靶向纳米颗粒以及它们的与真菌感染有关的用途
1.本技术要求2019年1月8日提交的美国临时申请第62/789,862号和2019年10月10日提交的美国临时申请第62/913,489号的优先权,这两项申请都通过引用以其整体并入本文。
背景技术:
2.数百种土著真菌会导致各种各样的疾病,包括曲霉病、芽生菌病、念珠菌病、球孢子菌病(山谷热)、隐球菌病、组织胞浆菌病、皮肤真菌病和肺孢子虫性肺炎(pcp),仅举几个例子。总的来说,致病性真菌感染许多不同的器官,但皮肤和肺是最常见的部位。一些真菌病只是致残,而另一些则危及生命。尽管对真菌感染病理学的理解有所进步,但目前诊断和治疗真菌感染的方法仍有不足。
技术实现要素:
3.本文提供了用于诊断、治疗或预防真菌感染的靶向纳米颗粒,例如脂质体。一些脂质体包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子。在本文提供的脂质体中,靶向分子被掺入到脂质体的外表面中,而抗真菌剂被包封在脂质体中。
4.进一步提供了治疗或预防受试者真菌感染的方法,其包括向患有真菌感染或有罹患真菌感染风险的受试者施用脂质体的多重体,其中所述多重体中的每个脂质体均包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子,其中靶向分子被掺入到脂质体的外表面中,而抗真菌剂被包封在脂质体中。
5.还提供了制备脂质体的多重体的方法,该脂质体包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子,其中靶向分子被掺入到每个脂质体的外表面,而抗真菌剂被包封在每个脂质体中。所述方法包括:(a)在约60℃下将抗真菌剂溶解或悬浮在溶剂中约10分钟至约30分钟;(b)通过将呈悬浮液形式的脂质体的多重体与步骤a)的抗真菌剂/溶剂溶液在约60℃下混合约3至约5小时,或在约37℃下混合约24
‑
120小时来将抗真菌剂包封到每个脂质体中;以及(c)通过使包含包封的抗真菌剂的脂质体与缀合至脂质的靶向分子在约60℃下接触约45分钟至约90分钟来将靶向分子掺入到每个脂质体的外表面中。
6.还提供了包含结合靶真菌抗原的靶向分子的脂质体,其中靶向分子被掺入到脂质体的外表面中,并且其中靶向分子连接至当靶向分子结合靶真菌抗原时产生信号的分子。在一些实例中,靶向分子连接至荧光蛋白的c末端和/或n末端片段或荧光蛋白的片段。
7.进一步提供了用于检测受试者中或受试者的样本中真菌感染的方法,其包括:a)使受试者或所述受试者的样本与脂质体的多重体接触,其中所述多重体中的每个脂质体均包含结合靶真菌抗原的靶向分子,其中靶向分子被掺入到脂质体的外表面中,并且其中靶向分子被连接至当靶向分子结合靶真菌抗原时产生信号的分子;以及b)检测信号,其中信号指示真菌感染的存在。
附图说明
8.本技术包括以下附图。附图旨在说明所述组合物和方法的某些实施方案和/或特征,并旨在补充所述组合物和方法的任何一项或多项描述。附图不限制所述组合物和方法的范围,除非书面描述明确表明情况如此。
9.图1a示出了编码示例性密码子优化的可溶性小鼠dectin
‑
1(sdectin
‑
1)的核酸序列(seq id no:1)。9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线。用于克隆到pet
‑
45b kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)中的位点分别加有下划线。gly ser(g,s)柔性接头残基的密码子以粗体示出,并且反应性lys(k)残基(aag)的密码子以粗体示出,赖氨酸密码子以斜体示出。小鼠sdectin
‑
1序列(clec7a,genbank登记号aas37670.1)以纯文本示出;ala密码子gct和终止密码子taa和tta加有下划线,终止密码子以粗体示出。替代基因名称mmsdectin1lyshis。核苷酸序列的长度为604个碱基对,其中597个碱基对编码长度为199个氨基酸的蛋白质。编码示例性密码子优化的小鼠sdectin
‑
1的核酸被克隆到pet
‑
45b 中。
10.图1b示出了由seq id no:1编码的氨基酸序列(seq id no:2)。这是包含小鼠sdectin
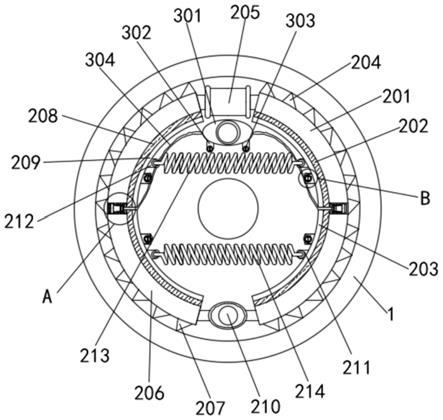
‑
1多肽的多肽。来自pet
‑
45b 的n末端氨基酸序列和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示)。小鼠sdectin
‑
1氨基酸残基以纯文本形式显示(seq id no:2的氨基酸23
‑
199),结束于以粗体显示的c末端ala残基(a),其密码子被用于将终止密码子和pacl位点放置于框架中。应当理解,任选地,本文公开的任何多肽序列中的终止密码子,如果不是该多肽衍生自的天然多肽的一部分,则可以被去除以产生不包括一个或多个终止密码子的多肽。包含小鼠sdectin
‑
1多肽的蛋白质长度为199个氨基酸,分子量(mw)为22,389.66g/mol。理论pi为7.74。应当理解,本文所述的任何包含亲和标签(例如(his)6亲和标签)的蛋白质都可以被修饰成去除了his标签。在一些实例中,本文所述的任何核苷酸序列可进一步包含用于亲和标签的翻译后和/或纯化后去除的蛋白酶切割位点。
11.图1c示出了编码示例性密码子优化的可溶性小鼠dectin
‑
2(sdectin
‑
2)的核酸序列(seq id no:3)。9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线。用于克隆到pet
‑
45b kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)中的位点分别加有下划线。gly ser(g,s)柔性接头残基的密码子以粗体显示,并且反应性lys(k)残基(aag)的密码子以粗体显示,赖氨酸密码子以斜体显示。来自clec6a小鼠dectin 2基因的密码子优化的sdectin
‑
2以纯文本形式显示,ala密码子(gct)和终止密码子taa和tta加有下划线,终止密码子以粗体显示。替代基因名称为mmsdectin2lyshis。核酸序列的长度为574个碱基对,其中567个碱基对编码长度为190个氨基酸的蛋白质。编码示例性密码子优化的小鼠sdectin
‑
2的核酸被克隆到pet
‑
45b 中。
12.图1d示出了由seq id no:3编码的氨基酸序列(seq id no:4)。此多肽包含小鼠sdectin
‑
2蛋白质。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示,赖氨酸以斜体显示。小鼠sdectin
‑
2氨基酸残基以纯文本形式显示(seq id no:4的氨基酸23
‑
189),结束于c末端ala残基(a)(粗体),其密码子用于将终止密码子和pacl位点放置于框架中。产生的包含小鼠sdectin
‑
2的多肽具有189个氨基酸,mw为21,699.25g/mol,理论pi为6.33。
13.图1e示出了编码示例性密码子优化的可溶性小鼠dectin
‑
3(sdectin
‑
3)的核酸序列(seq id no:5)。9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线。用于克隆到pet
‑
45b kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)中的位点分别加有下划线。gly ser(g,s)柔性接头残基的密码子以粗体显示。反应性赖氨酸(k)密码子(aag)以粗体显示,赖氨酸以斜体显示。来自clec4d小鼠dectin
‑
3基因(genbank登记号np_034949.3)的密码子优化的sdectin
‑
3以纯文本显示,ala密码子(gct)和终止密码子taa和tta加有下划线。终止密码子以粗体显示。替代基因名称为mmsdectin3lyshis。核苷酸序列的长度为604个碱基对,其中597个碱基对编码长度为199个氨基酸的蛋白质。编码示例性密码子优化的小鼠sdectin
‑
3的核酸被克隆到pet
‑
45b 中。
14.图1f示出了由seq id no:5编码的氨基酸序列(seq id no:6)。此多肽包含小鼠sdectin
‑
3蛋白质。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示,赖氨酸以斜体显示。小鼠sdectin
‑
3氨基酸残基以纯文本形式显示(seq id no:6的氨基酸23
‑
199),结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和pacl位点放置于框架中。该多肽的长度为199个氨基酸,mw为23,023.72g/mol,理论pi为6.52。
15.图1g示出了编码示例性密码子优化的可溶性人dectin
‑
1(sdectin
‑
1)的核酸序列(seq id no:7)。人sdectin
‑
1 dna序列是由载体pet
‑
45b 表达。9个密码子的载体pet
‑
45b 序列和his标签加有方框,起始密码子加有下划线。克隆位点bamhi(ggatcc)(seq id no:24)和paci(ttaattaa)(seq id no:21)分别加有下划线。肠激酶加工位点的密码子以小写字体显示,gly ser(g,s)柔性接头残基和反应性lys(k)残基(aaa和aag)的密码子以粗体显示(其中赖氨酸密码子以斜体显示)。人sdectin
‑
1序列(clec7a,genbank登记号nm_197947)以纯文本显示,密码子经优化用于表达。ala密码子gct和终止密码子taa和tta加有下划线,终止密码子为粗体。该序列的替代名称是hssdectin1lyshis。编码人sdectin
‑
1的核苷酸序列长度为649个碱基对,编码长度为214个氨基酸的多肽。编码示例性密码子优化的人sdectin
‑
1的核酸被克隆到pet
‑
45b 中。
16.图1h示出了由seq id no:7编码的氨基酸序列(seq id no:8)。这是包含人sdectin
‑
1蛋白质的多肽。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。肠激酶加工位点以小写字体显示。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示,赖氨酸以斜体显示。人sdectin
‑
1氨基酸残基以纯文本形式显示(seq id no:8的氨基酸35
‑
214),结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和pacl位点放置于框架中。该多肽的长度为214个氨基酸,mw为23,703.20g/mol,理论pi为6.22。
17.图1i示出了编码示例性密码子优化的可溶性人dectin
‑
2(sdectin
‑
2)的核酸序列(seq id no:9)。人sdectin
‑
2核苷酸序列是由载体pet
‑
45b 表达。核苷酸序列的长度为约580个碱基对,其中616个碱基编码长度为203个氨基酸的蛋白质。9个密码子的载体pet
‑
45b 序列(包括his标签)加有方框,起始密码子加有下划线。克隆位点bamhi(ggatcc)(seq id no:24)和paci(ttaattaa)(seq id no:21)分别加有下划线。肠激酶处理位点的密码子以小写字体显示。gly ser(g,s)柔性接头残基的密码子以粗体显示,并且反应性lys(k)残基(aag)以粗体显示,赖氨酸以斜体显示。来自clec6a人dectin 2基因(cdna genbank登记号
nm_001317999)的密码子优化的sdectin
‑
2以纯文本形式显示。ala密码子(gct)和终止密码子taa和tta加有下划线,终止密码子以粗体显示。替代基因名称为hssdectin2lyshis。编码示例性密码子优化的人sdectin
‑
2的核酸被克隆到pet
‑
45b 中。
18.图1j示出了由seq id no:9编码的氨基酸序列(seq id no:10)。此多肽包含人sdectin
‑
2蛋白质。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。肠激酶加工位点为以小写字体显示。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示,赖氨酸以斜体显示。人sdectin
‑
2氨基酸残基以纯文本形式显示(genbank登记号np_001007034.1)(seq id no:10的氨基酸36
‑
203),结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和pacl位点放置于框架中。该多肽的长度为203个氨基酸,mw为22,969g/mol,理论pi为5.91。
19.图1k示出了编码示例性密码子优化的可溶性人dectin
‑
3(sdectin
‑
3)的核酸序列(seq id no:11)。人sdectin
‑
3 dna序列是由大肠杆菌中的载体pet
‑
45b 表达。9个密码子的载体pet
‑
45b 序列以及hist标签加有方框,起始密码子加有下划线。用于克隆到pet
‑
45b bamhi(ggatcc)(seq id no:24)和paci(ttaattaa)(seq id no:21)中的位点分别加有下划线。肠激酶加工位点的密码子是以小写字体显示。gly ser(g,s)柔性接头残基的密码子以粗体显示,并且反应性lys(k)残基(aag)以粗体显示,赖氨酸以斜体显示。来自clec4d人dectin
‑
3基因的密码子优化的sdectin
‑
3(genbank登记号nm_080387)以纯文本形式显示。ala密码子(gct)和终止密码子taa和tta加有下划线,终止密码子以粗体显示。替代基因名称为hssdectin3lyshis。核苷酸序列长度为628个碱基对,编码长度为207个氨基酸的多肽。编码示例性密码子优化的人sdectin
‑
3的核酸被克隆到pet
‑
45b 中。
20.图1l示出了由seq id no:11编码的氨基酸序列(seq id no:12)。此多肽包含人dectin
‑
3蛋白。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。肠激酶加工位点为以小写字体显示。gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示,赖氨酸以斜体显示。人sdectin
‑
3氨基酸残基(genbank登记号np_525126)以纯文本形式显示(seq id no:12的氨基酸35
‑
207),结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和pacl位点放置于框架中。该蛋白质的长度为207个氨基酸,mw为23,662g/mol,理论pi为7.64。
21.图1m示出了编码与venus的n末端部分融合的示例性密码子优化的可溶性小鼠dectin
‑
1(sdectin
‑
1)的核酸序列(seq id no:13)。应当理解,不包含膜结构域的可溶性小鼠dectin
‑
1氨基酸序列可与来自另一物种的可溶性dectin
‑
1氨基酸序列,例如,如seq id no:8中列出的人dectin
‑
1氨基酸序列或其片段互换。来自小鼠或其他物种的dectin
‑
2和dectin
‑
3氨基酸序列或其片段也可用于本文所述的任何构建体以检测真菌感染。mmsdectin1vyn是在pet
‑
45b 中表达的密码子优化的dna序列,编码一半的bifc诊断物。序列:1,168个碱基对,其中1,161个碱基对编码长度为385个氨基酸的蛋白质。9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线。克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线。短gly ser gly柔性接头残基的密码子以粗体显示,接着是小写字母表示的编码venus vyn t154m序列(它是从genbank登记号aka95335修饰的突变venus蛋白的n末端半部)的核苷酸序列(465个碱基对),再接着是以粗体显示的长glyser间隔区(其中反应性赖氨酸以斜体显示),再接着是长度为528个残基
的小鼠sdectin
‑
1序列(以纯文本显示)(clec7a,genbank登记号aas37670.1),结束于终止密码子taa和tta(以粗体显示)。
22.图1n示出了由seq id no:13编码的氨基酸序列(seq id no:14)。此多肽包含小鼠mmsdectin1vyn蛋白。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no 22)亲和标签加有方框。第一个gly ser(gs)柔性接头以粗体显示,接着是以小写字母表示的venus荧光蛋白vyn的c末端部分,代表具有t154m突变的venus残基1
‑
155(seq id no:14的氨基酸15
‑
169)。这之后是长glyser间隔序列,该序列含有以斜体显示的用于偶联至脂质载体的反应性lys(k)残基,然后是以纯文本显示的小鼠sdectin
‑
1氨基酸残基(seq id no:14的氨基酸211
‑
387)。该多肽的长度为387个氨基酸,mw为42,181.62g/mol,预测pi为6.55。
23.图1o示出了编码与venus的c末端部分融合的示例性密码子优化的可溶性小鼠dectin
‑
1(sdectin
‑
1)的核酸序列(seq id no:15)。mmsdectin1vc是可在pet
‑
45b 中表达的密码子优化的dna序列,编码一半的bifc诊断物。长度:955,其中948编码长度为316个氨基酸的蛋白质。9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线。开始和结束处的克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线。gly ser(g,s)柔性接头残基的密码子以粗体显示。然后,用于venus的c末端氨基酸编码序列的编码序列(252个核苷酸)(a.a.残基155至238,由genbank登记号aka95335修饰并且含有t154m突变的蛋白质)以小写字母显示。然后,长glyser柔性接头以粗体显示,其中用于偶联至脂质载体的反应性赖氨酸以斜体显示。这之后是长度为528个残基的小鼠sdectin
‑
1序列(clec7a,genbank登记号naas37670.1)(以纯文本显示),结束于以粗体显示的终止密码子taa和tta。
24.图1p示出了由seq id no:15编码的氨基酸序列(seq id no:16)。此多肽包含小鼠mmsdectin1vc蛋白。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框。第一gly ser(gs)柔性接头序列以粗体字体显示。然后,代表venus残基155至238(seq id no:16的氨基酸15
‑
98)的venus荧光蛋白的c末端半部(genbank登记号aka95335)以小写字母显示。然后,含有反应性lys(k)残基(斜体)的长glyser柔性间隔区以粗体显示。小鼠sdectin
‑
1氨基酸残基以纯文本形式显示(氨基酸140
‑
316),接着是用于将终止密码子放置在框架中的c末端ala残基(a)。该多肽的长度为316个氨基酸,mw为34,081.21g/mol,预测pi为6.8。
25.图1q:mmdec2vyn,也是pet
‑
45b中表达的mmdec2vyn密码子优化的dna序列(seq id no:17)。长度:577bp,9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线,克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线,gly,ser(g,s)柔性接头残基的密码子以粗体显示,反应性lys(k)残基aag以斜体显示,来自小鼠dectin 2基因clec6a的针对大肠杆菌表达进行密码子优化的sdectin
‑
2序列以纯文本形式显示,后面是具有15个残基的富含gly ser的柔性间隔区,接着是以小写字母显示的venus vyn t154m(即,从aka95335修饰的突变venus蛋白的n末端半部)的长度为465个核苷酸的编码序列,一个ala(a)密码子gct和两个终止密码子taa和tta加有下划线,终止密码子以粗体显示。替代基因名称:mmsdectin2lyshis,长度:1084bp,少了用于终止和以下酶切位点的7bp,编码359个残基的蛋白质。但从genscript订购从kpnl位点ggtacc(seq id no:23)开始的较短1057bp型式用于亚克隆到pet
‑
45b 中。
26.图1r:正在合成的最终mmsdec2vyn蛋白(seq id no:18)。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框,gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示),166个小鼠sdectin
‑
2氨基酸残基以纯文本形式显示(seq id no:18的氨基酸23
‑
188),后面是具有15个残基的富含gly ser的柔性间隔区,具有t154m突变的venus残基1
‑
155(seq id no:18的氨基酸204
‑
359),结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和paci位点放置于框架中。总共359个氨基酸,mw 40,198g/mol。pi 6.04。o.d.2801.940/mg/ml。
27.图1s:mmdec2vc,也是在pet
‑
45b中表达的mmdec2vc密码子优化的dna序列(seq id no:19)。长度:577bp,9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加下划线,克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线,gly,ser(g,s)柔性接头残基的密码子以粗体显示且反应性lys(k)残基aag以斜体显示,来自小鼠dectin 2基因clec6a的针对大肠杆菌表达进行密码子优化的sdectin
‑
2序列以纯文本形式显示,后面是具有15个残基的富含gly ser的柔性间隔区,然后是以小写字母显示的用于venus的c末端氨基酸编码序列(a.a.残基155至238,从aka95335修饰的蛋白)的长度为252个核苷酸的编码序列,一个ala(a)密码子gct和两个终止密码子taa和tta加有下划线,终止密码子以粗体显示。替代基因名称:mmsdectin2lyshis。长度:871bp,少了用于终止密码子和切割位点的7bp,编码288个残基的蛋白质。但从genscript订购从kpnl位点ggtacc(seq id no:23)开始的较短844bp型式用于亚克隆到pet
‑
45b 中。
28.图1t:正在合成的最终mmsdec2vc2蛋白(seq id no:20)。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框,gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示),166个小鼠sdectin
‑
2氨基酸残基以纯文本显示(seq id no:20的氨基酸23
‑
188),后面是具有15个残基的富含gly ser的柔性间隔区,venus残基155至238(seq id no:20的氨基酸204
‑
288),接着是框架中的两个终止密码子和paci位点。总共288个氨基酸,mw为32,098g/mol。pi 6.16,o.d.280 2.02od/mg/ml。
29.图2示出了在细胞提取物中的和纯化后的可溶性dectin
‑
1(sdectin
‑
1)的sds page分析。sdectin
‑
1蛋白是在不进行iptg诱导的情况下由pet
‑
45b质粒在于luria肉汤中过夜生长的大肠杆菌bl21菌株中产生的,将其溶解在guhcl缓冲液中,通过ni
‑
nta树脂纯化并通过sds page检查。提取到还含有还原剂β巯基乙醇和triton
‑
x100去污剂的缓冲液中,相对于不含它们的缓冲液(右泳道)大幅提高了从不溶性包涵体(中心泳道)中的回收率。在用考马斯蓝染色的12%丙烯酰胺凝胶上检查蛋白质。用尿素缓冲液提取细胞产生很少的蛋白质。
30.图3是负载有sdectin
‑
1、两性霉素b和罗丹明的脂质体dec
‑
amb
‑
ll模型的示意图。将两性霉素b(amb,蓝色椭圆结构)嵌入到直径为100nm的脂质体的脂质双层中。将sdectin
‑
1(dec,绿色球状结构)与脂质载体dspe
‑
peg偶联,并也将dspe
‑
peg
‑
dec和红色荧光dhpe
‑
罗丹明(红色星形)都插入到脂质体膜中。sdectin
‑
1、罗丹明、amb和脂质体脂质的摩尔比为1:2:13:100。两个sdectin
‑
1单体(两个dsep
‑
peg
‑
dec分子)必须在膜中漂浮在一起,以与细胞壁β
‑
葡聚糖(红糖部分)结合。检查的两个脂质体对照是含有相等μg量的65kda bsa代替22kda sdectin
‑
1的bsa
‑
amb
‑
ll(即,0.33bsa:2:13:100摩尔比)和没有任何蛋白质涂层的
ambisome样脂质体(amb
‑
ll)(0:2:13:100摩尔比)。根据这些摩尔比、直径为100nm的脂质体的表面积以及已公布的每106nm2脂质双层5
×
106个脂质分子的估计值,估计每个脂质体中有大约3,000个罗丹明分子,每个dec
‑
amb
‑
ll中有大约1500个sdectin
‑
1单体。
31.图4a
‑
f显示sdectin
‑
1包被的dec
‑
amb
‑
ll与发芽中的烟曲霉(a.fumigatus)的膨大分生孢子和芽管强烈结合,而amb
‑
ll则不能如此。a.罗丹明红色荧光dec
‑
amb
‑
ll结合的烟曲霉(a.fumigatus)膨大分生孢子(白色箭头)和幼苗芽管。b.罗丹明红色荧光ambisome样amb
‑
ll没有结合。即使如此图像中所示增强了红色通道,也未检测到脂质体。最小的红点代表基于荧光观察到的各个100nm直径脂质体(橙色箭头)。大簇脂质体形成更亮的红色染色区。c至f。c和d是用dec
‑
amb
‑
ll染色的。e和f是用bsa
‑
amb
‑
ll染色的。a和b。让细胞在vmm 1%葡萄糖中在37℃下生长8小时。在脂质体稀释缓冲液ldb中进行标记60分钟。将所有三种脂质体制品均以1:100稀释,使得脂质体sdectin
‑
1和bsa蛋白的最终浓度为1ug/100ul。在仅针对细胞质egfp表达的绿色通道中以及在脂质体的红色通道中观察到幼苗。a和b是在油浸下以63x拍摄的。c至f是在倒置荧光显微镜上以20x拍摄的。
32.图5a
‑
f显示sdectin
‑
1包被的dec
‑
amb
‑
ll结合成熟烟曲霉(a.fumigatus)细胞的膨大分生孢子和菌丝,而非靶向ambisome样amb
‑
ll则不能如此。让烟曲霉(a.fumigatus)分生孢子在vmm 1%葡萄糖中在37℃下发芽并生长16小时,然后用荧光脂质体染色。a.至d.是用1:100稀释的罗丹明红色荧光dec
‑
amb
‑
ll染色,使sdectin
‑
1为1ug/100ul;e.和f.是用等量的红色荧光amb
‑
ll染色60分钟。a.单独的dic图像。b:组合的dic和红色荧光图像。a和b.显示罗丹明荧光dec
‑
amb
‑
ll与膨大分生孢子(白色箭头)和菌丝结合。c至f检查了细胞质绿色egfp与脂质体的红色荧光。最小的红点代表因其强荧光可见的各个100nm脂质体(橙色箭头)。c和d显示几乎所有分生孢子和大多数菌丝都被dec
‑
amb
‑
ll染色。e和f显示amb
‑
ll没有结合。a和b是在油浸下以63x拍摄,并合并了七个堆叠的图像。c至f是在倒置荧光显微镜上以20x拍摄的。
33.图6a
‑
f显示sdectin
‑
