1.本发明涉及来源于南极假丝酵母和玉米丝黑穗病菌脂肪酶突变体及其在不对称水解制备光学纯医药中间体中的应用,属于生物工程技术领域。
背景技术:
2.脂肪酶是一类多功能生物催化剂,可催化水解、酯化、转酯化等反应。由于其在水相、有机相中具有催化活性高、稳定性好、立体选择性好及广谱底物特异性等优点,广泛应用于酶催化合成工艺中,制备各类光学纯的高级医药中间体。在帕罗西汀(paroxetine)、依地普仑(escitalopram)、度洛西汀(duloxetine)、利斯的明(ricastigmine)和布洛芬(ibuprophen)等关键中间体制备工艺中均利用脂肪酶作为生物催化剂。研究表明(us20100087525),脂肪酶具有广底物谱,且调节反应温度和助溶剂可以显著影响其选择性和催化活性,表明其活性口袋具有较强的可塑性。因此可利用定向进化、半理性设计等技术对脂肪酶进行改造,提高其立体选择性和催化活性,进一步扩大脂肪酶应用范围。
3.定向进化(directed evolution)是在实验中模拟自然进化过程,通过快速突变和重组等方式人为的产生基因多样性,然后按照特定的需要和目的给予筛选压力,筛选出符合应用预期的酶蛋白。这一技术已被广泛应用于酶工程等生物以及其它交叉领域中,但酶催化的立体选择性改造的研究相对于其它酶学性质改造而言,发展缓慢,究其原因是酶的立体选择性改造筛选模型构建难度大,利用hplc或gc进行分析检测的效率难以匹配大体量突变文库的筛选,因此立体选择性改造的酶定向进化研究仍然是一个挑战。随着对蛋白质结构与功能理解的进步,对酶的改造逐渐转移至基于半理性设计技术构建的功能丰富的小型突变文库上。半理性设计是指利用蛋白质序列、结构、功能等相关信息以及计算机辅助来为酶的改造预筛选可能的有益靶点和氨基酸残基,从而构建高质量的突变文库,减少筛选工作量,提高酶改造效率。
4.利用脂肪酶制备手性中间体主要有手性拆分和不对称水解两种方案。拆分工艺由于需要对外消旋体进行光学拆分,因此目的构型产物理论收率低于50%;不对称水解工艺无需舍弃其中一种光学异构体,因此从根本上解决拆分工艺中对映异构体消旋回收造成的产能偏低及消旋反应收率问题,理论收率可达100%。近年来脂肪酶不对称水解获得了越来越多研究者的关注,但实际应用效果不尽如人意。zaida等人利用固定化脂肪酶不对称水解3-苯基戊二酸二乙酯制备(r)-5-乙氧基-5-氧代-3-苯基戊酸(tetrahedron:asymmetry 22(2011)2080-2084),以二氧六环为助溶剂,在低温条件下进行反应,产物e.e.值为92%。wei等人利用脂肪酶不对称水解3-(4-氟苯基)戊二酸二甲酯制备(r)-3-(4-氟苯基)-5-甲氧基-5-氧戊酸(journal of molecular catalysis b:enzymatic 97(2013)270-277),以二氧六环为助溶剂,在30℃进行反应,产物e.e.值仅为71.6%。liu等人选择3-(4-氟苯基)-戊二酸二甲酯为模型底物,筛选商品化脂肪酶进行不对称水解工艺研究,结果如表1所示,添加10%乙腈为助溶剂,室温条件下反应,产物立体选择性均无法达到工艺要求(process biochemistry 47(2012)1037-1041)。
5.表1文献报道脂肪酶筛选
6.脂肪酶e.e.(%)构型ppl56.3scrl11.1srnl3.6rhle26.4rlipase ak0-lipase ps3.7slipase ays0-lipozyme tl63.0snovozym 43591.8r
7.teva(us20100087525)报道了一条利用商品化脂肪酶不对称水解底物3-异丁基戊二酸二酯的工艺路线,通过流加底物3-异丁基戊二酸二甲酯和助溶剂叔戊醇,在-2℃低温进行反应,最终底物浓度约135g/l,转化率99%,产物为s构型,e.e.值96%。而当一次性投入3-异丁基戊二酸二甲酯-2℃低温进行反应时,产物e.e.值下降到89%。如果在反应体系中移除助溶剂叔戊醇,相同反应条件产物e.e.值降为70%。综合上述案例,脂肪酶不对称水解工艺仍存在以下问题:1)具有严格立体选择性的不对称水解脂肪酶来源不足,无法满足工艺开发要求;2)立体选择性不稳定,受反应温度、助溶剂和底物添加模式影响极大;3)受限于低温环境转化,脂肪酶催化活性远低于最适反应条件,酶使用成本上升,产能大幅降低;4)添加助溶剂也提高了工艺成本和产物分离提取难度,无法实现工业化放大。
