id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少80%序列同一性的序列或其活性片段。任选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少85%序列同一性的序列或其活性片段。任选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少90%序列同一性的序列或其活性片段。
8.任选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少95%序列同一性的序列或其活性片段。任选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少99%序列同一性的序列或其活性片段。任选地,外肽酶具有根据malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一的序列或其活性片段。
9.任选地,蛋白水解酶混合物具有第二外肽酶。优选地,第二外肽酶为氨肽酶。任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少70%序列同一性的序列或其氨肽酶活性片段。任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少80%序列同一性的序列或其氨肽酶活性片段。任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少85%序列同一性的序列或其氨肽酶活性片段。任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少90%序列同一性的序列或其氨肽酶活性片段。
10.任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少95%序列同一性的序列或其氨肽酶活性片段。任选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少99%序列同一性的序列或其氨肽酶活性片段。任选地,氨肽酶具有根据seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一的序列或其氨肽酶活性片段。任选地,氨肽酶具有根据seq id no:10的序列或其氨肽酶活性片段。
11.任选地,蛋白水解酶混合物还具有内肽酶。优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少70%序列同一性的序列或其内肽酶活性片段。任选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少80%序列同一性的序列或其内肽酶活性片段。任选地,内肽
酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少85%序列同一性的序列或其内肽酶活性片段。任选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少90%序列同一性的序列或其内肽酶活性片段。任选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少95%序列同一性的序列或其内肽酶活性片段。任选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少99%序列同一性的序列或其内肽酶活性片段。任选地,内肽酶具有根据seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一的序列或其内肽酶活性片段。