1包被的脂质体(dec
‑
amb
‑
ll)与白色念珠菌(candida albicans)和新型隐球菌(cryptococcus neoformans)细胞强结合。a、c和e是用在ldb中1:100稀释的dec
‑
amb
‑
ll标记的白色念珠菌(candida albicans)菌株sc5314和新型隐球菌(cryptococcus neoformans)菌株h99的明场图像。b、d和f是明场图像和红色荧光图像的组合,显示罗丹明标记的dec
‑
amb
‑
ll与这些细胞强结合。普通的、未包被的amb
‑
ll与这些细胞没有可检测到的结合。a和b是在油浸下以63x拍摄,c至f是在倒置荧光显微镜上以20x拍摄。
34.图7:sdectin
‑
1包被的dec
‑
amb
‑
ll与烟曲霉(a.fumigatus)的结合频率比对照amb
‑
ll或bsa
‑
amb
‑
ll高几个数量级,并且结合被可溶性β
‑
葡聚糖抑制。让4,500个烟曲霉(a.fumigatus)分生孢子的样本在vmm 1%葡萄糖中在37℃下发芽并生长36小时,在福尔马林中固定或检查活细胞,并在脂质体稀释缓冲液中1:100稀释脂质体的条件下孵育1小时。洗掉未结合的脂质体。如图4和图5所示,以20x拍摄红色荧光图像的多个视野。每个摄影视野包含大约25个膨大分生孢子和广泛的菌丝网络。a、b、c。标记福尔马林固定细胞。d、e、f。标记活细胞。g、h、i。1mg/ml海带多糖(可溶性β
‑
葡聚糖)与作为对照的1mg/ml蔗糖相比对固定细胞的dec
‑
amb
‑
ll标记的抑制作用。a、d和g。对红色荧光脂质体和脂质体簇的数量进行计数,对每个场取平均值,并以log10标度绘制。在每个棒上方和垂直轴上标示数字平均值。显示了标准误差。b、c、e、f、h和i中示出了用于构建棒图的脂质体的场实例。
35.图8a
‑
g显示在接近amb的ed50的amb浓度下,dec
‑
amb
‑
ll杀灭或抑制烟曲霉(a.fumigatus)生长的效率比amb
‑
ll要高很多。让4,500个烟曲霉(a.fumigatus)分生孢子的样本在96孔微量滴定板中在vogel最低培养基(vmm 1%葡萄糖)中发芽和生长,并同时用脂质体稀释缓冲液或向生长培养基递送指定浓度的amb(a
‑
d 3um amb,e.0.09um amb,f.0.18um amb)的脂质体制品处理。使用cell titer blue(ctb)试剂(a和c)(其测量总细胞酯酶活性)或通过菌丝长度(b和d)或通过发芽百分比(e和f)估计存活力和生长。减去培养基中含有ctb但缺少细胞和脂质体的孔的背景荧光。指示了标准误差。b和d中的插图示出了对经amb
‑
ll和dec
‑
amb
‑
ll处理的样本测定的菌丝长度的实例。b和d中一个菌丝长度单位等于5微米。a和b以及c和d比较了使用在不同缓冲液中制备和储存的两个完整生物学重复实验的结果。在a和b中,脂质体是在rn#5缓冲液(0.1m nah2po4、10mm三乙醇胺ph 7.2、1m l
‑
精氨酸、100mm nacl、5mm edta、5mm bme(β
‑
巯基乙醇))中制备,c至f是在rn#5缓冲液(0.1m nah2po4、10mm三乙醇胺ph 7.2、1m l
‑
精氨酸、100mm nacl、5mm edta、5mm bme)中制备,细胞生长56小时(c、d)或36小时(a、b、e、f)。g.基于分生孢子发芽百分比的剂量反应曲线。将分生孢子和递送不同浓度amb的脂质体在vmm 葡萄糖中一起平板接种于微量滴定板的孔中。在37℃下8小时后,将已发芽的分生孢子百分比量化。
36.图9a
‑
b显示,基于细胞质gfp荧光,在接近和低于amb的ed
50
的amb浓度下,dec
‑
amb
‑
ll比ambisome样amb
‑
ll更有效地抑制烟曲霉(a.fumigatus)细胞的生长。使4,500个烟曲霉(a.fumigatus)分生孢子的样本在96孔微量滴定板中于vogel最低培养基(vmm) 1%葡萄糖中发芽和生长,并将该样本用脂质体稀释缓冲液对照或递送2um amb(a.)和0.67amb(b.)的负载amb的脂质体制品的稀释液处理,并在37℃下孵育36小时。基于烟曲霉(a.fumigatus)a1163菌株中egfp报告基因的细胞质绿色荧光量测定细胞完整性。减去含培养基但缺少细胞和脂质体的孔的荧光背景。脂质体在使用前储存在rn#5缓冲液中,并用脂质体稀释缓冲液ldb稀释。
37.图10显示sdectin
‑
1包被的dec
‑
amb
‑
ll对hek293细胞的毒性低于未包被的ambisome样amb
‑
ll。将人胚肾hek293细胞用脂质体处理2小时,然后洗去脂质体。然后,将细胞再孵育16小时并进行测定。脂质体向培养基递送总共30或15um的amb。celltiter blue酯酶测定估计了细胞存活力和存活率。
38.图11示出了用于检测受试者或所述受试者的样本中真菌β
‑
葡聚糖和甘露聚糖多糖的固定化示例性dectin包被脂质体的示意图模型。dec
‑
bifc
‑
l是脂质体,该脂质体包被有500个融合至绿色荧光蛋白venus vyn155(例如dec
‑
vn
‑
)的n末端半部的sdectin单体(蓝色)以及500个融合至venus vc155(dec
‑
vc
‑
)的c末端半部的sdectin单体,其将快速识别并结合真菌β
‑
葡聚糖或甘露聚糖以形成sdectin二聚体,从而导致形成组装的venus,其将产生强烈的绿色双分子荧光互补(bifc)信号。这些脂质体仅在接触真菌细胞或释放的β
‑
葡聚糖或甘露聚糖时才会产生绿色荧光信号。当这些脂质体通过例如生物素
‑
链霉亲和素结合附着于不溶性基质上时,它们可以用于使用标准荧光仪器检测大体积血清中的低浓度真菌多糖。
39.图12是用于检测真菌β
‑
葡聚糖和甘露聚糖的dec
‑
hrp
‑
l(dectin包被的脂质体)的模型的示意图。将sdectin(dec,绿色球状结构)和辣根过氧化物酶(hrp,六边形)与脂质载体dspe
‑
peg或dspe偶联并插入到脂质体膜中。sdectin、hrp、peg和脂质体脂质的摩尔比为
约1:1:13:100。两个sdectin单体(两个dsep
‑
peg
‑
dec分子)必须在膜中漂浮在一起,以与细胞壁β
‑
葡聚糖或甘露聚糖(红糖部分)结合。在这些脂质体与固定的真菌样本结合后,洗掉多余的未结合脂质体。然后,添加hrp酶活性的底物(4
‑
氯
‑1‑
萘酚和过氧化物)。添加底物后,测量hrp产生的紫色沉淀以确定结合的dec
‑
hrp
‑
l的总量和真菌多糖的总量。
40.图13a示出了修饰的小鼠sdectin
‑
2 dna mmsdectin2lyshis dna序列(seq id no:3)。mmsdectin2lyshis的密码子优化的dna序列被克隆到pet
‑
45b中。ncbi bankit#mn104679。长度:577bp,9个密码子的载体pet
‑
45b 序列加有方框,起始密码子加有下划线,克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线,gly,ser(g,s)柔性接头残基的密码子以粗体显示,并且反应性lys(k)残基aag粗体以斜体显示,来自小鼠dectin 2基因clec6a的针对大肠杆菌表达进行密码子优化的sdectin
‑
2序列为纯文本形式显示,结束于paci位点中以粗体显示的一个ala(a)密码子gct和两个终止密码子taa和tta。编码序列和两个终止密码子的长度:574bp,编码189个残基的蛋白质。替代基因名称:mmsdectin2lyshis。
41.图13b示出了在大肠杆菌中合成的sdectin
‑
2(dec2)蛋白(seq id no:4)。来自pet
‑
45b 的n末端氨基酸肽序列和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框,gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示),166个小鼠sdectin
‑
2氨基酸残基以纯文本形式显示,结束于以粗体显示的c末端ala残基(a)。总共189个氨基酸,mw为21,763.29g/mol,理论pi为6.33。sdectin
‑
2序列代表来自天然小鼠dectin
‑
2序列的a.a.氨基酸残基44至209。替代蛋白质名称:mmsdectin2lyshis蛋白质。
42.图14示出了纯化的sdectin
‑
2的sds page分析。在考马斯蓝染色的12%凝胶上,通过sds page检查了不表达和表达sdectin
‑
2和纯化sdectin
‑
2蛋白的大肠杆菌bl21粗提取物。左侧指示了分子量标记和22kda的修饰sdectin
‑
2的近似分子量。
43.图15示出了负载罗丹明和两性霉素b的sdectin
‑
2包被的脂质体模型。将两性霉素b(amb,蓝色卵形结构)嵌入到100nm直径脂质体的脂质双层中。将sdectin
‑
2(dec2,绿色球状结构)与脂质载体dspe
‑
peg偶联,并将dec2
‑
peg
‑
dspe和红色荧光dhpe
‑
罗丹明(红色星形)通过其脂质部分dspe和dhpe插入到脂质体膜中。sdectin
‑
2、罗丹明、amb和脂质体脂质的摩尔比为1:2:11:100。两个sdectin
‑
2单体(两个dec2
‑
peg
‑
dspe分子)必须在膜中漂浮在一起以与真菌甘露聚糖(红糖部分)结合。根据这些摩尔比、直径100nm脂质体的表面积以及已公布的每106nm2脂质双层5
×
106个脂质分子的估计值,计算出在每个dec2
‑
amb
‑
ll中,每个脂质体中存在大约1,500个sdectin
‑
1单体、3,000个罗丹明分子和大约16,500个amb分子。
44.图16a
‑
f示出了结合于与多种多样的形态的白色念珠菌(candida albicans)细胞缔合的胞外基质的sdectin
‑
2包被脂质体dec2
‑
amb
‑
ll。a:酵母细胞。通过微分干涉对比显微镜(绿色,dic)和罗丹明荧光标记的dec2
‑
amb
‑
ll(红色)突出显示细胞。b&c:假菌丝(ps
‑
hyp)和菌丝(hyp)。通过细胞的内源性gfp荧光突出显示细胞。dec2
‑
amb
‑
ll以大簇形式与细胞外基质(ex)结合。d、e、f:成熟菌丝。d:明场显微镜示出了e中染色细胞菌丝周围的细胞外基质。e:组合明场图像和脂质体的红色荧光。f:与e中图像平行的额外图像再次强调了基质的典型染色。将所有细胞用1:200稀释到ldb2缓冲液中的dec2
‑
amb
‑
ll(例如,0.5μg sdectin
‑
2/100μl)染色1小时。指示了dec2
‑
amb
‑
ll染色(ex )或未染色或弱染色(ex
‑
)的细
胞外基质。箭头指示单个脂质体。照片是在油浸下以63x放大倍数拍摄的。几项独立的真菌细胞标记研究得到类似的图像。
45.图17a
‑
g示出了结合于与新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)细胞缔合的胞外基质的dec2
‑
amb
‑
ll。罗丹明红色荧光dec2
‑
amb
‑
ll以大簇形式结合于细胞外基质(ex),很少结合于这两个物种的细胞壁。a、b、c和d:新型隐球菌(c.neoformans)。用dec2
‑
amb
‑
ll和小鼠单克隆抗体18b7将酵母细胞与荚膜葡糖醛木甘聚糖(gxm)和二级山羊抗小鼠抗体alexa488(绿色)共染色。将所有细胞用1:200稀释到ldb2中的dec2
‑
amb
‑
ll(例如,0.5μgsdectin
‑
2/100μl)染色1小时。a:细胞的明场图像。b:gxm特异性抗体染色细胞的绿色荧光图像。c:b和d的合并荧光图像。d:红色荧光dec2
‑
amb
‑
ll。e、f和g:烟曲霉(a.fumigatus)。e和f:生长10小时并用dec2
‑
amb
‑
ll染色的幼苗的明场和组合荧光图像。g:生长24小时的成熟菌丝。将这两个物种用1:200稀释到ldb2中的dec2
‑
amb
‑
ll(例如,0.5μg sdectin
‑
2/100μl)染色1小时。指示了dec2
‑
amb
‑
ll染色(ex )或未染色或弱染色(ex
‑
)的细胞外基质或者未用18b7或dec2
‑
amb
‑
ll(ex
‑
/
‑
)染色的细胞外基质。照片是在油浸(板a
‑
f)下以63x或20x(板2g)放大倍数拍摄的。三项独立的真菌细胞标记研究得到类似的图像。
46.图18a
‑
d显示dec2
‑
rhod(罗丹明标记的dec2)和dec2
‑
amb
‑
ll以相似的模式与烟曲霉(a.fumigatus)菌丝细胞周围的胞外多糖基质结合。使烟曲霉(a.fumigatus)分生孢子在显微镜室载玻片上在vmm 1%葡萄糖 0.5%bsa中以低密度发芽,并在37℃下生长24小时并固定细胞,然后用罗丹明标记的dec2蛋白dec2
‑
rhod或dec2包被的脂质体染色一小时。a&b:dec2
‑
rhod。c&d:dec2
‑
amb
‑
ll。细胞是以20x拍摄,拍摄了微分干涉对比图像(dic,板a和c)以及组合dic和红色荧光图像(板b和d)。由于细胞高度分散,并且我们希望在一块板上示出几个示例细胞,因此这些图像是由从分开的摄影场拍摄并彼此相邻放置的细胞图像制成的合成图(参见移动到共同场中细胞的虚线轮廓)。图像代表90%的被检查真菌细胞集落。
47.图19a
‑
i显示sdectin
‑
2包被的dec2
‑
amb
‑
ll与白色念珠菌(candida albicans)、新型隐球菌(cryptococcus neoformans)和烟曲霉(a.fumigatus)细胞的结合效率比对照amb
‑
ll高一到两个数量级。将固定真菌细胞的密集场与脂质体在脂质体稀释缓冲液ldb2中的1:200稀释液(例如,0.5μg sdectin
‑
2/100μl)一起孵育1小时。一小时后洗掉未结合的脂质体。红色荧光图像的多个场是以20x拍摄,并估计图像j中荧光的面积。棒图的右侧示出了拍摄图像的实例。a、b和c:c.白色念珠菌(candida albicans),d、e和f:c.新型隐球菌(cryptococcus neoformans)。g、h和i:烟曲霉(a.fumigatus)。在a、d和g中,给出了与平均值的标准误差,并指示了倍数差异和p值以将dec2
‑
amb
‑
ll的结合与amb
‑
ll的结合区分开。
48.图20a
‑
h示出了dec2
‑
amb
‑
ll结合的特异性、稳定性和速率。a、b和c:结合特异性。白色念珠菌(c.albicans)的dec
‑
amb
‑
ll标记受可溶性酵母甘露聚糖抑制,但不受蔗糖或海带多糖的抑制。在1小时的染色程序中,每种多糖以10mg/ml加入。d、e和f:结合稳定性。将用于产生图3a
‑
c中的数据的用dec2
‑
amb
‑
ll和对照脂质体染色的白色念珠菌(candida albicans)细胞板留在pbs中,在暗处储存2个月,重新拍照,并重新量化脂质体结合的面积。g&h:结合速率。白色念珠菌(c.albicans)的成熟培养物由在24孔微量滴定板的表面上在vmm 20%fbs中生长的一些假菌丝和菌丝组成,被固定、封闭并用dec2
‑
amb
‑
ll处理指定时间。在所有三个实验中,dec2
‑
amb
‑
ll按1:200w/v ldb2(0.5μg,在100μl sdectin
‑
2中)稀释,并用ldb2洗涤4次。对于每个时间点,在倒置荧光显微镜上以20x放大倍数拍摄多张红色
荧光图像,并估计红色荧光脂质体染色的平均面积。a、d、g和h中示出了平均值的标准误差。在a和d中,在每个棒上方和垂直轴上面指示了所检查场的数字平均值和数量。在a和d中,指示了dec
‑
amb
‑
ll相对于甘露聚糖抑制(a)或相对于amb
‑
ll(d)的性能的倍数差异和p值。这些结果代表了两个生物学重复物。
49.图21a
‑
d示出了sdectin
‑
2包被的负载两性霉素b的脂质体对白色念珠菌(c.)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)的抑制和杀灭。a:白色念珠菌(c.albicans)与dec2
‑
amb
‑
ll。在96孔聚苯乙烯微量滴定板上在rpmi培养基 0.5%bsa中生长的处于假菌丝和早期菌丝阶段的细胞。将细胞用向如所示的培养基中递送1.0、0.5、0.25和0.12μm amb的脂质体处理30分钟,用培养基洗涤两次,再让该细胞生长16小时,然后使用celltiter
‑
blue(ctb)试剂测定它们的代谢活性。b:新型隐球菌(c.neoformans)与dec2
‑
amb
‑
ll。使新型隐球菌(c.neoformans)细胞在液体ypd培养基 0.5%bsa中在剧烈摇动下生长2小时,并用向如所示的培养基中递送0.4、0.2或0.1μm amb的脂质体处理4小时或过夜。将细胞稀释,平板接种在ypd培养基上,并从多个平板上对菌落形成单位(cfu)进行计数。c:烟曲霉(a.fumigatus)与dec2
‑
amb
‑
ll。使分生孢子在96孔聚苯乙烯微量滴定板中的vmm 葡萄糖 0.5%bsa中发芽9小时,用向如所示的培养基中递送0.5和0.25μm amb的脂质体处理2小时,用培养基洗涤两次,使该分生孢子生长过夜,然后在不含酚红指示剂的rpmi 0.5%bsa中使用ctb试剂测定它们的代谢活性。对照孔过度生长,菌丝从培养基中伸出,因此代谢活性低,甚至虽然这些孔中有更多细胞,但产生的信号也较低。d:烟曲霉(a.fumigatus)与dec1
‑
amb
‑
ll。测定条件与c中的测定相似,不同之处在于首先将脂质体稀释到ldb1缓冲液(pbs 0.5%bsa 1mm bme)中,然后才稀释到生长培养基中。对于a、c和d中的ctb测定,减去来自与ctb试剂一起孵育的培养基的荧光背景。示出了所有值的标准误差,并估计了倍数差异和p值,从而比较了amb
‑
ll与dec2
‑
amb
‑
ll的性能。两个或多个生物学重复物得出相似的结果。
50.图22a
‑
b示出了肺曲霉病的两种免疫抑制模型和用于检查dectin
‑
2靶向的负载抗真菌的脂质体功效的时间线。a:类固醇模型。b:白细胞减少模型。
51.图23示出了dectin靶向的负载抗真菌药物的脂质体。包被有例如dectin
‑
1或dectin
‑
2的碳水化合物识别结构域的负载抗真菌药物的脂质体特异性地结合至侵袭性真菌细胞和生物膜,并主动地向真菌细胞递送抗真菌药物。靶向脂质体与真菌细胞(右侧)的结合效率比非靶向负载药物的脂质体(例如,)或洗涤剂增溶的抗真菌药物(左侧)相比高出几个数量级。因此,靶向脂质体与非靶向脂质体相比,对动物细胞具有较低的相对亲和力,并在较低的总药物浓度下向真菌细胞递送高得多的剂量的抗真菌药物。通过减少杀灭真菌细胞所需的有效剂量,它们对动物细胞的毒性较小。
52.图24a
‑
b显示烟曲霉(a.fumigatus)菌株cea10的分生孢子快速发芽从而在cd1小鼠的肺中建立感染中心。在第0天(图21),给予cd1 swiss小鼠2
×
106个cea10分生孢子的口咽接种物。在第2天,感染后48小时,对小鼠实施安乐死,将手动制备的受感染肺的各个肺叶的薄切片(例如,厚度约0.5mm)固定在福尔马林中,并用钙荧光白染色真菌甲壳质一小时。在leica dm6000复合显微镜上,使用dapi滤光片组ex
a360
/em
a470
拍摄落射荧光摄影图像。a:大多数切片显示出一个大感染中心,其中有数百个染色的菌丝。以5x放大倍数拍摄照片。b:感染中心由菌丝延伸簇组成。没有观察到未发芽的分生孢子。以20x放大倍数拍摄照片。
53.图25a
‑
d显示,基于免疫抑制介导的曲霉病的类固醇小鼠模型,dec2
‑
amb
‑
ll在减少肺部真菌负荷方面比amb
‑
ll明显更有效。将免疫受到抑制的cd1 swiss小鼠(类固醇模型图21a)用2
×
106个烟曲霉(a.fumigatus)分生孢子cea10菌株(第0天)感染,并在次日(第1天)用递送0.2mg amb/kg小鼠体重的dec2
‑
amb
‑
ll或样amb
‑
ll脂质体或者用用于稀释脂质体的缓冲液(缓冲液对照)处理(参见图21a中的时间线)。将存活到第4天的每个处理组中三只小鼠安乐死,并通过两种独立的方法确定每个肺的真菌负荷。a
‑
c:菌落形成单位(cfu)。a:比较三个处理组每个肺的cfu平均数的棒图。b和c分别是以20x放大倍数拍摄的针对amb
‑
ll和dec2
‑
amb
‑
ll处理组检查的细胞的场的示例摄影图像。cfu是通过在富生长培养基上平板接种经匀浆化的肺组织、将平板孵育过夜并对真菌细胞微菌落进行计数来进行估计的。标准误差用线和须线表示。d:rdna的相对数量。烟曲霉(a.fumigatus)rdna基因间隔区(igs)的相对数量(rq)是通过对来自与图24a中测定的相同的三组三个肺的肺匀浆平行样本进行qpcr来确定的。将数据针对一只具有最高水平的缓冲液对照小鼠(即,该小鼠的rq设置为1.0)中烟曲霉(a.fumigatus)rdna的水平作归一化。
54.图26a
‑
b显示,基于免疫抑制介导的曲霉病的白细胞减少小鼠模型,dec2
‑
amb
‑
ll在减少肺部真菌负荷方面比amb
‑
ll明显更有效。将免疫受到抑制的cd1 swiss小鼠(模型图21b)用5
×
105个烟曲霉(a.fumigatus)分生孢子cea10菌株(第0天)感染,并在次日用递送0.2mg amb/kg小鼠体重的dec2
‑
amb
‑
ll或样amb
‑
ll脂质体或者用用于稀释脂质体的缓冲液(对照)处理(参见图21b中的时间线)。将每个处理组的三只小鼠安乐死,并通过两个独立的方法确定每个肺的真菌负荷。a:烟曲霉(a.fumigatus)菌落形成单位(cfu)。比较三个处理组每个肺的cfu平均数的棒图。标准误差用线和须线表示。b:真菌细胞rdna的相对数量。烟曲霉(a.fumigatus)rdna基因间隔区(igs)的相对数量(rq)是通过对来自与a中测定的相同的三组三个肺的肺匀浆平行样本进行qpcr来测定的。将数据针对一只具有最高水平的对照小鼠(即,该小鼠的rq为1.0)中烟曲霉(a.fumigatus)rdna的水平作归一化。
55.图26a
‑
b显示,基于免疫抑制介导的曲霉病的白细胞减少小鼠模型,dec2
‑
amb
‑
ll在减少肺部真菌负荷方面的效率比amb
‑
ll高两个数量级。将免疫受到抑制的cd1 swiss小鼠(模型图21b)用5
×
105个烟曲霉(a.fumigatus)分生孢子cea10菌株(第0天)感染,并在次日(第1天)用递送0.2mg amb/kg小鼠体重的dec2
‑
amb
‑
ll或样amb
‑
ll脂质体或者用用于稀释脂质体的缓冲液(缓冲液对照)处理(参见图21b中的时间线)。将每个处理组的三只小鼠安乐死,并通过两个独立的方法确定每个肺的真菌负荷。a:烟曲霉(a.fumigatus)菌落形成单位(cfu)。比较三个处理组每个肺的cfu平均数的棒图。标准误差用线和须线表示。b:真菌细胞rdna的相对数量。烟曲霉(a.fumigatus)rdna基因间隔区(igs)的相对数量(rq)是通过对来自与a中测定的相同的三组三个肺的肺匀浆平行样本进行qpcr来测定的。将数据针对一只具有最高水平的对照小鼠(即,该小鼠的rq为1.0)中烟曲霉(a.fumigatus)rdna的水平作归一化。
56.图27a
‑
d示出了用于真菌甘露聚糖bifc检测的dectin
‑
2 venus融合的dna编码序列和蛋白序列。a和c:分别为mmdec2vyn和mmdec2vc的dna序列。b和d:分别为dec2
‑
vyn和dec2
‑
vc的蛋白序列。
57.a:在pet
‑
45b中表达的mmdec2vyn密码子优化的dna序列(seq id no:17)。长度:577bp,9个密码子的载体pet
‑
45b 序列加有方框(其中起始密码子加有下划线),克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线,gly,ser(g,s)柔性接头残基的密码子以粗体显示并且反应性lys(k)残基aag以斜体显示,来自小鼠dectin 2基因clec6a的针对大肠杆菌表达进行密码子优化的sdectin
‑
2序列以纯文本形式显示,后面是具有15个残基的富含gly ser的柔性间隔区,接着是以小写字母显示的venus vyn t154m(从aka95335修饰的突变venus蛋白的n末端半部)的长度为465个核苷酸的编码序列,一个ala(a)密码子gct和两个终止密码子taa和tta加有下划线(其中终止密码子以粗体显示)。替代基因名称:mmsdectin2lyshis,长度:1084bp,少了终止和以下酶切位点的7bp(即,1077bp),编码359个残基的蛋白质。但从genscript订购的是从kpnl位点ggtacc(seq id no:23)开始的较短1057bp型式,用于亚克隆到pet
‑
45b 中。
58.b:正在合成的最终dec2
‑
vyn蛋白(seq id no:18)。来自pet
‑
45b 的n末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框,gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示),166个小鼠sdecetin
‑
1氨基酸残基以纯文本形式显示,后面是具有15个残基的的富含gly ser的柔性间隔区,接着是具有t154m突变的venus残基1
‑
155,结束于以粗体显示的c末端ala残基(a),其密码子用于将终止密码子和paci位点放置于框架中。总共359个氨基酸,mw 40,198g/mol。pi 6.04。o.d.2801.940/mg/ml。
59.c:mmdec2vc,也是在pet
‑
45b中表达的mmdec2vc密码子优化的dna序列(seq id no:19)。长度:577bp,9个密码子的载体pet
‑
45b 序列加有方框(其中起始密码子加有下划线),克隆位点kpni(ggtacc)(seq id no:23)和paci(ttaattaa)(seq id no:21)分别加有下划线,gly,ser(g,s)柔性接头残基的密码子以粗体显示并且反应性lys(k)残基aag以斜体显示,来自小鼠dectin 2基因clec6a的针对大肠杆菌表达进行密码子优化的sdectin
‑
2序列以纯文本形式显示,后面是具有15个残基的的富含gly ser的柔性间隔区,然后是以小写字母显示的用于venus的c末端氨基酸编码序列(a.a.残基155至238,从aka95335修饰的蛋白)的长度为252个核苷酸的编码序列,一个ala(a)密码子gct和两个终止密码子taa和tta加有下划线(其中终止密码子以粗体显示)。替代基因名称:mmsdectin2lyshis,长度:871bp,少了用于终止密码子和酶切位点的7bp,编码288个残基的蛋白质。但从genscript订购从kpnl位点ggtacc(seq id no:23)开始的较短844bp型式用于亚克隆到pet
‑
45b 。
60.d:正在合成的最终dec2
‑
vc蛋白(seq id no:20)。来自pet
‑
45b 的c末端氨基酸和(his)6(hhhhhh)(seq id no:22)亲和标签加有方框,gly ser(gs)柔性接头残基和反应性lys(k)残基以粗体显示(其中赖氨酸以斜体显示),166个小鼠sdecetin
‑
2氨基酸残基以纯文本形式显示,后面是具有15个残基的富含gly ser的柔性间隔区,venus残基155
‑
238,结束于以粗体显示的添加的c末端ala残基(a),该ala残基的密码子用于将终止密码子和paci位点放置于框架中。总共288个氨基酸,mw 32,098g/mol。pi 6.16,o.d.280 2.02od/mg/ml。
61.图28a和b示出了包被有与venus蛋白互补片段融合的dectin
‑
2的脂质体的蛋白质设计和模型,其用于检测含多糖的真菌甘露聚糖。a.所采用的诊断脂质体模型。包被有与绿色荧光蛋白venus vyn155的n末端片段(例如dec2
‑
vyn)融合的sdectin
‑
2单体(蓝色)的脂质体以及包被有与venus vc的c末端片段(dec2
‑
vc)融合的sdectin
‑
2单体的脂质体识别并
结合真菌甘露聚糖从而形成sdectin
‑
2二聚体,导致产生组装的venus蛋白和强烈的绿色双分子荧光互补(bifc)信号。这些真菌诊断脂质体仅在接触真菌细胞壁中的甘露聚糖、胞外多糖基质、生物膜或释放的含有甘露聚糖的可溶性多糖时才会产生绿色荧光信号。dec2