8.本研究通过半理性设计技术,以3-异丁基戊二酸二乙酯为模型底物,以南极假丝酵母脂肪酶(pseudozyma antarctia lipase,pal)为研究对象进行突变位点分析,针对立体选择性进行改造,获取正向单突变体;将正向突变位点进行组合后构建了系列突变株,能在常温条件,全水相体系,针对多种二酯进行立体选择性严格的不对称水解;利用基因挖掘技术克隆表达玉米丝黑穗病菌脂肪酶(sporisorium reilianum lipase,srl),整合pal正向突变位点后,在二酯底物不对称水解中表现了严格的立体选择性,为脂肪酶不对称水解工艺提供高效且稳定的生物催化剂,具备极大的产业化应用潜力。
技术实现要素:
9.本发明通过半理性设计技术提供了一系列南极假丝酵母pal和玉米丝黑穗病菌srl脂肪酶突变体,能避免低温反应条件并解除助溶剂依赖性,反应立体选择性和工艺稳定性显著提升,为一系列二酯底物的不对称水解工艺提供高效且稳定的生物催化剂。
10.本发明采用的技术方案是:
11.本发明所述的南极假丝酵母脂肪酶pal关键突变位点的选择是通过突变体库的构建及筛选确定的。
12.本发明提供一系列南极假丝酵母脂肪酶pal突变体,相对于野生型南极假丝酵母脂肪酶pal,所述突变体对应的突变位点及突变形式如表2所示的单点突变形式以及所有可能的组合方式所构成。
13.表2有益突变位点及其突变形式
14.氨基酸位点亲本氨基酸有益突变形式140lv,i,m,a,f,w,g,s,c,t,d,n,e,p,h,k154va,i,l,m,f,w,s,c,t,q,y281ai,l,m,f,v,s,c,t282av,i,l,m,f,g,s,c,n,k,y,r
15.本发明所述野生型南极假丝酵母脂肪酶pal氨基酸序列如seq id no:1所示,对应编码基因如seq id no:2所示。所述突变体l140x1,v154x2,a281x3,a282x4,l140x1/v154x2,l140x1/a281x3,l140x1/a282x4,v154x2/a281x3,v154x2/a282x4,a281x3/a282x4,l140x1/v154x2/a281x3,l140x1/v154x2/a282x4,l140x1/a281x3/a282x4,v154x2/a281x3/a282x4,l140x1/v154x2/a281x3/a282x4,其中x1表示l140位点的有益突变形式,x2表示v154位点的有益突变形式,x3表示a281位点的有益突变形式,x4表示a282位点的有益突变形式。
16.本发明所述野生型玉米丝黑穗病菌的脂肪酶srl氨基酸序列如seq id no:3所示,对应编码基因如seq id no:4所示。根据srl和pal脂肪酶一级结构和三级结构比对结果,srl脂肪酶145、159、287和288位分别对应pal脂肪酶140、154、281和282位氨基酸位点。所述srl脂肪酶突变体l145x1,v159x2,a287x3,g288x4,l145x1/v159x2,l145x1/a287x3,l145x1/g288x4,v159x2/a287x3,v159x2/g288x4,a287x3/g288x4,l145x1/v159x2/a287x3,v159x1/v159x2/g288x4,l145x1/a287x3/g288x4,v159x2/a287x3/g288x4,l145x1/v159x2/a287x3/g288x4,其中x1表示l145位点的有益突变形式,x2表示v159位点的有益突变形式,x3表示a287位点的有益突变形式,x4表示g288位点的有益突变形式。
17.本发明还涉及含有所述编码基因的重组载体,以及利用所述重组载体转化得到的重组基因工程菌。所述重组载体是用常规方法将本发明所述突变体核苷酸序列连接于各种载体上构建而成。所述载体可为本领域常规的各种载体,如各种质粒、噬菌体或病毒载体等。
18.本发明还提供一种含所述编码基因或所述重组载体的基因工程菌。所述基因工程菌可通过将本发明的重组表达载体转化至宿主微生物中获得。所述的宿主微生物可为本领域常规的各种宿主微生物,只要满足重组表达载体可以稳定自我复制且所携带的本发明的突变体基因可以有效表达,如各类酵母、大肠杆菌、芽孢杆菌、放线菌或霉菌等。
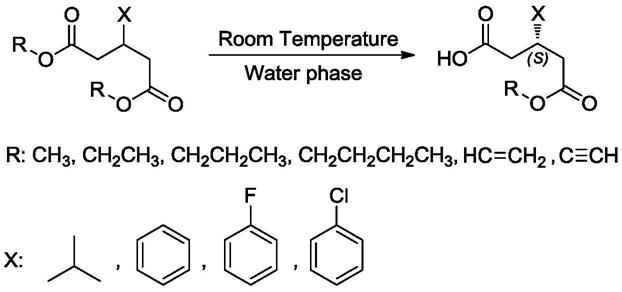
