870.1seq id no:47阿拉伯骆驼xp_010980763.1seq id no:48杀人鲸xp_004283641.1seq id no:49马xp_014588001.1seq id no:50小鼠np_001092768.1seq id no:51大鼠p04775.1seq id no:52兔xp_008256915.1seq id no:53鸡np_001280210.1seq id no:54眼镜王蛇(部分序列)ete69867.1seq id no:55绿海龟xp_007056690.1seq id no:56
[0160]
遗传修饰的啮齿动物
[0161]
在一些实施方案的一个方面,本公开内容涉及遗传修饰的啮齿动物,其中遗传修饰包含外源性scn基因的至少一部分向内源性scn9a基因座中的插入。
[0162]
在某些实施方案中,本公开内容提供了遗传修饰的啮齿动物,其基因组包含在内源性scn9a基因座处的核酸分子,其中所述核酸分子编码nav蛋白且包含外源性scn基因的至少一部分。
[0163]
本文中使用的术语“人源化的”包括被修饰成包含人序列。例如,人源化的基因座是已经被修饰成包含人序列(例如,基因区段或基因)的基因座(例如,内源性基因座)。
[0164]
本文中使用的术语“种系基因组”表示在动物形成所用的生殖细胞(例如,配子,例如,精子或卵子)中发现的基因组。种系基因组是动物中的细胞的基因组dna的来源。这样,在其种系基因组中具有修饰的动物(例如,小鼠或大鼠)被认为在其所有细胞的基因组dna中具有修饰。
[0165]
本文中使用的术语“替换”表示位置置换,其中第一核酸序列位于染色体中第二核酸序列的位置(例如,其中第二核酸序列在以前(例如,最初)位于染色体中,例如,在第二核酸序列的内源性基因座)。短语“替换”不要求第二核酸序列从例如基因座或染色体除去。在某些实施方案中,第二核酸序列和第一核酸序列是彼此相容的,因为,例如,第一和第二序列是彼此同源的,含有对应的元件(例如,蛋白编码元件、调节元件等),和/或具有类似的或相同的序列。在某些实施方案中,第一和/或第二核酸序列包括启动子、增强子、剪接供体位点、剪接受体位点、内含子、外显子、非翻译区(utr)中的一个或多个;在某些实施方案中,第一和/或第二核酸序列包括一个或多个编码序列。在某些实施方案中,第一核酸序列是第二核酸序列的同系物或变体(例如,突变体)。在某些实施方案中,第一核酸序列是第二序列的直系同源物或同系物。在某些实施方案中,第一核酸序列是或包含人核酸序列。在某些实施方案中,包括其中第一核酸序列是或包含人核酸序列,第二核酸序列是或包含啮齿动物序列(例如,小鼠或大鼠序列)。在某些实施方案中,包括其中第一核酸序列是或包含人核酸序列,第二核酸序列是或包含人序列。在某些实施方案中,第一核酸序列是第二序列的变体或突变体(即,与第二序列相比含有一个或多个序列差异(例如,置换)的序列)。如此放置的核酸序列可以包括一个或多个调节序列(例如,启动子、增强子、5'
‑
或3'
‑
非翻译区等),所述调节序列是用于获得如此放置的序列的源核酸序列的部分。例如,在各个实施方案中,第一核酸序列是用异源序列对内源序列的置换,其导致从如此放置的核酸序列(其包含异源序列)产生基因产物,但是不表达内源序列;第一核酸序列属于内源性基因组序列,其具有编码多肽的核酸序列,所述多肽具有与内源序列所编码的多肽类似的功能(例如,内源性基因组序列完全地或部分地编码非人可变区多肽,且dna片段完全地或部分地编码一个或多个人可变区多肽)。在各个实施方案中,人免疫球蛋白基因区段或其片段替换内源性非人免疫球蛋白基因区段或片段。
[0166]
本文中使用的术语“nav蛋白”包括(1)nav家族的天然存在的(野生型)电压门控钠通道,即,nav1.1、nav1.2、nav1.3、nav1.4、nav1.5、nav1.6、nav1.7、nav1.8和nav1.9,和(2)经工程改造的电压门控钠通道。经工程改造的nav蛋白维持天然存在的nav蛋白所特有的四个重复结构域结构,其中每个结构域含有六个跨膜区段,并且也作为电压门控钠通道(如天然存在的nav蛋白)起作用。经工程改造的电压门控钠通道的一个非限制性实施方案是嵌合蛋白,其包括nav1.2蛋白的细胞外结构域以及啮齿动物nav1.7蛋白的跨膜和细胞质结构域。
[0167]
本文中使用的术语“scn基因”包括编码天然存在的nav蛋白的核酸。“外源性scn基因”是指不存在于啮齿动物scn9a基因座内的scn基因,因为该基因座存在于自然界中。在某些实施方案中,外源性scn基因是并非啮齿动物scn9a的scn基因。在某些实施方案中,scn基因是或包含scn1a、scn2a、scn3a、scn4a、scn5a、scn8a、scn10a或scn11a基因。在某些实施方
no:4具有至少99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,人scn2a基因的编码序列编码包含与seq id no:4具有大于99%同一性的氨基酸序列的nav1.2蛋白。在某些实施方案中,人scn2a基因的编码序列编码包含与seq id no:4相同的氨基酸序列的nav1.2蛋白。
[0175]
在本文所公开的遗传修饰的啮齿动物的某些实施方案中,啮齿动物的基因组包含在内源性scn9a基因座处的核酸分子,其中所述核酸分子包含外源性scn基因的部分和内源性scn9a基因的部分,且其中所述核酸分子编码nav蛋白,所述nav蛋白包含由外源性scn基因编码的nav蛋白的部分。nav蛋白的“部分”意在包括对nav蛋白的至少2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20个或更多个氨基酸的连续序列的提及,但是短于全长nav蛋白。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少2个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少3个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少4个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少5个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少6个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少7个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少8个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少9个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少10个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少11个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少12个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少13个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少14个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少15个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少16个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少17个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少18个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少19个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分包含nav蛋白的至少20个氨基酸的连续序列。在某些实施方案中,nav蛋白的部分是nav蛋白的结构域,诸如细胞外结构域、跨膜结构域或细胞质结构域。
[0176]
在某些实施方案中,所述核酸分子包含外源性scn基因的部分,其编码由外源性scn基因编码的nav蛋白的细胞外结构域,使得在内源性scn9基因座处的核酸分子编码包含由外源性scn基因编码的nav蛋白的细胞外结构域的nav蛋白。
[0177]
在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因选自scn1a、scn2a、scn3a、scn4a、scn5a、scn 8a、scn10a和scn11a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn1a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn2a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸
分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn3a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn5a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn8a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn10a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域,其中所述外源性scn基因是scn11a基因。在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含scn2a基因的部分,所述部分编码由外源性scn2a基因编码的nav1.2蛋白的细胞外结构域。
[0178]
在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含来自不同于被修饰的啮齿动物物种的物种的外源性scn基因的部分,所述物种包括、但不限于人、黑猩猩、恒河猴、马来西亚飞行狐猴、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟和眼镜王蛇。