‑
vyn和dec2
‑
vc均与脂质载体dspe
‑
peg偶联并被插入到脂质体膜中。将dec2偶联的单体插入到脂质体中,使得总dec2蛋白和脂质体脂质处于1:100的摩尔比,或者具有1摩尔百分比的dec2。根据这个摩尔比和100nm直径的脂质体的表面积以及已公布的每106nm2脂质双层5
×
106个脂质分子的估计值
78
,我们计算出在每个dec2
‑
bifc试剂脂质体中大约有1,500个dec2单体(即,750个dec2
‑
vyn和750个dec2
‑
vc融合蛋白)。参见表2。这些脂质体可以通过例如生物素
‑
链霉亲和素结合(图右侧)容易地连接到不溶性基质上。在此构造中,dec2
‑
bifc试剂应使用标准荧光仪器检测释放到血清中的极低浓度的真菌甘露聚糖。b.dectin
‑
2 venus融合蛋白的设计。使两种单独的dectin
‑
2融合蛋白均在大肠杆菌中表达。使一个与突变的venus蛋白vyn的n末端融合,另一个与venus蛋白vc的c末端片段融合。已知这两个充分表征的互补片段在因相互作用的载体蛋白的组装而变得邻近时会组装并产生bifc信号(即,本文中的dec2二聚体)。不同长度的柔性(gly ser)
n
gly间隔区将功能结构域分开以允许运动和功能有一定的独立性。
62.图29显示dec2
‑
bifc试剂产生甘露聚糖特异性venus绿色荧光信号。在与dec2
‑
bifc试剂脂质体一起在23℃下孵育2小时期间,将每种多糖以1mg/ml添加到96孔微量滴定板的一列孔中。从所有其他样本中减去来自与葡聚糖一起孵育的平均背景信号。在预期的目标酵母α
‑
甘露聚糖和真菌β
‑
葡聚糖和蔗糖之间指示出了倍数差异。标准误差用线和须线表示。
63.图30a
‑
j显示dec2
‑
bifc试剂脂质体产生烟曲霉(a.fumigatus)细胞依赖性绿色荧光信号。使4,500个烟曲霉(a.fumigatus)分生孢子(cea10菌株)在赖氨酸包被的24孔微量滴定板上
在vmm 1%葡萄糖中在37℃
下发芽并生长72小时,在福尔马林中固定,并洗涤到ldb2缓冲液中。a
‑
h:将细胞与在脂质体稀释缓冲液ldb2中以1:100稀释的脂质体dec2
‑
bifc试剂一起孵育。最终的dec2蛋白浓度为约1ug/100ul。真菌细胞菌落是以20x放大倍数配对拍摄。示出了成对的明场图像和合并的荧光图像。示出与稀释缓冲液一起孵育的对照细胞和红色通道的i和j图像被简化,以示出来自绿色通道的可忽略不计的荧光背景。左侧的a、c、e、g、i是烟曲霉(a.fumigatus)细胞菌落的明场图像。右侧的b、d、f、h、j是与左侧相邻明场图像的venus绿色荧光蛋白图像合并的明场图像(红色)。30b中的白色箭头表示不产生信号的罕见菌丝。
具体实施方式
64.以下描述列举了本发明的组合物和方法的各个方面和实施方案。特定的实施方案并非旨在限定所述组合物和方法的范围。相反,实施方案仅提供至少包括在所公开的组合物和方法的范围内的非限制性实例。
65.在全球范围内,超过3亿人患有真菌感染。一些真菌疾病是急性且严重的。其他真菌感染是复发性的,一些是慢性的。每年分别有大约200,000、400,000和1,000,000例曲霉病、念珠菌病和隐球菌病,死亡率高得惊人。皮肤真菌感染、皮肤真菌病是最常见的真菌感染。据估计,全球4%至10%的人口(例如,超过3亿人)患有足部真菌感染(足藓感染(athlete’s foot infection)、tina pedia),尤其是脚趾甲感染(甲真菌病),其可能会致
残。
66.烟曲霉(a.fumigatus)和相关的曲霉属物种会引起曲霉病。罹患危及生命的曲霉病风险最大的患者存在例如由干细胞移植或器官移植引起的免疫系统减弱,或者患有各种肺部疾病,包括结核病、慢性阻塞性肺疾病(copd)、囊性纤维化或哮喘。在免疫受损的患者中,曲霉病是继念珠菌病之后的第二大常见真菌感染。与治疗侵袭性曲霉病相关的额外费用估计为每名儿童40,000美元,每名成人10,000美元。曲霉病患者用抗真菌药物,诸如两性霉素b、伊曲康唑、伏立康唑、氟康唑等进行治疗。然而,即使采用抗真菌疗法,免疫受损的曲霉病患者的一年生存率也仅为25%至60%。此外,所有已知的治疗曲霉病的抗真菌剂对人细胞都有很大的毒性(allen等人,antifungal agents for the treatment of systemic fungal infections in children.paediatrics&child health 15:603
‑
608(2010))。本文提供了靶向脂质体,其可改善抗真菌药物递送,并增强抗真菌剂对广泛范围的真菌病原体的治疗功效以及/或者降低抗真菌剂在被施用于受试者时的毒性。
67.纳米颗粒
68.本文提供了用于将药物递送至真菌细胞的纳米颗粒。如全文中所用,纳米颗粒可以是但不限于脂质纳米颗粒,例如,脂质体或非脂质体脂质纳米颗粒(例如,具有非水核的脂质纳米颗粒(lnp))、枝状聚合物、聚合物胶束、纳米胶囊或纳米球,仅举几例。例如,本文提供了纳米颗粒,例如脂质体,其包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子,其中靶向分子被掺入到纳米颗粒的外表面中,而抗真菌剂被包封在纳米颗粒中。用于使纳米颗粒(例如脂质体)靶向至本文所述的真菌的靶向分子可以使各种组合物的负载药物的纳米颗粒靶向至真菌细胞。其他实例包括但不限于氧化铁纳米颗粒、多糖凝胶纳米颗粒和二氧化硅纳米颗粒。
69.如本文所用,术语脂质体是指由至少一个脂质双层包围的水性或水性缓冲隔室。脂质体能够在由至少一个脂质双层包围的隔室(即内部空腔或空间)中携带水溶液、化合物、药物或其他物质。脂质体的尺寸,即直径可以变化。例如,脂质体可以具有约1000纳米(nm)或更小的尺寸。例如,脂质体颗粒可以具有约50nm至约1000nm、约50nm至约900nm、约50nm至约800nm、约50nm至约700nm、约50nm至约600nm、约50nm至约500nm、约50nm至约400nm、约50nm至约300nm、约50nm至约200nm或约50nm至约100nm的尺寸。脂质体包括包含用于包封一种剂(例如抗真菌剂)的隔室的脂质体、包含附着到或被掺入到脂质体外部的靶向分子的脂质体和包含包封的抗真菌剂的脂质体。包封的抗真菌剂是完全或部分位于脂质体内部空间的抗真菌剂。例如,在本文所述的任何脂质体中,至少约75%、80%、85%、90%、95%或99%的抗真菌剂被掺入到脂质体的内部空间中或者被掺入到脂质体的脂质双层中。
70.本文所述的任何纳米颗粒,例如脂质体,相对于脂质可以含有约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20摩尔或更高的百分比的抗真菌剂。换言之,纳米颗粒可以包含1:100、2:100、3:100、4:100、5:100、6:100、7:100、8:100、9:100、10:100、11:100、12:100、13:100、14:100、15:100、16:100、17:100、18:100、19:100、20:100或更高的抗真菌剂与脂质体脂质的摩尔比。如本文所用,摩尔比是两种组分的量(摩尔)之间的比值,例如靶向分子的摩尔数与脂质的摩尔数之间的比值(靶向分子的摩尔数:脂质的摩尔数)或抗真菌剂的摩尔数和脂质的摩尔数之间的比值(抗真菌剂的摩尔数:脂质的摩尔数)。并且类似地,纳米颗粒可以包含0.002:100、0.05:100、0.1:100、0.5:100、1:100、2:100、3:100、4:
100、5:100、10:100、15:100、20:100、25:100或更高的靶向蛋白与脂质体脂质的摩尔比。
71.还提供了本文所述的任何脂质体中的两种或更多种的多重体。例如,脂质体的多重体可以包含约2至约1
×
10
14
(100万亿)个脂质体。例如,多重体可以具有至少100、250、500、750、1000、5000、10,000、25,000、50,000、100,000、500,000、100万或更多的脂质体。脂质体可以通过本领域技术人员已知或后来发现的任何合适的方法制备。一般来说,脂质体可以通过薄膜水合技术以及随后的几个冻融循环来制备。脂质体混悬液还可以根据本领域技术人员已知的方法制备。akbarzadeh等人(“liposome:classification,preparation and applications,”nanoscale res.lett.8(1):102(2013))描述了制备脂质体的示例性方法,该文献通过引用方式整体并入本文。
72.一般而言,可以使用多种脂质成分来制造脂质体。这些脂质包括在生理ph值下以不带电形式或中性两性离子形式存在的中性脂质。此类脂质包括,例如,二酰基磷脂酰胆碱、二酰基磷脂酰乙醇胺、神经酰胺、鞘磷脂、二氢鞘磷脂、脑磷脂和脑苷脂。本文所述的任何脂质的合成衍生物也可以用于制造脂质纳米颗粒。脂质纳米颗粒还可包含甾醇,例如胆固醇。脂质纳米颗粒还可以包含在大约生理ph值下带有净正电荷的阳离子脂质。此类阳离子脂质包括但不限于n,n
‑
二油基
‑
n,n
‑
二甲基氯化铵(dodac);n
‑
(2,3
‑
二油基氧基)丙基
‑
n,n,n
‑
三乙基氯化铵(dotma);n,n
‑
二硬脂酰基
‑
n,n
‑
二甲基溴化铵(ddab);n
‑
(2,3
‑
二油酰氧基)丙基)
‑
n,n,n
‑
三甲基氯化铵(dotap);1,2
‑
二油基氧基
‑3‑
三甲氨基丙烷氯化物盐(dotap
·
cl);3
‑
β
‑
(n
‑
(n',n'
‑
二甲氨基乙烷)
‑
氨甲酰基)胆固醇(“dc
‑
chol”)、n
‑
(1
‑
(2,3
‑
二油基氧基)丙基)
‑
n
‑2‑
(精胺甲酰胺基)乙基)
‑
n,n
‑
二甲基
‑
三氟乙酸铵(“dospa”)、双十八烷基酰胺基甘氨酰基羧基精胺(dogs)、1,2
‑
二油酰基
‑
sn
‑3‑
磷酸乙醇胺(dope)、1,2
‑
二油酰基
‑3‑
二甲基铵丙烷(dodap)、n,n
‑
二甲基
‑
2,3
‑
二油基氧基)丙胺(dodma)和n
‑
(1,2
‑
二肉豆蔻酰氧基丙
‑3‑
基)
‑
n,n
‑
二甲基
‑
n
‑
羟乙基溴化铵(dmrie)。阴离子脂质也适用于本文所述的脂质纳米颗粒。这些脂质包括但不限于磷脂酰甘油、心磷脂、二酰基磷脂酰丝氨酸、二酰基磷脂酸、n
‑
十二酰磷脂酰乙醇胺、n
‑
琥珀酰磷脂酰乙醇胺、n
‑
戊二酰磷脂酰乙醇胺、赖氨酰磷脂酰甘油以及与中性脂质连接的其他阴离子修饰基团。
73.在一些实例中,脂质体包括磷脂酰胆碱、磷脂酰乙醇胺、磷脂酰丝氨酸、磷脂酰肌醇、磷脂酸、磷脂酰甘油、棕榈酰油酰磷脂酰胆碱、溶血磷脂酰胆碱、溶血磷脂酰乙醇胺、二肉豆蔻酰磷脂酰胆碱(dmpc)、二棕榈酰磷脂酰胆碱(dppc)、二油酰磷脂酰胆碱、二硬脂酰磷脂酰胆碱(dspc)、二亚油酰磷脂酰胆碱、1,2
‑
二硬脂酰基
‑
sn
‑
甘油
‑3‑
磷酸乙醇胺(dspe)缀合的聚乙二醇(dspe
‑
peg)、鞘磷脂、胆固醇或其任何组合。在一些实例中,peg可为peg
‑
分子量(mw500)至peg
‑
mw20000。除了本文所述脂质体的组分外,本文所述的任何脂质均可与结合真菌细胞上抗原的靶向分子或其片段缀合。在一些实例中,本文所述的任何脂质的聚乙二醇化型式均可与结合真菌细胞上的抗原(例如,真菌细胞壁抗原或真菌细胞胞外多糖基质抗原)的靶向分子或其片段缀合。
74.如全文中所用,靶向分子是对真菌细胞上的抗原具有结合亲和力,任选地具有特异性结合亲和力的分子,并且可以包括但不限于抗体、多肽、肽、适体或小分子。如全文中所用,真菌细胞上的抗原或靶真菌细胞抗原可以是在真菌细胞周期的任何阶段与真菌细胞缔合的任何抗原。例如但不限于,抗原可以是与真菌细胞缔合的抗原(例如,包埋到真菌细胞壁中的抗原、附着于真菌细胞壁的抗原或真菌细胞表面抗原)。与真菌细胞缔合的抗原也可
以直接或间接地与真菌细胞结合,例如直接或间接地与真菌细胞壁结合。抗原也可以是由真菌细胞产生或与真菌细胞缔合的真菌胞外多糖基质(例如生物膜)的抗原。在一些实例中,胞外多糖基质附着或结合至真菌细胞或真菌细胞群。应理解,与真菌细胞缔合的胞外多糖基质可以但不一定由与它缔合的真菌细胞或真菌细胞群产生。如全文中所用,抗原可以是但不限于蛋白质、脂质或碳水化合物。
75.如全文中所用,“多肽”、“蛋白”以及“肽”在本文中可互换用于指代氨基酸残基的聚合物。如本文所用,该术语涵盖其中氨基酸残基通过共价肽键连接的任何长度的氨基酸链,包括全长蛋白质。
76.如全文中所用,术语“核酸”是指单链或双链形式的脱氧核糖核酸(dna)或核糖核酸(rna)及其聚合物。除非特别限制,否则该术语涵盖含有天然核苷酸的已知类似物的核酸,所述核酸具有与参考核酸相似的结合性质,并且以与天然存在的核苷酸相似的方式代谢。除非另有说明,否则特定的核酸序列还隐含地包括其保守修饰的变体(例如,简并密码子替换)、等位基因、直系同源物、snp和互补序列以及明确指示的序列。具体而言,简并密码子替换可通过生成其中一个或多个选定(或所有)密码子的第三个位置被混合碱基和/或脱氧肌苷残基替换的序列来实现(batzer等人,nucleic acid res.19:5081(1991);ohtsuka等人,j.biol.chem.260:2605
‑
2608(1985);以及rossolini等人,mol.cell.probes 8:91
‑
98(1994))。
77.在列举特异性核酸或多肽序列的每个情况下,还提供了包含与所叙述的序列具有至少70%(例如70%、75%、80%、85%、90%、95%、99%)同一性的序列的实施方案。关于序列的同一性和相似性被定义为在比对序列并在必要时引入空位以实现最大的序列同一性百分比后,候选序列中与起始氨基酸残基同一的氨基酸残基(即,相同的残基)的百分比。例如,本文提供了与seq id no:1
‑
20具有至少70%(例如70%、75%、80%、85%、90%、95%、99%)同一性的多肽和核酸序列。还提供了不包括seq id no:1
‑
20中列出的组氨酸标签和/或接头序列的多肽和核酸序列。本文还提供了与不包括seq id no:1
‑
20中列出的组氨酸标签和/或接头序列的多肽和核酸序列具有至少70%(例如70%、75%、80%、85%、90%、95%、99%)同一性的多肽和核酸序列。还提供了编码本文所述多肽的核酸。用于比较的序列比对方法是本领域众所周知的。用于比较的最佳序列比对可以例如通过smith和waterman的局部同源性算法(adv.appl.math.2:482,1970)、needleman和wunsch的同源性比对算法(j.mol.biol.48:443,1970)、pearson和lipman关于相似性方法的研究(proc.natl.acad.sci.usa85:2444,1988)、这些算法的计算机化实现方式(例如,wisconsin genetics software package,genetics computer group,575science dr.,madison,wis.中的gap、bestfit、fasta和tfasta),或人工比对和目视检查(参见,例如,ausubel等人,current protocols in molecular biology(1995supplement))进行。
78.本文公开的任何多肽可以包含一个或多个保守性氨基酸替换。作为非限制性实例,下面的列表总结了通常可能能在不导致相应变体的生物活性显著改变的情况下进行的可能替换:
79.1)丙氨酸(a)、丝氨酸(s)、苏氨酸(t)、缬氨酸(v)、甘氨酸(g)和脯氨酸(p);
80.2)门冬氨酸(d)、谷氨酸(e);
81.3)门冬酰胺(n)、谷氨酰胺(q);
82.4)精氨酸(r)、赖氨酸(k)、组氨酸(h);
83.5)异亮氨酸(i)、亮氨酸(l)、蛋氨酸(m)、缬氨酸(v)以及
84.6)苯丙氨酸(f)、酪氨酸(y)、色氨酸(w)。
85.还可参见creighton,proteins,w.h.freeman and co.(1984)。
86.在制作此类改变/替换时,还可以考虑氨基酸的亲水指数。亲水性氨基酸指数在赋予蛋白质交互式生物学功能中的重要性在本领域中是普遍了解的(kyte和doolittle;(1982)j mol biol.157(1):105
‑
32)。公认的是,氨基酸的相对亲水特征促成所得蛋白质的二级结构,其反过来限定了蛋白质与其它分子(例如酶、底物、受体、dna、抗体、抗原等)的相互作用。
87.本文提供的任何多肽,例如可溶性dectin
‑
1、dectin
‑
2或dectin
‑
3单体,可以包含n末端或c末端缺失或截短。例如,可以从本文提供的任何多肽的n末端或c末端删除1、2、3、4、5、6、7、8、9、10个或更多个氨基酸,但仍保留至少一个功能,例如,与另一种可溶性dectin单体的二聚化。如附图中所示,可溶性dectin单体多肽序列以纯文本形式示出。如上所述,这些不包括组氨酸标签或接头序列的序列可以通过从多肽的n末端和/或c末端删除1、2、3、4、5、6、7、8、9、10个或更多个氨基酸来截短。
88.还提供了包含本文所述的任何多肽的组合物。例如,本文提供了包含多肽的组合物,所述多肽包含可溶性dectin
‑
1、dectin
‑
2或dectin
‑
3单体多肽或其片段。任选地,所述多肽可以包含以下或者由以下组成:seq id no:2的氨基酸23
‑
199、seq id no:4的氨基酸23
‑
189、seq id no:6的氨基酸23
‑
100、seq id no:8的氨基酸35
‑
214、seq id no:10的氨基酸36
‑
203或seq id no:12的氨基酸35
‑
207。还提供了包含以下或者由以下组成的多肽的片段:seq id no:2的氨基酸23
‑
199、seq id no:4的氨基酸23
‑
189、seq id no:6的氨基酸23
‑
100、seq id no:8的氨基酸35
‑
214、seq id no:10的氨基酸36
‑
203或seq id no:12的氨基酸35
‑
207。例如但非限制性地,还提供了包含以下的多肽的在n末端或c末端上含有约1、2、3、4、5、6、7、8、9、10或更多个氨基酸的缺失的片段:seq id no:2的氨基酸23
‑
199、seq id no:4的氨基酸23
‑
189、seq id no:6的氨基酸23
‑
100、seq id no:8的氨基酸35
‑
214、seq id no:10的氨基酸36
‑
203或seq id no:12的氨基酸35
‑
207。任选地,所述多肽与荧光部分,例如但不限于罗丹明连接或缀合。如实施例部分中所述,所述组合物可以包含缓冲液,例如,含有约0.5m至约1.5m l
‑
精氨酸的复性缓冲液。例如但非限制性地,所述组合物可以包含ph值为7.2的含有以下的缓冲液:介于约0.05和0.15m之间的nah2po4、介于约10mm和20mm之间的三乙醇胺、介于约0.5m和1.5m之间的l
‑
精氨酸、介于约50和200mm之间的nacl、介于约2.5mm至7.5mm之间的edta和介于约0.25至7.5mm之间的bme。任选地,所述组合物可以包含ph值为7.2的含有以下的缓冲液:约0.1m nah2po4、约10mm三乙醇胺、约1m l
‑
精氨酸、约100mm nacl、约5mm edta和5mm bme。还提供了包括任一种所述组合物的试剂盒。任选地,试剂盒包含变性缓冲液或还原缓冲液,例如,包含β
‑
巯基乙醇的还原缓冲液。还提供了包括本文所述的任何脂质体的试剂盒。
89.如全文中所用,术语“抗体”包括但不限于纳米体、任何类别的完整免疫球蛋白(即,完整抗体)(包括多克隆和单克隆抗体)以及保留结合其特异性抗原的能力的抗体片段。还有用的是抗体片段和抗原结合蛋白(单链抗体)的缀合物,如例如美国专利第4,704,692号中所述的,其内容通过引用方式整体并入本文。
90.如全文中所用,适体是选择性地结合靶抗原的寡核苷酸(单链dna或单链rna)或肽分子。参见,例如,lakhin等人,“aptamers:problems,solutions and prospects,”acta naturae 5(4):34
‑
43(2013);以及reverdatto等人,“peptide aptamers:development and applications,”curr.top med.chem.15(12):1082
‑
101(2015)),其通过引用方式整体并入本文。
91.如本文所用,术语“特异性地结合”或“选择性地结合”意指与非特异性或非选择性相互作用可测量地不同的结合。特异性结合可以例如通过与对照分子的结合相比,确定分子与靶抗原的结合进行测量。特异性结合可以通过与类似于靶抗原的对照分子诸如过量的未标记靶抗原竞争来确定。在这种情况下,如果标记靶标与探针的结合受到过量未标记靶抗原竞争性地抑制,则指示特异性结合。
92.任选地,可以将两个靶向分子至约10,000个靶向分子掺入到本文提供的脂质体中。例如,可以将约5个到约100个、约5个到约200个、约5个到约300个、约5个到约400个、约5个到约500个、约5个到约600个、约5个到约700个、约5个到约800个、约5个到约900个、约5个至约1000个、约5个至约1100个、约5个至约1200个、约5个至约1300个、约5个至约1400个、约5个至约1500个、约5个至约1600个、约5个至约1700个、约5个至约1800个、约5个至约1900个、约5个至约2000个、约5个至约2250个、约5个至约2500个、约5个至约3000个靶向分子、约5个至约3500个、约5个至约4000个靶向分子、约5个至约4500个靶向分子、约5个至约5000个靶向分子、约5个至约5500个靶向分子、约5个至约6000个靶向分子、约5个至约6500个靶向分子、约5个至约7000个靶向分子、约5个至约7500个靶向分子、约5个至约8000个靶向分子、约5个至约8500个靶向分子、约5个至约9000个靶向分子、约5个至约9500个靶向分子或约5个至约10,000个靶向分子掺入到一个或多个本文所述的脂质体中。在一些实例中,将约2个分子至约3,000个靶向分子掺入到直径100nm的纳米颗粒中。本领域技术人员不知道如何计算可以掺入到纳米颗粒中的靶向分子的数量,例如介于约2个至10,000个靶向分子之间或更多个靶向分子,这取决于纳米颗粒的尺寸。
93.如全文中所用,将靶向分子掺入到脂质体的外表面中意味着靶向分子被掺入到脂质体的外脂质双层中或附着于脂质体上。掺入可以通过将靶向分子插入或嵌入到脂质双层中而发生。与脂质体的附着可以例如通过与被掺入到脂质体的外脂质双层中的分子的亲和性而发生。例如,脂质体可以包被有生物素(例如,被插入到脂质双层中的dspe
‑
peg
‑
生物素)以及与链霉亲和素连接的靶向分子。替代地,靶向分子可以缀合到脂质体的外表面。靶向分子可以通过本领域已知的许多方法缀合至脂质体(例如,arruebo等人,“antibody
‑
conjugated nanoparticles for biomedical applications,”journal of nanomaterials vol.2009,论文id 439389(2009))。脂质体还可以通过链霉亲和素/生物素键、硫醇/马来酰亚胺化学、叠氮化物/炔化学、四嗪/环辛炔化学和其他点击化学缀合至靶向分子。这些化学操控物(handles)是在亚磷酰胺合成过程中或合成后制备的。如本文所用,术语“点击化学”是指主要旨在将所选底物与特异性生物分子结合的生物相容性反应。点击化学反应不受水的干扰,产生极少且无毒的副产物,并且特征在于高热力学驱动力,该高热力学驱动力可快速且不可逆地驱动它在高反应特异性的情况下获得单一反应产物的高产率。
94.靶向分子可以结合一种或多种类型的真菌细胞或真菌细胞群上的抗原,所述真菌
细胞包括但不限于来自动物真菌病原体(例如,人真菌病原体)和植物病原体的细胞。人真菌病原体的实例包括但不限于链格孢菌(alternaria alternata)、曲霉菌属物种(诸如烟曲霉(a.fumigatus))、芽生菌属物种(诸如皮炎芽生菌(b.dermatitidis))、念珠菌属物种(诸如白色念珠菌(a.fumigatus)、光滑念珠菌(c.glabrata)、克柔氏念珠菌(c.krusei)、耳念珠菌(c.auris))、球孢子菌属物种(诸如粗球孢子菌(c.immitis)和波萨球孢子菌(c.posadasii))、隐球菌属物种(诸如格特隐球菌(c.gattii)和新型隐球菌(c.neoformans))、组织胞浆菌属物种(诸如荚膜组织胞浆菌(h.capsulatum);肺囊虫属物种(诸如耶氏肺囊虫(p.jirovecii)、孢子丝菌属物种(诸如申克孢子丝菌(s.schenckii)、马尔尼菲蓝状菌(talaromyces marneffei)(以前称为马尔尼菲青霉菌(penicillium marneffei))和红色毛癣菌(trichophyton rubrum)。
95.植物真菌病原体的实例包括但不限于大豆锈病菌(aecidium glycines)、桑树锈病菌(aecidium mori)、日本链格孢菌(alternaria japonica)、稻毛链格孢菌(alternaria padwickii)、小麦链格孢菌(alternaria triticina)、鸭梨链格孢菌(alternaria yaliinficiens)、龟纹淀粉韧革菌(amylostereum areolatum)、apiognomonia erythrostoma、arkoola nigra、水稻香亭病菌(balansia oryzae
‑
sativae)、轮纹病菌(botryosphaeria berengeriana)、丽赤壳菌(calonectria pseudonaviculata
‑
黄杨木枯梢病菌
‑
丽赤壳菌、栎枯萎病菌(ceratocystis fagacearum)、白蜡枯梢病菌(chalara fraxinea)、冷杉金锈菌(chrysomyxa abietis)、喜马金锈菌(chrysomyxa himalensis)、杜鹃金锈菌(chrysomyxa rhododendri)、小菌核病菌(ciborinia allii)、梭状麦角菌(claviceps fusiformis)、大叶麦角菌(claviceps gigantea)、玉蜀黍麦角菌(claviceps sorghi)、高粱麦角菌(claviceps sorghicola)、松芍柱锈菌(cronartium flaccidum)、成堆暗被盘菌(crumenulopsis sororia)、茄间座壳菌(diaporthe vexans)、小双胞腔菌
‑
蚕豆叶壳二胞菌(didymella fabae
‑
ascochyta fabae)、肯氏内锈菌(endophyllum kaernbachii)、坏损外担菌(exobasidium vexans)、gerwasia imperialis、gerwasia mayorii、gerwasia rosae、gerwasia rubi、gerwasia variabilis、goplana dioscoreae、柔果胶锈菌(gymnosporangium miyabei)、山田胶锈菌(gymnosporangium yamadae)、尖戟孢锈菌(hamaspora acutissima)、澳洲戟孢锈菌(hamaspora australis)、桥冈戟孢锈菌(hamaspora hashiokai)、长戟孢锈菌(hamaspora longissimi)、鲁比戟孢锈菌(hamaspora rubi
‑
sieboldii)、中国戟孢锈菌(hamaspora sinica)、玉米枯萎头孢霉(harpophora maydis)、中国咖啡锈菌(hemileia vastatrix)、日本不多胞锈菌(kuehneola japonica)、kuehneola loeseneriana、落叶松癌肿病菌(lachnellula willkommii)、翼状半帚孢(leptographium wingfieldii)、mainsia rubi、黄花柳栅锈菌(melampsora capraearum)、松柳栅锈菌(melampsora larici
‑
epitea)、落叶松五蕊柳栅锈菌(melampsora larici
‑
pentandrae)、落叶松五蕊柳栅锈菌(melampsora larici
‑
