pifs的暗处稳定剂在促进双子叶植物顶端弯钩发育中的应用
技术领域
1.本发明涉及植物培育技术领域,特别是涉及一种pifs的暗处稳定剂在促进双子叶植物顶端弯钩发育中的应用。
背景技术:
2.双子叶植物的顶端弯钩是植物早期暗形态建成的关键结构之一,由暗处生长的黄化苗的下胚轴上端弯曲形成,其能够在植物幼芽的出土阶段保护幼嫩的子叶免受土壤机械压力的损伤,从而帮助埋藏于土壤中的种子顺利出土。衡量顶端弯钩发育程度的指标主要是下胚轴上端弯曲的角度,顶端弯钩发育有缺陷的突变体植物,包括乙烯相关的突变体以及完全不能形成顶端弯钩的hls1
‑
1突变体,会有显著的出土障碍。
3.顶端弯钩的发育通常被分为三个阶段:形成(发育早期从0度弯曲到180度),维持(角度维持在180度,过程中下胚轴迅速伸长帮助植物破土而出),以及见光后打开(见光后角度迅速减小到0度的过程)。自上世纪九十年代开始,顶端弯钩加剧作为一种乙烯处理的标志表型被人们发现,此后研究人员们对顶端弯钩的激素调控网络进行了深入的研究,并发现乙烯、赤霉素等植物激素能够正向促进顶端弯钩,而茉莉素、水杨酸等激素能够拮抗顶端弯钩的发育。此外,针对顶端弯钩发育阶段的调控十分重要,若其角度在前期形成阶段加剧过于明显,反而会将子叶暴露在土壤中,不利于植物保护幼嫩的子叶,同时也不利于顶端弯钩在见光后的迅速打开,以及后续子叶张开、光合作用等过程。
4.因此,通过化学调控剂对顶端弯钩不同发育阶段进行动态调控,能够帮助植物幼苗出土并进行接下来的光合作用与早期生命活动,提升种子的出土率和幼苗的成活率有重要的应用价值。
技术实现要素:
5.本技术经研究发现,细胞分裂素能够通过促进乙烯合成而促进乙烯下游转录因子ein3/eil1的积累来增强顶端弯钩早期的形成;并且细胞分裂素还可以通过在暗处稳定转录因子pifs(phytochrome interacting factors,简称pifs)进行转录后水平的稳定,从而促进顶端弯钩在暗处(例如土壤或培养基)的维持,协助幼苗出土,且细胞分裂素不会在光下显著影响pifs的降解过程,因而不会延迟顶端弯钩的见光后打开,不会抑制后续的光合作用。因此,细胞分裂素或其前体是一种pifs的暗处稳定剂的良好活性成分,可促进双子叶植物顶端弯钩发育。
6.基于此,有必要提供一种pifs的暗处稳定剂在促进双子叶植物顶端弯钩发育中的应用,所述暗处稳定剂包括活性成分,所述活性成分包括细胞分裂素和/或所述细胞分裂素的前体。
7.此外,还提供一种双子叶植物幼苗的制备方法。该制备方法的种子的出土率高且存活率高。
8.一种双子叶植物幼苗的制备方法,包括以下步骤:
9.将双子叶植物的种子置于含有pifs的暗处稳定剂的基质中后培养,制备幼苗,其中,所述暗处稳定剂包括活性成分,所述活性成分包括细胞分裂素和/或/或所述细胞分裂素的前体。
10.在其中一个实施例中,所述活性成分包括细胞分裂素,所述细胞分裂素选自激动素、6
‑
苄氨基嘌呤和反式玉米素中的至少一种。
11.在其中一个实施例中,制备幼苗的步骤包括:
12.将所述双子叶植物的种子置于含有pifs的暗处稳定剂的基质中,以使所述种子吸胀;及
13.将吸胀后的种子暗培养后光照培养,制备幼苗。
14.在其中一个实施例中,所述活性成分在所述基质中的浓度为0.1μm~30μm。
15.在其中一个实施例中,在将所述吸胀后的种子暗培养之前,还包括催芽的步骤。
16.在其中一个实施例中,所述催芽的步骤包括:将所述吸胀后的种子置于光照下4小时~8小时。
17.在其中一个实施例中,所述暗培养的时间为72小时~108小时;
18.及/或,所述暗培养的温度为18℃~25℃;
19.及/或,所述光照培养的温度为18℃~25℃;
20.及/或,所述双子叶植物为拟南芥、大豆或番茄。
21.在其中一个实施例中,在所述将双子叶植物的种子置于含有pifs的暗处稳定剂的基质中后培养的步骤之前,还包括对所述种子进行消毒处理的步骤。
附图说明
22.图1为实施例1中的拟南芥乙烯不敏感突变体ein3 eil1种子和col
‑
0型拟南芥种子经acc和kr分别处理后在黑暗中培养3.5天的植株;
23.图2为实施例2中的拟南芥乙烯不敏感突变体ein3 eil1种子和col