[0179]
在某些实施方案中,在内源性啮齿动物scn9a基因座处的核酸分子包含外源性scn2a基因的部分,所述部分编码由外源性scn2a基因编码的nav1.2蛋白的细胞外结构域,其中所述外源性scn2a基因来自选自人、黑猩猩、恒河猴、马来西亚飞行狐猴、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟和眼镜王蛇的物种。在某些实施方案中,所述外源性scn2a基因是人scn2a基因,且在内源性啮齿动物scn9a基因座处的核酸分子包含人scn2a基因的部分,所述部分编码人nav1.2蛋白的细胞外结构域。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少95%、至少96%、至少97%、至少98%、至少99%或大于99%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少95%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少96%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少97%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少98%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有至少99%同一性的氨基酸序列。在某些实施方案中,所述人nav1.2蛋白包含与seq id no:4具有大于99%同一性的氨基酸序列。在具体实施方案中,人scn2a基因编码包含与seq id no:4相同的氨基酸序列的nav1.2蛋白。在图1d中描绘了seq id no:4的人nav1.2蛋白的细胞外结构域(在细胞外结构域与跨膜或细胞质结构域之间的连接部可以从图1d中描绘的那些偏移1
‑
2个氨基酸)。
[0180]
在某些实施方案中,除了编码细胞外结构域的外源性scn基因的部分以外,在内源性啮齿动物scn9a基因座处的核酸分子也包含编码内源性啮齿动物nav1.7蛋白的跨膜和细胞质结构域的内源性啮齿动物scn9a基因的部分。在某些实施方案中,所述啮齿动物是小鼠,且在内源性小鼠scn9a基因座处的核酸分子包含编码内源性小鼠nav1.7蛋白的跨膜和细胞质结构域的内源性小鼠scn9a基因的部分。在某些实施方案中,内源性小鼠scn9a基因
编码与seq id no:2具有至少95%、至少96%、至少97%、至少98%、至少99%或大于99%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有至少95%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有至少96%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有至少97%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有至少98%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有至少99%同一性的小鼠nav1.7蛋白。在某些实施方案中,内源性小鼠scn9a基因编码与seq id no:2具有大于99%同一性的小鼠nav1.7蛋白。在一个具体实施方案中,内源性小鼠nav1.7蛋白包含与seq id no:2相同的氨基酸序列。在图1d中描绘了seq id no:2的小鼠nav1.7蛋白的跨膜和细胞质结构域(在细胞外结构域与跨膜或细胞质结构域之间的连接部可以从图1d中描绘的那些偏移1
‑
2个氨基酸)。
[0181]
在某些实施方案中,所述啮齿动物是大鼠,且在内源性大鼠scn9a基因座处的核酸分子包含编码内源性大鼠nav1.7蛋白的跨膜和细胞质结构域的内源性大鼠scn9a基因的部分。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少95%、至少96%、至少97%、至少98%、至少99%或大于99%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少95%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少96%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少97%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少98%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有至少99%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠scn9a基因编码与seq id no:39具有大于99%同一性的大鼠nav1.7蛋白。在某些实施方案中,内源性大鼠nav1.7蛋白包含与seq id no:39相同的氨基酸序列。
[0182]
在本文公开的遗传修饰的啮齿动物的某些实施方案中,存在于内源性scn9a基因座中的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子是cdna分子。在某些实施方案中,存在于内源性scn9a基因座中的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子是基因组dna。
[0183]
在某些实施方案中,存在于内源性scn9a基因座中的编码nav蛋白且包含外源性scn基因的至少一部分的核酸可以源自遗传修饰,其中在内源性啮齿动物scn9a基因座处的内源性scn9a基因已经被完全或部分的外源性scn基因完全或部分替换。在某些实施方案中,包含内源性啮齿动物scn9a基因的编码序列(例如,从atg密码子至终止密码子)的基因组片段已经被外源性scn基因的编码序列(例如,从atg密码子至终止密码子,在基因组dna或cdna中)替换;且在某些实施方案中,所述外源性scn基因是scn2a基因,例如,来自人、黑猩猩、恒河猴、马来西亚飞行狐猴、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟或眼镜王蛇的scn2a基因。在某些实施方案中,所述外源性scn基因是人scn2a基因。在某些实施方案中,编码内源性啮齿动物nav1.7蛋白的一个或多个或全部细胞外结构域的内源性啮齿动物scn9a基因的部分已经被编码由外源性scn基因编码的nav蛋白的对应细胞外结构域的外源性scn基因的部分替换。在某些实施方案中,适合用于替换内源性啮齿动物
scn9a基因的外源性scn基因包括上文描述的那些中的任一种,例如,scn1a、scn2a、scn3a、scn4a、scn5a、scn 8a、scn10a或scn11a基因,或来自不同于被修饰的啮齿动物的动物物种的scn9a基因;且在某些实施方案中,所述外源性scn基因是scn2a基因,例如,来自人、黑猩猩、恒河猴、马来西亚飞行狐猴、兔、马、阿拉伯骆驼、杀人鲸、牛、绵羊、大鼠、小鼠、狗、鸡、绿海龟或眼镜王蛇的scn2a基因。在某些实施方案中,所述外源性scn基因是人scn2a基因。
[0184]
在某些实施方案中,在内源性scn9a基因座处的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子可操作地连接至5'转录调节序列(例如,启动子和/或增强子)。在某些实施方案中,在内源性scn9a基因座处的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子可操作地连接至内源性scn9a基因的5'非翻译区(5'utr)。在某些实施方案中,所述核酸分子可操作地连接至外源性scn基因的5'非翻译区(5'utr)。在某些实施方案中,在内源性scn9a基因座处的编码nav蛋白且包含外源性啮齿动物scn基因的至少一部分的核酸分子可操作地连接至内源性scn9a基因的5'utr和5'转录调节序列(例如,启动子和/或增强子)。
[0185]
在某些实施方案中,在内源性scn9a基因座处的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子可操作地连接至内源性scn9a基因的3'调节序列,例如,3'utr。在某些实施方案中,在内源性scn9a基因座处的编码nav蛋白且包含外源性scn基因的至少一部分的核酸分子包含外源性scn基因的3'utr。在某些实施方案中,所述核酸分子包含外源性scn基因的3'utr和在3'utr之外的外源性scn基因的另外基因组序列,例如,30
‑
500bp或更大的基因组序列,所述基因组序列存在于在外源性scn基因的3'utr的下游紧邻处的外源性scn基因基因座中。
[0186]
在某些实施方案中,遗传修饰的啮齿动物就遗传修饰而言是异源的,即,就在内源性scn9a基因座处的包含外源性scn基因的至少一部分的核酸分子而言是异源的。在某些实施方案中,遗传修饰的啮齿动物就遗传修饰而言是纯合的,即,就在内源性scn9a基因座处的包含外源性scn基因的至少一部分的核酸分子而言是纯合的。
[0187]
在某些实施方案中,本文中公开的遗传修饰的啮齿动物不能表达内源性啮齿动物nav1.7蛋白,例如,由于对内源性啮齿动物scn9a基因座的遗传修饰或内源性啮齿动物scn9a基因的灭活(例如,完全或部分缺失)。
[0188]
在某些实施方案中,如本文中所述的遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组中包含(例如,通过杂交育种或多基因靶向策略):(i)人源化的免疫球蛋白重链基因座,其包含一个或多个人v
h
基因区段、一个或多个人d
h
基因区段和一个或多个人j
h
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的hoh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座处是杂合的。
[0189]
在某些实施方案中,包含人源化的hoh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含重链,其中每个重链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人重链可变结构域。