pentandrae)、串珠霉(monilia polystroma)、果生链核盘菌(monilinia fructigena)、美赭痂锈菌(ochropsora ariae)、柚木周丝单胞锈菌(olivea tectonae)、ophiostoma longicollum、peronospora digitalis、菊花霜霉(peronospora radii)、葡萄层锈菌(phakopsora ampelopsidis)、真葡萄层锈菌(phakopsora euvitis)、山马蟥层锈菌(phakopsora meibomiae)、由豆薯层锈菌(phakopsora pachyrhizi)、维管束茎点霉(phoma tracheiphila)、尖形多孢锈菌
(phragmidium acuminatum)、北极多孢锈菌(phragmidium arcticum)、阿里山多孢锈菌(phragmidium arisanense)、phragmidium assamense、巴克莱多孢锈菌(phragmidium barclayi)、球茎多孢锈菌(phragmidium bulbosum)、巴氏多孢锈菌(phragmidium butleri)、台湾多孢锈菌(phragmidium formosanum)、灰斑多孢锈菌(phragmidium griseum)、平冢多孢锈菌(phragmidium hiratsukanum)、堪察加多胞锈菌(phragmidium kamtschatkae)、纳布多孢锈菌(phragmidium nambuanum)、少隔多孢锈菌(phragmidium pauciloculare)、phragmidium rosae
‑
moschatae、蔷薇多胞锈菌(phragmidium rosae
‑
multiflorae)、玫瑰多孢锈菌(phragmidium rosae
‑
rugosae)、蓬垒多胞锈菌(phragmidium rubi
‑
thunbergii)、山田多胞锈菌(phragmidium yamadanum)、玉米黑痣菌(phyllachora maydis)、黄莲木帽孢锈菌(pileolaria pistaciae)、笃耨香帽孢锈菌(pileolaria terebinthi)、凤仙花霜霉(plasmopara obducens)、柑橘斑点病菌(pseudocercospora angolensis)、puccinia agrophila、黄杨柄锈菌(puccinia buxi)、红柄锈菌(puccinia erythropus)、唐菖蒲柄锈菌(puccinia gladioli)、甘油柄锈菌(puccinia glyceriae)、萱草柄锈菌(puccinia hemerocallidis)、水平柄锈菌(puccinia horiana)、屈恩柄锈菌(puccinia kuehnii)、puccinia mccleanii、黑顶柄锈菌(puccinia melanocephala)、芒柄锈菌(puccinia miscanthi)、马铃薯柄锈菌(puccinia pittieriana)、番石榴柄锈菌(puccinia psidii)、寡纹柄锈菌(puccinia substriata)、长叶柄锈菌(puccinia veronicae
‑
longifoliae)、榛膨痂锈菌(pucciniastrum coryli)、霍姆斯刚毛瘤(setomelanomma holmii)、一品红痂圆孢(sphaceloma poinsettiae)、sporisporium pulverulentum、sporisorium sacchari、马铃薯黑粉病菌(thecaphora solani)、杉李盖痂锈菌(thekopsora areolate)、冰草条黑粉菌(urocystis agropyri)、唐菖蒲单胞锈菌(uromyces gladioli)、uromyces nyikensis、唐菖蒲横点锈病菌(uromyces transversalis)和uromycladium tepperianum。应当理解,除了检测、治疗或预防受试者的真菌感染外,本文提供的任何方法均还可用于检测、治疗或预防植物的真菌感染。例如但非限制性地,可以在植物的叶、茎、根、花瓣、萼片、雄蕊、心皮和种子的表面或粉碎样本或提取物上检测、治疗或预防真菌感染。
96.在一些实例中,靶向分子是c型凝集素受体或其片段。在其他实例中,靶向分子是凝集素结合蛋白或其片段。本文所述的靶向分子的片段是这样的多肽,其可与真菌细胞上的靶抗原以与所述片段来源自的蛋白或多肽相同或不同的结合亲和力结合。可以用作靶向分子的c型凝集素受体的实例包括dectin
‑
1(clec7a,小鼠genbank登记号:aas37670和人genbank登记号:np_922938)、dectin
‑
2(clec6a小鼠genbank登记号:np_064385和人genbank登记号:q6eig7)、dectin
‑
3(clec4d小鼠genbank登记号:np_034949和人genbank登记号:np_034949)和dectin
‑
1、dectin
‑
2、dectin
‑
3的片段。包含与真菌细胞上的β
‑
葡聚糖和/或甘露聚糖结合的氨基酸序列的dectin
‑
1、dectin
‑
2、dectin
‑
3的片段也可以用作靶向分子。示例性靶向分子包括但不限于seq id no:2,其包含小鼠dectin
‑
1β
‑
葡聚糖结合片段的氨基酸序列。seq id no:2包含可以被去除的n末端his标签。用于制备可溶性dectin
‑
1(例如,seq id no:7
‑
14)的任何核酸构建体可以进一步包含选择性蛋白酶切割位点以在dectin
‑
1多肽表达后去除n末端his标签序列。这些包括但不限于由烟草蚀刻病毒(tev)蛋白酶、肠肽酶、凝血酶、因子xa和鼻病毒3c蛋白酶识别的肽蛋白酶加工位点。另一个示例性
靶向分子是包含seq id no:6的可溶性dectin
‑
3多肽。seq id no:6包含小鼠dectin
‑
3的氨基酸44至219。另外的示例性靶向分子包括可溶性人dectin
‑
1、dectin
‑
2和dectin
‑
3多肽,其分别包含seq id no:8、10和12。seq id no:8、10和12分别包含人dectin
‑
1的氨基酸69至248、人dectin
‑
2的氨基酸42至209和人dectin
‑
3的氨基酸44至215。还提供了人dectin
‑
1的氨基酸69至248、人dectin
‑
2的氨基酸42至209和人dectin
‑
3的氨基酸44至215的片段,例如,包含人dectin
‑
1的氨基酸69至248、人dectin
‑
2的氨基酸42至209和人dectin
‑
3的氨基酸44至215,或者由其组成的多肽的在c末端和/或n末端含有1、2、3、4、5、6、7、8、9、10或更多个氨基酸的缺失的片段,这些片段可以用于任何所述的脂质体、多肽或组合物中。
97.dectin
‑
1是在t细胞中表达的跨膜受体,并且由小鼠和人中的clec7a(c型凝集素结构域包含7a,β
‑
葡聚糖受体,gr)基因编码。dectin
‑
1与真菌细胞壁中的各种β
‑
葡聚糖结合,并且是跨膜发出致病和非致病真菌表面上存在裸露细胞壁组分的信号以刺激先天免疫应答的主要受体。虽然人和小鼠dectin
‑
1分别是长度为244和247个氨基酸的质膜蛋白,但存在产生较短人类亚型的mrna剪接变体。dectin
‑
1作为单体漂浮在膜上,但作为二聚体与β
‑
葡聚糖结合,如图3中所示的dectin
‑
1靶向脂质体的设计中所模仿的。长度为176个氨基酸(20kda)的细胞外c末端结构域可以作为可溶性sdectin
‑
1单独操作,并含有β葡聚糖结合结构域。β1
→
3葡聚糖是结构上不同的类别的多糖,因此,sdectin
‑
1以不同的方式结合各种β
‑
葡聚糖,kd亲和常数介于2.6mm至2.2pm之间。具有泛真菌结合活性的dectin
‑
1及其片段,例如,β
‑
葡聚糖结合片段是示例性靶向分子,其可用于杀灭一种或多种类型的真菌细胞,例如但不限于,曲霉属、念珠菌属和/或隐球菌属细胞。
98.可以用于脂质体的真菌多糖靶向的其他具有甘露聚糖和甘露糖结合结构域的哺乳动物蛋白质为甘露糖受体c型i(人mrc1,ng_047011)、甘露糖结合蛋白2(mbl2,ng_033955)和c型凝集素结构域家族4成员l(cd209、clec4l或dc
‑
sign,人ng_012167)或其片段。可用作靶向分子的具有真菌碳水化合物结合结构域的非哺乳动物蛋白的实例包括但不限于细菌和昆虫葡聚糖结合蛋白cbm11(例如,来自甲醇类芽孢杆菌(paenibacillus methanolicus)的typ77495)和cbm39(例如,来自毕氏卵角蚁(ooceraea biori)的eza53410)、细菌甘露聚糖结合蛋白cbm27(例如,来自纤维素类芽孢杆菌(paenibacillus cellulosilyticus)的pwv98652)、cbm35(例如,来自类芽孢杆菌属(paenibacillus sp.)ov191的pye65467)、cbm46(例如,来自类芽孢杆菌(paenibacillus sp.)598k的gbf77546)和mvl(例如,来自伯氏金黄杆菌(chryseobacterium bernardetii)的azb35797)或其片段。
99.可用作靶向分子的具有几丁质结合结构域的哺乳动物蛋白质的实例包括但不限于几丁质酶
‑
1(chit1,人ng_012867)、几丁质酶
‑
3样
‑
1(hcgp39、chi3l1、人ng_013056)、几丁质酶
‑
3样
‑
2(chi3l2,人nm_001025197)、几丁质酶酸性(chia,人类基因卡编号gc01p111291)和壳二糖酶(ctbs,人类基因卡gc01m084549)或其片段。可用作靶向分子的具有几丁质结合结构域的非哺乳动物蛋白的实例包括但不限于细菌cbm5(例如,来自木聚糖赖氨酸杆菌(lysinibacillus xylanilyticus)的tdx99194)和植物和/或真菌cbm18(例如,来自苜蓿轮枝菌(verticillium alfalfae)的ayu56549)和cbm19(例如,来自布拉克须霉(phycomyces blakesleeanus)nrr1555的xp_018290979)。
100.可以掺入或包封在本文所述的靶向脂质体中的抗真菌剂包括但不限于多烯或唑类抗真菌剂。多烯抗真菌剂的实例包括但不限于两性霉素b(amb)、念珠菌素、菲律宾菌素、
哈霉素、那他霉素、制霉菌素、日立霉素(hitachimycin)和龟裂杀菌素(rimocidin)。唑类抗真菌剂的实例包括但不限于咪唑类(例如,联苯苄唑、布托康唑、克霉唑、益康唑、芬替康唑、异康唑、酮康唑、卢立康唑、咪康唑、奥莫康唑、奥昔康唑、舍他康唑、舒康唑和噻康唑)、三唑类(例如,阿巴康唑、艾氟康唑、环氧康那唑、氟康唑、艾沙康唑、伊曲康唑、泊沙康唑、丙环唑、雷夫康唑、特康唑、伏立康唑)、噻唑类(例如,阿巴芬净)和棘球白素(例如,卡泊芬净、米卡芬净和阿尼芬净)。
101.两性霉素b(amb)是用于包括曲霉病在内的许多种类的真菌感染的最常用剂。两性霉素b的副作用包括神经毒性和/或肾毒性和/或肝毒性,并且经常导致患者死亡。amb是疏水的,并被嵌入到脂质体的脂质双层中,如图3所示。商业非靶向球形负载amb的脂质体amb
‑
ll通常被称为ambisomes。与第二最常用的amb产品脱氧胆酸盐洗涤剂增溶的amb相比,amb
‑
ll更有效地穿透到各种器官,穿透细胞壁并在略微较高、更有效的amb剂量下显示降低的毒性。然而,amb
‑
ll仍然会产生amb人毒性,诸如在50%的患者中产生肾毒性。当用amb
‑
ll治疗受感染的小鼠时,大的真菌细胞群通常会得以保留。这种大的残留群体可能是用洗涤剂增溶的amb和amb
‑
ll治疗的人患者在治疗后具有高复发率和后续死亡率的原因。本文提供的靶向脂质体被设计用于有效地靶向真菌细胞以及/或者降低抗真菌剂例如amb的毒性。
102.在一些实例中,所述抗真菌药物的浓度与掺入到或包封在这样的脂质体(其未包含掺入到其外表面中的靶向分子或其片段)中的抗真菌药物的浓度相比降低,其中所述靶向分子结合真菌细胞上的抗原。通过形成蛋白质包被的脂质体(即,用多肽,例如c
‑
凝集素型受体或其片段包被脂质体),可以使用较低浓度的抗真菌剂来治疗或预防真菌感染,从而降低与向受试者施用抗真菌药物相关的毒性。较低的毒性允许在更长时间内延长使用靶向抗真菌剂,这将使真菌负荷减少至超出目前实现的较差的减少。较低的毒性可允许预防性地使用本文所述的靶向脂质体,例如,作为鼻喷雾剂,以在确立肺部感染之前对它们进行预防。
103.在一些脂质体中,抗真菌药物的浓度降低或下降至少约5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%或这些百分比之间的任何百分比。在一些实例中,毒性的降低是体外和/或体内肾细胞毒性和/或肝细胞毒性的降低。在另一个实例中,脂质体中amb浓度的降低是从相对于脂质体脂质的约11摩尔百分比降低至相对于脂质体脂质的约1至10摩尔百分比。在一些靶向脂质体中,抗真菌剂的浓度相对于脂质体脂质百分比,可以在约1至约20摩尔百分比的抗真菌剂的范围内。例如,抗真菌剂的浓度相对于脂质体脂质百分比可以是约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20摩尔百分比的抗真菌剂。
104.在一些实例中,所述靶向脂质体与这样的脂质体(其未包含结合真菌细胞上的抗原的靶向分子,例如,掺入到脂质体外表面中的靶向分子)相比对动物细胞(例如人细胞)具有降低的亲和力和/或较小的毒性。在一些实例中,所述靶向脂质体对受试者肺中的真菌细胞的亲和力与所述靶向脂质体对受试者的肺、肾或肝细胞的亲和力相比较高。因此,本文提供的脂质体可以用于向受试者递送抗真菌剂,同时最大程度地减低所述抗真菌剂对非真菌细胞(例如人肺、肾或肝细胞)的影响,从而降低抗真菌剂的毒性。包含本文所述的抗真菌剂的任何脂质体都可以用于体外、离体或体内降低或减少真菌感染。
105.制备靶向脂质体的方法
106.本文提供了制备包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子的脂质体的多重体的方法,其中靶向分子被掺入到每个脂质体的外表面中,而抗真菌剂被包封在每个脂质体中,所述方法包括以下步骤:(a)在约60℃下将抗真菌剂在溶剂中溶解约10分钟至约30分钟;(b)通过将呈悬浮液形式的脂质体的多重体与步骤(a)的抗真菌剂/溶剂溶液,在约60℃下混合约3至约5小时,或在约37℃下混合约24
‑
120小时来将抗真菌剂包封在每个脂质体中;以及(c)通过使包含经包封的抗真菌剂的脂质体与缀合至脂质的靶向分子在60℃下接触约45分钟至约90分钟来将靶向分子掺入到每个脂质体的外表面中。
107.在制备本文提供的脂质体的方法中,抗真菌剂可以溶解在任何合适的溶剂中。根据抗真菌剂及其性质,本领域技术人员会知道如何选择有效的溶解溶剂。在一些实例中,将抗真菌剂例如两性霉素b溶解在含水dmso或甲酰胺中。也可以使用其他疏水性溶剂或双疏溶剂。任选地,可以将抗真菌剂溶解约10至约120分钟。例如,可以将抗真菌剂溶解约10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119或120分钟。
108.任选地,可以将抗真菌剂在约55℃至约65℃的温度下,例如,在约55、56、57、58、59、60、61、62、63、64或65℃下溶解。任选地,可以通过使呈悬浮液形式的脂质体的多重体与抗真菌剂/溶剂溶液(溶解的抗真菌剂)在约55℃至约65℃的温度下,例如,在约55、56、57、58、59、60、61、62、63、64或65℃下混合约3至约5小时来将抗真菌剂包封到每个脂质体中。在另一个实例中,通过使呈悬浮液形式的脂质体的多重体与抗真菌剂/溶剂溶液(溶解的抗真菌剂)在约35℃至约45℃下,例如,在35、36、37、38、39、40、41、42、43、44或45℃下混合约24
‑
120小时来将抗真菌剂包封到每个脂质体中。在另一个实例中,通过使呈悬浮液形式的脂质体的多重体与抗真菌剂/溶剂溶液(溶解的抗真菌剂)在约35℃至约45℃下,例如,在35、36、37、38、39、40、41、42、43、44或45℃下混合约72
‑
100小时来将抗真菌剂是包封到每个脂质体中。
109.任选地,在制备本文所述的脂质体的任何方法中,将选自由dectin
‑
1、dectin
‑
2和dectin 3或其结合片段组成的组的c型凝集素受体或其片段维持在包含精氨酸的复性缓冲液中,并在掺入到脂质体中之前进行变性。任选地,制备本文所述的脂质体的任何方法可以进一步包括将包含抗真菌剂和靶向分子的脂质体储存在包含精氨酸的复性缓冲液中。任选地,复性缓冲液可以包含约0.5至约1.5m的精氨酸。任选地,复性缓冲液可以包含约0.1m nah2po4、约10mm三乙醇胺、约1m l
‑
精氨酸、约100mm nacl、约5mm edta和5mm bme,ph 7.2。
110.在一些方法中,所述靶向分子缀合至脂质。缀合至所述靶向分子的脂质可以是聚乙二醇化或非聚乙二醇化脂质。可缀合至靶向分子的脂质的实例以及可掺入到靶向脂质体中的抗真菌剂的实例在上文中有阐述。在一些实例中,掺入到每个脂质体的外表面中的靶向分子是c型凝集素受体,例如,dectin
‑
1、dectin
‑
2、dectin
‑
3或其片段,几丁质结合蛋白,胞外多糖结合蛋白或其片段,或抗体。
111.通过所描述的方法制备的靶向脂质体的多重体可以产生包含任何数量的脂质体,例如,约两个至约100,000,000个脂质体的多重体。应当理解,在制备靶向脂质体的过程中,
可能存在一些未包封抗真菌剂的脂质体或者不具有被掺入到其外表面中的靶向分子的脂质体。因此,本文提供了靶向脂质体的多重体,其中至少70%、80%、90%、95%、99%、99.5%或99.9%的脂质体含有经包封的抗真菌剂和结合真菌细胞上的抗原的靶向分子,其中所述靶向分子被掺入到脂质体的外表面中。
112.真菌感染检测
113.本文还提供了包含结合靶真菌细胞抗原的靶向分子和信号产生分子的脂质体,其中靶向分子被掺入到脂质体的外表面中,并且信号产生分子在靶向分子结合靶真菌细胞抗原时产生可检测的信号。在一些实例中,信号产生分子连接或附着于靶向分子。在其他实例中,信号产生分子被掺入到脂质体的外表面中或附着于脂质体的外表面上。这些脂质体可以用于体内、离体或体外检测真菌或真菌感染的存在。可以检测真菌细胞或一种或多种真菌细胞,即真菌细胞群体。可以直接或间接检测可检测的信号。例如,信号产生分子可以是直接检测的荧光染料、标记或探针(例如,罗丹明、荧光素、绿色荧光蛋白、吖啶橙等)。
114.在一些实例中,靶向分子连接至可使用成像技术(包括但不限于磁共振成像、放射线照相术、正电子发射断层扫描(pet)、计算机体层摄影(ct)扫描,仅具几个例子)直接体内检测的分子。可以用于使用体内成像检测真菌感染的分子的实例包括但不限于金属蛋白、铁蛋白、转铁蛋白、水通道蛋白和化学交换饱和转移(cest)报告物,仅举几个例子。参见,例如,silindir等人,“liposomes and their applications in molecular imaging,”j.drug target 20(5):401
‑
415(2012);以及mukherjee等人,“biomolecular mri reporters:evolution of new mechanisms,”prog.nucl.magn reson.spectrosc.102
‑
103:32
‑
42(2017))。在另一个实例中,靶向分子连接至可以使用二抗间接检测的一抗或其片段(例如,抗体的fc片段)。在一些实例中,靶真菌抗原位于真菌细胞上。在其他实例中,靶真菌抗原例如真菌细胞壁成分是从真菌细胞释放到生物样本中。
115.在一些实例中,脂质体本身在与真菌细胞抗原结合时产生信号。此信号可以在脂质体结合后的一个或多个步骤中产生。单步信号发出系统的实例如图11和28a中所示,其中脂质体可能会或可能不会任选地附着于固体支持物上。在此实施例中,使受试者或生物样本与包含融合蛋白(其包含连接至荧光蛋白的c末端片段的可溶性dectin(靶向分子))的脂质体和包含连接至荧光蛋白的n末端片段的可溶性dectin(靶向分子)的脂质体接触。荧光蛋白的实例包括但不限于黄色荧光蛋白(yfp,例如venus)、绿色荧光蛋白(gfp)和红色荧光蛋白(rfp)以及这些蛋白质的衍生物,例如突变衍生物。参见,例如,chudakov等人,“fluorescent proteins and their applications in imaging living cells and tissues,”physiological reviews 90(3):1103
‑
1163(2010);以及specht等人,“a critical and comparative review of fluorescent tools for live
‑
cell imaging,”annual review of physiology79:93
‑
117(2017))。
116.在一些实例中,靶向分子例如可溶性dectin
‑
1、dectin
‑
2、dectin
‑
3或其片段在结合真菌细胞上的β
‑
葡聚糖或甘露聚糖后在脂质体的表面上形成二聚体。当其表面上存在连接至荧光蛋白的n末端片段的第一靶向分子和连接至荧光蛋白的c末端片段的第二靶向分子的脂质体与受真菌感染的受试者(例如,受试者的眼、耳、喉、阴道、鼻道、皮肤或指甲,仅举几个例子)或受试者的样本(例如,尿、血液、血清、泪液、痰液、肺灌洗液、组织刮片或匀浆)接触时,掺入到脂质体表面中的可溶性dectin单体在与真菌细胞上存在的或来源于真
pharmacology of neuropeptide y y1/y5 receptor heterodimers,”mol.pharm.87:718
‑
732(2015))。将在与真菌葡聚糖或甘露聚糖或真菌本身结合时产生荧光信号的dectin包被脂质体的示意性模型在图11和图28a中示出。这种构建体可实现单步测定,其中脂质体与真菌多糖的结合和随后的信号产生都发生在同一个单一临床步骤中,无需洗涤步骤或添加额外的试剂。参见,例如,kilpatrick等人,“a g protein
‑
coupled receptor dimer imaging assay reveals selectively modified pharmacology of neuropeptide y y1/y5 receptor heterodimers,”mol.pharm.87:718
‑
732(2015))。
121.本文进一步提供了融合多肽,所述融合多肽包含结合靶真菌细胞抗原的靶向分子和荧光多肽的n末端或c末端部分。这些融合多肽可以用于检测真菌感染。在一些实例中,所述靶向分子是c型凝集素受体、抗体、真菌细胞壁结合蛋白、几丁质结合蛋白或其片段。在一些实例中,所述靶向分子是dectin
‑
1、dectin
‑
2、dectin
‑
3或其片段。在一些实例中,所述荧光蛋白是黄色荧光蛋白或其衍生物。
122.本文提供了用于检测受试者中或受试者的样本中真菌感染的方法,其包括:a)使受试者或受试者的样本与脂质体的多重体接触,其中所述多重体中的每个脂质体均包含结合靶真菌细胞抗原的靶向分子和信号产生分子,其中靶向分子被掺入到脂质体的外表面中,并且信号产生分子在靶向分子结合靶真菌细胞抗原时生成可检测的信号;以及b)检测信号,其中信号指示真菌感染的存在。这种方法可以用于体内、离体或体外检测真菌感染的存在。在一些实例中,真菌细胞抗原是细胞上的真菌细胞抗原。在其他实例中,真菌细胞抗原是可溶性真菌细胞抗原,例如生物样本中的β
‑
葡聚糖或甘露聚糖。在一些实例中,靶向分子连接至信号产生分子。在其他实例中,信号产生分子被掺入到脂质体的外表面中或附着于脂质体的外表面上。在一些实例中,靶向分子连接至信号产生酶,例如hrp、萤光素酶、β
‑
葡萄糖醛酸酶和β
‑
半乳糖苷酶。在其他实例中,靶向分子连接至荧光蛋白,例如罗丹明、gfp、yfp、rfp等。荧光蛋白的片段,例如,任何荧光蛋白的n末端或c末端,可以连接至靶向分子。在其他实例中,靶向分子连接至抗体或其片段。
123.本文提供了用于检测受试者或受试者的样本中真菌感染的方法,其包括:(a)使受试者或受试者的样本与脂质体的多重体接触,其中所述多重体中的每个脂质体均包含连接至荧光蛋白的n末端片段的靶向分子和连接至荧光蛋白的c末端片段的靶向分子,其中靶向分子结合真菌细胞上的靶抗原,以及(b)检测由荧光蛋白的n末端和c末端片段之间的相互作用产生的荧光信号,其中信号指示真菌感染的存在。在一些方法中,使用紫外光或荧光显微镜或用于双分子荧光互补的定量荧光检测(即,bifc分析)的仪器来检测荧光信号。
124.在一些用于检测真菌感染的方法中,所述多重体中的每个脂质体均包含:a)至少约500个连接至荧光蛋白n末端片段的靶向分子;b)至少约500个连接至荧光蛋白c末端片段的靶向分子。例如,每个脂质体可以包含至少约250、500、100、1500、2000或2500个连接至荧光蛋白的n末端片段的靶向分子和至少约250、500、100、1500、2000或2500个连接至荧光蛋白的c末端片段的靶向分子。
125.进一步提供了用于检测受试者或受试者的样本中的真菌感染的方法,其包括:a)使受试者或受试者的样本与包含结合靶真菌细胞抗原的靶向分子和荧光多肽的n末端部分的第一融合多肽多重体和包含结合靶真菌细胞抗原的靶向分子和荧光多肽的c末端部分的第二融合多肽多重体接触;以及b)检测由荧光蛋白的n末端和c末端片段之间的相互作用产
生的荧光信号,其中信号指示真菌感染的存在。
126.在一些实例中,所述脂质体的多重体在固体支持物上被固定化。固体支持材料的非限制性实例包括玻璃、改性或功能化玻璃、塑料(包括丙烯酸酯、聚苯乙烯和苯乙烯与其他材料的共聚物、聚丙烯、聚乙烯、聚丁烯、聚氨酯或teflonj、尼龙、硝基纤维素、多糖、树脂)、二氧化硅或基于二氧化硅的材料(包括硅和改性硅)、碳、金属、无机玻璃和塑料。固体支持物的大小和形状可以变化。固体支持物可以是平面的,固体支持物可以是孔,或者替代地,固体支持物可以是珠粒或载片。在一些实例中,固体支持物是多孔板的孔。在其他实例中,固体支持物可以是磁珠粒、基于琼脂糖的树脂或琼脂糖珠粒。在其他实例中,固体支持物包含非琼脂糖色谱介质、整料(monoliths)或纳米颗粒。例如,色谱介质可以是例如甲基丙烯酸酯、纤维素或玻璃。在其他实例中,纳米颗粒是金纳米颗粒或磁性纳米颗粒。
127.如全文中所用,受试者意指个体。受试者可以是成人受试者或儿科受试者。儿科受试者包括年龄范围从出生到十八岁的受试者。因此,年龄小于约10岁、5岁、2岁、1岁、6个月、3个月、1个月、1周或1天的儿科受试者也包括在受试者内。优选地,受试者是哺乳动物,诸如灵长类动物,并且更优选是人。非人类灵长类动物也是受试者。术语“受试者”包括驯养动物(诸如猫、狗等)、家畜(例如牛、马、猪、绵羊、山羊等)和实验室动物(例如,雪貂、毛丝鼠、小鼠、兔、大鼠、沙鼠、豚鼠等)。因此,本文考虑了兽医用途和医疗制剂。
128.如本文所用,生物样本是源自受试者的样本,并且包括但不限于任何细胞、组织或生物流体。样本可以是但不限于血液、血浆、血清、痰液、尿液、唾液、支气管肺泡灌洗液、活检物(例如从器官组织分离的组织或细胞,例如,从肺、肝肾、皮肤等分离的组织或细胞)、阴道分泌物、鼻腔分泌物、皮肤、胃分泌物或骨髓标本。
129.受试者也可以是植物或来自植物的种子。生物样本也可以来自植物,但不限于任何细胞、组织或植物渗出液。样本可以是但不限于叶、茎、根、花瓣、萼片、雄蕊、心皮和种子的表面或它们的粉碎样本或提取物。
130.治疗或预防真菌感染的方法
131.还提供了用于治疗或预防受试者的真菌感染的方法。所述方法包括对患有真菌感染或有罹患真菌感染风险的受试者施用有效量的本文所述的任何脂质体的多重体,其中所述多重体中的每个脂质体均包含抗真菌剂和结合真菌细胞上的靶抗原的靶向分子,其中靶向分子被掺入到脂质体的外表面中,而抗真菌剂被包封在脂质体中。本文提供的任何脂质体或脂质体的多重体可以呈药物组合物的形式。