12.任选地,蛋白质材料为植物来源蛋白质、动物来源蛋白质、鱼类来源蛋白质、昆虫来源蛋白质或微生物来源蛋白质。任选地,蛋白质材料包含谷蛋白、大豆蛋白、乳蛋白、卵蛋白、乳清蛋白、酪蛋白、肉、血红蛋白或肌球蛋白。
13.任选地,蛋白水解酶混合物至少具有如上所述对在次末n
‑
末端具有脯氨酸的肽具有特异性的外肽酶、第二外肽酶和内肽酶。任选地,这些酶用于同时处理蛋白质材料。任选地,在不同时间使用这些酶。
14.任选地,用于生产蛋白质水解产物的方法是用于生产具有升高水平的谷氨酸的水解产物。任选地,蛋白水解酶混合物具有谷氨酰胺酶。任选地,谷氨酰胺酶具有与seq id no:29具有至少70%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有与seq id no:29具有至少80%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有与seq id no:29具有至少85%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有与seq id no:29具有至少90%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有与seq id no:29具有至少95%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有与seq id no:29具有至少99%序列同一性的序列或其谷氨酰胺酶活性片段。任选地,谷氨酰胺酶具有根据seq id no:29的序列或其谷氨酰胺酶活性片段。根据本发明的这一方面,蛋白质材料任选地为谷蛋白。
15.任选地,用于生产蛋白质水解产物的方法是用于生产具有升高水平的脯氨酸的水解产物。
16.在本发明的另一方面,提供了一种蛋白质水解产物,所述蛋白质水解产物是根据以上披露的任一方法生产的。
17.在本发明的另一方面,提供了一种食物产品,所述食物产品具有如上所述的蛋白质水解产物。
18.生物学序列简述
19.seq id no:1列出了全长malpro11的蛋白质序列。
20.seq id no:2列出了全长mcipro4的蛋白质序列。
21.seq id no:3列出了全长tcipro1的蛋白质序列。
22.seq id no:4列出了全长fvepro4的蛋白质序列。
23.seq id no:5列出了全长ssppro2的蛋白质序列。
24.seq id no:6为pgxt
‑
malpro11、pgxt
‑
mcipro4和pgxt
‑
tcipro1中额外的5’dna片段的dna序列。
25.seq id no:7列出了预测的前导区截短的fvepro4的蛋白质序列。
26.seq id no:8列出了预测的前导区截短的ssppro2的蛋白质序列。
27.seq id no:9列出了五肽底物的蛋白质序列。
28.seq id no:10列出了预测的前导区截短的acpepn2 tri035的蛋白质序列。
29.seq id no:11列出了预测的前导区截短的氨肽酶tr031的蛋白质序列。
30.seq id no:12列出了预测的前导区截短的氨肽酶tr032的蛋白质序列。
31.seq id no:13列出了预测的前导区截短的氨肽酶tr033的蛋白质序列。
32.seq id no:14列出了预测的前导区截短的氨肽酶tr034的蛋白质序列。
33.seq id no:15列出了预测的前导区截短的氨肽酶tr036的蛋白质序列。
34.seq id no:16列出了预测的前导区截短的氨肽酶tr037的蛋白质序列。