‑
0型拟南芥种子经不同浓度的kr和kt分别处理后的植株表型分析结果;
24.图3~图4为实施例3中的col
‑
0型拟南芥种子和拟南芥不同突变体的种子经acc、kr和kt分别处理后的顶端弯钩角度和植株;
25.图5为实施例3中的顶端弯钩中tcs基因的表达情况;
26.图6~图7为实施例3中的col
‑
0型拟南芥种子和拟南芥乙烯不敏感突变体ein3 eil1种子经acc、kr、kt、6
‑
ba和trans
‑
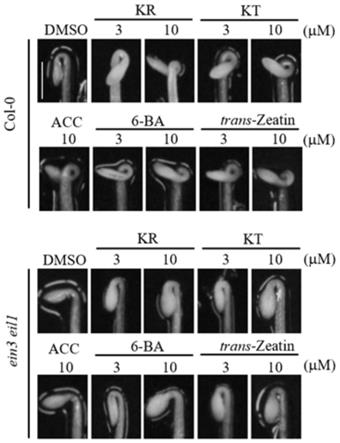
zeatin分别处理后的顶端弯钩角度和植株;
27.图8为实施例4中的拟南芥乙烯不敏感突变体ein3 eil1种子经dmso和kt处理的顶端弯钩形成时期的表型图;
28.图9为实施例4中的拟南芥乙烯不敏感突变体ein3 eil1种子经dmso、acc和kt处理下生长3天、3.5天、4天、4.5天和5天的顶端弯钩角度;
29.图10为实施例4中的拟南芥乙烯不敏感突变体ein3 eil1种子经dmso、acc和kt处理后植株全苗中的hlsi/pp2a的表达情况;
30.图11为实施例4中的拟南芥乙烯不敏感突变体ein3 eil1种子经acc和kt分别处理后的生长素在植株的分布情况;
31.图12为实施例4中的col
‑
0型拟南芥种子、拟南芥乙烯不敏感突变体ein3eil1种子
和拟南芥pifq突变的种子经acc和kt分别处理后的顶端弯钩的角度随时间的的变化图;
32.图13~图14为实施例5中的转基因植物pif4ox/pifq的种子经kt和acc分别处理后的植株和顶端弯钩角度随光照时间的变化结果;
33.图15~图16为实施例5中的转基因植物pif5ox/pifq的种子经kt和acc分别处理后的植株和顶端弯钩角度随光照时间的变化结果;
34.图17为实施例5中的的转基因植物pif4ox/pifq的种子经kt处理后其植株中gfp蛋白和hsp90蛋白含量随光照时间的变化情况;
35.图18为实施例5中的的转基因植物pif5ox/pifq的种子经kt处理后其植株中gfp蛋白和hsp90蛋白含量随光照时间的变化情况。
具体实施方式
36.为了便于理解本发明,下面将对本发明进行更全面的描述,本发明可以以许多不同的形式来实现,并不限于本文所描述的实施例。相反地,提供这些实施例的目的是使本发明公开内容更加透彻全面。术语“第一”、“第二”等仅用于描述目的,而不能理解为指示或暗示相对重要性。
37.除非另有定义,本文所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。本文中在本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不是旨在于限制本发明。本文中,dmso是指二甲基亚砜的简称;acc是1
‑
aminocyclopropane
‑1‑
carboxylic acid的缩写,为乙烯合成前体;kt是kinetin的缩写,指激动素,kt的化学结构式为:kr是kinetin riboside的缩写,是核糖化的激动素,kr的化学结构式为:
38.通常,术语“植物”涉及通过细胞分裂而特征性地生长的植物界的任何各种光合作用的多细胞的生物,含有叶绿体,并且具有由纤维素组成的细胞壁。具体而言,植物旨在包括但不限于被子植物,可以为双子叶植物。
39.本技术经研究发现,细胞分裂素能够通过促进乙烯合成而促进乙烯下游转录因子ein3/eil1的积累来增强顶端弯钩早期的形成;并且细胞分裂素还可以通过在暗处稳定
pifs进行转录后水平的稳定,从而促进顶端弯钩在暗处(例如土壤或培养基)的维持,协助幼苗出土,且细胞分裂素不会在光下显著影响pifs的降解过程,因而不会延迟顶端弯钩的见光后打开,不会抑制后续的光合作用。
40.基于上述,本技术一实施方式提供了一种pifs的暗处稳定剂在促进双子叶植物顶端弯钩发育中的应用,该暗处稳定剂的活性成分包括细胞分裂素和/或该细胞分裂素的前体。需要说明的是,在本文中,细胞分裂素的前体是指能在种子发育成幼苗过程中被转化成细胞分裂素的物质。在一些实施例中,细胞分裂素的前体为核糖化的细胞分裂素。例如,激动素的前体为核糖化的激动素。可选地,pifs为pif4和/或pif5。