[0190]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的loh基因座”。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座处是杂合的。
[0191]
在某些实施方案中,包含人源化的loh基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含免疫球蛋白链,其中每个免疫球蛋白链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人轻链可变结构域。
[0192]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座(例如,κ和/或λ),所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个免疫球蛋白轻链恒定区基因的上游(例如,与其可操作地连接)。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vκ基因区段和一个或多个人jκ基因区段。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vλ基因区段和一个或多个人jλ基因区段。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cκ。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cλ。
[0193]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人vκ基因区段和一个或多个人jκ基因区段,所述区段是在cκ基因的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的kok基因座”。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的kok基因座处是杂合的。
[0194]
在某些实施方案中,包含人源化的kok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含κ轻链,其中每个κ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人κ轻链可变结构域。
[0195]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lol基因座”。在某
些实施方案中,人源化的lol基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因和一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个小鼠cλ基因包含小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lol基因座处是杂合的。
[0196]
在某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。在某些实施方案中,包含人源化的lol基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
[0197]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在cκ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lok基因座”。在某些实施方案中,人源化的lok基因座的cκ基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lok基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lok基因座处是杂合的。
[0198]
在某些实施方案中,包含人源化的lok基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含轻链,其中每个轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人λ轻链可变结构域。
[0199]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lik基因座”。在某些实施方案中,人源化的lik基因座的cλ基因是啮齿动物(例如,大鼠或小鼠)cλ基因。在某些实施方案中,人源化的lik基因座的cλ基因是小鼠cλ1基因。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的lik基因座处是杂合的。
[0200]
在某些实施方案中,包含人源化的lik基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。
[0201]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,
其种系基因组)中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个人cλ基因上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。在某些实施方案中,这样的人源化的免疫球蛋白κ轻链基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是纯合的。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)就这样的人源化的免疫球蛋白κ轻链基因座而言是杂合的。在某些实施方案中,包含这样的人源化的免疫球蛋白κ轻链基因座的遗传修饰的啮齿动物(例如,大鼠或小鼠)产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
[0202]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0203]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
[0204]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0205]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0206]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0207]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
[0208]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0209]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在其基因组(例如,其种系基因组)中包含人源化的loh基因座和人源化的lik基因座。在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0210]
在某些实施方案中,本文描述的啮齿动物(例如,大鼠或小鼠)是如在例如美国专利号8,502,018、8,642,835、8,697,940、8,791,323、9,226,484和wo2019/113065中所述,它们都通过引用整体并入本文。可以按照本领域容易获得的方案进行育种(或“杂交”或“杂交育种”);参见,例如,jove science education database.lab animal research,fundamentals of breeding and weaning,jove,cambridge,ma,(2018)(视频文章);breeding strategies for maintaining colonies of laboratory mice,a jackson laboratory resource manual,2007the jackson laboratory;都通过引用并入本文。可替换地,可以将经工程改造的igλ轻链基因座工程改造进包含人源化的igh基因座和/或人源化的igκ基因座的es细胞中,并将得到的es细胞用于产生啮齿动物,或可以使包含人源化的igλ轻链基因座的啮齿动物与包含人源化的igh基因座和/或人源化的igκ基因座的另一种啮齿动物繁殖。包含人源化的igh基因座和/或人源化的igκ基因座的各种啮齿动物是已知的,例如,品系(参见,例如,美国专利号8,502,018和/或8,642,835;通过引用整体并入本文)、xenomouse
tm
品系(参见,例如,mendez,m.j.等人,1997,nat.genetics 15(2):146
‑
56和jakobovits,a.等人,1995,ann.ny acad.sci.764:525
‑
35,通过引用整体并入)。
[0211]
在某些实施方案中,本文描述的啮齿动物包含在以下文献中描述的有限免疫球蛋白轻链基因座:美国专利号9,796,788、9,969,814;美国专利申请公开号2011/0195454a1、2012/0021409a1、2012/0192300a1、2013/0045492a1、2013/0185821a1、2013/0302836a1;国际专利申请公开号wo 2011/097603、wo 2012/148873、wo 2013/134263、wo 2013/184761、wo 2014/160179、wo 2014/160202;它们中的每一篇特此通过引用整体并入。在某些实施方案中,本文描述的啮齿动物包含在wo2019/113065、wo2017214089、us20180125043和美国专利号9,035,128、9,066,502、9,163,092、9,150,662、9,334,333、9,006,511、9,029,628、9,206,261、9,012,717、9,394,373、9,206,262、9,206,263、9,226,484、9,540,452和9,399,683中描述的免疫球蛋白轻链基因座。
[0212]