35.seq id no:17列出了预测的前导区截短的氨肽酶tr038的蛋白质序列。
36.seq id no:18列出了成熟枯草杆菌蛋白酶a的蛋白质序列。
37.seq id no:19列出了成熟枯草杆菌蛋白酶bpn’的蛋白质序列。
38.seq id no:20列出了成熟迟缓枯草杆菌蛋白酶(subtilisin lentus)的蛋白质序列。
39.seq id no:21列出了成熟嗜热菌蛋白酶的蛋白质序列。
40.seq id no:22列出了成熟芽孢杆菌溶素(bacillolysin)的蛋白质序列。
41.seq id no:23列出了成熟木霉属胃蛋白酶(trichodermapepsin)的蛋白质序列。
42.seq id no:23列出了成熟木霉属胃蛋白酶(trichodermapepsin)的蛋白质序列。
43.seq id no:24列出了成熟菠萝蛋白酶(bromealin)的蛋白质序列。
44.seq id no:25列出了成熟曲霉属胃蛋白酶(aspergillopepsin)的蛋白质序列。
45.seq id no:26列出了成熟胰蛋白酶1的蛋白质序列。
46.seq id no:27列出了成熟胰凝乳蛋白酶a的蛋白质序列。
47.seq id no:28列出了预测的前导区截短的氨肽酶tr063的蛋白质序列。
48.seq id no:29列出了全长谷氨酰胺酶的蛋白质序列。
附图说明
49.图3a描绘了phe
‑
pro上纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的剂量反应曲线。
50.图3b描绘了ser
‑
pro上纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的剂量反应曲线。
51.图4描绘了纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的ph曲线。
52.图5描绘了纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的温度曲线。
53.图6描绘了纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的热稳定性测试。
54.图7描绘了纯化的malpro11、mcipro4、tcipro1、fvepro4和ssppro2的gln
‑
pro
‑
gln
‑
gln
‑
pro水解分析。
55.图8示出了不同剂量的ssppro2对于由谷蛋白预水解产物在与acpepn2和谷氨酰胺酶一起孵育19h后形成游离谷氨酸的作用。参考:含有谷蛋白预水解产物 谷氨酰胺酶。acpepn2含有谷蛋白预水解产物 谷氨酰胺酶 acpepn2。最后两个样品含有与acpepn2相同的内容,但还额外地含有131μg/ml或392μg/ml的预水解产物。
56.图9与图8相同,但为孵育26h后。
57.图10示出了不同x
‑
proap对于谷氨酸产率的作用。在50℃下,与预水解产物、谷氨酰胺酶和所提及的酶一起孵育24h。在所有情况下,x
‑
proap的剂量均为312μg/ml的预水解产物。
58.图11示出了aox
‑
proap和hx
‑
proap对于谷氨酸产率的作用。在50℃下,与预水解产物、谷氨酰胺酶和所提及的酶一起孵育42h。在所有情况下,x
‑
proap的剂量均为15μg/ml的预水解产物。
59.图12示出了水解产物的重叠色谱图。实线:预水解产物与谷氨酰胺酶和acpepn2一起孵育26h。虚线:预水解产物与谷氨酰胺酶、acpepn2和ssppro2一起孵育26h。图中指示出氨基酸(aa’s)主要洗脱的时间间隔以及dp2至dp5主要洗脱的间隔。