41.可选地,暗处稳定剂的活性成分包括选自激动素(kt)、核糖化的激动素(kr)、6
‑
苄氨基嘌呤(6
‑
ba)和反式玉米素(trans
‑
zeatin)中的至少一种。在一些实施例中,暗处稳定剂为细胞分裂素和/或细胞分裂素的前体。此时,暗处稳定剂只有活性成分,不包括其他辅料。在另一些实施例中,暗处稳定剂除活性成分外,还包括其他辅料。
42.此外,本技术一实施方式还提供了一种双子叶植物幼苗的制备方法,该制备方法包括步骤s110~步骤s120:
43.步骤s110、对双子叶植物的种子进行消毒处理。
44.可选地,双子叶植物为拟南芥、大豆或番茄。在本实施方式中,双子叶植物为拟南芥。
45.可选地,采用乙醇对双子叶植物的种子进行消毒。具体地,对双子叶植物的种子进行消毒处理的步骤包括:将种子在75%乙醇中浸泡10分钟以上。进一步地,在消毒处理的步骤中,浸泡的时间为10分钟~25分钟。在一个具体示例中,浸泡的时间为15分钟。可以理解的是,在其他实施例中,还可以采用其他消毒试剂对双子叶植物的种子进行消毒,对应地,消毒的步骤也适应性地调整。
46.可以理解的是,在一些实施例中,步骤s110可以省略。例如,购买的种子为已经经消毒处理的种子。
47.步骤s120、将双子叶植物的种子置于含有pifs的暗处稳定剂的基质中,以使种子吸胀。
48.具体地,pifs的暗处稳定剂如上述,包括活性成分,该活性成分包括细胞分裂素和/或该细胞分裂素的前体。进一步地,细胞分裂素的前体为核糖化的细胞分裂素。
49.可选地,暗处稳定剂的活性成分包括选自激动素(kt)、核糖化的激动素(kr)、6
‑
苄氨基嘌呤(6
‑
ba)和反式玉米素(trans
‑
zeatin)中的至少一种。
50.可选地,pifs的暗处稳定剂的活性成分在基质中的浓度为0.1μm~30μm。进一步地,pifs的暗处稳定剂的活性成分在基质中的浓度为1μm~10μm。
51.在一个可选地具体示例中,双子叶植物的种子为col
‑
0型拟南芥的种子,pifs的暗处稳定剂的活性成分在基质中的浓度为5μm~15μm。
52.在另一个可选地具体示例中,双子叶植物的种子为拟南芥乙烯不敏感突变体ein3 eil1的种子,pifs的暗处稳定剂的活性成分在培养基中的浓度为0.1μm~5μm。
53.可选地,基质为土壤、培养基或溶剂。可以理解的是,基质不限于上述,还可以是其他物质,能为种子吸胀提供水且能使种子与pifs的暗处稳定剂充分接触即可。
54.步骤s130、将吸胀后的种子暗培养后光照培养,培育至幼苗。
55.具体地,将吸胀后的种子埋于基质中暗培养后光照培养。可选地,此处的基质为土壤或培养基。
56.暗培养是指将吸胀后的种子置于黑暗的环境中培养。例如,将吸胀后的种子埋于土壤中。可选地,暗培养的时间为72小时~108小时。进一步地,暗培养的时间为84小时~96小时。可选地,暗培养的温度为18℃~25℃。进一步地,暗培养的温度为21℃~23℃。可以理解的是,在其他实施例中,暗培养的时间和温度可以根据具体植物进行调整。可以理解的是,在暗培养阶段,若用透明的容器承装透明的基质培养种子时,培养容器中的基质应避光。
57.光照培养是指将埋有种子的基质置于光照下培养。可选地,光照培养的温度为18℃~25℃。进一步地,光照培养的温度为21℃~23℃。可选地,光照培养的时间为1小时~24小时。进一步地,光照培养的时间为1小时~10小时。可以理解的是,在其他实施例中,光照培养的时间和温度可以根据具体植物进行调整。
58.在一些实施例中,在将吸胀后的种子埋于基质中培育之前,还包括催芽的步骤。可选地,催芽的步骤包括:将吸胀后的种子置于光照下4小时~8小时。可选地,催芽时的温度为21℃~23℃。
59.可以理解的是,在一些实施例中,步骤s120和s130可以合并。此时,将双子叶植物的种子埋于含有pifs的暗处稳定剂的培养基培养基质中后,培育至幼苗。
60.上述双子叶植物幼苗的制备方法通过采用含有pifs的暗处稳定剂对种子进行处理,使得顶端弯钩在出土之前,可以维持在180度左右,帮助出土,保护幼嫩的子叶免受土壤机械压力的损伤,且在见光之后,暗处稳定剂不影响pifs的降解,顶端弯钩可以迅速减小到0度左右,不影响后续的光合作用,进而提高种子的出土率和幼苗的成活率。