在某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座缺少功能性内源性啮齿动物adam6基因。在某些实施方案中,本文提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)表达一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或
功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段,所述核苷酸序列被包括在与人源化的免疫球蛋白重链(例如,hoh或loh)基因座相同的染色体上。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座的基因组(例如,种系基因组),所述基因座包含一个或多个核苷酸序列,所述核苷酸序列编码一种或多种啮齿动物adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有替代人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。在某些实施方案中,所提供的遗传修饰的啮齿动物(例如,大鼠或小鼠)具有替换人adam6假基因的包含一个或多个核苷酸序列的基因组(例如,种系基因组),所述核苷酸序列编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段。
[0213]
在某些实施方案中,所提供的遗传修饰的啮齿动物具有基因组(例如,种系基因组),其含有包含第一和第二人v
h
基因区段的一个或多个人v
h
基因区段、以及在第一人v
h
基因区段和第二人v
h
基因区段之间的编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列。在某些实施方案中,第一人v
h
基因区段是v
h1‑
2且第二人v
h
基因区段是v
h6‑
1。
[0214]
在某些实施方案中,编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其功能性直系同源物、功能性同系物或功能片段的一个或多个核苷酸序列是在人v
h
基因区段和人d
h
基因区段之间。
[0215]
在某些实施方案中,编码一种或多种啮齿动物adam6多肽的一个或多个核苷酸序列恢复或增强雄性啮齿动物的能育性。
[0216]
在某些实施方案中,本文描述的啮齿动物包含如在美国专利号8,642,835、9,932,408、8,687,940和9,944,716中所述的adam6基因。在某些实施方案中,所述重链基因座包含功能性的,例如,adam6a基因,adam6b基因,或二者。在某些实施方案中,遗传修饰的非人动物的基因组进一步包含功能性的,例如,adam6a基因,adam6b基因,或二者,其不位于小鼠重链可变区基因区段之间。在美国专利号8,642,835和8,697,940中描述了表达adam6a和/或adam6b的示例性啮齿动物,它们中的每一篇特此通过引用整体并入。
[0217]
在某些实施方案中,遗传修饰的啮齿动物(例如,大鼠或小鼠)包含外源性末端脱氧核苷酸转移酶(tdt)基因。在某些实施方案中,与没有外源性tdt基因的啮齿动物相比,包含外源性末端脱氧核苷酸转移酶(tdt)基因的啮齿动物(例如,大鼠或小鼠)可以具有增加的抗原受体多样性。
[0218]
在某些实施方案中,如本文中所述的啮齿动物具有包含可操作地连接至转录控制元件的外源性末端脱氧核苷酸基转移酶(tdt)基因的基因组。
[0219]
在某些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任意组合。
[0220]
在某些实施方案中,外源性tdt位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
[0221]
在某些实施方案中,tdt是人tdt。在某些实施方案中,tdt是tdt的短异形体(tdts)。
[0222]
在某些实施方案中,本公开内容的啮齿动物包括,例如,小鼠、大鼠和仓鼠。在某些实施方案中,啮齿动物选自鼠总科(muroidea)。在某些实施方案中,本公开内容的啮齿动物来自选自丽仓鼠科(calomyscidae)(例如,小鼠
‑
样仓鼠)、仓鼠科(cricetidae)(例如,仓鼠、新世界大鼠和小鼠、田鼠)、鼠科(muridae)(例如,真小鼠和大鼠、沙鼠、刺毛鼠、冠鼠)、马岛鼠科(nesomyidae)(攀鼠、岩鼠、白尾大鼠、马达加斯加大鼠和小鼠)、刺睡鼠科(platacanthomyidae)(例如,多刺睡鼠)和鼹形鼠科(spalacidae)(例如,鼹鼠、竹鼠和鼢鼠)的科。在某些实施方案中,本公开内容的啮齿动物选自真小鼠或大鼠(鼠科)、沙鼠、刺毛鼠和冠鼠。在某些实施方案中,本公开内容的小鼠来自鼠科的成员。
[0223]
遗传修饰的啮齿动物的表型
[0224]
在某些实施方案中,遗传修饰的啮齿动物(其基因组包含在内源性scn9a基因座处的核酸分子,其中所述核酸分子能够编码包含外源性scn基因的至少一部分的nav蛋白)在遗传修饰的啮齿动物中表达nav蛋白。在某些实施方案中,以与对照啮齿动物(即在内源性scn9a基因座处没有遗传修饰的啮齿动物)中的啮齿动物nav1.7蛋白相当或基本相同的模式表达nav蛋白。已知啮齿动物nav1.7蛋白在背根神经节的伤害性(疼痛)神经元、交感神经元和神经内分泌细胞中表达。在某些实施方案中,以与对照啮齿动物(即在内源性scn9a基因座处没有遗传修饰的啮齿动物)中的啮齿动物nav1.7蛋白相当或基本相同的水平表达nav蛋白。术语“相当”是指,被对比的模式或水平可能彼此不同,但允许相互对比,以便根据观察到的差异或相似之处合理得出结论;并且提及水平的术语“基本相同”是指,被对比的水平彼此相差不超过20%。
[0225]
在某些实施方案中,遗传修饰的啮齿动物在用nav1.7免疫原(例如,人nav1.7免疫原)免疫接种能够提高体液免疫应答。nav1.7免疫原可以是蛋白免疫原、dna免疫原或它们的组合。基于血清中对nav1.7蛋白特异性的抗体的滴度,可以确定啮齿动物中的体液免疫应答。可以采用多种测定法来确定抗体滴度,包括基于elisa和流式细胞计量术的测定法(参见,例如,david h.margulies,induction of immune responses,current protocols in immunology,89,1,(2.0.1
‑
2.0.3)(2010);henri v.van der heyde等人,“analysis of antigen
‑
specific antibodies and their isotypes in experimental malaria,”cytometry,第71a(4)卷:242
‑
250(2007);二者通过引用并入本文。在某些实施方案中,测定利用在细胞表面上表达或被工程改造成表达nav1.7的细胞,并且抗体滴度可通过测量与细胞结合的抗体来确定。在某些实施方案中,所述细胞是被工程改造成表达人nav1.7蛋白的hek细胞。在某些实施方案中,将抗体滴度定义为内插的血清稀释因子,其结合信号是背景的2倍。在某些实施方案中,与对照啮齿动物(即,没有遗传修饰的啮齿动物,即,不具有插入在内源性啮齿动物scn9a基因座处的外源性scn基因的至少一部分)相比,本文所公开的遗传修饰的啮齿动物以至少5、10、20、30、40、50、60、70、80、90或100倍或更大的滴度产生针对nav1.7(例如,人nav1.7)的抗体。
[0226]
在某些实施方案中,啮齿动物能够产生对nav1.7蛋白(例如,人nav1.7蛋白)特异
性的抗体。在某些实施方案中,基于抗体与被工程改造成表达nav1.7蛋白的细胞系的结合相对于与没有nav1.7蛋白的工程改造表达的亲本细胞系的结合之比,确定抗体特异性。在某些实施方案中,在至少2、3、4、5、6、7、8、9、10、15、20的比率或大于20的比率,抗体具有对nav1.7(例如,但不限于,人nav1.7)的特异性。
[0227]
在某些实施方案中,本文所公开的遗传修饰的啮齿动物表现出对热刺激的受损应答。可以测量对热刺激的应答,例如,在hargreaves试验(一种试验,其测量啮齿动物对指向后爪的辐射热刺激的缩回潜伏期;参见,例如,shields等人,journal of neuroscience,2018,38(47):10180
–
10201)中,或在设置在有害温度(例如,52.5℃或55℃)的热板设备中(参见,例如,shields等人.2018,出处同上)。在某些实施方案中,本文所公开的遗传修饰的啮齿动物(诸如向mnav1.7 ko小鼠中的hnav1.2 ki)表现出与野生型啮齿动物相比延长的对热刺激(例如,在hargreaves试验中的辐射热刺激)的应答潜伏期,例如,遗传修饰的啮齿动物需要与野生型啮齿动物相比长至少25%、50%、75%或100%的时间来响应热刺激。
[0228]
在某些实施方案中,本文所公开的遗传修饰的啮齿动物表现出减少的对组胺的痒应答。通过在啮齿动物的颈背皮内注射组胺并测量指定时间段内的搔抓发作次数,可以确定痒应答(参见,例如,参见,例如,shields等人,2018,出处同上)。在某些实施方案中,本文所公开的遗传修饰的啮齿动物(诸如在mnav1.7小鼠中的hnav1.2 ki)通过在一段时间(诸如15分钟、20分钟、25分钟或30分钟)内比野生型啮齿动物少至少25%、50%、75%或100%的搔抓发作次数,表现出减少的对组胺的痒应答。
[0229]
遗传修饰的啮齿动物组织和细胞
[0230]
在一些实施方案的另一个方面,本文中公开了分离的啮齿动物细胞或组织,其包含如本文中所述的在内源性啮齿动物scn9a基因座处的遗传修饰。
[0231]
在某些实施方案中,啮齿动物组织是脂肪、膀胱、脑、乳房、骨髓、眼、心脏、肠、肾、肝、肺、淋巴结、肌肉、胰腺、血浆、血清、皮肤、脾、胃、胸腺、睾丸、卵子或它们的组合。
[0232]
在某些实施方案中,啮齿动物细胞是淋巴细胞。在某些实施方案中,细胞选自b细胞、树突细胞、巨噬细胞、单核细胞和t细胞。
[0233]
在某些实施方案中,将本文描述的遗传修饰的啮齿动物的b细胞用于生产结合nav1.7(例如,人nav1.7)的抗体。例如,可以从本文描述的啮齿动物分离b细胞,并直接用于或永生化用于产生杂交瘤。这样的啮齿动物可以用nav1.7免疫原(dna或蛋白)免疫,然后从啮齿动物分离b细胞。可以关于与表达nav1.7(例如,人nav1.7)的细胞的结合来筛选b细胞和/或杂交瘤。可以从这样的细胞克隆抗体并测序,并用于产生候选治疗剂。
[0234]