60.图13示出了水解产物的重叠色谱图。实线:预水解产物与谷氨酰胺酶和acpepn2一起孵育26h。虚线:预水解产物与谷氨酰胺酶、acpepn2和hx
‑
proap一起孵育26h。图中指示出氨基酸(aa’s)主要洗脱的时间间隔以及dp2至dp5主要洗脱的间隔
具体实施方式
61.除非另有说明,否则本发明教导的实践将使用在本领域技术范围内的分子生物学(包括重组技术)、微生物学、细胞生物学和生物化学的常规技术。此类技术在以下文献中得到充分解释,例如,molecular cloning: a laboratory manual[分子克隆:实验室手册],第二版(sambrook等人,1989);oligonucleotide synthesis[寡核苷酸合成](m.j.gait编辑,1984);current protocols in molecular biology[分子生物学当前方案](f.m.ausubel等人,编辑,1994);pcr:the polymerase chain reaction[pcr:聚合酶链式反应](mullis等人,编辑,1994);gene transfer and expression:a laboratory manual[基因转移和表达:实验室手册](kriegler,1990),和the alcohol textbook[醇教科书](ingledew等人,编辑,第五版,2009),以及essentials of carbohydrate chemistry and biochemistry[碳水化合物化学与生物化学基础](lindhorste,2007)。
[0062]
除非在本文中另有定义,本文所用的全部技术术语和科学术语具有与本发明教导所属领域的普通技术人员通常所理解的相同含义。singleton等人,dictionary of microbiology and molecular biology[微生物学和分子生物学词典],第二版,john wiley and sons[约翰威利父子公司],纽约(1994),以及hale和markham,the harper collins dictionary of biology[哈珀柯林斯生物学词典],harper perennial[哈珀永久出版社],纽约(1991)为技术人员提供了本发明中所使用的许多术语的通用词典。与本文所述的那些类似或等效的任何方法和材料可用于本发明教导的实践或测试中。
[0063]
本文提供的数值范围包括限定范围的数值在内。
[0064]
定义
[0065]
关于多肽,术语“野生型”、“亲本”或“参考”是指在一个或多个氨基酸位置处不包含人为取代、插入或缺失的天然存在的多肽。类似地,关于多核苷酸,术语“野生型”、“亲本”或“参考”是指不包含人为核苷变化的天然存在的多核苷酸。然而,注意编码野生型、亲本、或参考多肽的多核苷酸不限于天然存在的多核苷酸,并且涵盖编码野生型、亲本、或参考多肽的任何多核苷酸。
[0066]
对野生型多肽的提及应理解为包括多肽的成熟形式。“成熟”多肽或其变体是其中不存在信号序列的多肽或变体,例如,在多肽表达期间或之后从未成熟形式的多肽切割。
[0067]
关于多肽,术语“变体”是指与指定的野生型、亲本或参考多肽不同的多肽,因为它包括一种或多种天然存在的或人为的氨基酸取代、插入或缺失。类似地,关于多核苷酸,术语“变体”是指在核苷酸序列中与指定的野生型、亲本或参考多核苷酸不同的多核苷酸。野生型、亲本或参考多肽或多核苷酸的特性将从上下文中显而易见。
[0068]
当关于主题细胞、核酸、蛋白质或载体使用时,术语“重组”指示受试者已经从其天然状态被修饰。因此,例如,重组细胞表达在天然(非重组)形式的细胞中未发现的基因,或以不同于自然界发现的水平或在不同于自然界发现的条件下表达天然基因。重组核酸与天然序列不同在于一个或多个核苷酸,和/或有效地连接到异源序列,例如表达载体中的异源启动子。重组蛋白质与天然序列的不同可在于一个或多个氨基酸,和/或与异源序列融合。包含编码蛋白酶的核酸的载体是重组载体。