132.所述方法可以用于治疗或预防任何动物(例如人)的真菌感染。人真菌感染的实例包括但不限于链格孢菌(alternaria alternata)、曲霉菌属物种(诸如烟曲霉(a.fumigatus))、芽生菌属物种(诸如皮炎芽生菌(b.dermatitidis))、念珠菌属物种(诸如白色念珠菌(a.fumigatus)、光滑念珠菌(c.glabrata)、克柔氏念珠菌(c.krusei)、耳念珠菌(c.auris))、球孢子菌属物种(诸如粗球孢子菌(c.immitis)和波萨球孢子菌(c.posadasii))、隐球菌属物种(诸如格特隐球菌(c.gattii)和新型隐球菌(c.neoformans))、组织胞浆菌属物种(诸如荚膜组织胞浆菌(h.capsulatum);肺囊虫属物种(诸如耶氏肺囊虫(p.jirovecii)、孢子丝菌属物种(诸如申克孢子丝菌(s.schenckii)、马尔尼菲蓝状菌(talaromyces marneffei)(以前称为马尔尼菲青霉菌(penicillium marneffei))和红色毛癣菌(trichophyton rubrum)。
133.在全文中,治疗(treat)、治疗(treating)和治疗(treatment)是指减少或延迟真菌感染的一种或多种影响或症状的方法。受试者可以被诊断患有真菌感染。治疗也可以指减少潜在病理而不是仅仅症状的方法。对受试者施用的效果可以具有但不限于以下的效果:减少疾病的一种或多种症状、减少疾病的严重程度、使疾病完全消融或延迟一种或多种症状的发作或恶化。例如,如果受试者的疾病的一种或多种症状与治疗前的受试者相比或与对照受试者或对照值相比时减少约10%,则所公开的方法被视为治疗。因此,所述减少可以是约10%、20%、30%、40%、50%、60%、70%、80%、90%、100%或之间任何数量的减少。
134.如本文所用,预防(prevent)、预防(preventing)或预防(prevention)意指排除、延迟、防止、避免、预先阻止、中止或阻碍疾病或病症的发作、发生率、严重程度或复发的方法。例如,如果易发生真菌感染或易发生真菌感染复发的受试者的真菌感染的发作、发生率、严重程度或复发与易发生真菌感染或易发生真菌感染复发的未接受治疗的对照受试者相比减少或延迟,则所公开的方法被视为预防。因此,真菌感染的发作、发生率、严重程度或复发的减少或延迟可以是约10%、20%、30%、40%、50%、60%、70%、80%、90%、100%或之间任何数量的减少。
135.在一些方法中,受试者免疫受损。例如,受试者可以是接受过干细胞、器官、组织或骨髓移植的受试者,患有癌症的受试者,接受癌症疗法(例如,化学疗法、免疫疗法或放射疗法)的受试者,服用皮质类固醇的受试者,感染hiv或患有获得性免疫缺陷综合征的受试者,患有肝炎的受试者,具有b细胞缺陷的受试者,或具有t细胞缺陷的受试者,仅举几个例子。
136.在一些方法中,受试者患有一种或多种影响受试者肺功能的病症,例如肺纤维化、肺炎、哮喘、慢性阻塞性肺疾病(copd)、囊性纤维化、结核病、肺气肿或结节病。
137.本文提供的方法任选地包括选择患有真菌感染或有真菌感染风险的受试者。本领域技术人员知道如何诊断患有真菌感染的受试者。例如,可以进行医学检查。还可以使用以下一项或多项测试:临床样本的显微镜检查、组织病理学测试、培养物测试和血清学测试。还可以使用临床样本的分子诊断和抗原检测(参见,例如,kozel和wickes“fungal diagnostics”,cold spring harb.perspect.med.4(4):a019299(2014))。
138.本文提供的方法任选地进一步包括向受试者施用有效量的第二治疗剂或疗法。所述第二治疗剂或疗法可以在所述脂质体的多重体的施用之前、同时或之后向受试者施用。在一些方法中,所述第二治疗疗法是外科手术。在一些方法中,所述第二治疗剂是第二抗真菌剂。所述抗真菌剂可以是上述多烯抗真菌剂、唑类抗真菌剂、咪唑类、三唑类或棘球白素中的任一种。
139.药物组合物
140.如全文中所用的术语“有效量”被定义为产生所需生理反应例如治疗或预防真菌感染所需的任何量。施用的剂量范围是大到足以产生使疾病或病症的一种或多种症状受到影响(例如,减少或延迟)的期望效果的那些剂量范围。所述剂量不应大到引起显著的不良副作用,诸如不需要的交叉反应、不需要的细胞死亡等。通常,所述剂量将随抑制剂的类型,受试者的种类、年龄、体重、一般健康状况、性别和饮食,施用方式和时间,排泄速度,药物组合,以及具体病况的严重程度而变化,并且可以由本领域的技术人员来确定。如出现任何禁忌症,剂量可以由个人医生进行调整。剂量可以变化并且可以按一次剂量施用或者按每天施用或以延长的间隔施用的多次剂量施用。
141.本文所述的任何脂质体可以以组合物,例如药物组合物的形式提供。所述组合物可以包含一种或多种本文所公开的脂质体。任选地,包含一种或多种脂质体的组合物位于试剂盒中。药物组合物包括,例如,包含治疗有效量的本文所述的任何脂质体和药物载体的药物组合物。术语“载体”是指这样的化合物、组合物、物质或结构,其在与化合物或组合物组合时有助于或促进所述化合物或组合物的制备、储存、施用、递送、有效性、选择性或任何其他特征以达到它的预期用途或目的。例如,载体可被选择成使活性成分的任何降解最小化,并使在受试者中的任何不良副作用最小化。此类药学上可接受的载体包括无菌生物相容性药用载体,包括但不限于盐水、缓冲盐水、人工脑脊髓液、右旋糖和水。
142.包含本文所述的任何脂质体的药物组合物可以根据标准技术制备,并且进一步包含药学上可接受的载体。通常,生理盐水将被用作药学上可接受的载体。其他合适的载体包括例如水、缓冲水或盐水、0.4%盐水、0.3%甘氨酸、右旋糖等,包括用于增强稳定性的糖蛋白,诸如白蛋白、脂蛋白和球蛋白。这些组合物通常是无菌的。所述药物组合物还可以包含药学上可接受的赋形剂。此类赋形剂包括本身不会诱导对接受所述组合物的个体有害的免疫应答且可被施用而不会产生不当毒性的任何药剂。药学上可接受的赋形剂包括但不限于液体,诸如水、盐水、甘油、糖和乙醇。药学上可接受的盐可以被包括在其中,例如,无机酸盐,诸如盐酸盐、氢溴酸盐、磷酸盐、硫酸盐等;以及有机酸的盐,诸如乙酸盐、丙酸盐、丙二酸盐、苯甲酸盐等。另外,此类媒介物中可能存在辅助物质,诸如润湿剂或乳化剂、ph缓冲物质等。药学上可接受的载体、赋形剂和包含这些物质的制剂的制备描述于例如,remington:the science and practice of pharmacy,第22版,loyd v.allen等人,编者,pharmaceutical press(2012)。
143.水溶液可以被包装以供使用或者可在无菌条件下被过滤并冻干,可将冻干制品在施用前与无菌水溶液混合。所述组合物可以含有接近生理条件所需的药学上可接受的辅助物质,诸如ph调节和缓冲剂、张力调节剂等,例如,醋酸钠、乳酸钠、氯化钠、氯化钾和氯化钙。此外,脂质体悬浮液可包含脂质保护剂,其保护脂质在储存时免受自由基和脂质过氧化损伤。亲脂性自由基猝灭剂诸如α
‑
生育酚和水溶性铁特异性螯合剂,诸如铁链霉素(ferrioxamine)是合适的。
144.药物制剂中脂质体的浓度可以大幅变化,即从小于约0.05重量%,通常为或至少约2
‑
5重量%至多达10重量%至30重量%变化,并且将主要根据选择的特定施用模式,通过流体体积、粘度进行选择。或者脂质体可以被干燥或冻干,并在使用时在水或缓冲液中重新悬浮至所需浓度。脂质体的量或施用的脂质体中活性剂的量取决于所使用的特定标记、所诊断的疾病状态和临床医生的判断,但通常在约0.01与约150mg每千克体重之间,优选介于约0.1与约20mg/kg体重之间、介于约0.1至约10mg/kg体重之间或介于约0.1至约5mg/kg体重之间,其可以按单次剂量或以各个剂量(诸如每天1至4次)的形式施用。施用可以被执行1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20天或更多天。本领域技术人员可根据所述剂的具体特征和接受它的受试者,如下文所述调整剂量。
145.本文公开的组合物以多种方式施用,这取决于是否需要局部或全身治疗以及待治疗的区域。所述组合物通过包括以下的几种施用途径中的任一种施用:经口、鼻内、经吸入、经喷雾器、肠胃外、静脉内、腹膜内、颅内、脊柱内、鞘内、心室内、肌内、皮下、腔内或经皮。药物组合物还可以例如通过局部施用或局部注射被局部递送到需要治疗的区域。药物组合物
还可以通过泵或在手术部位递送。本文所述的任何施用方法的有效剂量可以由来源于体外或动物模型测试系统的剂量
‑
反应曲线外推。
146.公开了可以用于所公开的方法和组合物、可以与所公开的方法和组合物联合使用、可以用于制备所公开的方法和组合物或是所公开方法和组合物的产物的材料、组合物和组分。本文公开了这些以及其它材料,并且应当理解当公开这些材料的组合、子集、相互作用、组等时,虽然可能不能明确地公开对这些化合物的各种单独和集合的组合和排列中的每一个的具体提及,但是每个均被具体考虑并在本文被描述。例如,如果公开和讨论了方法,并且讨论了可以对方法中包括的许多分子进行的许多修饰,则该方法的每个组合和排列以及可能的修改均被具体地考虑,除非明确指示与之相反。同样,还具体考虑并公开了这些的任何子集或组合。此概念适用于本公开的所有方面,包括但不限于使用所公开的组合物的方法中的步骤。因此,如果有多种可以执行的附加步骤,应当理解,这些附加步骤中的每一个都可以按所公开方法的任何特定方法步骤或方法步骤组合来执行,并且每个此类组合或组合的子集均被具体地考虑并且被视为公开。
147.本文引用的出版物以及引用它们的材料都通过引用方式明确地整体并入。
148.实施例
149.提供以下实施例只是为了说明,而不是要进行限制。本领域的技术人员将容易地识别多种可被改变或修改以产生基本上相同或类似的结果的非关键参数。
150.实施例1
151.dectin
‑1152.真菌生长
153.烟曲霉(aspergillus fumigatus)菌株a1163是用kang等人描述的携带绿色荧光蛋白egfp的质粒pbv126(“a dual selection based,targeted gene replacement tool for magnaporthe grisea and fusarium oxysporum.fungal genet biol 42:483
‑
492(2005))在稻瘟病菌核糖体蛋白27启动子控制下转化的,并且在一些实验中被用于监测真菌细胞以产生菌株aek012。使烟曲霉(a.fumigatus)孢子在平板上在vogel最低培养基(vmm,1%葡萄糖、1.5%琼脂)中生长7天,并在pbs 0.1%吐温中收集分生孢子。为进行荧光脂质体定位和生长抑制与杀伤测定,将20,000和4,500个aek012分生孢子分别平板接种于24孔和96孔聚
‑
l
‑
赖氨酸包被的平板上的vmm,1%葡萄糖,0.5%bsa中,在37℃下保持从8小时到4天不等的时间(2001第7章。using microscopy to explore the duplication cycle,p 119
‑
125.in talbot nj(编者),molecular and cellular biology of filamentous fungi:a practical approach oxford university press,university of exeter,exeter,uk;以及sasaki等人,(heterochromatin controls gammah2a localization in neurospora crassa.eukaryot cell 13:990
‑
1000(2014))。使白色念珠菌(candida albicans)sc5314和新型隐球菌(cryptococcus neoformans)h99在马铃薯右旋糖液体培养基中预生长过夜。然后,将细胞用无菌水洗涤3次并再悬浮于roswell park memorial institute(rpmi)培养基中,并使其在聚
‑
l
‑
赖氨酸包被的平板上在37℃下生长10小时。所有真菌细胞生长均在bsl2实验室中进行。为进行经脂质体染色的真菌的荧光成像,将所有三种真菌用pbs洗涤3次,在含4%甲醛的pbs中固定15至60分钟,洗涤一次并在4℃下储存在pbs中。
154.可溶性dectin
‑
1的生产
155.具有被克隆到pet
‑
45b(genscript)中的ms
‑
sdectin
‑
1的示例性密码子优化的大肠杆菌(e.coli)表达构建体的序列示于图1中。该构建体编码经略微修饰的198aa长的sdectin
‑
1蛋白,该蛋白含有载体指定的n末端(his)6亲和标签、柔性间隔区、两个赖氨酸残基、另一个柔性间隔区,其后是c末端176aa鼠类sdectin
‑
1结构域。从在不进行iptg诱导的情况下在luria肉汤中过夜生长的1l细菌培养物(bl21菌株)开始,获得大约45mg/l的22kda sdectin
‑
1(图2)。从在ph=8.0的6m盐酸胍(guhcl,fisher bioreagents bp178),0.1mna2hpo4/nah2po4,10mm三乙醇胺,100mm nacl,5mm bme,0.1%triton
‑
x100中的细胞沉淀物中提取sdectin
‑
1。在这种缓冲液中,将sdectin
‑
1与镍亲和树脂(qiagen,#30210)结合,在调节至ph 6.3的这种缓冲液中洗涤,并在调节至ph 4.5的这种缓冲液中洗脱。立即用1m ph 10.0m三乙醇胺将洗脱蛋白质的ph值中和至ph 7.2用于长期储存。回收了40mg纯度高于95%的蛋白质(图2)。将在添加有新鲜5mm bme的这种相同guhcl缓冲液中的6ug/ul的sdectin
‑
1样本用三乙醇胺进一步调节至ph 8.3,并在23℃下与4摩尔过量的脂质载体试剂dspe
‑
peg
‑
3400
‑
nhs(nanosoft polymers,1544
‑
3400)反应1小时,以制备dspe
‑
peg
‑
dec。对在复性和储存缓冲液rn#5(0.1m nah2po4、10mm三乙醇胺,ph 7.2,1m l
‑
精氨酸、100mm nacl、5mm edta、5mm bme)中的bio
‑
gel p
‑
6丙烯酰胺树脂(bio
‑
rad#150
‑
0740)执行凝胶排阻色谱法以去除未掺入的dspe
‑
peg和guhcl。rn#5的组成是通过测试大量温和的变性且拥挤(crowding)的缓冲液凭经验确定,大部分所述缓冲液导致修饰的dectin
‑
1蛋白在几天后从溶液中脱落。当储存在rn#5中时,该蛋白质无限期地保留在溶液中,如果用bme新鲜还原,则当从rn#5稀释到正常生物缓冲液中时,它可能很容易复性为活性碳水化合物结合形式。dspe
‑
peg
‑
bsa是由牛血清白蛋白bsa(sigma,a
‑
8022)通过相同的方案制备。
156.两性霉素b、sdectin
‑
1、bsa和罗丹明向脂质体中的远程加载
157.无菌聚乙二醇化脂质体是从formumax sci.inc.获得(dspc:chol:mpeg2000
‑
dspe,50:45:5mole%,直径100nm,含60umol/ml脂质的脂质体悬浮液,~4
×
10
12
个脂质体/ml,#f10203a)。商业ambisomes(注射用两性霉素b脂质体,gilead,avanti)含有大约11摩尔百分比的amb。将相对于脂质体脂质的11摩尔百分比的两性霉素b(amb,sigma a4888)远程加载到小批量脂质体中,以制备在整个研究中使用的ambisome样amb
‑
ll。例如,通过在60℃下加热10至20分钟并偶尔混合来将amb(1.8mg,1.95umol)溶解在13ul dmso中,以制备油状透明棕色amb溶液。将250ul无菌脂质体悬浮液(15微摩尔脂质体脂质)加入amb油中,并在旋转平台上于37℃混合96小时,此时大部分amb被嵌入到脂质体中。如通过在a406处的分析所量化,未掺入的amb(0.3umol)保留在油相中,而1.65umol结合在脂质体中,使得脂质体含有相对于脂质的摩尔数的11摩尔百分比的amb。在单独的制备物中,对biogel a
‑
0.5m琼脂糖树脂(biorad 151
‑
0140)执行凝胶排阻色谱法并检查在a406处的排除的流分,确认脂质体中保留了11摩尔百分比的amb。通过从含有较高或较低量amb的油相开始,可以将较大或较小量的amb加载到脂质体中。这些脂质体含有大约11摩尔百分比的amb,在全文中称为amb
‑
ll。
158.通过在60℃下孵育60分钟来使rn#5缓冲液中的dspe
‑
peg
‑
sdectin
‑
1和dspe
‑
peg
‑
bsa缀合物经由其dspe部分,以相对于脂质体脂质摩尔数的1.0和0.33摩尔百分比的蛋白质整合到amb
‑
ll的磷脂双层膜中,以制备dec
‑
amb
‑
ll和bsa
‑
amb
‑
ll。在此相同的60℃孵育期
间,红色荧光标签、罗丹明丽丝胺b
‑
dhpe三乙醇胺盐(invitrogen,#l1392)也以相对于脂质体脂质的2摩尔百分比被掺入(yao等人,phlip(r)
‑
mediated delivery of pegylated liposomes to cancer cells.j control release 167:228
‑
237(2013);he等人,immunoliposome
‑
pcr:a generic ultrasensitive quantitative antigen detection system.j nanobiotechnology 10:26(2012);以及garrett等人,liposomes fuse with sperm cells and induce activation by delivery of impermeant agents.biochim biophys acta 1417:77
‑
88(1999))。对biogel a
‑
0.5m树脂的凝胶排阻色谱确认,在这些摩尔比下,罗丹明
‑
dhpe和dspe
‑
peg
‑
蛋白质在脂质体中的插入基本上是定量的。在4℃下储存的dec
‑
amb
‑
ll保持结合特异性约2个月。在自由条件下储存可以延长货架期。
159.脂质体结合的显微镜检查
160.将福尔马林固定的或活的真菌或动物细胞与dec
‑
amb
‑
ll、bsa
‑
amb
‑
ll和amb
‑
ll脂质体一起在脂质体稀释缓冲液ldb(pbs ph 7.2,0.5%bsa、5mm bme)中孵育,并在相同的ldb中孵育15分钟至2小时后(参见各附图)洗去未被结合的脂质体。罗丹明红色荧光脂质体、绿色egfp烟曲霉(a.fumigatus)和微分干涉对比(dic)照明细胞的图像是在leica dm6000b自动显微镜上,以63x在油浸下拍摄(图4a、4b、5a、5b、6a、6b)。从板中取出细胞并铺展在显微镜载玻片上。以一微米的间隔记录七张z堆栈图像,并将其在adobe photoshop cc2018中合并。在olympus ix70倒置显微镜和olympus pen e
‑
pl7数码相机上,以10x、20x或40x直接拍摄微量滴定板上细胞的明场和/或红色和/或绿色荧光图像,并在photoshop中合并明场和/或彩色图层(图4c
‑
4f、5c
‑
5f、6c
‑
6f)。
161.细胞生长和存活力测定
162.将脂质体储备液以800um amb储存,将其首先在脂质体稀释缓冲液(ldb)或生长培养基中稀释2至20倍,然后在含有细胞的生长培养基中进一步稀释10倍或20倍,以便在指定浓度下使用。总稀释倍数通常为250倍至4,000倍。按照生产商的说明书(promega,文档#g8080)如下进行celltiter
‑
blue(ctb)细胞存活力测定:使用20ul底物处理100或200ul在生长培养基中的真菌或动物细胞,并在37℃下孵育4小时,然后通过加入50ul 3%sds来中止反应。在biotek synergy ht微量滴定板读数器中测量酯酶ctb产物(ex485/em590)的红色荧光。对于每个数据点,对来自六个孔的数据进行平均并计算标准误差(图8a、8c)。发芽(图8e
‑
8f)和菌丝长度(图8b、8d)测定的数据是从以10x和/或20x拍摄的多张拍摄图像中手动收集的。在将细胞固定在4%甲醛和pbs中后,在ex495/em520的微量滴定板读数器上对表达egfp的烟曲霉(a.fumigatus)a1163细胞存活力进行绿色荧光测定(图9)。
163.结果
164.负载两性霉素b的sdectin
‑
1包被的脂质体的制备
165.在聚乙二醇化脂质体中远程加载相对于脂质体脂质的摩尔数的11摩尔百分比的amb,以制备对照负载amb的脂质体amb
‑
ll,其在结构和amb浓度上与商业ambisomes(gilead ambisomes)相似。将sdectin
‑
1(dec,图1、图2)和牛血清白蛋白(bsa)用聚乙二醇化脂质载体dspe
‑
peg修饰。将1摩尔百分比的dspe
‑
peg
‑
dec掺入到amb
‑
ll中,以制备sdectin
‑
1包被的dec
‑
amb
‑
ll(图3),并将0.33摩尔百分比的dspe
‑
peg
‑
bsa掺入到amb
‑
ll中,以制备bsa
‑
amb
‑
ll。sdectin
‑
1(mw 22kda)和bsa(mw 65kda)的这种摩尔比导致这两组脂质体上面包被了等同微克量的蛋白质。因为这些蛋白质包被的脂质体是由相同的amb
‑
ll制成,所以所有
三种脂质体制品相对于脂质摩尔数都含有11摩尔百分比的amb。将两摩尔百分比的dhpe
‑
罗丹明加载到所有三类脂质体中,以制备红色荧光amb
‑
ll、bsa
‑
amb
‑
ll和dec
‑
amb
‑
ll。
166.sdectin
‑
1包被的脂质体dec
‑
amb
‑
ll与真菌细胞强烈结合
167.在对烟曲霉(a.fumigatus)幼苗进行的测定中,罗丹明红色荧光dec
‑
amb
‑
ll与膨大的分生孢子和芽管强烈结合,如图4所示。sdectin
‑
1靶向脂质体通常大量结合或聚集到特定区域。虽然100nm脂质体太小而无法通过光学显微镜分辨,但单个脂质体可见为大小有些均匀的小红色荧光点(橙色箭头,图4a)。因为每个脂质体均含有超过一千个罗丹明分子(图3),所以它们每个均发出强到足以作为单个脂质体可视化的荧光。基本上所有幼苗都结合dec
‑
amb
‑
ll(图4c和4d)。未检测到与未发芽分生孢子的结合(未显示)。ambisome样amb
‑
ll(图4b)和牛血清白蛋白包被的脂质体,bsa
‑
amb
‑
ll(图4e和4f)未与分生孢子或芽管可检测地结合。dec
‑
amb
‑
ll的最大标记在15至30分钟内达到,并且当平板在pbs中在4℃下在暗处储存时,与细胞结合的dec
‑
amb
‑
ll的强红色荧光信号可维持数周。
168.dec
‑
amb
‑
ll还与来自更成熟细胞的膨大分生孢子和菌丝结合,如图5所示。同样,sdectin
‑
1靶向的脂质体通常以团块形式结合,但一些大小相当均匀的各个小红点是可见的(橙色箭头,图5a),它们似乎是单个荧光脂质体。与芽管的标记不同,dec
‑
amb
‑
ll仅与成熟菌丝的子集结合(图5c和5d),如关于各种sdectin
‑
1制品的结合的早期报告中所示。amb
‑
ll与成熟菌丝(图5e和5f)和bsa
‑
amb
‑
ll(未示出)都没有可检测地结合。最后,dec
‑
amb
‑
ll还标记了白色念珠菌(candida albicans)菌丝和新型隐球菌(cryptococcus neoformans)h99细胞(图6)。简而言之,dectin包被的负载两性霉素b的脂质体与真菌细胞有效地结合,而未包被的ambisome样脂质体和bsa包被的脂质体则不能有效地结合。
169.dec
‑
amb
‑
ll和对照ll与固定真菌细胞和活真菌细胞的结合是通过在洗掉未结合的脂质体后计算与烟曲霉(a.fumigatus)菌丝的密集区域结合的单个荧光脂质体和荧光脂质体团块的数量来进行量化。图7a
‑
f显示dec
‑
amb
‑
ll与固定菌丝和活菌丝结合的频率比对照脂质体,bsa
‑
amb
‑
ll或amb
‑
ll高100倍以上。图7g
‑
i显示通过添加可溶性β
‑
葡聚糖,海带多糖可将sdectin
‑
1包被的dec
‑
amb
‑
ll的结合抑制50倍以上,但蔗糖则不能抑制该结合,证明与真菌细胞的结合是β
‑
葡聚糖特异性的。
170.dec
‑
amb
‑
ll对真菌的杀灭和生长抑制
171.用这样的脂质体(其递送的amb浓度接近针对各种烟曲霉(a.fumigatus)菌株估计的2至3um amb的ed
50
以及估计的0.5至1um的mic)处理烟曲霉(a.fumigatus)后,执行各种真菌细胞生长和存活力测定。在这些实验中的大多数中,使4,500个分生孢子发芽并将其与负载药物的脂质体一起在96孔微量滴定板中孵育36至56小时。图8显示靶向dec
‑
amb
‑
ll(dectin
‑
1包被的负载amb的脂质体)比bsa
‑
amb
‑
ll或未包被的amb
‑
ll更有效地杀灭烟曲霉(a.fumigatus)细胞或抑制烟曲霉细胞的生长。使用celltiter blue(ctb)试剂进行的测定表明,用递送3um amb的dec
‑
amb
‑
ll处理细胞能杀灭烟曲霉(a.fumigatus),杀灭效果比携带相同量的药物的ambisome样amb
‑
ll或bsa包被的脂质体bsa
‑
amb
‑
ll高一个数量级(图8a)。ctb试剂测定作为细胞完整性和存活力的替代的总细胞质酯酶活性。作为对脂质体活性进行评分的第二种方法,检查菌丝长度。菌丝长度测定给出了令人惊讶的相似结果,表明递送3um amb的dec
‑
amb
‑
ll在抑制菌丝生长方面比amb
‑
ll或bsa
‑
amb
‑
ll有效得多(图8b)。在使用不同的amb远程加载方法、独立的s
‑
dectin
‑
1和bsa和罗丹明加载以及不同的脂质体
稀释缓冲液进行的完整的生物学重复实验中,在递送3um amb时获得了类似但稍微不那么显著的结果(图8c和8d)。ctb试剂和菌丝长度测定表明,dec
‑
amb
‑
ll在杀灭烟曲霉(a.fumigatus)或抑制烟曲霉的生长方面的有效性比amb
‑
ll高几乎一个数量级。
172.使用脂质体amb活性的附加测定,其测量在各种脂质体制品存在下发芽的分生孢子的百分比(图8e和8f)。递送低至0.09um和0.187um amb的dec
‑
amb
‑
ll抑制烟曲霉(a.fumigatus)分生孢子发芽的有效性比amb
‑
ll或bsa
‑
amb
‑
ll高出几倍。
173.使用脂质体活性的第四种测定法,检查了由表达egfp的烟曲霉(a.fumigatus)aek012菌株产生的内源性绿色荧光信号(图9)。在测定绿色荧光水平时,递送2um或0.67um amb的dec
‑
amb
‑
ll在抑制真菌细胞生长方面的有效性再次比amb
‑
ll或bsa
‑
amb
‑
ll更有效。总之,各种测定表明靶向dec
‑
amb
‑
ll比未包被的amb
‑
ll或bsa包被的bsa
‑
amb
‑
ll更有效地杀灭烟曲霉(a.fumigatus)细胞或者抑制烟曲霉细胞的生长或发芽。
174.测定分生孢子发芽百分比的剂量
‑
反应曲线示于图8g中,说明了三种类型的脂质体的性能存在很大差异。dec
‑
amb
‑
ll优于其他两种类型的脂质体,并且活性与浓度成正比,并且它们在广泛的浓度范围上优于amb
‑
ll。bsa
‑
amb
‑
ll的表现总是最差,可能是因为它们阻断了脂质体膜与真菌质膜的随机通道,该通道允许amb
‑
ll结合。这些测定的时间安排影响这些曲线的精确形状,其中早期进行的测定可以更好地解决最低浓度下的差异,而更长的孵育时间通过允许更多高度抑制的细胞生长来解决最高浓度amb的差异。这使得很难在dec
‑
amb
‑
ll优于对照脂质体的整个浓度范围内测定脂质体制品之间的最佳差异。但总体而言,这三类脂质体之间的关系始终保持不变。
175.dec
‑
amb
‑
ll的动物细胞结合和毒性的降低
176.人胚肾hek293细胞存活力的cell titer blue测定表明,脱氧胆酸盐胶束悬浮液中的amb
‑
ll和amb对hek293细胞的毒性高于dec
‑
amb
‑
ll或bsa
‑
amb
‑
ll(图10)。据推测,用蛋白质包被脂质体会减慢动物细胞的质膜对脂质体的吸收。用递送15或30微摩尔amb的各种制品处理细胞两小时,洗掉含有amb的多余物质,将该细胞在37℃c下生长过夜,然后对其进行测定。
177.诊断法
178.包被有500个靶向分子(例如通过柔性接头融合至venus绿色荧光蛋白的n末端半部的sdectin
‑
1单体(例如dec1
‑
vn))并且还包被有500个靶向分子(例如通过柔性接头融合至venus的c末端半部的sdectin
‑
1单体(dec1
‑
vc))的脂质体将快速识别并结合低浓度的真菌β
‑
葡聚糖从而形成sdectin
‑