19.本发明还涉及所述的脂肪酶及其突变体在不对称水解x-取代戊二酸二酯制备s-单酯产物的应用。所述应用是将底物、突变体酶和缓冲液投入到反应器中进行催化反应;所述底物浓度为1-300g/l;所述缓冲液为ph 4.0-9.0的柠檬酸-柠檬酸钠,磷酸盐和tris-hcl缓冲液等;所述应用的反应温度为10-40℃。反应式如下所示。
[0020][0021]
本发明的有益效果主要体现在:获得了一系列能在常温条件(10-40℃)、纯水相等工业化可接受条件下不对称水解前手性底物制备(s)-3-异丁基-5-甲氧基-5-氧戊酸、(s)-3-异丁基-5-乙氧基-5-氧戊酸、(s)-3-(4-氟苯基)-5-甲氧基-5-氧戊酸和(s)-3-(4-氯苯基)-5-甲氧基-5-氧戊酸等中间体的高选择性脂肪酶及其突变体,反应e.e.>95%,转化率>99%,为首次报道常温水相条件专一性不对称水解一系列二酯底物生成s-单酯产物的脂肪酶。将突变体应用于各种医药中间体不对称水解工艺后,可在制备工艺中避免“拆分-消旋回收”工艺,提高产能和底物利用率高,立体选择性严格,可显著提高生产效率,具备极大的工业化应用潜力。
附图说明
[0022]
图1为活性口袋结构分析及突变位点选择
具体实施方式
[0023]
基因来源:本专利所涉及的pseudozyma antarctia脂肪酶pal基因和sporisorium reilianum脂肪酶srl基因均通过全基因合成获得,所涉及的突变体均为定点突变所得。
[0024]
酶的三维结构分析:酶的三维结构分析采用pymol可视化软件。
[0025]
对映体选择性测定:反应产物(s)-3-异丁基-5-甲氧基-5-氧戊酸、(s)-3-异丁基-5-乙氧基-5-氧戊酸、(s)-3-(4-氟苯基)-5-甲氧基-5-氧戊酸和(s)-3-(4-氯苯基)-5-甲氧基-5-氧戊酸及其相应的对映异构体采用高效液相色谱法测定,检测条件参考us 20100087525。
[0026]
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
[0027]
实施例1:突变位点的选择
[0028]
脂肪酶pal的活性口袋由酰基口袋和醇基口袋组成,两个口袋的连接处有催化三联体d187-h224-s105以及氧负离子洞。构成酰基口袋的氨基酸残基t138、l140、v154和i189以及构成醇基结合口袋的氨基酸残基l278、a281、a282可能影响酶与底物的结合构象,如图1所示,选取上述7个位点作为候选突变位点。
[0029]
实施例2:重组表达载体及工程菌的构建
[0030]
选取pal(seq id no:1)和srl(seq id no:3)脂肪酶序列,根据表达载体pet22b的特点设计了ecor i和hind iii,通过基因工程的常规操作以全基因合成的方法合成了脂肪酶基因pal(seq id no:2)和srl(seq id no:4)。利用ecor i和hind iii限制性内切酶对实
施例1中合成的pal和srl基因片段进行双酶切以及回收处理,并利用t4 dna连接酶将该片段与用相同的限制性内切酶处理的商品化载体pet22b在16℃下连接过夜,从而构建得到重组表达载体pet22b-pal和pet22b-srl。将构建完成的重组表达载体转化至e.coli bl21(de3)感受态细胞中,涂布于含终浓度50μg/ml氨苄青霉素的lb平板,37℃下培养过夜;于平板上长出的菌落中随机挑取克隆进行菌落pcr鉴定,阳性克隆测序验证,结果表明重组菌e.coli bl21(de3)/pet22b-pal和e.coli bl21(de3)/pet22b-srl已成功构建。
[0031]
实施例3:含有重组脂肪酶或其突变体菌体细胞的制备
[0032]
将基因工程菌e.coli bl21(de3)/pet22b-pal和e.coli bl21(de3)/pet22b-srl或其突变体接种至含有50μg/ml氨苄青霉素的2
×
yt培养基中,37℃培养至菌体浓度od
600
值为0.4-0.6,再向2
×
yt液体培养基中加入终浓度0.1mmol/l的iptg,18℃诱导培养18-24h后,将培养液于4℃、8000rpm离心5min,弃去上清液,收集含有重组pal和srl脂肪酶或其突变体的湿菌体。
[0033]
实施例4:候选位点氨基酸对对映体选择性影响效果初筛
[0034]