61.具体实施例
62.以下结合具体实施例进行详细说明。以下实施例如未特殊说明,则不包括除不可避免的杂质外的其他组分。实施例中采用试剂和仪器如非特别说明,均为本领域常规选择。实施例中未注明具体条件的实验方法,按照常规条件,例如文献、书本中所述的条件或者生产厂家推荐的方法实现。在下文和附图中,“apical hook angle”指顶端弯钩角度;“hypocotyl”是指下胚轴长度;“root length”指根长“degrees”是角度的单位,
°
。
63.在以下实施例中,如无特殊说明,1/2ms(murashige and skoog)培养基中添加的kt、kr或acc均是用dmso溶解后添加的,且dmso在培养基中的终浓度为0.1%(v/v)。例如,含10μm acc的1/2ms培养基是指用dmso溶解acc后加到1/2ms培养基中,acc在培养基中的浓度为10μm,dmso在培养基中的浓度为0.1%(v/v)。需要说明的是,下文中的“0.1%dmso”均是指体积百分浓度为0.1%的dmso。
64.实施例1
65.(1)将干燥的拟南芥乙烯不敏感突变体ein3 eil1(简称ein3 eil1)的种子在75%乙醇中浸泡15分钟以消毒,后用无菌水冲洗三遍后点于含0.1%dmso的1/2ms(murashige and skoog)培养基、含10μm acc的1/2ms培养基和含10μm kr的培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天(82小时)。之后对植物进行表型分析。结果如图1所示,图1中的比例尺的1mm。
66.(2)将干燥的col
‑
0型拟南芥种子(也即是哥伦比亚0型拟南芥种子,为野生型拟南芥种子)在75%乙醇中浸泡15分钟以消毒,后用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基、含10μm acc和0.1%dmso的1/2ms培养基和含10μm kr和0.1%dmso的培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天(82小时)。结果如图1所示,图1中的比例尺的长度为1mm。
67.由图1可知,kr将乙烯不敏感突变体ein3 eil1的顶端弯钩角度从90度增大到180度左右,这说明kr不依赖于乙烯信号调控顶端弯钩发育。
68.实施例2
69.将干燥的col
‑
0型拟南芥种子和拟南芥乙烯不敏感突变体ein3 eil1的种子分别在75%乙醇中浸泡15分钟以消毒;然后分别用无菌水冲洗三遍后点于含有不同浓度kr的1/2ms培养基和含有不同浓度kt的1/2ms培养基中。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天。之后对植物进行表型分析(测尖端弯钩角度、下胚轴长度和根长),结果如表1和图2所示。图2中,“chemical concentrations”指化合物浓度,也即是kt或kr的浓度。
70.表1
[0071][0072]
由图2可以看出,kt和kr对拟南芥种子的作用效果(顶端弯钩弯曲)表现出了很高的一致性,且其促进ein3 eil1顶端弯钩的最适浓度约为1μm,而此时其对于野生型(col
‑
0)的顶端弯钩无明显促进作用,而在高浓度(10μm)下kt和kr均会使野生型植株的顶端弯钩出现加剧。因此,通过对kr和kt的施加浓度进行控制,可以实现对顶端弯钩不同时段(形成与维持)的精确调控。
[0073]
由表1可知,顶端弯钩角度对kt和kr的响应比下胚轴和根更加敏感,而且可以看出kt和kr对下胚轴和根的促进主要依赖于乙烯(表现为突变体ein3eil1需要更高的浓度来达到相同的效果),而顶端弯钩则不是。并且,从图2和表1可看出,在施加了kt和kr之后,拟南芥野生型col
‑