在某些实施方案中,提供了从如本文中所述的分离的啮齿动物细胞或啮齿动物组织制备的永生化细胞。来自本文中公开的啮齿动物的细胞可以临时分离和使用,或者可以在培养物中维持多代。在某些实施方案中,将来自本文中公开的啮齿动物的细胞永生化(例如,通过使用病毒、细胞融合等)并在培养物中无限维持(例如,在系列培养中)。
[0235]
在某些实施方案中,提供了啮齿动物胚胎干(es)细胞,其基因组包含如本文中所述的在内源性scn9a基因座处的遗传修饰。啮齿动物es细胞可以用于制备啮齿动物胚胎和啮齿动物。
[0236]
在某些实施方案中,啮齿动物es细胞是小鼠胚胎干细胞,且在某些实施方案中来自129品系、c57bl品系、或其混合物。在某些实施方案中,啮齿动物es细胞是小鼠胚胎干细
胞且是129和c57bl品系的混合物。在某些实施方案中,啮齿动物es细胞是大鼠胚胎干细胞。
[0237]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白重链基因座,所述基因座包含一个或多个人v
h
基因区段、一个或多个人d
h
基因区段和一个或多个人j
h
基因区段,所述区段是在一个或多个啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因(例如,一个或多个内源性啮齿动物(例如,大鼠或小鼠)免疫球蛋白重链恒定区基因)的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的hoh基因座”。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座处是杂合的。
[0238]
在某些实施方案中,分离的啮齿动物细胞是包含人源化的hoh基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含重链,其中每个重链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)重链恒定结构域的人重链可变结构域。
[0239]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人v
l
基因区段和一个或多个人j
l
基因区段,所述区段是在一个或多个免疫球蛋白轻链恒定区基因的上游(例如,与其可操作地连接)。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vκ基因区段和一个或多个人jκ基因区段。在某些实施方案中,一个或多个人v
l
基因区段和一个或多个人j
l
基因区段是一个或多个人vλ基因区段和一个或多个人jλ基因区段。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cκ。在某些实施方案中,一个或多个免疫球蛋白轻链恒定区基因是或包含cλ。
[0240]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含一个或多个人vκ基因区段和一个或多个人jκ基因区段,所述区段是在cκ基因的上游(例如,与其可操作地连接)。这样的人源化的免疫球蛋白重链基因座在本文中被称作“人源化的kok基因座”。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的kok基因座的免疫球蛋白κ轻链恒定区基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的kok基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的kok基因座处是杂合的。
[0241]
在某些实施方案中,分离的啮齿动物细胞是包含人源化的kok基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含κ轻链,其中每个κ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人κ轻链可变结构域。
[0242]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lol基因座”。在某些实施方案中,人源化的lol基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因。在某些实施方
案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个cλ基因包含一个或多个人cλ基因和一个或多个小鼠cλ基因。在某些实施方案中,人源化的lol基因座的一个或多个小鼠cλ基因包含小鼠cλ1基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lol基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lol基因座处是杂合的。
[0243]
在某些实施方案中,分离的啮齿动物细胞是包含人源化的lol基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。在某些实施方案中,分离的啮齿动物细胞是包含人源化的lol基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
[0244]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白轻链基因座,所述基因座包含在cκ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lok基因座”。在某些实施方案中,人源化的lok基因座的cκ基因是啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,人源化的lok基因座的cκ基因是在内源性免疫球蛋白κ轻链基因座处的内源性啮齿动物(例如,大鼠或小鼠)cκ基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lok基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lok基因座处是杂合的。
[0245]
在某些实施方案中,分离的啮齿动物细胞是包含人源化的lok基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含轻链,其中每个轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)κ轻链恒定结构域的人λ轻链可变结构域。
[0246]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在cλ基因的上游(例如,与其可操作地连接)的一个或多个人vλ基因区段和一个或多个人jλ基因区段。这样的人源化的免疫球蛋白轻链基因座在本文中被称作“人源化的lik基因座”。在某些实施方案中,人源化的lik基因座的cλ基因是啮齿动物(例如,大鼠或小鼠)cλ基因。在某些实施方案中,人源化的lik基因座的cλ基因是小鼠cλ1基因。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lik基因座处是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的lik基因座处是杂合的。
[0247]
在某些实施方案中,分离的啮齿动物细胞是包含人源化的lik基因座的b细胞或脾细胞且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至啮齿动物(例如,大鼠或小鼠)λ轻链恒定结构域的人λ轻链可变结构域。
[0248]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的免疫球蛋白κ轻链基因座,所述基因座包含在一个或多个人jλ基因区段和一个或多个人cλ基因上游(例如,与其可操作地连接)的一个或多个人vλ基因区段。在某些实施方案中,
这样的人源化的免疫球蛋白κ轻链基因座的一个或多个人jλ基因区段和一个或多个cλ基因存在于jλ
‑
cλ簇中。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织就这样的人源化的免疫球蛋白κ轻链基因座而言是纯合的。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织就这样的人源化的免疫球蛋白κ轻链基因座而言是杂合的。在某些实施方案中,分离的啮齿动物细胞是b细胞或脾细胞,其包含这样的人源化的免疫球蛋白κ轻链基因座,且产生抗体,例如,响应于抗原刺激,所述抗体尤其包含λ轻链,其中每个λ轻链包含可操作地连接至人λ轻链恒定结构域的人λ轻链可变结构域。
[0249]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0250]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
[0251]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0252]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0253]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0254]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
[0255]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0256]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在其基因组中包含人源化的loh基因座和人源化的lik基因座。在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0257]