1二聚体和组装的venus,从而产生强烈的绿色双分子荧光互补(bifc)信号。这些示例性bifc报告基因构建体的dna和蛋白序列参见seq id no:13、14、15、16(图1m、1n、1o和1p)。对于附加序列,也可参见针对dectin
‑
2 bifc融合的seq id no:17、18、19、20(图1q、1r、1s、1t和图27d中的图示)。在本文提供的任何构建体中,可以使用靶向序列的n末端或c末端片段。例如,dectin
‑
1、dectin
‑
2或dectin
‑
3的n末端或c末端片段。在一些实例中,每个脂质体上存在数百或数千个sdectin
‑
1单体确保了检测的快速性和灵敏度。该测定依赖于以下事实:dectins仅以二聚体形式与细胞上或溶液中的葡聚糖或甘露聚糖紧密且不可逆地结合。
179.任选地,当这些真菌靶向脂质体附着于不溶性基质时,它们可以使用标准荧光仪器检测体外大体积、对照缓冲液和血清中甚至较低浓度的真菌细胞表面分子,例如,β
‑
葡聚
糖。当它们暴露于大体积的样本材料中时,与不溶性基质的结合可使荧光信号集中在小体积中以获得更灵敏的检测。除血清外,固定化脂质体可以测定尿液、肺灌洗液或增溶组织提取物中的真菌和可溶性真菌细胞壁材料。
180.在一些实例中,靶向脂质体可以用于检测真菌甘露聚糖。这可以通过例如给脂质体包被大约500个融合至荧光蛋白的n末端半部的sdectin
‑
2单体(例如,venus dec2
‑
vn)并且给相同的脂质体包被大约500个融合至补充荧光蛋白的c末端半部的sdectin
‑
2或dectin
‑
3单体(venus,dec2
‑
vc、dec3
‑
vc)来实现。dectin
‑
2的同源二聚体与真菌壁甘露聚糖强烈结合,但单体不能显著结合。dectin
‑
2和dectin
‑
3之间的异二聚体可以比它们的单独同源二聚体更强烈地结合甘露聚糖。
181.该系统将使得测定:(1)能够在血清与脂质体基质组合后60分钟内完成;(2)能够在一个步骤中完成;(3)能够高度灵敏地检测极低浓度的多糖;(4)便宜;(5)需要最少的操作员培训和专业知识;以及/或者(6)能够检测几乎所有的真菌病原体。这些脂质体应该能够对血液、血清、尿液、肺渗出液、肺、眼睛、咽喉、阴道、皮肤以及手指甲和脚趾甲中的侵袭性真菌进行一步测定。图11示出了用于一步检测真菌β
‑
葡聚糖和甘露聚糖多糖的自由漂浮的或固定化dectin包被脂质体的一种可能模型。一步检测意指脂质体与靶细胞壁成分的结合以及信号产生是生化相关的,即无需额外的处理步骤或试剂即可进行检测。
182.小鼠或人sdectin
‑
1的生化生产是复杂的,因为蛋白质在水性缓冲液中很容易聚集并变得不溶和无活性。如本文所示,sdectin的溶解性问题通过组合多种途径得以克服,所述多种途径包括使用非常短的带电肽标签,在蛋白质提取、纯化和化学修饰过程中纳入6mguhcl,通过在含有蛋白质增溶剂1m精氨酸的缓冲液中执行复性、脂质体加载和储存,以及纳入巯基还原剂。
183.先前的报告显示,小鼠sdectin
‑
1与烟曲霉(a.fumigatus)膨大分生孢子和芽管有效结合,但对成熟菌丝的结合(如果有的话)效率低下,而对未发芽的分生孢子则完全无效(steele等人,the beta
‑
glucan receptor dectin
‑
1recognizes specific morphologies of aspergillus fumigatus.plos pathog 1:e42(2005))。缺乏菌丝结合可能是由于较老的静息细胞表面上β
‑
葡聚糖水平大大降低,或者可能是成熟细胞壁中的β
‑
葡聚糖不太容易接近。在本文中,sdectin
‑
1包被的荧光dec
‑
amb
‑
ll与膨大分生孢子、芽管和菌丝子集有效结合,表明存在于脂质体表面上的本文所述的经修饰的sdectin
‑
1保留了其对β
‑
葡聚糖的正常亲和力。这些结合数据首次证明,sdectin
‑
1的化学修饰形式(例如,dspe
‑
peg
‑
dectin
‑
1)可以维持其真菌细胞结合特异性。此外,荧光dec
‑
amb
‑
ll结合迅速,并能与细胞稳定结合数周。据报道,纯化的sdectin
‑
1与白色念珠菌(candida albicans)圆形酵母细胞结合,并在亲本细胞和成熟芽之间区域的菌丝中,而不与菌丝结合。尽管与烟曲霉(a.fumigatus)菌丝的结合相比dec
‑
amb
‑
ll与白色念珠菌(candida albicans)的结合效率低下,但我们证明了dec
‑
amb
‑
ll在沿着白色念珠菌(candida albicans)菌丝的多个位点处有合理的结合(图6)。可能是与五聚体igm抗体的亲合力相似,包被有超过一千个sdectin
‑
1分子的脂质体的更大的亲合力保证了快速结合和被结合脂质体的非常缓慢的释放。每个脂质体上存在数千个罗丹明分子,这也可以增加检测明确荧光信号的机会。在使用包含bsa阻断剂的不同结合缓冲液以及各种孵育期的大量实验中,从未检测到未包被的amb
‑
ll或bsa
‑
amb
‑
ll对真菌细胞的显著亲和力。尽管在未将bsa阻断剂纳入孵育中的初步实验中,bsa
‑
amb
‑
ll与烟曲霉
(a.fumigatus)膨大分生孢子结合较弱,
184.但曲霉属、念珠菌属和隐球菌属物种属于三个进化上完全不同的真菌群,即半子囊菌纲、真子囊菌纲和膜菌纲,它们与共同祖先相隔数亿年。dec
‑
amb
‑
ll与所有三个真菌群都特异性结合,表明sdectin
‑
1靶向脂质体可以接近在许多病原真菌的外细胞壁中发现的β葡聚糖。然而,与曲霉属相比,dectin
‑
1与念珠菌的结合相对较弱,而dectin
‑
2的结合则较强。这表明真菌病原体的稳健泛真菌检测可能需要检测葡聚糖和甘露聚糖二者的测定法。
185.如本文所示,在使用不同细胞测定方法的各种生物学实验性重复实验中,dec
‑
amb
‑
ll比递送相同水平的amb的ambisome样amb
‑
ll更有效地杀灭或抑制烟曲霉(a.fumigatus)细胞。在我们所有的实验中,dec
‑
amb
‑
ll在测试的各种amb浓度(其接近或低于所估计的3um的ed
50
)中比对照脂质体的杀真菌能力高几倍至超过一个数量级。检测到dec
‑
amb
‑
ll的活性显著高于amb
‑
ll,甚至在amb浓度低至0.094um amb(远低于amb的mic)时。这些研究表明,dec
‑
amb
‑
ll显著降低了amb杀灭烟曲霉(a.fumigatus)和抑制烟曲霉生长的ed
50
和mic。
186.未包被脂质体的脂质膜与动物细胞(其与真菌细胞不同,不受细胞壁保护)的脂质膜被动结合。然而,脂质体的蛋白包衣以及用小鼠或人血清白蛋白进行的包衣,尤其减少了脂质体从动物模型中的清除率,因此增加了脂质体的半衰期。据推测,蛋白包衣减缓了脂质体膜与质膜的直接相互作用,从而减少药物的被动递送。用sdectin
‑
1对脂质体进行蛋白包被可减少脂质体膜与动物细胞质膜的直接相互作用,从而减少它们与动物细胞的相互作用和融合。此处描述的结果(其显示dec
‑
amb
‑
ll相对于amb
‑
ll对人肾细胞的毒性降低)与该观点一致。
187.总之,当任选地与聚乙二醇化脂质载体缀合并作为单体插入到脂质体中时,sdectin
‑
1能够形成功能复合物并有效结合多种多样的真菌物种的细胞壁中的β
‑
葡聚糖。对递送从0.094至3um的amb浓度的dec
‑
amb
‑
ll进行的多重生长抑制和存活力测定表明,sdectin
‑
1靶向脂质体使脂质体amb的ed50降低至远低于如通过我们的amb
‑
ll建模的针对非靶向ambisomes报告的ed50。此外,与ambisome样脂质体相比,dec
‑
amb
‑
ll对动物细胞的亲和力较低且对动物细胞的毒性较低。综合考虑这些结果,使用sdectin
‑
1包被的脂质体作为泛真菌载体来靶向抗真菌治疗是合理的。
188.实施例2
189.dectin
‑2190.细胞培养
191.将在adh1启动子控制下表达gfp的白色念珠菌(candida albicans)cai4、烟曲霉(a.fumigatus)a1163和野生型新型隐球菌h99
‑
α在呈液体形式的vogel最低培养基(vmm) 1%葡萄糖(85) 0.5%bsa或不含红色指示染料的rpmi 1640培养基(thermofisher sku
‑
11835
‑
030) 0.5%bsa或ypd(1%酵母提取物、2%蛋白胨、2%右旋糖)中在振荡下或在24或96孔聚苯乙烯微量滴定板上或在玻璃显微镜室载玻片上生长,并在37℃下孵育3至36小时。对于烟曲霉(a.fumigatus),将平板用聚
‑
l
‑
赖氨酸预包被,对于所有三种物种,玻璃显微镜载玻片都被预包被。所有真菌细胞生长均在bsl2实验室中进行。在用荧光脂质体处理细胞以用于结合的显微镜分析之前,将真菌细胞用pbs洗涤三次,在含4%甲醛的pbs中固定60分钟,洗涤三次,并在4℃下储存在pbs中。
192.使人结肠直肠腺癌细胞系ht
‑
29(atcc htb
‑
38)和人胚肾细胞系hek
‑
293(atcc crl
‑
1573)在96孔微量滴定板中,在不含红色指示染料加10%胎牛血清的rpmi培养基中在37℃培养箱中在补加有5%co2的空气中生长。使用在培养基中以1:10稀释的ctb试剂并在37℃下孵育60至90min,每次处理8孔,测定所述细胞系在过夜抗真菌处理后的存活力和代谢活性。
193.sdectin
‑
2的生产和化学修饰
194.dectin
‑
2的羧基末端含有其甘露聚糖识别结构域sdectin
‑
2。图13示出了密码子优化的大肠杆菌表达构建体的序列,其具有由genscript合成并克隆到pet
‑
45b中的mmsdectin
‑
2lyshis。长度为577个碱基对的dna序列编码轻微修饰的189a.a.长度的sdectin
‑
2蛋白,其含有载体指定的n末端(his)6亲和标签、附加的柔性glyser间隔区、含有用于交联的甘氨酸残基的序列lysglylys、另一个柔性间隔区,随后是c末端166a.a.长度的鼠sdectin
‑
2结构域。使经修饰的sdectin
‑
2蛋白dec2在大肠杆菌中表达,并如上文所述的那样被纯化用于小鼠sdectin
‑
1,然后在sephacryl s
‑
100hr(ge healthcare,#17061210)上执行额外的凝胶排阻色谱步骤。在用考马斯蓝染色的sds page凝胶上显示了蛋白质,如图14所示。使用1m ph 10三乙醇胺将该相同guhcl缓冲液中5μg/ul的sdectin
‑
2样本(其中新添加有5mm 2
‑
巯基乙醇)调节至ph 8.3,并使其在23℃下与4摩尔过量的dspe
‑
peg
‑
3400
‑
nhs(1,2
‑
二硬脂酰
‑
sn
‑
甘油
‑3‑
磷酸乙醇胺(dspe)缀合的聚乙二醇(peg),来自nanosoft polymers,1544
‑
3400)的反应性琥珀酰亚胺酯nhs部分反应1小时,以制备dec2
‑
peg
‑
dspe(补充图s1)。在复性和储存缓冲液rn#5(0.1m nah2po4、10mm三乙醇胺,ph 8.0,1m l
‑
精氨酸、100mm nacl、5mm edta、5mm 2
‑
巯基乙醇)中通过bio
‑
gel p
‑
6丙烯酰胺树脂(bio
‑
rad#150
‑
0740)执行分子排阻色谱以去除未掺入的dspe
‑
peg和guhcl(51)。bsa
‑
peg
‑
dspe是按相同方案用bsa(牛血清白蛋白,sigma,a
‑
8022)制备,但在dspe
‑
peg标记过程中使用不含guhcl的缓冲液并在bio
‑
gel p6层析过程中使用l
‑
精氨酸。罗丹明标记的sdectin
‑
2(dec2
‑
rhod)是通过用于制备dec2
‑
peg
‑
dspe的相同程序在相同的guhcl缓冲液中制备,但是标记是用4摩尔过量的罗丹明
‑
nhs试剂(thermo fisher#46406)在sdectin
‑
2上进行的。在rn#5缓冲液中,在bio
‑
gel p2树脂上通过分子排阻色谱法从dec2
‑
rhod中除去水解的未结合罗丹明试剂和不需要的盐。与dec2
‑
peg
‑
dspe一样,rn#5将dec2
‑
rhod保持在这样的状态下,在该状态下dec2
‑
rhod在被稀释到较温和的生物缓冲液中时很容易复性为活性碳水化合物结合形式。
195.amb、sdectin
‑
2、bsa和罗丹明向脂质体中的远程加载
196.从来自formumax sci.inc.的无菌聚乙二醇化脂质体(dspc:chol:mpeg2000
‑
dspe,formumax#f10203a)开始,相对于100%脂质体脂质(amb,sigma a4888)制备了具有11摩尔百分比amb的小批量脂质体,以制备如上所述的amb
‑
ll(参见表1)。
197.表1
[0198][0199]
与上述方案并行地,通过在60℃下孵育30分钟来使在rn#5缓冲液和pbs中的dec2
‑
peg
‑
dspe
‑
和bsa
‑
peg
‑
dspe缀合物经由其脂质dspe部分整合到amb
‑
ll的磷脂双层膜中,以制备dec2
‑
amb
‑
ll和bsa
‑
amb
‑
ll。在这些相同的60℃孵育期间,还将2摩尔百分比的红色荧光标签罗丹明丽丝胺b
‑
dhpe(invitrogen,#l1392)掺入到sdectin
‑
2包被和bsa包被的脂质体和amb
‑
ll中。将dec2
‑
amb
‑
ll的重复样本在bio
‑
gel a 0.5m树脂(bio
‑
rad,#151
‑
0140)上进行凝胶排阻色谱。从树脂中有效地排除了荧光脂质体。由于在凝胶中所包含的低分子量流分中未检测到sdectin
‑
2(2.2od a
280
/mg/ml)或罗丹明,因此我们得出结论,两者均可有效地加载至脂质体中。在每次用于结合或杀灭测定之前,向dec2
‑
amb
‑
ll中添加新鲜的2mm bme。在rn#5中以4℃储存的dec2
‑
amb
‑
ll似乎保留完整的真菌细胞结合特异性和杀灭活性至少12个月。
[0200]
与真菌细胞结合的脂质体和dec2
‑
rhod的显微镜检查
[0201]
将福尔马林固定的真菌细胞与脂质体一起在23℃下在脂质体稀释缓冲液ldb2(20mm hepes、10mm三乙醇胺、150mm nacl、10mm cacl2、1mmβ
‑
巯基乙醇(bme)、5%bsa,ph 8.0)中孵育,其中bme为新鲜添加的。将脂质体储备液按1:200稀释,然后将其与细胞一起孵育,使得sdectin
‑
2蛋白浓度为0.5μg/100μl。15分钟、1小时或更长时间的孵育后,通过更换4次ldb2洗去未被结合的脂质体。罗丹明红色荧光脂质体、绿色荧光细胞和微分干涉对比
(dic)照明细胞的合并图像是在leica dm6000b自动显微镜上在油浸下按63x从在显微镜载玻片上生长的细胞拍摄的。将dec2
‑
rhod储备液也按1:200稀释,然后将其与细胞一起孵育,使得sdectin
‑
2蛋白浓度也为0.5μg/100μl。在olympus ix70倒置显微镜上使用olympus pen e
‑
pl7数码相机按20x拍摄微量滴定板上细胞的明场、dic和红色(ex560/em645)以及绿色(ex500/em535)荧光图像,并在photoshop中合并明场和/或荧光彩色层。将未修改的红色荧光tif图像的8位灰度副本放入image j(imagej.nih.gov/ij),使用image>adjust>threshold>apply仅捕获脂质体照亮的红色荧光区域,并使用analyze>measure将每个图像的区域数据放入文件中,如此对20x放大倍数下脂质体结合的面积进行量化。通常分析6
‑
10张照片图像,并计算大多数面积估计值的平均值。然而,由于不同摄影视野中新型隐球菌(c.neoformans)细胞之间的染色强度差异很大,因此每次处理分析了90张图像。还在leica dm6000b自动显微镜上,以20x或在油浸下以63x拍摄显微镜室载玻片中生长的细胞的明场、dic和荧光图像。
[0202]
葡糖苷酸木脂甘露聚糖(gxm)特异性单克隆抗体18b7获自sigma
‑
aldrich(mabf2069),以1:200稀释倍数(0.5μg/100μl)使用,用也以1:200稀释的山羊抗小鼠第二抗体alexa488(life technologies,a11001)显色,并用gfp滤光片(ex500/em535)拍照。
[0203]
脂质体处理后的生长抑制和存活力测定
[0204]
将脂质体储备液以615至800μm amb储存,通常先30至600倍稀释到脂质体稀释缓冲液ldb2中,然后以1:11稀释到生长培养基中,以达到指示的最终杀菌剂浓度范围2μm到0.1μm。对照细胞接受等量的ldb2。白色念珠菌(candida albicans)和烟曲霉(a.fumigatus)的celltiter
‑
blue(ctb)细胞存活力和代谢活性测定按照我们最近针对烟曲霉(a.fumigatus)的描述的那样进行:与ctb试剂一起孵育3至4小时,并在bio
‑
tek synergy ht荧光微量滴定板读数器中分析96孔板。从实验孔中减去对照孔的荧光背景。对于每个数据点,取八个孔的数据的平均值。这些测定有大量背景,并且如果在vmm中测定细胞,则不太灵敏。当使用此方案或针对该物种发布的其他ctb协议对新型隐球菌(cryptococcus neoformans)进行平行细胞存活力测定时,我们无法检测到来自ctb试剂的任何荧光信号。作为对新型隐球菌(c.neoformans)和白色念珠菌(candida albicans)细胞存活力的替代测量,通过使1ml细胞在ypd中生长、在指定的生长时间内添加负载药物的脂质体、稀释所述细胞、在ypd上平板接种和计数菌落形成单位(cfu)而进行测定。通过向培养基中加入50μg/ml碘化丙啶并在37℃下孵育60min来测定经负载杀菌剂的脂质体处理后在ypd中生长的细胞中死亡新型隐球菌(c.neoformans)细胞的分数。移除培养基,用pbs替换,以便使用红色荧光蛋白通道(ex560/em595)进行荧光显微镜检查,并相对于染色和未染色细胞的总数对死亡染色细胞的百分比进行评分。在使用dec1
‑
amb
‑
ll的实验中,所有脂质体都用ldb1(pbs 5%bsa 1mm bme)(51)而不是ldb2稀释。
[0205]
dectin
‑
2是由人和小鼠c型lectin受体基因clec6a编码。dectin
‑
2结合甘露糖蛋白中的α
‑
甘露聚糖以及n连接和o连接的甘露聚糖(42
‑
46)。dectin
‑
2在一些淋巴细胞的质膜中表达,它的甘露聚糖结合结构域(sdectin
‑
2)在这些细胞的外部,且它的信号传导结构域在细胞质中。dectin
‑
2起到先天免疫受体的作用,向宿主发出活动性真菌感染信号。
[0206]
如实施例1中所阐述,dectin
‑
1包被的负载两性霉素b(amb)的脂质体靶向内细胞壁中的β
‑
葡聚糖,并有效结合多种细胞类型的烟曲霉(a.fumigatus)和新型隐球菌
(cryptococcus neoformans)酵母细胞。dectin
‑
1包被的负载amb的脂质体(dec1
‑
amb
‑
ll)可有效地抑制和杀灭烟曲霉(a.fumigatus)细胞。然而,dectin
‑
1脂质体与白色念珠菌(candida albicans)的结合很差,据推测是由于厚的甘露聚糖和甘露糖蛋白外层的存在掩盖了它们的β
‑
葡聚糖。在本文中,给负载amb的脂质体包被小鼠dectin
‑
2的甘露聚糖结合结构域sdectin
‑
2。与非靶向负载药物的脂质体相比,sdectin
‑
2包被的负载amb的脂质体与白色念珠菌(candida albicans)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)有效结合,并显著降低细胞生长和存活力。
[0207]
结果
[0208]
负载杀菌剂的sdectin
‑
2包被的荧光脂质体的制备
[0209]
图15中示出了本文构建的负载杀菌剂的sdectin
‑
2包被的脂质体的模型。脂质体构建方法和脂质体组合物与上述sdectin
‑
1包被的脂质体密切相似。将相对于脂质体脂质摩尔数的11摩尔百分比的两性霉素b(amb)远程加载到聚乙二醇化脂质体的膜中,以制备负载amb的脂质体amb
‑
ll。作为参考,广泛使用的商业化负载amb的非靶向脂质体产品含有相对于脂质体脂质的摩尔数的10.6摩尔百分比的amb(表1)。鼠类sdectin
‑
2序列被设计为在其氨基末端含有小赖氨酸标签(图13)。使它在大肠杆菌中表达(图14),并使纯化的sdectin
‑
2蛋白经由这种赖氨酸标签缀合至nhs
‑
peg
‑
dspe,从而制成dec2
‑
peg
‑
dspe。然后,将dec2
‑
peg
‑
dspe经由其dspe脂质部分以相对于脂质体脂质的摩尔数的1摩尔百分比的蛋白分子(每个脂质体1,500个sdectin
‑
2分子)掺入到amb
‑
ll中,以制备dec2
‑
amb
‑
ll。类似地,并且作为蛋白质包被的脂质体对照,将0.33摩尔百分比的牛血清白蛋白经由脂质载体掺入到amb
‑
ll中以制备bsa
‑
amb
‑
ll。这导致这两组脂质体的表面上有等同μg量的22kda sdectin
‑
2和66kda bsa蛋白。与商业产品非常相似的未包被amb
‑
ll也可用作脂质体对照。也将2摩尔百分比的dhpe
‑
罗丹明掺入到所有三种脂质体制品的脂质体膜中。因此,所有三组脂质体都含有相同的11摩尔百分比的amb和2摩尔百分比的罗丹明。将dec2
‑
amb
‑
ll的组成与表1中的bsa
‑
amb
‑
ll、amb
‑
ll、和amb/胶束的组成进行比较。
[0210]
sdectin
‑
2包被的脂质体dec2
‑
amb
‑
ll与多种多样的真菌物种的结合比对照脂质体更强
[0211]
sdectin
‑
2包被的红色荧光dec2
‑
amb
‑
ll与白色念珠菌(candida albicans)酵母细胞、假菌丝和菌丝强烈结合(图16)。绝大多数sdectin
‑
2包被的脂质体以大簇形式结合至与这些细胞缔合的细胞外多糖基质。此外,dec2
‑
amb
‑
ll与这些细胞周围的细胞外基质(ex )的大的子集结合,而基质的一些区域明显不结合sdectin
‑
2包被的脂质体(ex
‑
)(图16e)。虽然100nm脂质体太小而无法通过光学显微镜分辨,但每个脂质体的估计3,000个罗丹明分子(图15)可以使来自单独的脂质体的荧光信号可视化。很少看到单独的dec2
‑
amb
‑
ll(白色箭头,图15a)或脂质体簇直接与白色念珠菌(candida albicans)细胞壁结合。相比之下,单独的sdectin
‑
1包被的脂质体经常与烟曲霉(a.fumigatus)细胞的细胞壁结合。
[0212]
dec2
‑
amb
‑
ll与新型隐球菌(cryptococcus neoformans)酵母细胞强烈结合(图17)。单克隆抗体18b7对新型隐球菌(c.neoformans)的荚膜和胞外多糖中发现的葡糖醛木甘聚糖(gxm)具有特异性。抗体18b7染色了大部分(图17b)但不是所有的细胞荚膜以及大部分但不是所有在明场图像中可见的胞外多糖(ex ,图17a)。dec2
‑
amb
‑
ll与胞外多糖基质中
大部分18b7染色的gxm区域强烈共染色,但不染色荚膜的187b标记的gxm(图17c、17d)。此外,有一些细胞外基质区域没有用18b7或dec2
‑
amb
‑
ll染色(ex
‑
/
‑
,图17a)。
[0213]
dec2
‑
amb
‑
ll也以大簇被结合到烟曲霉(a.fumigatus)发芽分生孢子和菌丝产生的胞外多糖基质上(图17e、17f、17g)。同样,很少的结合(如果有的话)与细胞壁本身相关联。此外,似乎存在胞外多糖基质未被染色或染色不良的区域(例如,图17e和17f)。
[0214]
因为dec2
‑
amb
‑
ll与被检查的所有三种真菌物种的细胞壁的紧密交联多糖内的甘露聚糖结合不良或根本不结合,我们认为我们的脂质体的100纳米直径尺寸限制了它们的进入,或者在存在于脂质体膜中时,sdectin
‑
2在某种程度上受到了限制而无法获得全部活性。溶液中的旋转直径dec2
‑
amb
‑
ll甚至大于它们的物理尺寸估计值,这是它们包被有sdectin
‑
2蛋白和相关的水分子。sdectin
‑
2本身具有22kda的原子量,因此溶液中的旋转直径可能估计为约4纳米。制备了罗丹明偶联的sdectin
‑
2、dec2
‑
rhod。罗丹明(0.48kd)和一个或两个罗丹明偶联分子的原子量对dec2
‑
rhod的这种尺寸估计值几乎没有影响。红色荧光dec2
‑
rhod与烟曲霉(a.fumigatus)菌丝细胞周围的大部分胞外多糖基质强烈结合(图18,平板a和b)。结合模式和结合强度与dec2
‑
amb
‑
ll的结合模式和结合强度无法区分(图18,平板c和d)。
[0215]
与未包被的amb
‑
ll和bsa包被的bsa
‑
amb
‑
ll对照相比,对dec2
‑
amb
‑
ll与真菌细胞结合的效率进行量化。荧光脂质体信号的面积是从与真菌细胞几乎汇合的脂质体染色培养物拍摄的多个红色荧光摄影图像测量的(图19)。dec2
‑
amb
‑
ll与白色念珠菌(c.albicans)假菌丝和菌丝、新型隐球菌(c.neoformans)酵母细胞和烟曲霉(a.fumigatus)菌丝结合的效率比amb
‑
ll或bsa
‑
amb
‑
ll高50到150倍(图19a、19d、19g)。为进行这些测量而定量的荧光脂质体摄影图像的实例与每个棒图相邻列出(图19b、19c、19e、19f、19h、19i)。
[0216]
使用与白色念珠菌(c.albicans)假菌丝和菌丝结合的荧光脂质体区域的相同定量测定法,表征了dec2
‑
amb
‑
ll结合的特异性、稳定性和速率(图20)。在结合测定过程中,通过纳入增溶酵母甘露聚糖可将dec2
‑
amb
‑
ll标记抑制75%,但相同浓度的可溶性β
‑
葡聚糖海带多糖或含有二糖蔗糖的葡萄糖
‑
果糖则不能如此(图20a、20b、20c)。该结果确认sdectin
‑
2靶向脂质体与细胞外基质的结合是甘露聚糖特异性的,这与已公布的sdectin
‑
2的碳水化合物特异性一致。dec2
‑
amb
‑
ll与白色念珠菌(c.albicans)细胞结合的稳定性是通过获取图19a中检查的dec2
‑
amb
‑
ll染色制品,并将它们在4℃的磷酸盐缓冲盐水中于暗处储存进行检查。2个月后,对这些细胞进行重新拍照并量化脂质体染色。dec2
‑
amb
‑
ll与细胞结合的荧光强度仍然很强,估计比amb
‑
ll的非特异性结合强50倍(图20d、20e、20f)。该结果表明dec2
‑
amb
‑
ll本身相对稳定,并且它们与细胞的结合也相对稳定。dec2
‑
amb
‑
ll结合速率是通过在洗掉未结合的脂质体之前,将白色念珠菌(c.albicans)假菌丝和菌丝细胞的密集区域暴露于脂质体中10秒至90分钟的时间来估计的(图20g、20h)。dec2
‑
amb
‑
ll标记的面积在前15分钟呈指数迅速增加(图20g),然后减慢,但在90分钟后似乎没有完成(图20h)。
[0217]
总之,dectin
‑
2包被的amb脂质体dec2
‑
amb
‑
ll与体外生长的白色念珠菌(c.albicans)细胞外基质中存在的真菌甘露聚糖特异性地稳定而快速地结合,而几乎没有观察到对照脂质体的非特异性结合。只有痕量的dec2
‑
amb
‑
ll与细胞壁结合。dec2
‑
amb
‑
ll也能有效地与新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)细胞周围的细胞外基质的部分结合。
[0218]
dec2
‑
amb
‑
ll的生长抑制和杀伤
[0219]
在用sdectin
‑
2包被的脂质体和对照脂质体处理白色念珠菌(c.albicans)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)的生长活跃的培养物后,进行了各种真菌细胞生长和存活力测定,所述sdectin
‑
2包被的脂质体和对照脂质体递送的amb浓度接近该杀真菌剂的最低抑菌浓度(mic)(图21)。根据amb的测定条件和递送方法,针对这些物种估计的amb的mic范围为0.06至1.3μm。
[0220]
将4,000个白色念珠菌(c.albicans)酵母细胞接种到96孔微量滴定板的各个孔中,使其生长6小时至假菌丝和早期菌丝阶段,并用负载药物的脂质体处理细胞。孵育30分钟后,洗去脂质体,再使细胞生长16小时。图21a显示递送1μm amb至0.125μm amb的靶向dec2