0和ein3 eil1的顶端弯钩均出现了不同程度的加剧,且ein3 eil1的顶端弯钩角度相较野生型对kt或kr更加敏感(ec50更小)。在达到最大浓度之前,顶端弯钩角度随着kt和kr浓度的增加而增加,表现出了浓度依赖性。由此说明,细胞分裂素存在乙烯不依赖的方式调控顶端弯钩。
[0074]
实施例3
[0075]
将干燥的col
‑
0型拟南芥种子、拟南芥细胞分裂素受体双重突变体ahk2/3(简称ahk2/3)的种子、拟南芥细胞分裂素受体双重突变体ahk2/4(简称ahk2/4)的种子、拟南芥细胞分裂素受体双重突变体ahk3/4(简称ahk3/4)的种子和拟南芥细胞分裂素下游转录因子
的三重突变体arr1/11/12(简称arr1/11/12)的种子分别在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm acc的1/2ms培养基、含10μm kr的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天。之后对植物拍照并进行表型分析(测尖端弯钩角度),结果如图3和图4所示,图4中的比例尺表示1mm。
[0076]
另外,将干燥的转基因拟南芥种子(tcs::gfp)在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm kr的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天。之后将植物制片并在激光共聚焦显微镜((zeiss lsm
‑
880)下观察(激发波长:488nm,检测的发射波长为510
–
525nm)。结果如图5所示,图5中的比例尺的长度表示50μm。
[0077]
由图3和图4可知,细胞分裂素处理的这些突变体的顶端弯钩角度均小于kt和kr处理的野生型(col
‑
0)。由图5可知,kt和kr均可以促进tcs的表达(tcs是一个细胞分裂素信号响应原件,即有细胞分裂素信号激活的地方就会有绿色荧光蛋白被表达出来),这也说明了kt和kr均可以激活细胞分裂素信号。由图3~图5可知,kt和kr的作用效果在细胞分裂素信号突变体中均出现了一定程度的减弱,且其均能够激活细胞分裂素信号报告基因tcs::gfp的表达。
[0078]
由上可知,kt和kr都是通过细胞分裂素信号来调控顶端弯钩的,而且经本技术验证,除了kt之外,如6
‑
苄氨基嘌呤(6
‑
ba),反式玉米素(trans
‑
zeatin)等经典的细胞分裂素均有与kr相似的功能(图6~图7,图7中的比例尺的长度表示1mm)。由此可知,细胞存在乙烯不依赖的方式调控顶端弯钩。
[0079]
实施例4
[0080]
将干燥的col
‑
0型拟南芥种子、拟南芥乙烯不敏感突变体ein3 eil1种子分别在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)或含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下,在植物表型动态观测平台(dynaplant)上于黑暗中培养48小时后,每隔一小时拍摄一张照片,结果如图8所示(图8中的“hai”表示萌发后小时,hours after germination)。
[0081]
将干燥的col
‑
0型拟南芥种子、拟南芥乙烯不敏感突变体ein3 eil1种子分别在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)含10μm acc的1/2ms培养基、含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下,从吸胀后三天开始,每隔12小时记录一次表型(顶端弯钩角度),到吸胀后五天截止。结果如图9所示(图9中的“dai”表示吸胀后小时,days after imbibition)。
[0082]
hls1基因表达水平检测:(1)样品准备:将干燥的拟南芥乙烯不敏感突变体ein3 eil1种子在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μmacc的1/2ms培养基、含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小