在某些实施方案中,本文提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含缺乏内源性啮齿动物adam6基因的人源化的免疫球蛋白重链(例如,hoh或loh)基因座。在某些实施方案中,本文提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含编码一种或多种啮齿动物adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织表达一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列,所述核苷酸序列被包括在与人源化的免疫球蛋白重链(例如,hoh或loh)基因座相同的染色体上。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含人源化的免疫球蛋白重链(例如,hoh或loh)基因座,所述基因座包含编码一种或多种啮齿动物adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有替代人adam6假基因的基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有替换人adam6假基因的基因组,所述基因组包含编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。
[0258]
在某些实施方案中,所提供的分离的啮齿动物细胞或啮齿动物组织具有基因组,所述基因组包含含有第一和第二人v
h
基因区段的一个或多个人v
h
基因区段、以及在第一人v
h
基因区段和第二人v
h
基因区段之间的编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列。在某些实施方案中,第一人v
h
基因区段是v
h1‑
2且第二人v
h
基因区段是v
h6‑
1。
[0259]
在某些实施方案中,编码一种或多种啮齿动物(例如,大鼠或小鼠)adam6多肽、其直系同源物、同系物或片段的一个或多个核苷酸序列是在人v
h
基因区段和人d
h
基因区段之间。
[0260]
在某些实施方案中,分离的啮齿动物细胞或啮齿动物组织包含外源性末端脱氧核苷酸转移酶(tdt)基因。在某些实施方案中,分离的啮齿动物细胞是包含外源性末端脱氧核苷酸转移酶(tdt)基因的b细胞或脾细胞,且当与不具有外源性tdt基因的分离的啮齿动物细胞(例如,b细胞或脾细胞)相比时可以具有增加的抗原受体多样性。
[0261]
在某些实施方案中,如本文中所述的分离的啮齿动物细胞或啮齿动物组织具有包含可操作地连接至转录控制元件的外源性末端脱氧核苷酸基转移酶(tdt)基因的基因组。
[0262]
在某些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任意组合。
[0263]
在某些实施方案中,外源性tdt位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
[0264]
在某些实施方案中,tdt是人tdt。在某些实施方案中,tdt是tdt的短异形体(tdts)。
[0265]
用于制备遗传修饰的啮齿动物的组合物和方法
[0266]
在一些实施方案的一个方面,本文公开了用于制备上述遗传修饰的啮齿动物的方法,以及适合用于制备遗传修饰的啮齿动物的核酸载体。
[0267]
在某些实施方案中,本文中公开了靶向载体(或核酸构建体),其包含期望整合进啮齿动物scn9a基因座中的外源性scn基因。在某些实施方案中,本文中公开了靶向载体(或核酸构建体),其包含期望整合进啮齿动物scn9a基因座中的外源性scn基因的至少一部分。在某些实施方案中,靶载体包含外源性scn基因的一部分,该部分编码由外源性scn基因编码的nav蛋白的细胞外结构域的连续氨基酸。在某些实施方案中,靶载体包含外源性scn基因的部分,所述部分编码由外源性scn基因编码的nav蛋白的细胞外结构域的氨基酸,并且也包含内源性scn9a基因的部分,所述部分编码跨膜和细胞质结构域的氨基酸,其中跨膜和细胞质结构域彼此可操作地连接。在某些实施方案中,靶载体包含外源性scn基因的编码序列。在某些实施方案中,所述靶向载体也包括侧接要整合进啮齿动物scn9a基因座中的核苷酸序列的5'和3'啮齿动物序列(也被称作同源性臂),其介导所述核苷酸序列向靶啮齿动物scn9a基因座中的同源重组和整合。在某些实施方案中,所述同源性臂包含核苷酸序列,其侧接在要被替换的靶啮齿动物基因座处的核苷酸序列。在一个示例性实施方案中,来自内源性啮齿动物scn9a基因的从起始密码子至终止密码子的编码序列被人scn2a基因的编码序列替换,5'侧翼序列可以包括在内源性啮齿动物scn9a基因的atg密码子上游的序列,且3'侧翼序列可以包括在内源性啮齿动物scn9a基因的终止密码子下游的序列。
[0268]
在某些实施方案中,靶向载体包含选择标记基因。在某些实施方案中,靶向载体包含一个或多个位点特异性的重组位点。在某些实施方案中,靶向载体包含选择标记基因,其侧接位点特异性的重组位点,使得选择标记基因可以由于位点直接的重组而被删除。
[0269]
在示例性实施方案中,使用细菌同源重组和技术(参见,例如,美国6,586,251和valenzuela等人(2003)nature biotech.21(6):652
‑
659;都通过引用整体并入本文),可以修饰携带啮齿动物scn9a基因的啮齿动物基因组片段的细菌人工染色体(bac)克隆。所以,从原始bac克隆删除啮齿动物scn9a基因组序列,并插入外源性scn核苷酸序列,从而产生经修饰的携带外源性scn核苷酸序列的bac克隆,其侧接5'和3'啮齿动物同源性臂。经修饰的bac克隆一旦线性化就可以引入啮齿动物胚胎干(es)细胞中。
[0270]
在某些实施方案中,本发明提供了如本文中所述的靶向载体用于制备经修饰的啮齿动物胚胎干(es)细胞的用途。例如,可以将靶向载体引入啮齿动物es细胞,例如,通过电穿孔。在本领域中已经描述了小鼠es细胞和大鼠es细胞。参见,例如,us 7,576,259、us 7,659,442、us 7,294,754和us 2008
‑
0078000 a1,它们描述了小鼠es细胞和用于制备遗传修饰的小鼠的方法;us 2014/0235933 a1(regeneron pharmaceuticals inc.),us 2014/0310828 a1(regeneron pharmaceuticals inc.),tong等人(2010)nature 467:211
‑
215,和tong等人(2011)nat protoc.6(6):doi:10.1038/nprot.2011.338,它们描述了大鼠es细胞和用于制备遗传上修饰的大鼠的方法,它们可以用于制备经修饰的啮齿动物胚胎,所述胚胎又可以用于制备啮齿动物。
[0271]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座和人源化的kok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座和人源化的lol基因座。在某些实施方案中,
本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的hoh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0272]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座、人源化的kok基因座和人源化的lik基因座。
[0273]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座和人源化的lok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的hoh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0274]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的hoh基因座和人源化的lik基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的hoh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0275]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座和人源化的kok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座和人源化的lol基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lol基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的loh基因座、人源化的kok基因座、人源化的lol基因座或它们的组合处是纯合的。
[0276]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座、人源化的kok基因座和人源化的lik基因座。
[0277]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座和人源化的lok基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的loh基因座、人源化的lok基因座或它们的组合处是纯合的。
[0278]
在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在其基因组中包含人源化的loh基因座和人源化的lik基因座。在某些实施方案中,本文描述的经修饰的啮齿动物es细胞在人源化的loh基因座、人源化的lik基因座或它们的组合处是纯合的。
[0279]
在某些实施方案中,可以选择具有整合进基因组中的外源性scn核苷酸序列的es细胞。在某些实施方案中,基于啮齿动物等位基因的缺失和/或外源性核苷酸序列测定的获得,选择es细胞。在某些实施方案中,然后通过使用方法(参见,例如,us 7,576,259、us 7,659,442、us 7,294,754和us 2008
‑
0078000 a1),或在us 2014/0235933 a1和us 2014/0310828 a1中描述的方法,将选择的es细胞用作供体es细胞以注射进桑椹胚前阶段胚胎(例如,8
‑
细胞阶段胚胎)中。在某些实施方案中,经修饰的啮齿动物es细胞包含人源化的免疫球蛋白重链和/或轻链基因座和外源性scn序列,且经修饰的啮齿动物es细胞可以引入胚胎中。将包含供体es细胞的胚胎温育直到胚泡期,并然后植入代孕母体以产生完全衍生自供体es细胞的f0啮齿动物。使用啮齿动物scn9a序列的缺失和/或外源性scn序列测定的获得,通过分离自剪断的尾巴的dna的基因分型,可以鉴定带有外源性scn
核苷酸序列的啮齿动物幼崽。