‑
amb
‑
ll杀灭或抑制白色念珠菌(c.albicans)细胞的效率比递送相同浓度amb的未包被amb
‑
ll或bsa
‑
amb
‑
ll高90倍至3倍。鉴于在这些实验中细胞仅暴露于脂质体药物中30分钟,因此差异是显著的。这些数据是使用celltiter
‑
blue试剂获得的,以评估作为细胞完整性和存活力的替代的细胞质还原酶活性。死细胞或代谢失活的细胞不会将刃天青底物还原为荧光试卤灵产物。用脂质体连续处理白色念珠菌(c.albicans)细胞整个16小时时间或用更高的药物浓度处理导致所有三种负载药物的脂质体样本的细胞死亡过多,无法明确解答不同脂质体制品之间的差异。六个月时间的dec2
‑
amb
‑
ll制品保留了它们的全部抗真菌活性,只要将它们是被新还原的。还测定了脂质体处理后的活细胞数目。将在丰富培养基中的液体培养物中生长的白色念珠菌(c.albicans)酵母细胞与递送2μm amb的脂质体一起孵育。30分钟后洗去脂质体。再生长6小时后,将培养物稀释并用丰富培养基测定琼脂平板上的集落形成单位(cfu)。基于cfu,dec2
‑
amb
‑
ll在抑制或杀灭液体中的白色念珠菌(c.albicans)酵母细胞方面的有效性比amb
‑
ll或bsa
‑
amb
‑
ll的有效性高3倍(图13a)。
[0221]
将液体中生长的新型隐球菌(c.neoformans)酵母细胞用递送0.4μm amb的脂质体处理4小时,并用递送0.4、0.2和0.1μm amb的脂质体处理过夜(图21b)。在每次处理结束时,稀释细胞并用丰富培养基在琼脂板上测定集落形成单位(cfu)。dec2
‑
amb
‑
ll在杀灭新型隐球菌(c.neoformans)方面的有效性比amb
‑
ll或bsa
‑
amb
‑
ll的有效性高2.5至11倍,最佳处理为0.2μm amb过夜。作为替代测定法,用递送1μm amb的脂质体处理微量滴定板上生长的新型隐球菌(cryptococcus neoformans)酵母细胞5小时。通过将细胞与碘化丙啶一起孵育,立即测定细胞的细胞死亡。碘化丙啶进入死细胞,但不会进入活细胞,并在嵌入到双链dna或双链rna的短区域中时发出红色荧光。碘化丙啶测定表明,在这些处理条件下,dec2
‑
amb
‑
ll在杀灭新型隐球菌(c.neoformans)细胞方面的有效性比未包被的amb
‑
ll高出5倍(图13b、13c、13d)。
[0222]
用递送接近并低于针对估计的mic 0.5μm的amb浓度的脂质体处理烟曲霉(a.fumigatus)。当芽管首先开始从95%分生孢子汇合时,分生孢子发芽并生长至非常早期的幼苗阶段。然后,用含amb的脂质体或脂质体稀释缓冲液处理细胞2小时,并用生长培养基洗去未结合的脂质体。使细胞再生长19小时,并用celltiter blue试剂测定细胞的存活力和代谢活性。递送0.5μm和0.25μm amb的dec2
‑
amb
‑
ll杀灭烟曲霉(a.fumigatus)或抑制烟曲霉生长的效率分别比amb
‑
ll高出20倍和36倍(图21c)。应当注意的是,经稀释缓冲液对照处理的烟曲霉(a.fumigatus)细胞在该测定过程中过度生长,并在微量滴定孔中产生厚的菌丝垫。因此,来自这些对照细胞的代谢活性和celltiter blue信号低。
[0223]
靶向amb的负载dectin
‑
1的脂质体也能有效结合烟曲霉(a.fumigatus)膨大分生孢子、幼苗和菌丝细胞,并抑制和杀灭这些细胞,但它们结合的是β
‑
葡聚糖而不是α
‑
甘露聚糖。在这项先前的研究中,在整个测定过程中将细胞与脂质体连续孵育,并不进行洗涤。为了更直接地比较两种dectin的药物靶向效率,使用本文中用于dec2
‑
amb
‑
ll的相同测定设计检查dec1
‑
amb
‑
ll,2小时后洗去脂质体,但是最初将脂质体稀释到ldb1中并使细胞生长16小时。递送0.5μm和0.25μm amb的dec2
‑
amb
‑
ll杀灭烟曲霉(a.fumigatus)或抑制烟曲霉生长的效率分别比amb
‑
ll高出28倍和5倍(图21d)。使用这种测定条件,dectin
‑
1和dectin
‑
2靶向负载amb的脂质体的结果非常相似。
[0224]
dec2
‑
amb
‑
ll对动物细胞的毒性
[0225]
将快速生长的人hek 293和人ht
‑
29细胞培养物用各自递送15μm amb的各种脂质体和脱氧胆酸盐胶束悬浮液处理过夜。基于代谢活性和存活力的celltiter
‑
blue测定,dec2
‑
amb
‑
ll的毒性比amb
‑
ll或bsa
‑
amb
‑
ll高10%至20%,且毒性比amb脱氧胆酸盐(amb/doc)胶束低2至5倍(图14)。当这些细胞被处理成接受较低的amb浓度,例如,3μm amb时,只有amb/doc胶束表现出可测量的毒性。当通过荧光显微镜检查时,这三种脂质体制品似乎对这些细胞系中的每一个都没有任何特殊的亲和力。
[0226]
总之,将dectin
‑
2的n末端结构域sdectin
‑
2与脂质载体偶联,并且将该缀合物插入脂质体中,使每个单体的c末端碳水化合物识别结构域(crd)从脂质体膜朝外。考虑到针对这些dec2
‑
amb
‑
ll观察到与三种不同的人真菌病原体的细胞外基质有效结合,sdectin
‑
2单体必须在构象上是自由的,以形成有效的甘露聚糖结合所需的功能二聚体,c型凝集素受体往往与它们的某些底物结合较弱。已公布的针对50%的sdectin
‑
2与甘露聚糖相关多糖结合估计的有效浓度(ec50)估计值范围为约20mm(甘露糖)、2mm(甘露聚糖
‑
α
‑1‑2‑
甘露聚糖)至150m(甘露聚糖)。与dectin
‑
2最接近的旁系同源物dectin
‑
1相比,这确实是弱结合,dectin
‑
1结合各种β
‑
葡聚糖的ec50范围为2mm至2.2皮摩尔。每个脂质体上的约1,500个sdectin
‑
2单体产生的更高亲合力可能会导致针对dec2
‑
amb
‑
ll与真菌细胞的结合所观察到的快速、强烈和稳定的结合。
[0227]
dec2
‑
amb
‑
ll与所有三个物种的细胞外基质有效结合的数据与它们的胞外多糖中甘露聚糖丰富的含量一致。然而,白色念珠菌(candida albicans)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)基质的某些区域没有被sdectin
‑
2包被的脂质体染色或染色不良,表明基质中甘露聚糖的分布是异质的,或者这些区域中的甘露聚糖被掩蔽而免于暴露于脂质体sdectin
‑
2中。结果表明dec2
‑
amb
‑
ll与胞外多糖甘露聚糖的结合并不比尺寸小得多的的dec2
‑
rhod的结合受到更多限制,表明尺寸限制可能不是主要限制。
[0228]
没有令人信服的证据表明,dec2
‑
amb
‑
ll与这些真菌物种中任一物种的细胞壁的结合高于痕量水平。尽管细胞壁甘露聚糖含量在很大程度上基于生长培养基和化学分析方法而变化,但新型隐球菌(c.neoformans)的估计细胞壁甘露聚糖多糖含量为22%,白色念珠菌(candida albicans)的估计细胞壁甘露聚糖多糖含量为40%,烟曲霉(a.fumigatus)的估计细胞壁甘露聚糖多糖含量为15至41%。尽管罗丹明标记的sdectin
‑
2 dec2
‑
rhod以与dec2
‑
amb
‑
ll相似的强度和模式与烟曲霉(a.fumigatus)的胞外多糖基质结合,但dec2
‑
rhod并未染色细胞壁。因此,再一次地,大很多的dec2
‑
amb
‑
ll的尺寸障碍似乎并不能解释它们的细胞壁结合缺失。因此,这些发现表明细胞壁甘露聚糖可能被化学掩蔽而免于
sdectin
‑
2介导的脂质体结合。该结果与白色念珠菌(candida albicans)细胞壁β
‑
葡聚糖掩蔽免于sdectin
‑
1结合和免于dec1
‑
amb
‑
ll结合相似。
[0229]
白色念珠菌(candida albicans)可在单细胞卵形酵母和多细胞椭圆形假菌丝和细长菌丝形态之间可逆地转换。所有三个阶段都能够产生细胞外基质并粘附到宿主组织上。所有三种被结合的dec2
‑
amb
‑
ll的基质,表明负载杀菌剂的dectin
‑
2包被的脂质体疗法具有降低白色念珠菌(candida albicans)的毒力的潜力。
[0230]
dec2
‑
amb
‑
ll抑制和杀灭白色念珠菌(candida albicans)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)的效率比递送相同浓度的amb的普通未包被的amb
‑
ll或bsa包被的bsa
‑
amb
‑
ll高得多。基于ctb试剂的代谢活性测定、基于cfu的细胞生长测定和死细胞的碘化丙啶染色的组合确认发生了细胞的抑制和杀灭。与dec2
‑
amb
‑
ll一起孵育低至30分钟到几个小时,会导致显著的杀灭。当递送接近或低于针对amb报告的mic值的amb浓度(在该浓度下,等效的未包被amb
‑
ll对细胞抑制或存活率有很小的影响或没有影响)时,dec2
‑
amb
‑
ll抑制或杀灭真菌细胞的效率高3倍至90倍。dec2
‑
amb
‑
ll与所有三个物种的结合比amb
‑
ll高50
‑
150倍,并且在某些试验条件下,抑制和杀灭它们的效率比amb
‑
ll高11
‑
94倍。
[0231]
amb或其他脂质体包装的治疗性杀真菌剂的mic显著降低应导致减少的杀真菌剂剂量和降低的给药频率,从而降低宿主毒性。本文证明了,当递送15μm amb(其比这里用于杀灭真菌细胞的amb浓度高15至150倍)时,dec2
‑
amb
‑
ll对动物细胞没有特别的毒性。
[0232]
这些研究是用小鼠sdectin
‑
2蛋白序列进行,以避免未来在念珠菌病、曲霉病和隐球菌病小鼠模型中测试sdectin
‑
2靶向抗真菌剂期间出现sdectin
‑
2免疫原性的问题。人sdectin
‑
2蛋白序列与小鼠蛋白有72%同一,仅短两个氨基酸。因此,操纵人类蛋白质以靶向负载杀菌剂的治疗性脂质体以用于临床研究是可能的。
[0233]
总之,迫切需要新的抗真菌疗法。dec2
‑
amb
‑
ll与由白色念珠菌(c.albicans)、新型隐球菌(c.neoformans)和烟曲霉(a.fumigatus)的不同细胞阶段产生的细胞外基质有效结合。递送接近或低于amb对所有三个物种的生长抑制和杀灭的mic的amb浓度的dec2
‑
amb
‑
ll证明了,脂质体包装的药物的sdectin
‑
2靶向使抗真菌作用与非靶向脂质体amb相比提高了一个或多个数量级。可以合理地提出,靶向真菌细胞的负载药物的脂质体作为具有广泛应用的泛抗真菌疗法具有巨大潜力。
[0234]
实施例3
[0235]
dectin
‑
2包被的抗真菌脂质体抑制真菌负荷在肺曲霉病小鼠模型中
[0236]
真菌细胞培养物
[0237]
烟曲霉(a.fumigatus)cea10(cbs 144.89,atcc mya1163)的人临床分离株先前已用于曲霉病小鼠模型(参见desobeaux和cray,“rodent models of invasive aspergillosis due to aspergillus fumigatus:still a long path toward standardization,”front microbiol.8,841(2017))。分生孢子是通过使cea10在37℃下在含有vogel最低培养基(vmm) 1%葡萄糖 100ug/ml卡那霉素和氨苄西林中的每一种的1.5%琼脂板上生长6天来制备。通过轻微振摇平板及玻璃珠粒和磷酸盐缓冲盐水 0.05%吐温20来收获分生孢子。经无菌40微米尼龙网过滤器(fisher sci.#22363547,hampton nh)过滤分生孢子,按1xg沉降过夜以浓缩分生孢子,并在血细胞计数器中测定分生孢子细
胞密度。发芽率被证明为接近100%。
[0238]
抗代谢药和类固醇制品
[0239]
在ph 7.4盐水中制备35mg/ml环磷酰胺(cayman#13849)储备液,并以175mg/kg小鼠体重递送。在二水中以22.4mg/ml制备醋酸可的松悬浮液(cayman chemical co.,#23798,ann arbor,mi)并以112mg/kg递送。在dmso中以40mg/ml制备曲安西龙(millipore sigma#t6376,burlington,ma)的储备液,并将其储存在4℃。就在以40mg/kg腹膜内注射之前,将该储备液在pbs中作1:4v/v稀释以制备水悬浮液。
[0240]
免疫抑制介导的肺曲霉病小鼠模型
[0241]
7周龄的远交雌性cd1(cd
‑
1igs)瑞士小鼠获自charles river labs(每只25g至30g)。cd1小鼠已用于大多数实验性曲霉病
56
的研究。类固醇模型(图22a):通过在第
‑
1天(d
‑
1)和第3天腹膜内注射40mg/kg小鼠体重的曲安西龙
63
来对小鼠进行免疫抑制(图22)。白细胞减少模型(图22b):在第
‑
3天使用单次ip注射175mg/kg环磷酰胺,然后在第
‑
1天单次皮下注射40mg/kg曲安西龙,对小鼠进行免疫抑制。由于这些小鼠将在第4天被安乐死以测定真菌负荷,因此没有给予它们后续的免疫抑制剂注射。
[0242]
在第1天,通过口咽抽吸50μl分生孢子样本来使免疫受到抑制的小鼠受到感染(图22)。症状的进展首先是理毛行为和折叠皮毛,然后是轻度嗜睡。一旦小鼠表现出严重的嗜睡和/或它们的体重减轻了25%,它们就会被宣布临床死亡,并在麻醉后通过颈椎脱臼对它们实施安乐死。对于真菌负荷实验,所有动物在第4天被安乐死。所有小鼠方案都符合美国联邦政府和乔治亚大学机构动物护理和使用委员会(iacuc)概述的非人类动物伦理治疗指南。
[0243]
用钙荧光白rc(blankophor bbh sv
‑
2560bayer,corp.)染色手工制备的小鼠肺切片中的真菌几丁质。通过将5mg溶解在218μldmso中并在4℃下避光储存来制备25mm储备液。在通过将储备液1:1,000稀释到pbs 5%dmso中至终浓度为25um的钙荧光白而制备的溶液中染色组织。
[0244]
通过集落形成单位(cfu)和实时定量pcr(qpcr)估计真菌负荷。
[0245]
将从存活至第4天的动物切除的肺称重,并切碎成数百个直径约1mm 3
的碎片,然后将这些碎片混合,这样可以使用切碎的组织的子集对整个肺进行准确取样。cfu:将25mg肺组织在200μl pbs中匀浆化,并通过在ypd(酵母马铃薯右旋糖)琼脂平板上与无菌玻璃珠粒一起摇动来铺展,该琼脂平板含有100μg/ml的卡那霉素和氨苄西林中的每一种。在37℃下孵育16小时后,对微量真菌集落进行计数,并在evos成像系统上以4x放大倍数拍摄一些菌落(图25b、25c)。根据整个平板相对于每个显微镜视野的面积和每个肺的重量,校正图25a和图26a中报告的cfu数量。qpcr:使用qiagen的25a和图26a中报告的cfu数量。qpcr:使用qiagen的血样和组织试剂盒(#69504,hilden,germany),从每个肺的25mg平行样本中提取dna。根据生产商的说明,将组织与180ul缓冲液atl和20ul蛋白酶k混合。此时,修改方案以便通过添加玻璃珠粒并用珠粒磨器(retsch mm300 laboratory mill)在室温下以中等速度振动样本10分钟来破碎真菌细胞壁。匀浆是透明液体,表明肺细胞和真菌细胞都完全溶解在atl缓冲液中,但它含有一些漂浮的脂质。将该材料经qiagen shredder自旋柱(货号79654)过滤以去除漂浮的脂质。此时,我们返回生产商的dna制备方案,从推荐的56℃孵育10分钟开始。通常从25mg肺组织中获得10μg dna。与先前描述的途径
65
相似,定量实时pcr(qpcr)被用于估计100ng肺dna样本中烟
曲霉(a.fumigatus)rrna重复序列的量。针对烟曲霉(a.fumigatus)的rdna基因中的基因间间隔区(igs)设计了几种新引物对。产生最低循环阈值(ct)和单个解离峰的最佳引物对具有以下序列(af18srrna2s正向引物5
′‑
ggatcgggcggtgtttctatga和af18srrna2a反向引物5
′‑
ttctttaagtttcagccttgcgaccat)。当检查未感染的肺组织时,即使经过45个pcr循环后,该引物对也未产生可检测的产物。真菌rdna igs的相对数量(rq)是使用dct方法(see livak等人,“analysis of relative gene expression data using real
‑
time quantitative pcr and the 2(
‑
delta delta c(t))method,”methods 25,402
‑
408(2001)),通过将所有ct值针对由感染肺样本对照确定的最低ct值作归一化来确定。
[0246]
结果
[0247]
如本文所述,开发了一种使用c型凝集素受体dectin
‑
1和dectin
‑
2的抗真菌药物的新递送系统,以分别使负载药物的脂质体靶向葡聚糖和甘露聚糖,即大多数真菌病原体的细胞壁和胞外多糖基质中发现的多糖。与非靶向的样脂质体药物相比,在一些情况下,dectin
‑
2包被的amb脂质体将90%抑制和杀灭烟曲霉(a.fumigatus)的脂质体amb体外有效剂量降低了约10至90倍。在本文中,使用免疫抑制介导的小鼠肺曲霉病的类固醇和白细胞减少模型来研究该新型治疗剂的体内功效。这些实验是以接近文献中报告的由递送的amb最低抑菌浓度0.2mg amb/kg小鼠体重的浓度进行的。对于这两种模型,在本文中证明了,dectin
‑
2靶向的负载amb的脂质体使肺部真菌负荷与递送相同的低amb浓度的非靶向样脂质体(amb
‑
ll)相比减少了超过一到两个数量级。通过显著降低抗真菌药物的有效剂量,靶向泛抗真菌脂质体有能力创造新的临床范式,以更安全地治疗多种多样的真菌疾病,无论它们是侵袭性感染还是局部感染。
[0248]
如本文所述,两种c型凝集素受体dectin
‑
1(dec1)和dectin
‑
2(dec2)的碳水化合物识别结构域分别被用于使负载抗真菌药物的脂质体靶向真菌β
‑
葡聚糖和α
‑
甘露聚糖。dectin靶向脂质体与烟曲霉(a.fumigatus)、白色念珠菌(c.albicans)和新型隐球菌(c.neoformans)的细胞壁和胞外多糖基质特异性结合。非靶向脂质体不会特异性结合,只能被动地将抗真菌剂递送至真菌细胞。如本文所示,dec1包被和dec2包被的负载两性霉素b(amb)的脂质体dec1
‑
amb
‑
ll和dec2
‑
amb
‑
ll有效地结合所有发育烟曲霉(a.fumigatus)的阶段,包括未膨大和膨大的孢子、幼苗和菌丝。它们抑制和/或杀灭烟曲霉(a.fumigatus)的效率比样脂质体amb
‑
ll高约10倍至约90倍,并将药物的体外有效剂量降低一个数量级。本文研究了这种真菌细胞靶向技术在曲霉病小鼠模型中是否体内有效。图23中的图表(右侧)例示了dectin包被的负载抗真菌药物的脂质体与真菌细胞及其胞外多糖基质结合,因此将药物特异性地靶向至真菌细胞附近。这也使抗真菌剂集中在远离哺乳动物细胞表面的地方。相比之下,非靶向药物被动地递送抗真菌剂至所有细胞(图23,左侧)。
[0249]
烟曲霉(a.fumigatus)是曲霉病的主要致病菌,曲霉病是四种最危及生命的真菌病之一。在全球范围内,估计有大约300,000例曲霉病急性病例。仅在2017年,美国曲霉菌病的治疗费用为每名患者58,000至105,000美元,每年美国曲霉菌感染的医疗费用总计15亿美元。曲霉病占治疗所有真菌感染费用的17%。烟曲霉(a.fumigatus)是常见的土壤生物,但也存在于家庭和工作场所。大多数人每天吸入数千个孢子而没有受到感染。罹患危及生命的侵袭性真菌感染(诸如曲霉病)的风险最大的患者通常具有减弱的免疫系统和/或患有
各种肺部疾病,这增加了真菌感染的机会。在免疫受损的患者中,曲霉病是继念珠菌病之后的第二大常见真菌感染。此外,易受各种机会性真菌感染的免疫受损个体的数量因使用免疫抑制剂的癌症、干细胞和器官移植患者升高而正在增加。
[0250]
曲霉病患者用抗真菌药物诸如两性霉素b(amb)、卡泊芬净和各种唑类药物进行治疗。几乎所有的抗真菌剂都是疏水性的,它们的低水溶性给药物递送带来了问题。具有被嵌入到脂质双层膜中的amb的脂质体药物制剂,诸如或等效物,诸如amb
‑
ll,能更有效地渗透到各种器官,穿透真菌细胞壁,并在比经洗涤剂增溶的amb(例如,amb
‑
doc)更高的amb剂量下显示降低的肾毒性和更小的输注毒性。及其等效物amb
‑
ll往往用于杀灭驻留于生物膜中的烟曲霉(a.fumigatus)。dectin靶向技术可以提高在生物膜中的抗真菌活性,因为dectin与帮助形成生物膜的胞外多糖基质结合。然而,所有抗真菌剂,无论是否包装在脂质体中,都因缺乏足够的杀真菌作用、耐药真菌种类增加以及对细胞和器官的宿主毒性而存在严重的局限性。例如,脂质体amb会导致肾毒性(参见,例如,allen u.“antifungal agents for the treatment of systemic fungal infections in children”paediatrics&child health 15,603
‑
608(2010);以及dupont b.“overview of the lipid formulations of amphotericin b”j antimicrob chemother 49 suppl 1,31
‑
36(2002)。由于两性霉素b的副作用,临床医生经常使用昵称“加倍可怕(amphoterrible)”来描述两性霉素b。即使采用药物治疗,曲霉菌病患者一年生存期的病死率通常在10%至仅90%的范围内,具体取决于患者的潜在状况。对于大约10%的患者,侵袭性曲霉病会发展为脑曲霉病,脑曲霉病是一种死亡率高达99%的脑部感染。
[0251]
使用本文所述的基于dectin的技术的目标之一是证明dectin
‑
2包被的脂质体药物在免疫抑制介导的曲霉病小鼠模型中抑制和杀灭体内烟曲霉(a.fumigatus)的功效增加(desoubeaux等人,“animal models of aspergillosis”comp med 68,109
‑
123(2018))。在以接近非靶向样amb
‑
ll递送的amb最低抑制浓度的浓度工作时,发现相对于递送相同amb浓度的amb
‑
ll而言,dec2
‑
amb
‑
ll显著降低了肺部真菌负荷。
[0252]
不同的出版物表明,需要使用在1
×
104至1
×
107烟曲霉(a.fumigatus)菌株cea10分生孢子范围内的鼻内接种量来诱导免疫受到抑制的cd1远交瑞士小鼠的致命曲霉病感染(参见desoubeaux等人;以及herbst等人,“a new and clinically relevant murine model of solid
‑
organ transplant aspergillosis”dis model mech 6,643
‑
651(2013))。当递送至小鼠肺部时,cea10与更常用的烟曲霉(a.fumigatus)af293菌株相比发芽更快,并显示出更强的毒力。在体内模型中,作为或样amb
‑
ll递送的amb的被报告的最低抑菌浓度(mic)在从0.06至1.0mg/kg小鼠体重的广泛范围内,并且似乎因烟曲霉(a.fumigatus)菌株而大幅变化。通过在amb
‑
ll的低amb剂量下工作,我们试图证明体内靶向真菌细胞的dectin包被的脂质体相对于体内非靶向脂质体具有最大程度地改进的表现。在接近amb
‑
ll的体外mic的浓度下工作时,培养物中烟曲霉(a.fumigatus)生长和存活力的治疗(先前证明为1或2个数量级)提高了dectin靶向脂质体的体外性能。本文所述的实验表明,样amb
‑
ll递送的0.2mg amb/kg足以测量到与受感染的对照小鼠相比,肺中烟曲霉(a.fumigatus)水平的轻微下降。
[0253]
健康小鼠与健康人一样,对曲霉属某些种的感染具有天然抵抗力。因此,小鼠必须
免疫受到抑制才会罹患急性曲霉病。使用了两种不同的免疫抑制小鼠模型,即类固醇模型和白细胞减少模型,它们的差异在于被抑制的免疫细胞功能的质量和免疫抑制的严重程度。图22示出了用于检查dectin
‑
2包被的抗真菌脂质体在肺曲霉病小鼠模型中的功效的两种方案和时间线。简而言之,在类固醇模型中,cd1小鼠是用合成类固醇曲安西龙或可的松进行免疫抑制(图22a),而在白细胞减少模型(图22b)中,小鼠是用抗代谢物环磷酰胺和类固醇进行免疫抑制。在免疫抑制介导的曲霉病的类固醇和白细胞减少模型中,免疫受到抑制的小鼠在第0天(d0)分别接受2
×
106和100,000个烟曲霉(a.fumigatus)分生孢子菌株cea10的口咽接种。到第2天(感染后第2天),在检查的小鼠肺的几乎每个肺叶中都观察到至少一个直径约1mm或更大的大感染中心。这些感染中心由短菌丝簇组成(图24)。在真菌接种后的第1天(图22),将感染的小鼠随机分成三组。一个处理组接受递送0.2mg amb/kg小鼠体重的dectin
‑
2包被的dec2
‑
amb
‑
ll,第二个脂质体处理组接受也递送0.2mg amb/kg的非靶向amb
‑
ll;对照模拟品处理组仅接受脂质体稀释缓冲液。所有分生孢子和脂质体都通过如先前所述用于肺疾病模型的口咽递送方法(参见de vooght等人,“oropharyngeal aspiration:an alternative route for challenging in a mouse model of chemical
‑
induced asthma,”toxicology 259,84
‑
89(2009))施用给小鼠。
[0254]
与非靶向amb
‑
ll相比,dec2
‑
amb
‑
ll治疗显著降低了真菌负荷。
[0255]
检查了抗真菌脂质体治疗对肺部真菌负荷的影响。首先,采用免疫抑制介导的曲霉病的类固醇模型(图22a)。在第1天和第3天将cd1瑞士小鼠用曲安西龙进行免疫抑制。在第0天,它们感染了2
×
106个烟曲霉(a.fumigatus)分生孢子。在第1天用dec2
‑
amb
‑
ll或者用递送0.2mg amb/kg小鼠体重的样amb
‑
ll脂质体或者用用于稀释脂质体的对照缓冲液治疗小鼠。在第1天和第4天之间,每个处理组中的一些动物已经死亡,但每个处理组中至少有三只动物存活到第4天。在第4天,从每组中随机选择三只存活的动物进行安乐死,收集它们的肺并称重。通过两种方法检查肺部的真菌负荷(图25)。将经匀浆化的肺组织平板接种在丰富生长培养基上,16小时后,估计每个肺的集落形成单位(cfu)的平均数量,如图25a所示。用dec2
‑
amb
‑
ll处理的小鼠显示出的能够形成集落的活真菌细胞数量比经amb
‑
ll处理的小鼠低12.5倍,而比经脂质体稀释缓冲液处理的对照小鼠低20.5倍。用于生成这些数据的真菌细胞微集落场的图像实例,参见图25b(经amb
‑
ll处理的小鼠)和图25c(经dec2
‑
amb
‑
ll处理的小鼠)。为了对由dec2
‑
amb
‑
ll引起的真菌负荷减少进行独立估计,使用定量聚合酶链反应qpcr来测量来自三个处理组中每一组的相同三只小鼠的经匀浆化的肺组织的平行样本上烟曲霉(a.fumigatus)核糖体rdna基因拷贝的相对数量(图25d)。根据rdna的相对数量估计值,用dec2
‑
amb
‑
ll治疗使每肺的真菌负荷比用样amb
‑
ll处理的小鼠的水平低22倍,而比对照小鼠的水平低40倍。在这两个真菌负荷实验中递送的amb浓度均为0.2mg/kg,处于所报告的对曲霉病小鼠模型中烟曲霉(a.fumigatus)真菌负荷的mic的范围内。观察到与对照动物相比,amb
‑
ll的真菌负荷下降了约40%,表明浓度处于非靶向药物mic的高端。
[0256]
其次,采用了免疫抑制介导的曲霉病的白细胞减少小鼠模型。该模型使小鼠甚至更容易感染曲霉病,并且需要较低剂量的分生孢子来建立急性感染(图22b)。在第
‑
3天和第
‑
1天用曲安西龙对cd1瑞士小鼠进行免疫抑制。在第0天,使它们感染5
×
105个烟曲霉
(a.fumigatus)分生孢子。在第1天用递送0.2mg amb/kg小鼠体重的靶向dec2
‑
amb
‑
ll或非靶向样amb
‑