时以促进萌发后置于22℃条件下,于暗处培养5天后在暗中收取植物样品冻存于液氮中,供后续进行荧光定量pcr实验。(2)荧光定量pcr实验:用eastup super total rna extraction kit(promega shanghai)试剂盒进行rna提取并用m
‑
mlv逆转录酶进行反转录后,用light cycler 480system(roche)仪器与sybr green mix(takara)酶进行荧光定量pcr实验以检测rna的转录水平,结果如图10所示。
[0083]
生长素信号不对称分布观测:将干燥的转基因拟南芥种子(dr5::gfp/ein3eil1)在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm acc的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养3.5天。之后将植物制片并在激光共聚焦显微镜((zeiss lsm
‑
880)下观察(激发波长:488nm,检测的发射波长为510
–
525nm)。结果如图11所示。
[0084]
由图8可知,细胞分裂素能够以乙烯不依赖的方式促进顶端弯钩在暗处的维持;由图9可知,细胞分裂素促进乙烯不敏感突变体ein3 eil1顶端弯钩维持的效果至少能够达到培养后五天之久;由图10可知,细胞分裂素有不依赖于乙烯促进顶端弯钩发育的途径,该途径能够在ein3 eil1突变体中促进顶端弯钩发育相关基因hls1的表达;由图11可知,细胞分裂素有不依赖于乙烯促进顶端弯钩发育的途径,该途径能够促进生长素的不对称分布,从而导致顶端弯钩的维持。由图11可以排除细胞分裂素本身通过对细胞分裂产生影响从而导致顶端弯钩处的不对称生长。
[0085]
由图8~图11可知,细胞分裂素对顶端弯钩的维持有着很强的促进作用,其能够使ein3 eil1在培养后五天仍然保持闭合(180度)的状态,同时也会激活hookless1的表达以及生长素的不对称分布(两个顶端弯钩发育的最基础通路),相较之下,未经处理的植物顶端弯钩在此时以及几乎完全打开。
[0086]
突变体表型动态观测实验:将干燥的col
‑
0型拟南芥种子、拟南芥乙烯不敏感突变体ein3 eil1、以及拟南芥光受体互作因子四重突变体pifq分别在75%乙醇中浸泡15分钟以消毒;然后均分别用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm acc的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养。之后从吸胀后3天开始,每隔12小时变统计一次不同突变体在不同处理下的表型(顶端弯钩的角度),结果如图12所示。图12中pifq是指pif的四重突变体(在col
‑
0的基础上缺失了pif1基因、pif3基因、pif4基因和pif5基因,简称pifq)。
[0087]
在图12中,野生型col
‑
0中,kt处理组和acc处理组的顶端弯钩角度表现出了相似程度的加剧(260度左右),而kt处理组的顶端弯钩角度维持了更长的时间。此外,在ein3 eil1中,kt仍然具有此功能。在pifq中,无论是否有kt处理,pifq的顶端弯钩角度均不能在暗处维持。由此可知,细胞分裂素(kt)对顶端弯钩的维持作用不依赖于乙烯信号,但是依赖于pif的存在。也即是说,与另一种能够强烈地促进顶端弯钩加剧的激素乙烯相比,细胞分裂素在促进顶端弯钩维持的方面表现出了更加持久的特性,能够显著的延长其在暗处维持的时间,且这种效果依赖于pifs蛋白的存在。
[0088]
实施例5
[0089]
为了验证细胞分裂素的处理不会阻止顶端弯钩的见光后打开,从而抑制后续的发育以及光合作用,利用过表达pif4和pif5的转基因植物进行了实验。具体过程包括如下步骤:
[0090]