[0280]
在某些实施方案中,可以将在内源性scn9a基因座处的遗传修饰杂合的啮齿动物杂交(或杂交育种)以产生纯合的啮齿动物,例如,通过遵循本领域中容易获得的育种方案;参见,例如,jove science education database.lab animal research,fundamentals of breeding and weaning,jove,cambridge,ma,(2018)(视频文章);breeding strategies for maintaining colonies of laboratory mice,a jackson laboratory resource manual,2007the jackson laboratory;都通过引用整体并入本文。
[0281]
在某些实施方案中,包含在内源性scn9a基因座处的遗传修饰的啮齿动物可以与包含人或人源化的免疫球蛋白重链和/或轻链基因座的啮齿动物杂交以获得包含如上所述的在内源性scn9a基因座处的遗传修饰和人或人源化的免疫球蛋白重链和/或轻链基因座的啮齿动物。在某些实施方案中,包含人源化的免疫球蛋白重链和/或轻链基因座的啮齿动物es细胞可用于接受外源性scn序列以整合到内源性scn9a基因座中,并且所得到的遗传修饰的啮齿动物es细胞可用于制备遗传修饰的啮齿动物,其包含在内源性scn9a基因座处的遗传修饰和人源化的免疫球蛋白重链和/或轻链基因座。
[0282]
采用遗传修饰的啮齿动物的方法
[0283]
在某些实施方案中,本文描述的遗传修饰的啮齿动物用于产生针对nav1.7蛋白(例如,人nav1.7蛋白)的抗体。
[0284]
在某些实施方案中,通过经由各个途径(例如,但不限于,静脉内或腹膜内途径)将nav1.7免疫原(例如,人nav1.7)施用给本文描述的啮齿动物,可以产生抗体。nav1.7免疫原是蛋白免疫原(即,nav1.7蛋白或其片段)、dna免疫原(能够在接受者啮齿动物中表达nav1.7蛋白或其片段的dna,例如,病毒载体)或它们的组合。在某些实施方案中,所述免疫原是在大肠杆菌中或在真核(例如,酵母)或哺乳动物细胞(例如,中国仓鼠卵巢(cho)细胞)中表达的重组nav1.7蛋白。在某些实施方案中,使用标准佐剂可以施用一次或多次加强注射。加强注射可以使用相同的nav1.7免疫原,或从原始蛋白免疫原切换到dna免疫原,或反之亦然。从经免疫的啮齿动物回收淋巴细胞(诸如b
‑
细胞),并可以直接筛选,或可以与骨髓瘤细胞系融合以制备永生化的杂交瘤细胞系,然后将其筛选以鉴定产生对nav1.7特异性的抗体的细胞。筛选可以是基于相对于与不表达nav1.7蛋白的亲代细胞(hek细胞)的结合,评价候选抗体与被工程改造成表达nav1.7蛋白的细胞(例如,被工程改造成表达人nav1.7的hek细胞)的结合。在某些实施方案中,在指定的抗体浓度与被工程改造成表达nav1.7蛋白的细胞的结合相对于与不表达nav1.7蛋白的亲代细胞的结合的比率被用于测量抗体的特异性,并且如果所述比率是至少2、3、4、5、6、7、8、9或10或大于10的比率,可以将抗体鉴定为nav1.7蛋白的特异性结合剂。
[0285]
可以分离编码所鉴定的细胞的重链和轻链的可变区的dna,并将其连接到合乎需要的重和轻恒定区。可以在细胞、诸如cho细胞中生产这样的抗体蛋白。
[0286]
本说明书通过以下实施例进一步举例说明,不应将其解释为以任何方式进行限制。所有引用的参考文献(包括贯穿本技术引用的参考文献、授权专利和公开的专利申请)的内容特此明确地通过引用并入。
[0287]
实施例
[0288]
实施例1.包含在小鼠nav1.7敲出(ko)中的人nav1.2敲入(ki)的小鼠品系的制备
[0289]
本实施例解释了制备遗传修饰的啮齿动物(例如,小鼠)的示例性方法,其中啮齿动物scn基因(例如,小鼠scn9a基因,其编码小鼠nav1.7蛋白)被来自不同物种的scn基因完全地或部分地替换(例如,人scn2a基因,其编码人nav1.2蛋白)。
[0290]
使用细菌人工染色体(bac)克隆和技术(参见,例如,美国专利号6,586,251和valenzuela等人(2003)high
‑
throughput engineering of the mouse genome coupled with high
‑
resolution expression analysis,nature biotech.21(6):652
‑
659,其通过引用整体并入本文),如下构建用于修饰内源性小鼠scn9a基因的靶向载体。
[0291]
简而言之,使用含有来自bac克隆rp11
‑
422d18的96,735bp的人scn2a基因组dna和4809bp的自删除新霉素盒的dna片段(loxp
‑
mprm1
‑
crei
‑
pa
‑
hub1
‑
em7
‑
neo
‑
pa
‑
loxp),通过细菌细胞中的同源重组来修饰含有小鼠scn9a基因的细菌人工染色体(bac)克隆rp23
‑
454h3。人scn2a基因组dna含有人scn2a atg至超出终止密码子的2734bp,其包括在所述盒前面紧挨着的人3'utr之后的约250bp的3'人序列。作为同源重组的结果,在bac克隆rp23
‑
454h3中的84,847bp的小鼠核苷酸序列(从atg起始密码子至小鼠scn9a基因的终止密码子)被96,735bp的人序列替换,随后是所述盒。得到的经修饰的bac克隆(具有57kb的5'同源性臂和43kb的3'同源性臂,后者侧接人scn2a基因组dna和自删除盒)用作靶向载体以修饰内源性小鼠scn9a基因。参见图1a
‑
1b。
[0292]
通过电穿孔将经修饰的bac克隆引入小鼠胚胎干(es)细胞。通过测定(valenzuela等人,出处同上)来鉴定含有替代在内源性小鼠scn9a基因座处的小鼠scn9a基因的人scn2a基因(“在小鼠scn9a ko中的人scn2a ki”或“在mnav1.7 ko中的hnav1.2 ki”)的正靶向的es细胞,所述测定检测人序列的存在并证实小鼠序列的缺失和/或保留。表5描述了在测定中使用的引物和探针。也参见图1a,该图描绘了在测定中使用的引物和探针的位置。成功修饰的scn9a基因座的核苷酸序列如seq id no:20所示。在选择具有期望修饰的靶向的es细胞克隆以后,通过引入cre重组酶,例如,通过电穿孔,可以切离新霉素选择盒。可替换地,通过使从es克隆产生的后代与表达cre重组酶的缺失啮齿动物品系杂交,可以除去新霉素选择盒。在删除盒以后的经修饰的scn9a基因座如图1c所示,其中连接部序列显示在图1c的底部。
[0293]
表5
[0294][0295]
使用方法(参见,例如,美国专利号7,294,754和poueymirou等人,2007,nature biotech.25(1):91
‑
99)将选择的es细胞克隆(具有或没有盒)用于植入雌性小鼠以产生一窝在基因组中含有人源化的scn9a基因座的幼崽。使用检测人序列的存在的等位基因测定的改进(valenzuela等人,出处同上),通过分离自剪断的尾巴的dna的基因分型,再次证实和鉴定带有这样的遗传修饰的小鼠。通过使杂合的动物杂交,制备就人源化的scn9a基因座而言纯合的动物。
[0296]
实施例2.在mnav1.7敲出(ko)中的hnav1.2敲入(ki)/vi
‑
3小鼠的免疫接种和对免疫原的血清抗体应答的分析。
[0297]
免疫接种.将在mnav1.7 ko中的人nav1.2敲入(ki)/vi
‑
3小鼠用编码全长人nav1.7蛋白的全长dna或人nav1.7蛋白免疫。使用标准的佐剂,在不同的时间间隔经由不同的途径强化小鼠。在免疫接种开始之前和在免疫原强化后定期给小鼠抽血,并在各种抗原上测定抗
‑
血清滴度。
[0298]
抗
‑
血清滴度测定.使用meso scale discovery(msd)细胞结合elisa测定血清中针对各种免疫原的抗体滴度。在37℃给九十六
‑
孔碳表面平板涂布在pbs中的40,000个细胞/孔的hek293/hnav1.7
‑
gfp(来自sanofi,sa)、hek293/hnav1.7(millipore)和hek293亲代细胞1小时。倾析细胞涂布溶液,并将平板用150μl的在pbs中的2%牛血清白蛋白(bsa,sigma
‑
aldrich)在室温(rt)封闭1h。使用洗板机(来自molecular devices的2000)将平板用pbs洗涤三次。将免疫前和免疫抗
‑
血清在1%bsa
‑
pbs中连续稀释三倍,并加入平板在室温保持1h。将平板洗涤,然后将山羊抗
‑
小鼠igg
‑
fc钌缀合的第二抗体以1μg/ml加入平板并在室温温育1小时。将平板洗涤,并通过加入150μl/孔的msd的4x无表面活性剂的read buffer t(稀释至1x)进行显影,并在msd sector
tm
成像仪6000仪器上读出。使用graphpad prism软件计算抗
‑
血清滴度。将滴度定义为内插的血清稀释因子,其结合信号是
背景的2倍。
[0299]
结果.在用蛋白或dna免疫原免疫接种以后,研究了在mnav1.7 ko中的hnav1.2 ki/vi
‑
3小鼠中的体液免疫应答。来自用蛋白免疫的小鼠的抗血清在nav1.7过表达细胞上显示出高的特异性滴度,与亲代细胞的结合较低(图2)。对最初用dna免疫的低应答小鼠施用蛋白强化,这导致在nav1.7工程改造的细胞上引发高特异性滴度(图2)。
[0300]
实施例3.来自在mnav1.7 ko中的人nav1.2 ki/vi小鼠的抗体的电化学发光细胞结合
‑
来自初步筛选的上清液和纯化的抗体。
[0301]
实验规程
[0302]
将在实施例1中描述的在mnav1.7 ko中的人nav1.2 ki/vi小鼠用纯化的去污剂增溶的人nav1.7蛋白免疫。通过来自这些免疫的小鼠的脾细胞与小鼠骨髓瘤p3x63ag8.653细胞的融合,制备单克隆抗体。使用基于电化学发光(ecl)的检测,关于它们的结合人nav1.7表达细胞的能力,评价来自杂交瘤的上清液。通过在nav1.7工程改造的细胞和参考细胞系上的结合的对比,关于特异性来评价正nav1.7细胞结合剂。24个nav1.7
‑
特异性的杂交瘤的子集是通过流式细胞计量术筛分的单个细胞,将其扩增,并纯化抗体。确定这些抗体的特异性地结合被工程改造成表达nav1.7的细胞的能力。
[0303]
简而言之,被工程改造成表达人nav1.7的人胚胎肾细胞(hek293)得自两个来源:sanofi(sa 293/gfp
‑
hnav1.7,缩写“sa”,如在图2中所示)和millipore(millipore 293/hnav1.7,缩写“millipore”,如在图2中所示)。来自atcc的hek293细胞用作nav1.7基线参考,因为它们具有低水平的nav1.7 mrna,如通过taqman分析所确定的。先前分离的抗