ll脂质体处理小鼠或者用用于稀释脂质体的对照缓冲液处理小鼠。在4天(d4),除一只动物外,全部三个处理组中所有的动物都还活着。一个经对照缓冲液处理的动物在第4天上午死亡,剩余的对照动物和一个经amb
‑
ll处理的小鼠表现出理毛行为减少。amb
‑
ll和dec2
‑
amb
‑
ll处理组中的所有剩余动物看起来都相对健康。随机选择来自每个处理组的三只存活动物,将它们安乐死,收集它们的肺并通过刚刚描述的两种方法估计真菌负荷(图26)。用dec2
‑
amb
‑
ll处理的小鼠表现出的每肺集落形成单位(cfu)平均数比经amb
‑
ll处理的小鼠低100倍(图26a)。显然,使抗真菌脂质体靶向真菌细胞显著提高了包装在脂质体中的amb的功效。经amb
‑
ll处理的小鼠的cfu平均数较低,但与经对照缓冲液处理的动物在统计学上无法区分。amb
‑
ll处理组的高真菌负荷是因一只已表现出理毛行为减少的小鼠具有非常高的真菌滴度这一事实造成的。相比之下,一只经dec2
‑
amb
‑
ll处理的小鼠没有可检测到的真菌集落,导致该处理组的平均cfu非常低。作为对真菌负荷的独立估计,还检查了从来自相同小鼠的经匀浆化的肺组织的平行样本制备的dna上烟曲霉(a.fumigatus)rdna基因拷贝的相对数量(图26b)。用dec2
‑
amb
‑
ll处理使每肺的真菌负荷比用样amb
‑
ll处理的小鼠的水平低600倍。显然,利用dectin
‑
2使脂质体amb靶向真菌细胞可显著提高药物性能,以减少肺部真菌负荷。
[0257]
如本文所述,通过在使脂质体药物靶向真菌细胞及其胞外多糖基质中的甘露聚糖的泛真菌递送系统中使用dectin
‑
2的c型凝集素碳水化合物识别结构域,开发了脂质体抗体靶向的替代方案(图23)。有一种担忧是,由于两个主要原因,抗体作为使脂质体靶向真菌病原体的途径可能不如dection包被的脂质体那么有效。首先,高亲和力单克隆抗体可能由于特异性过大而仅与葡聚糖、甘露聚糖、木糖葡聚糖交联物和甘露糖蛋白的特定子集反应,因此仅靶向真菌物种的小子集或者甚至是同一物种的真菌细胞的子集。dectin靶向脂质体对多种多样的侵袭性真菌病原体的泛抗真菌活性应该使它们更值得广泛且昂贵的临床开发。其次,开发了用于以生产相同摩尔浓度单克隆抗体的成本的百分之几的成本在大肠杆菌(e.coli)中生产dectin
‑
1和dectin
‑
2、dec1和dec2的功能性碳水化合物识别结构域的不太繁琐的方法。再次,低试剂成本应鼓励用于治疗急性侵袭性真菌病和表面真菌感染的改进的抗真菌剂的临床开发。
[0258]
本文提供的数据表明,相对于递送相同的低amb剂量的非靶向样amb
‑
ll,dec2
‑
amb
‑
ll具有显著增强的抗真菌活性。使用两种不同的免疫抑制介导的小鼠曲霉病模型,dec2
‑
amb
‑
ll使肺部真菌负荷相对于等效物amb
‑
ll减少了一到两个数量级。预期用dec2
‑
amb
‑
ll治疗将在本文所述小鼠模型中提供改善的小鼠存活率。真菌负荷的显著减少应该可以推导出改善的患者长期生存率。
[0259]
同样重要的是,使用仅递送0.2mg amb/kg小鼠体重(该浓度接近所报告的在曲霉病小鼠模型中脂质体amb在体内对抗烟曲霉(a.fumigatus)的mic的下限)的dec2
‑
amb
‑
ll显示了显著效应。在使用这两种模型进行的本文所述的研究中,递送0.2mg amb/kg的amb
‑
ll对真菌负荷有温和的影响或没有影响。检查或amb
‑
ll在曲霉病肺部模型中的功效的研究通常需要5至20mg amb/kg的浓度才能对肺部真菌负荷产生最大的抗真菌作用并显著提高小鼠的存活率。此外,往往需要在感染后几天内提供多剂amb
‑
ll,以确保显著
降低真菌负荷和小鼠存活率。但在本文中我们证明了,由靶向dec2
‑
amb
‑
ll递送的低剂量amb使真菌负荷显著减少,比目前文献中大多数关于在小鼠模型中产生的减少真菌负荷的报告更显著。通过显著降低amb的有效剂量和减少控制曲霉病所需的剂量数量,dectin
‑
2靶向脂质体amb药物,诸如dec2
‑
amb
‑
ll,应该能以不会导致人器官毒性的浓度有效控制曲霉属感染。
[0260]
进一步地,本文报告的有关使用仅递送0.2mg amb/kg的dectin
‑
2靶向脂质体并获得真菌负荷数量级减少的数据表明,使负载抗真菌剂的脂质体直接靶向真菌细胞可能比靶向邻近的肺细胞和组织更有效。负载抗真菌药物的脂质体与真菌细胞的紧密邻近很可能会增加递送至这些细胞的药物局部浓度。本文提供的体外数据表明,dectin
‑
2主要靶向烟曲霉(a.fumigatus)胞外多糖基质中的α
‑
甘露聚糖,而不是其细胞壁的甘露聚糖含量。我们尚未证实,dectin
‑
2主要靶向小鼠模型的体内烟曲霉(a.fumigatus)胞外多糖基质。
[0261]
有几种不同的曲霉病小鼠模型可以模拟最易感曲霉病的各种患者群体所遭受的不同类型的免疫抑制。在本文中,使用免疫抑制的类固醇和白细胞减少模型来介导真菌感染(图22)。在类固醇模型中,将小鼠用合成的糖皮质激素曲安西龙进行预处理,糖皮质激素曲安西龙通过抑制一些t细胞和b细胞活性来破坏适应性免疫反应,从而增加烟曲霉(a.fumigatus)菌丝向肺组织中的增殖。类固醇免疫抑制小鼠模型有效地模拟了接受实体器官移植的免疫受到抑制的患者和一些接受干细胞移植的患者的免疫缺陷状态,这些患者特别容易感染曲霉病(参见desoubeaux等人)。然而,该模型的缺点是中性粒细胞过度募集到受影响的组织,尤其是肺部,从而导致剧烈的炎症反应。由于免疫系统仅部分失效,因此需要相对较大的真菌接种量,至少2
×
106个分生孢子,以确保100%的小鼠罹患急性曲霉病。大多数用在500,000至106个分生孢子范围内的较小接种量处理的小鼠在未进行抗真菌处理的情况下存活,而在另一个极端下,几乎所有用5
×
106处理的小鼠到第3天时死亡。在类固醇模型中,一些接种小鼠早在第1天和第2天死亡,这向我们表明,在脂质体治疗有时间生效之前,小鼠可能已经死于炎症和感染。尽管如此,与递送低剂量amb的非靶向amb
‑
ll相比,靶向真菌细胞的dec2
‑
amb
‑
ll减少了肺部真菌负荷。
[0262]
在下一组实验中,采用白细胞减少小鼠模型(图22b),其中小鼠用环磷酰胺和曲安西龙进行预处理。环磷酰胺是dna合成抑制剂和抗代谢药,可诱导包括b细胞和记忆t细胞在内的各种白细胞群的凋亡和快速减少。这被添加到类固醇曲安西龙的免疫抑制作用中。该模型消除了大部分先天性和免疫反应,并且对真菌细胞增殖的限制比类固醇模型要少得多。严重的白细胞减少症类似于造血干细胞移植接受者的免疫受到抑制的状态。由于这些小鼠会出现严重的白细胞减少症,因此可以使用较小的真菌接种量,从而减缓最初的疾病进展。希望减缓感染的初始进展将使amb
‑
ll或dec2
‑
amb
‑
ll的治疗有时间减少真菌负荷,并在未来的实验中提高小鼠的存活率。
[0263]
这两种模型都产生了统计学上令人信服的证据,即,靶向真菌细胞的脂质体相对于递送低剂量amb的非靶向脂质体减少了肺部的真菌负荷。如图26所示的在低剂量amb下肺部真菌负荷的这种显著减少无疑将能推导出小鼠存活率的提高和动物毒性的降低。
[0264]
如本文所示,dec2
‑
amb
‑
ll与体外生长的烟曲霉(a.fumigatus)、白色念珠菌(c.albicans)和新型隐球菌(c.neoformans)的胞外多糖基质中的甘露聚糖有效结合,并提高了杀灭所有三种真菌物种的功效。上述实验表明,dectin
‑
2靶向脂质体介导的抗真菌药
物递送显著提高了抑制和杀灭两种不同的免疫抑制介导的曲霉病小鼠模型体内烟曲霉(a.fumigatus)的功效。考虑到显示出对多种真菌病原体的有效活性的体外数据以及本文提供的关于烟曲霉(a.fumigatus)的体内小鼠数据,抗真菌药物的dectin
‑
2靶向应具有对抗包括念珠菌病和隐球菌病在内的许多其他侵袭性真菌疾病,以及可能对抗皮肤真菌病(例如由红色毛癣菌引起的足藓感染和指甲甲真菌病)的泛抗真菌应用。
[0265]
实施例4
[0266]
诊断法
[0267]
细胞培养
[0268]
使烟曲霉(a.fumigatus)菌株cea10(atcc mya1163)在预包被有聚
‑
l
‑
赖氨酸的24孔聚苯乙烯微量滴定板中的vogel最低培养基(vmm) 1%葡萄糖
77
或rpmi 1640培养基 1%葡萄糖(无红色指示剂)(thermofisher sku
‑
11835
‑
030,waltham,ma)中在37℃下生长12至16小时。将细胞用pbs(150mm nacl、10mm磷酸盐,ph 7.4)洗涤,用4%甲醛固定45分钟并用pbs洗涤3次,然后将该细胞与dec2
‑
bifc试剂脂质体一起孵育。
[0269]
venus融合蛋白
[0270]
密码子优化的大肠杆菌(e.coli)表达构建体dec2
‑
vyn和dec2
‑
vc的序列,连同它们产生的蛋白质序列和一些预测的蛋白质特性示于图27中。长度为15个氨基酸的富含glyser的柔性间隔区将dec2序列与venus片段分开。短至2个氨基酸的间隔区通常用于bifc构建体,诸如pet
‑
bifc。应当理解,在这些构建体中可以使用约2、3、4、5、6、7、8、9、10、11、12、13、14、15个氨基酸或更多的氨基酸的间隔区。在这个实例中,相对较长的间隔区被用来阻止融合蛋白的互补vyn和vc部分在脂质体上被放置成彼此邻近时自发缔合。这两种基因序列均由genscript合成并被亚克隆到pet
‑
45b表达载体中。
在于37℃
下进行6小时的iptg诱导后,经修饰的蛋白质在大肠杆菌的bl21菌株中表达。如之前针对小鼠sdectin
‑
1所述的那样,将两者都从细胞中提取出来,在镍亲和柱上纯化并储存在变性缓冲液#1(ph=8.0,6m guhcl(fisher bioreagents bp178)、0.1mna2hpo4/nah2po4、10mm三乙醇胺、100mm nacl、5mm 2
‑
巯基乙醇、0.1%triton
‑
x100)中。
[0271]
dec2
‑
bifc试剂构建
[0272]
用1m ph 10三乙醇胺将5μg/μl的在这种相同guhcl缓冲液(新添加有5mm 2
‑
巯基乙醇)中的两种蛋白质样本调节至ph 8.3,并使其与4摩尔过量的dspe
‑
peg
‑
3400
‑
nhs(1,2
‑
二硬脂酰
‑
sn
‑
甘油
‑3‑
磷酸乙醇胺(dspe)缀合的聚乙二醇(peg)(来自nanosoft polymers,1544
‑
3400)的反应性琥珀酰亚胺酯nhs部分在23℃下反应1小时。peg
‑
dspe部分的peg部分使这些疏水性蛋白稍微更容易溶解,而dspe是允许插入到脂质体膜中的脂质。通过bio
‑
gel p
‑
6丙烯酰胺树脂(bio
‑
rad#150
‑
0740)在复性和储存缓冲液rn#5(0.1m nah2po4、10mm三乙醇胺,ph 8.0,1m l
‑
精氨酸、100mm nacl、5mm edta、5mm 2
‑
巯基乙醇)中执行分子排阻色谱以去除未掺入的dspe
‑
peg和guhcl
69
。将这两种修饰的蛋白质以大约5μg/μl的浓度储存在rn#5中,直到用于活性测定或掺入到脂质体中。在通过透析去除guhcl后,使用sds
‑
page检查蛋白质纯度水平。
[0273]
dec2
‑
vyn和dec2
‑
vc向脂质体中的远程加载
[0274]
从来自formumax sci.inc.的无菌聚乙二醇化脂质体(dspc:chol:mpeg2000
‑
dspe,formumax#f10203a)开始,通过加入相对于用于制备dec2
‑
bifc试剂(表2)的100%脂
质体脂质而言的0.5摩尔百分比的仍在rn#5缓冲液中的dspe
‑
peg修饰的dec2
‑
vyn和dec2
‑
vc中的每一种来在60℃下在pbs中孵育30min,如此制备小批次脂质体。表2给出了以dec2
‑
vyn和dec2
‑
vc的摩尔数表示的本文讨论的dec2
‑
bifc试剂脂质体的化学组成,其中脂质体脂质的总量代表100摩尔百分比,并对该化学组成与最近描述的负载两性霉素b的dec2
‑
amb
‑
ll进行了比较。将dec2
‑
bifc试剂脂质体稀释到脂质体稀释缓冲液#2(ldb2、20mm hepes、10mm三乙醇胺、150mm nacl、10mm cacl2、1mmβ
‑
巯基乙醇(bme),5%bsa ph 8.0)中,然后用于荧光测定。
[0275]
表2
[0276][0277]
dec2
‑
bifc试剂与可溶性多糖和细胞的结合
[0278]
无细胞测定法。在pbs中按10mg/ml制备来自酿酒酵母(saccharomyces cerevisiae)的可溶性甘露聚糖(sigma,货号#084k3789)、来自掌状海带(laminaria digitate)的海带多糖(sigma
‑
aldrich,货号#l
‑
9634)、蔗糖(sigma)以及从乳杆菌属某些种纯化并且被尺寸分级为40,000分子量的葡聚糖
‑
t40(pharmacosmos,货号#551000409007)的多糖储备液。然后将它们以1:10稀释到含有dec2
‑
bifc试剂脂质体的ldb2缓冲液中,然后在室温下孵育。
[0279]
基于细胞的测定法。将福尔马林固定的真菌细胞与脂质体一起在23℃下在脂质体稀释缓冲液ldb2(20mm hepes、10mm三乙醇胺、150mm nacl、10mm cacl2、1mmβ
‑
巯基乙醇(bme)、5%bsa,ph 8.0)中孵育,其中bme为新鲜添加的。将dec2
‑
bifc脂质体稀释到ldb2缓冲液中,然后与细胞一起孵育,使dec2蛋白质组分的浓度w/v分别介于2μg/100μl、1.0μg/100μl或0.5μg/100μl之间。孵育在23℃或4℃下进行。采用gfp滤光片在olympus ix70倒置
显微镜上以20x拍摄用venus绿色荧光dec2
‑
bifc试剂脂质体染色的真菌集落图像,在明场中拍摄平行图像。在adobe photoshop中通过从明场图像中减去绿色和蓝色通道数据并从绿色荧光通道中加回绿色通道数据来制备合并图像。
[0280]
结果
[0281]
dectin
‑
2在淋巴细胞膜中作为单体漂移,但必须形成细胞外c型凝集素受体结构域的二聚体,以结合真菌细胞壁、胞外多糖基质、生物膜和释放到组织和血液中的多糖片段中的α
‑
甘露聚糖。二聚化给免疫系统发出真菌感染信号。我们利用dectin
‑
2的二聚化特性来设计基于双分子荧光互补(bifc)的无细胞泛真菌检测系统。使dectin
‑
2(dec2)的碳水化合物识别结构域与绿色荧光蛋白venus的两个互补片段,vyn和vc.dec2
‑
vyn融合。将dec2
‑
venus融合蛋白用脂质载体dspe
‑
peg进行修饰,并作为单体在脂质体膜中漂浮在一起以制备dec2
‑
bifc试剂。dec2
‑
bifc试剂在结合烟曲霉(a.fumigatus)时产生真菌细胞特异性绿色荧光信号。没有产生无细胞特异性的荧光信号。该试剂在与可溶性多糖结合时产生甘露聚糖特异性信号。未来的工作将探索在与白色念珠菌(candida albicans)和新型隐球菌(cryptococcus neoformans)结合后产生的信号。dec2
‑
bifc技术具有作为危及生命的侵袭性真菌感染的简单、快速、单步诊断法的潜力。
[0282]
侵袭性真菌感染常被误诊为非真菌相关疾病。抗真菌药物疗法的延迟显著增加了患者的死亡风险。以下是将曲霉病、念珠菌病和隐球菌病误诊为非真菌病的实例。曲霉病和结核病(tb)的许多症状是相同的,导致最初误诊并且施用抗菌药物治疗tb而不是用抗真菌药物治疗。类似地,侵袭性念珠菌病经常被误诊为细菌感染,并且用广谱抗菌药物治疗这些患者。此外,用抗细菌药物治疗患者会促进念珠菌属某些种过度生长,并增加患者获得侵袭性念珠菌病急性病例的机会。隐球菌脑膜炎经常被误诊为脑癌,因此不会立即用抗真菌药物治疗患者。本文提供了一种用于鉴别真菌感染并将它们与具有相似症状的非真菌疾病相区分的可靠、快速、灵敏的泛真菌诊断法。
[0283]
目前诊断真菌感染的方法在可靠性、快速性和敏感性方面存在严重的可鉴别问题。诊断念珠菌病的最常用技术是能够检测低至1个细胞每毫升的体外培养法。然而,细胞培养技术,甚至pcr未能在50%的患有该疾病的患者中检测到念珠菌病,因为在感染后不久,白色念珠菌(candida albicans)酵母细胞可以隐藏在宿主组织中,从而逃避基于真菌细胞的血清检测。由于曲霉属和念珠菌属物种将真菌特异性多糖释放到血清、痰、支气管肺泡灌洗液和尿液中,因此已经设计了真菌多糖的测定,特别是甘露聚糖和半乳甘露聚糖以及β
‑
葡聚糖的免疫测定法。然而,目前用于急性曲霉病的甘露聚糖和半乳甘露聚糖测定(例如,platelia aspergillus,bio
‑
rad)和β
‑
葡聚糖泛真菌测定(例如,fungitel,beacon diagnostics)主要是多步免疫elisa夹心测定法。这些商业化测定法不是严格可靠的,要么未能检测到感染,要么对很大一部分患者产生假阳性。此外,这些测定法不容易适应即时检测(point
‑
of
‑
care)设施,进一步延迟了诊断。隐球菌脑膜炎是使用基于抗体的elisa和隐球菌抗原的侧向流测定(lateral flow assay)以及真菌dna的pcr测定来诊断的,但这些测定在应用于患者脑脊髓液样本时也有类似的局限性。同样,早期检测将会挽救许多侵袭性真菌病患者的生命。显然,迫切需要更可靠、简单、快速、单步、即时检测的真菌诊断法,以减少对侵袭性真菌病患者的错误处置。
[0284]
dectin
‑
2(clec6a基因)是c型lectin结构域,其含有在小鼠和人中一些类别的淋
巴细胞表面上表达的跨膜受体。dectin
‑
2在与真菌α
‑
甘露聚糖和甘露糖蛋白结合时形成二聚体或多聚体,从而向先天性和适应性免疫系统发出真菌感染的信号。虽然细胞膜受体与高分子量多形真菌多糖相互作用的亲和常数很难确定,但从各出版物获取的sdectin
‑2‑
蛋白质融合体对模型甘露聚糖的亲和力估计值表明,它对含甘露聚糖的多糖只有适度的亲和力。然而,野生型与clec4n
‑
/
‑
缺失小鼠的反应比较表明,dectin
‑
2信号传导对1ng/ml的甘露聚糖聚合物浓度的反应最大,这意味着亲和常数kd在亚纳摩尔范围内。
[0285]
如本文所示,sdectin
‑
2(dec2)包被的脂质体与烟曲霉(a.fumigatus)、白色念珠菌(c.albicans)和新型隐球菌(c.neoformans)的胞外多糖基质非常快速、有效且相对不可逆地结合。因此,开发了利用dec2的新泛真菌诊断法。念珠菌属、隐球菌属和曲霉属物种属于三种进化上不同的真菌类别,分别是酵母纲(saccharomycetes)(子囊菌门)、银耳纲(tremellomycetes)(担子菌门)和散囊菌纲(eurotiomycetes)(子囊菌门)。据估计,它们在0.8至13亿年前的真菌王国进化过程中相对较早地与共同祖先分离。因为dec2包被的脂质体与所有三种菌属的细胞外基质特异性结合,这表明在大多数病原真菌的细胞外基质中发现的甘露聚糖在结构上足够保守从而能够被基于dec2的诊断法识别。
[0286]
本文中利用了dec2在与含甘露聚糖的多糖结合后形成二聚体的性质,以便通过双分子荧光互补(bifc)产生荧光信号。制备与荧光蛋白venus的互补片段融合的dec2,并将其在相同的脂质体中组合在一起。所得的dec2
‑
bifc试剂脂质体在与烟曲霉(a.fumigatus)细胞结合后会产生清晰的荧光信号。该技术可以为多种多样的病原真菌、它们的胞外多糖基质、生物膜和释放的多糖提供简单、快速的单步即时测定。
[0287]
图28中示出了使用dec2的基于脂质体的单步检测系统的模型。它基于dec2的二聚化,因为它与真菌甘露聚糖和bifc结合。产生了两种单独的sdectin
‑
2蛋白。第一种蛋白质与绿色荧光蛋白venus vyn的n末端部分(venus残基1
‑
155、突变体t154m)融合以制备dec2
‑
vyn(图28a、28b)。vyn venus突变片段的优点是与c末端venus片段的自发缔合减少,在与c末端片段缔合后产生的信号强,而相对于其他荧光蛋白的n末端片段或未修饰的vn片段而言优点很少。产生的第二种蛋白质是与venus vc的c末端部分(venus残基155
‑
238)融合从而制备dec2
‑
vc的sdectin
‑
2(图28a、28b)。该vc片段之前已在成功的bifc应用中与vyn合作。图27中示出了两种编码dna和两种bifc融合蛋白dec2
‑
vyn和dec2
‑
vc的确切序列。这两种编码序列都在大肠杆菌(e.coli)中表达,将蛋白质提取到盐酸胍变性缓冲液中,并使用本文所述的方法通过亲和层析进行纯化。虽然仍然变性,但融合蛋白的互补对各自通过游离赖氨酸残基与胺反应性双疏脂质试剂dspe
‑
peg
‑
nhs偶联。然后,将dspe
‑
peg修饰的蛋白质经由它们偶联的dspe部分以相等的摩尔比插入到相同的100纳米直径脂质体的膜中,以制备dec2
‑
bifc试剂脂质体(图28a)。插入到每个100nm脂质体中的dec2融合蛋白的数量限制于相对于脂质体脂质分子数量的1摩尔百分比的dec2或1,500个dec2分子。换言之,每个脂质体中存在大约750个dec2
‑
vyn和750个dec2
‑
vc蛋白分子。这里的目标是使用足够低的蛋白质浓度以避免两个venus片段自发缔合(这种缔合可能会在不存在任何甘露聚糖结合的情况下产生高背景荧光信号),但使用足够高浓度的dec2来促进有效结合、二聚化以及试剂遇到真菌甘露聚糖时产生强信号。有人提出,在脂质体膜中漂浮在一起的dec2
‑
vyn和dec2
‑
vc单体几乎只在它们结合真菌细胞甘露聚糖时才会形成二聚体或多聚体。还有人提出,dec2二聚化将促进venus荧光蛋白
‑
vyn和
‑
vc的互补部分的组装,以产生bifc信号。
[0288]
执行无细胞微量滴定板测定,以证明dec2
‑
bifc试剂脂质体产生了对多糖α
‑
甘露聚糖具有特异性的荧光信号。递送1μg/100μlvenus的dec2
‑
bifc试剂脂质体
[0289]
将培养基中的l dec2蛋白与含有α
‑
甘露聚糖的可溶性酵母甘露聚糖、与海带多糖(可溶性β
‑
葡聚糖)、与蔗糖(葡萄糖和果糖的二糖)以及与葡聚糖(α
‑
葡聚糖)一起孵育。在荧光微量滴定板读数器中,查看gfp通道的信号。图29显示,相对于含有其他多糖的孔,在含有甘露聚糖的孔中观察到统计学上明显更高的水平的venus绿色荧光(p=<0.01)。这些结果支持信号是相对甘露聚糖特异性的这一观点,正如对dectin
‑
2所预期的。两小时后观察到最大信号,并随着孵育时间的延长而降低。
[0290]
执行了基于细胞的显微镜测定,以证明dec2
‑
bifc试剂脂质体产生真菌细胞特异性荧光信号。使烟曲霉(a.fumigatus)分生孢子在聚
‑
l
‑
赖氨酸包被的微量滴定板上发芽,过夜生长至早期菌丝阶段,从而形成直径为300至500微米的小集落。将细胞固定在福尔马林中并洗涤到脂质体稀释缓冲液中。将细胞与向孵育培养基中递送2μg/100μl、1μg/100μl和0.5μg/100μl的总dec2蛋白浓度的dec2
‑
bifc试剂一起孵育。
[0291]
在倒置荧光显微镜的gfp通道(ex515/em528)中,很容易看到bifc信号。图30示出了dec2
‑
bifc试剂(1μg dec2蛋白/100μl)与烟曲霉(a.fumigatus)集落结合时形成的荧光信号。bifc荧光信号仅在与真菌细胞缔合时观察到,因此是真菌细胞特异性的(图30b、d、f、h)。一百多个真菌细胞集落中每一个都检查到结合的试剂。每个真菌集落的中心处信号最强。请注意,每个集落的中心都有最厚的细胞层和最老的细胞,并且有最多的时间产生富含甘露聚糖的粘附性胞外多糖,以帮助它们粘附到赖氨酸包被的平板表面。我们之前已经证明,dec2特别擅长结合由烟曲霉(a.fumigatus)产生的胞外多糖,并且胞外多糖沉积应增强与dec2
‑
bifc试剂的结合。存在沿其大部分长度被染色的长菌丝。在孵育两到三天后观察到最大信号。有一些真菌菌丝不显示荧光信号并且可能不结合试剂的实例(白色箭头,图30b)。使用递送2μg dec2/100μl的脂质体也检测到相似强度的真菌细胞特异性信号,在0.5μg dec2/100μl时,信号稍弱。在细胞之间(图30b、d、f、h)或在缺乏真菌细胞的对照孔中没有观察到荧光。与缓冲液一起孵育的对照细胞没有显示绿色荧光信号(图30i和j)。
[0292]
venus作为bifc构建体中荧光蛋白的选择
[0293]
由于两个原因,venus被选为构建bifc泛真菌诊断法的最佳荧光蛋白。首先,选择venus是因为在几个得到最佳研究的黄色、绿色和天蓝色荧光蛋白中,当未被伴侣蛋白强迫在一起时,venus的两个片段(c末端片段vc和突变的n末端片段vyn)具有最低的报告的自发缔合率。自发缔合会产生独立于融合蛋白与所需靶标的结合的荧光信号,并且是bifc技术自其诞生以来的一大难题。低自发缔合率被认为对于开发不产生假阳性(即在不存在真菌细胞甘露聚糖的情况下的荧光信号)的诊断试剂是必不可少的。其次,venus是最亮的荧光蛋白之一,并且源自非常亮的黄色的荧光蛋白(yfp)家族中的蛋白质。量子产率(每个被吸收光子释放的光子)在0.5至0.6的范围内(例如,理论100%产率的50%至60%)
70
。这与罗丹明的量子产率0.7相似,罗丹明是荧光细胞化学常用的最具荧光性的低分子量荧光分子之一。
[0294]
bifc试剂脂质体
[0295]
出于多种原因,设计了含有dec2
‑
vyn和dec2
‑
vc的dec2
‑
bifc试剂脂质体,而不是使用呈溶液形式的两种可溶性蛋白质dec2
‑
vyn和dec2
‑
vc。首先,dec2单体相对不溶,并且
通过将它们储存在6m盐酸胍缓冲液中,可以使它们保持可溶状态。其次,dspe
‑
peg部分的添加稳定了并部分增溶了dec2单体,并使得它们能够被储存在温和变性的含1m精氨酸的缓冲液中一段时间。这种修饰对于将修饰的蛋白质插入到脂质体中是必不可少的。然而,该两种蛋白质的这种修饰形式有可能用作不含脂质体的诊断试剂。第三,使用脂质体时,dec2单体在较容易操纵的稳定试剂中的局部浓度能被控制。这里的目标还是要达到高dec2浓度,但不能太高以致于导致连接的venus片段自发缔合和明显可测量的荧光背景。例如,在这些初始实验中,发现1,500个dec2单体(即750个dec2
‑
vyn加750个dec2
‑
vc单体)是有效的。甚至将脂质体在4℃下储存2个月后,也没有观察到自发信号形成,应理解单体的浓度可以增加或减少。第四,并且也许是最重要的,试剂脂质体的亲合力特性应该使荧光信号强度呈指数增加。亲合力的潜力是通过在每个脂质体上具有多个甘露聚糖结合位点而产生的,诸如针对五聚igm抗体所观察到的。一旦一个dec2
‑
vyn和dec2
‑
vc对与富含甘露聚糖的真菌多糖中的甘露聚糖结合,当结合扩散到脂质体上相邻的dec2分子与富含甘露聚糖的多糖中的相邻甘露聚糖部分的结合时,信号应放大。一旦开始发展,亲和力也应稳定该信号。可溶性bifc诊断试剂无法提供这些优势。本文列出的数据是用dec2
‑
bifc试剂脂质体进行的。结果显然是阳性的,毫无疑问,由该试剂产生的荧光信号是真菌细胞特异性的。
[0296]
在其他实例中,可以使用更短的间隔区来强迫vyn和vc片段更有效地在一起,以更快地形成信号。本文所述的系统可以用于在dec2
‑
bifc试剂与真菌样本组合后约30分钟、40分钟或60分钟内获得最大信号。
[0297]
dectin
‑
2的碳水化合物识别结构域与α
‑
甘露聚糖的紧密结合伴随着体内白细胞表面上的二聚体形成。这种由细胞外碳水化合物识别结构域dec2形成二聚体的特性已成功地被用于开发体外工作的真菌细胞特异性诊断法。使dec2与荧光蛋白venus的两个互补片段融合,以制成dec2
‑
bifc试剂脂质体。只要通过将dec2
‑
bifc试剂脂质体与烟曲霉(a.fumigatus)细胞一起孵育,它们就会产生清晰的真菌细胞特异性绿色荧光信号。因此,这种技术可以用作简单、单步、即时检测的真菌诊断法,其将减少对侵袭性真菌病患者的错误处置,并可能用作浅表真菌病的诊断法。
[0298]
此外,有数十种可形成多聚体的c型凝集素结合蛋白,因为它们结合多种多样的靶真菌多糖,因此这些多糖可用于扩展这种诊断技术(参见,例如,hardison等人,“c
‑
type lectin receptors orchestrate antifungal immunity,”nat immunol.13,817
‑
822(2012))。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。