(1)转基因植物光处理表型实验:将干燥的拟南芥pif4过表达株系(pif4ox/pifq)的种子和将干燥的拟南芥pif5过表达株系(pif5ox/pifq)的种子分别在75%乙醇中浸泡15分钟以消毒;然后用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm acc的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养,暗培养3.5天后将植物置于持续光照下0小时,6小时,12小时后分别记录表型(顶端弯钩角度)。实验结果如图13~图16所示,其中:图13为pif4ox/pifq的表型数据统计图(图13中“hae”表示见光后小时:hours after exposure,下文同理);图14为图13中不同处理组具有代表性的植株表型图;图15为pif5ox/pifq的表型数据统计图;图16为图15中不同处理组具有代表性的植株表型图。
[0091]
(2)蛋白质免疫印迹实验:将干燥的拟南芥pif4过表达株系(pif4ox/pifq)的种子和干燥的拟南芥pif5过表达株系(pif5ox/pifq)的种子分别在75%乙醇中浸泡15分钟以消毒;然后用无菌水冲洗三遍后点于含0.1%dmso的1/2ms培养基(作为空白对照)、含10μm acc的1/2ms培养基和含10μm kt的1/2ms培养基上。之后将种子和培养基一起置于四度冰箱中吸胀三天。随后取出板子,置于光下照射4小时以促进萌发后置于22℃条件下黑暗中培养,暗培养3.5天后,将dmso处理组的植物分别用光照处理0分钟、15分钟、30分钟和45分钟后收取植物样品冻存于液氮中,将kt处理组的植物分别用光照处理0分钟、15分钟、30分钟、45分钟和60分钟后收取植物样品冻存于液氮中。
[0092]
将植物的总蛋白分别用2
×
sds buffer提取(60mm tris
‑
hcl,ph 6.8,25%213glycerol,2%sds,14.4mβ
‑
mercaptoethanol,0.1%bromophenol,and 1mdtt),之后再7.5%sds聚丙烯酰胺凝胶上电泳,随后被电转移至pvdf薄膜上用对应的anti
‑
gfp或anti
‑
actin抗体进行杂交和显影,结果如图17和图18所示。在图17和图18中,“l0m”表示见光0min的组别,“l15m”表示见光15min的组别,其他类推。
[0093]
由图13~图16可知,kt处理的转基因植物pif4ox/pifq以及pif5ox/pifq,虽然早期能够表现出极强的顶端弯钩加剧,但在光照处理6小时后顶端弯钩便有了明显的打开趋势,角度基本减半,且在12小时后减小到了90度左右。相较之下,乙烯处理的转基因植物pif4ox/pifq以及pif5ox/pifq,虽然也能够诱导顶端弯钩加剧,但其对光的响应比细胞分裂素更加迟钝,在光照6小时的情况下并没有表现出明显的打开趋势,只有在处理12小时的情况下回到了180度左右的角度。
[0094]
由图17和图18可知,kt能够在暗处(光处理0h,0hae)促进pifs的积累,而当植物见光时,kt的处理也并未显著地延缓pifs的降解,在大约见光一小时后,pifs便会降解殆尽,这也与表型上的结论高度相似。
[0095]
由上可知,细胞分裂素能够在暗处促进顶端弯钩维持,并在光下迅速打开的特性为植物幼苗的出土提供了强有力的保障,且不会影响其后续的光合作用以及光下的生长。
[0096]
以上所述实施例的各技术特征可以进行任意的组合,为使描述简洁,未对上述实施例中的各个技术特征所有可能的组合都进行描述,然而,只要这些技术特征的组合不存
在矛盾,都应当认为是本说明书记载的范围。
[0097]
以上所述实施例仅表达了本发明的几种实施方式,便于具体和详细地理解本发明的技术方案,但并不能因此而理解为对发明专利保护范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。应当理解的是,在本领域技术人员在本发明提供的技术方案的基础上,通过合乎逻辑的分析、推理或有限的试验得到的技术方案,均在本发明所附权利要求的保护范围内。因此,本发明专利的保护范围应以所附权利要求的内容为准,说明书及附图可以用于解释权利要求的内容。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。