‑
人nav1.7抗体用作nav1.7阳性细胞结合对照。无关的小鼠igg抗体(抗
‑
hcd48 migg1或抗
‑
higg4 migg2a对照)用作测定中的阴性结合对照。
[0304]
根据下述规程进行实验。将来自上述系的细胞在不含ca
2
/mg
2
的1xpbs缓冲液中冲洗一次,并在37℃与无酶细胞解离溶液(enzyme free cell dissociation solution)一起温育10分钟以使细胞脱离烧瓶。将所有细胞用含有ca
2
/mg
2
的1xpbs洗涤一次,并用cellometer
tm auto t4细胞计数器(nexcelom bioscience llc,lawrence,ma)计数。将大约2.0x104个hek293、sa 293/gfp
‑
hnav1.7和millipore 293/hnav1.7细胞分别接种在96
‑
孔碳电极平板(multi
‑
array高结合平板,meso scale discovery(msd,rockville,md))上并在37℃温育1小时。将非特异性结合位点用在含有ca
2
/mg
2
的1xpbs中的2%bsa(w/v)在室温(rt)封闭1小时。向平板结合的hek293、sa 293/gfp
‑
hnav1.7和millipore 293/hnav1.7细胞,以在pbs 0.5%bsa中的1:20的固定稀释度,加入抗
‑
nav1.7上清液或对照抗体的溶液作为单个点。对于纯化的抗体,一式两份地加入从1.7pm至100nm范围内的系列稀释物和不存在抗体的溶液。将平板在室温温育1小时,然后使用具有细胞洗涤头的aquamax2000洗板机(mds analytical technologies,sunnyvale,ca)洗涤以除去未结合的抗体。用对fcγ片段特异性的sulfo
‑
tag
tm
‑
缀合的山羊多克隆抗
‑
人igg抗体(jackson immunoresearch,west grove,pa)在室温检测平板结合的抗体1小时。将平板洗涤,并根据生产商的说明书用read buffer(msd,rockville,md)显影,用sector imager(msd,rockville,md)记录发光信号。记录以相对光单位(rlu)测量的发光强度以指示在浓度范围处的每种抗体的结合强度。
[0305]
对于3080个样品的初筛,将在sa 293/gfp
‑
hnav1.7或millipore 293/hnav1.7细胞系中具有大于300rlu的直接结合信号的上清液评分为阳性。用上述规程在所有三个细胞
系中进一步试验145个阳性样品以确定特异性比。与hek293细胞相比在人nav1.7
‑
表达细胞上具有大于或等于2的结合比的抗体被分类为nav1.7
‑
特异性结合剂,且计数值如表6所示。
[0306]
对于纯化的抗体,在人nav1.7表达细胞上在1.2nm抗体检测到的结合信号相对于结合hek293细胞的相同抗体浓度的比率显示在表7中,并用作nav1.7结合的特异性的指示。在sa 293/gfp
‑
hnav1.7或millipore 293/hnav1.7细胞上具有>150rlu的结合信号并且具有与hek293细胞相比大于或等于2的比率的抗体被归类为nav1.7
‑
特异性结合剂。具有小于2的结合比或<150rlu的结合信号的抗体被归类为非特异性结合剂。
[0307]
结果总结和结论
[0308]
使来自nav1.7免疫的在mnav1.7 ko中的人nav1.2 ki/vi小鼠的三个脾融合以产生杂交瘤。使用电化学发光(ecl)关于人nav1.7细胞结合和特异性评价来自那些细胞的上清液。使抗体结合人nav1.7
‑
表达细胞、293/gfp
‑
hnav1.7或millipore 293/hnav1.7,并且在一些实验中结合参考细胞系hek293,并用sulfo
‑
tag
tm
‑
缀合的抗
‑
小鼠igg多克隆抗体检测。
[0309]
如表6中的结果所示,以1:20稀释度试验了3080个杂交瘤上清液,其中145个结合sa 293/gfp
‑
hnav1.7和/或millipore 293/hnav1.7细胞,通过ecl检测到大于或等于300rlu的信号。所有三种融合体产生了nav1.7阳性细胞结合剂。随后关于与两个nav1.7细胞系以及参考hek293细胞的结合试验了145个上清液。60个阳性杂交瘤特异性地结合nav1.7细胞,与sa 293/gfp
‑
hnav1.7或millipore 293/hnav1.7细胞的结合相对于hek293细胞的比率大于2倍。那60个上清液中的52个特异性地结合两个细胞系。三个融合体中的两个产生了通过ecl确定的nav1.7
‑
特异性的杂交瘤。
[0310]
60个杂交瘤中的24个的子集是通过流式细胞计量术分选的单个细胞,将其扩增,并纯化抗体和在两个结合实验之一中评估nav1.7细胞特异性结合。在表7中,报告了1.2nm抗体与sa 293/gfp
‑
hnav1.7和millipore 293/hnav1.7细胞相对于hek293细胞的结合的比率。24种抗体中的20种与两种nav1.7细胞系特异性结合,对sa 293/gfp
‑
hnav1.7的结合比对hek293细胞的结合高2.4至57.9倍,且对millipore 293/hnav1.7细胞的结合比对hek293细胞的结合高2.5
‑
44.5倍。24种抗体中的四种是非特异性的,具有<150rlu的结合信号,和/或对nav1.7细胞相对于参照细胞小于二的结合比率。先前分离的阳性nav1.7对照抗体对sa 293/gfp
‑
hnav1.7的结合比对hek293细胞的结合高平均高23.5倍,且对millipore 293/hnav1.7细胞的结合比对hek293细胞的结合的平均高17.3倍。同种型对照抗体具有<150rlu的结合,并且如预期的那样对所有细胞具有接近相等的结合。
[0311]
表6.杂交瘤上清液初筛总结
[0312][0313][0314]
表7
[0315][0316][0317]
实施例4
[0318]
收集选定的抗
‑
nav1.7杂交瘤克隆并使用promega16系统分离总rna。
接着,使用smartscribe
tm
逆转录酶(clontech)和对小鼠重链igg1、igg2a、igg2b、igg3和小鼠κ轻链的小鼠恒定区特异性的反向引物以及模板转换寡物smarter ii a寡物(trombetta等人.2014,pmid:24984854,通过引用整体并入本文),进行逆转录以产生含有人可变结构域和小鼠恒定区序列的部分的cdna。使用ampure xp珠子(beckman coulter genomics)纯化cdna和随后的pcr产物。然后使用具有illumina连接序列的对smarter ii a寡物特异性的引物(5
’‑
tcgtcggcagcgtcagatgtgtataagagacag
‑3’
,seq id no:57)和具有illumina连接序列的对小鼠恒定区特异性的反向引物(5
’‑
gtctcgtgggctcggagatgtgtataagagacag
‑3’
,seq id no:58),通过pcr扩增纯化的cdna。使用具有用于多路测序的索引序列的引物,通过pcr进一步扩增片段。将pcr产物纯化、处理和合并后,通过miseq测序仪(illumina)分析进行测序。
[0319]
表8显示了抗
‑
nav1.7单克隆抗体中的v基因使用与velocimmune小鼠中的v基因使用的对比,其在美国专利号8,502,018和8,642,835(通过引用整体并入本文)中有所描述。
[0320]
表8
[0321]
[0322][0323]
实施例5.在mnav1.7 ko中的人nav1.2 ki/vi小鼠具有对热刺激的受损应答和对组胺的降低的痒应答。
[0324]
方法
[0325]
热板
‑
将小鼠置于热板装置(iitc,woodland hills,ca)上。在52.5℃或55℃(2种不同温度相隔10天进行试验)记录跳跃、立起和/或舔后爪的潜伏期。
[0326]
hargreaves
‑
使用hargreaves设备(iitc,woodland hills,ca)测量热痛觉过敏。在试验前至少60分钟将小鼠置于plexiglas室中。在试验期间记录3次对施加到左后爪的辐射热刺激做出应答的热潜伏期,并使用总体平均潜伏期测量。
[0327]
痒
‑
在试验前使小鼠习惯plexiglas室至少15分钟。在颈背的肩胛骨之间给小鼠真皮内注射150μg二盐酸组胺(15μl在pbs中,sigma,目录号1309009)。向上定向以观察室底部的摄像机(noldus)记录注射后长达25分钟的活动。关于组胺注射后的总搔抓发作,对视频文件进行手动评分。
[0328]
结果
[0329]
关于其对急性热刺激的应答,试验了在实施例1中描述的在mnav1.7 ko中的hnav1.2 ki的小鼠。首先,关于其对指向后爪的辐射热刺激的缩回潜伏期,试验了小鼠,也被称作hargreaves试验。在小鼠nav1.7中的hnav1.2的小鼠显示出显著延长的对热刺激的应答潜伏期(在小鼠nav1.7中的hnav1.2小鼠为22.9
±
0.9s,n=15,与此相比,wt小鼠为12.3
±
0.5s,n=19,未配对student氏t检验,p<0.0001);参见图7a。接着,在热板设备上在2个有害温度52.5℃和55℃试验小鼠(2个温度相隔10天进行试验)。表达在小鼠nav1.7中的hnav1.2的小鼠在任一温度都没有应答;所有小鼠都达到了30秒的截止时间,在此时停止试验以防止组织损伤,而wt小鼠在两种温度快速地表现出防伤害应答(在55℃为6.5
±
0.5s,n=9,和在52.5℃为10.4
±
0.6s);参见图7b。
[0330]
为了试验表达在小鼠nav1.7基因座中的hnav1.2小鼠的小鼠是否对瘙痒原具有受损的痒应答,将组胺(150μg)真皮内地注射到小鼠的颈背中,并记录搔抓发作最多25分钟。在mnav1.7中的hnav1.2小鼠表现出比wt小鼠少3.7倍的搔抓发作(在mnav1.7中的hnav1.2小鼠为24
±
11次发作,与此相比,wt为81
±
20次发作,未配对student氏t检验p=0.047);参见图7c。
[0331]
在本说明书中引用了各种出版物,包括专利、专利申请、公开的专利申请、登录号、技术文章和学术文章。这些引用的出版物中的每一篇通过引用以其整体并为了所有目的并入本文件中。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。