针对实施例1中确定的7个候选位点,通过定点突变构建初筛突变文库,利用高保真酶primerstar扩增,pcr产物直接用dpn i消化,转化e.coli bl21(de3),涂布amp平板筛选,测序验证后获得突变体,如实施例3所述获得相应的突变酶,以3-异丁基戊二酸二乙酯为模型底物,探查每个候选位点氨基酸变化对对映体选择性的影响。以野生型pal为对照,对pal突变体的对映体选择性进行检测,结果发现140、154、281和282位点改造对映体选择性影响显著,因此对以上4个位点进行进一步定点饱和突变。针对srl脂肪酶,候选位点也通过pal对映体选择性研究结果确定,影响显著的位点为145、159、287和288位。
[0035]
实施例5:脂肪酶定点饱和突变文库构建及选择性测定
[0036]
利用定点突变构建pal脂肪酶140、154、281、282位和srl脂肪酶145、159、287、288位系列点突变体。分别以pet22b-pal和pet22b-srl质粒为模板,利用高保真酶primerstar扩增,pcr产物直接用dpn i消化,转化e.coli bl21(de3),涂布amp平板筛选,测序验证后获得突变体,如实施例3所述获得相应突变酶。以3-异丁基戊二酸二乙酯为模型底物,对所获得系列突变体对映体选择性进行比较,结果如表3所示。srl脂肪酶145、159、287、288位系列单点突变体对映体选择性变化规律分别与pal脂肪酶140、154、281、282点突变相同,在对应位点氨基酸相同的条件下,srl突变体的对映体选择性均比pal突变体稍差。
[0037]
表3脂肪酶对映体选择性测定
[0038]
[0039]
注:产物构型为r型,e.e.值标记为
“-”
;产物构型为s型,e.e.值0-30%标记为“ ”;e.e.值30-60%标记为“ ”;e.e.值60-90%标记为“ ”;e.e.值>90%标记为“ ”[0040]
实施例6:脂肪酶组合突变体转化制备(s)-3-异丁基-5-甲氧基-5-氧戊酸
[0041]
根据实施例5单点突变构建组合突变菌株e.coli bl21(de3)/pet22b-pal(l140v/a281v),以实施例3方法制备菌体进行催化反应。在50ml转化瓶中加入9.9ml 100mm缓冲液(ph 8.0),加入底物3-异丁基戊二酸二甲酯100μl,0.39g湿菌体,反应温度30℃,摇床转速150rpm,反应结束后用正己烷萃取,hplc分析转化率及反应萃取液中(s)-3-异丁基-5-甲氧基-5-氧戊酸的对映体比例,结果显示转化率>99%,产物(s)-3-异丁基-5-甲氧基-5-氧戊酸e.e.>95%。
[0042]
实施例7:脂肪酶组合突变体转化制备(s)-3-异丁基-5-乙氧基-5-氧戊酸
[0043]
根据实施例5单点突变构建组合突变菌株e.coli bl21(de3)/pet22b-srl(l145m/v159l/a287v),以实施例3方法制备菌体进行催化反应。在50ml转化瓶中加入9.9ml 100mm缓冲液(ph 8.0),加入底物3-异丁基戊二酸二乙酯100μl,0.39g湿菌体,反应条件及样品处理方式同实施例6。检测结果显示转化率>99%,产物(s)-3-异丁基-5-乙氧基-5-氧戊酸e.e.>95%。
[0044]
实施例8:脂肪酶组合突变体转化制备(s)-3-(4-氟苯基)-5-甲氧基-5-氧戊酸
[0045]
根据实施例5单点突变构建组合突变菌株e.coli bl21(de3)/pet22b-pal(l140i/a281v),以实施例3方法制备菌体进行催化反应。在50ml转化瓶中加入9.9ml 100mm缓冲液(ph 8.0),加入底物3-(4-氟苯基)-戊二酸二甲酯0.1g,0.39g湿菌体,反应条件及样品处理方式同实施例6。检测结果显示转化率>99%,产物(s)-3-(4-氟苯基)-5-甲氧基-5-氧戊酸e.e.>95%。
[0046]
实施例9:脂肪酶组合突变体转化制备(s)-3-(4-氯苯基)-5-甲氧基-5-氧戊酸
[0047]
根据实施例5单点突变构建组合突变菌株e.coli bl21(de3)/pet22b-srl(v159a/a287i/g288w),以实施例3方法制备菌体进行催化反应。在50ml转化瓶中加入9.9ml 100mm缓冲液(ph 8.0),加入底物3-(4-氯苯基)-戊二酸二甲酯0.1g,0.39g湿菌体,反应条件及样品处理方式同实施例6。检测结果显示转化率>99%,产物(s)-3-(4-氯苯基)-5-甲氧基-5-氧戊酸e.e.>95%。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。