[0069]
术语“回收的”、“分离的”和“单独的”是指从如天然存在的与其天然相关的至少一种其他材料或组分中除去的化合物、蛋白质(多肽)、细胞、核酸、氨基酸、或者其他指定材料或组分。其“分离的”多肽包括但不限于含有在异源宿主细胞中表达的分泌多肽的培养液。
[0070]
术语“纯化的”是指处于相对纯状态的材料(例如,分离的多肽或多核苷酸),例如,至少约90%纯、至少约95%纯、至少约98%纯或甚至至少约99%纯。
[0071]
术语“富集的”是指处于约50%纯、至少约60%纯、至少约70%纯、或甚至至少约70%纯的材料(例如,分离的多肽或多核苷酸)。
[0072]
关于酶的“ph范围”是指在其下酶显示催化活性的ph值的范围。
[0073]
关于酶的术语“ph稳定”和“ph稳定性”涉及在一个宽范围内的ph值下,酶保持预定时间段(例如,15min.、30min.、1小时)的活性的能力。
[0074]
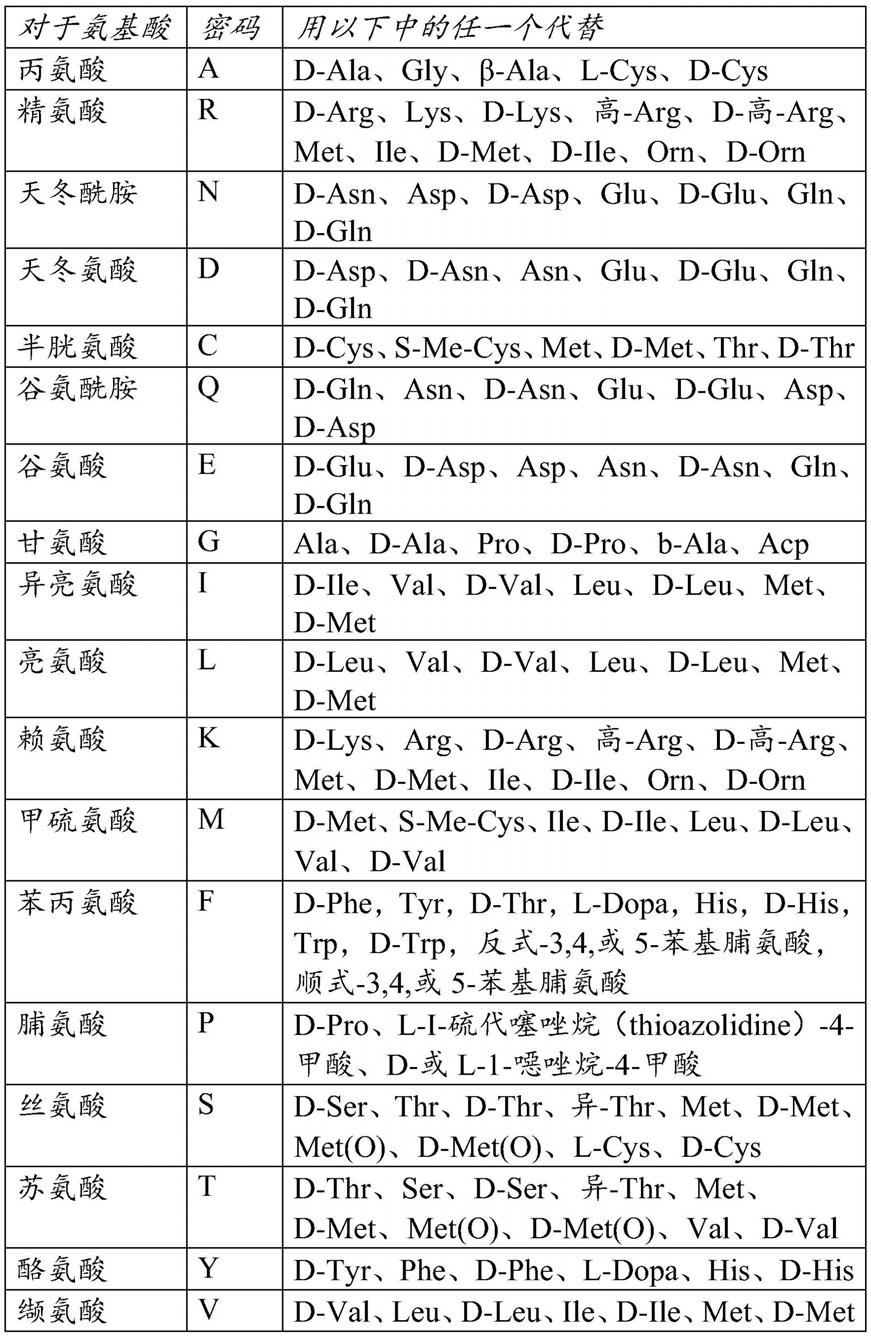
术语“氨基酸序列”与术语“多肽”“蛋白质”和“肽”同义,并且可互换地使用。当此类氨基酸序列显示出活性时,它们可以被称为“酶”。使用针对氨基酸残基的常规单字母或三字母密码,采用标准氨基端
‑
至
‑
羧基端取向(即n
→
c)表示氨基酸序列。
[0075]
术语“核酸”涵盖能够编码多肽的dna、rna、异源双链体、以及合成分子。核酸可以是单链的或双链的,并且可以是化学修饰。术语“核酸”和“多核苷酸”可互换地使用。由于遗传密码是简并的,可以使用多于一个密码子来编码具体的氨基酸,并且本发明的组合物和方法涵盖编码具体氨基酸序列的核苷酸序列。除非另有说明,否则核酸序列以5
′‑
至
‑3′
取向呈现。
[0076]“杂交”是指如在印迹杂交技术和pcr技术期间发生的,一条核酸链与互补链形成双链体(即碱基对)的过程。严格杂交条件通过在以下条件下杂交来例证:65℃和0.1x ssc(其中1x ssc=0.15m nacl、0.015m柠檬酸三钠,ph 7.0)。杂交的双链核酸的特征在于熔融
温度(t
m
),其中一半杂交的核酸与互补链不配对。双链体内错配的核苷酸降低t
m
。非常严格杂交条件涉及68℃和0.1x ssc。
[0077]“合成”分子是通过体外化学或酶合成而不是通过生物产生的。
[0078]
关于细胞使用的术语“转化”、“稳定转化”和“转基因”意指细胞含有整合到其基因组中或作为通过多代维系的附加体的非天然(例如异源)核酸序列。
[0079]
在将核酸序列插入细胞的上下文中,术语“引入”意指本领域已知的“转染”、“转化”或“转导”。
[0080]“宿主菌株”或“宿主细胞”是已经引入了表达载体、噬菌体、病毒或其他dna构建体,包括编码目的多肽(例如,蛋白酶)的多核苷酸的生物。示例性宿主菌株是能够表达目的多肽的微生物细胞(例如,细菌、丝状真菌和酵母)。术语“宿主细胞”包括从细胞产生的原生质体。
[0081]
关于多核苷酸或蛋白质的术语“异源”是指不是天然存在于宿主细胞中的多核苷酸或蛋白质。
[0082]
关于多核苷酸或蛋白质的术语“内源”是指天然存在于宿主细胞中的多核苷酸或蛋白质。
[0083]
术语“表达”是指基于核酸序列产生多肽的过程。所述过程包括转录和翻译两者。
[0084]“选择性标记”或“可选择标记”是指能够在宿主中被表达以促进选择携带所述基因的宿主细胞的基因。可选择标记的实例包括但不限于在宿主细胞上赋予代谢优势(如营养优势)的抗微生物剂(例如,潮霉素、博来霉素或氯霉素)和/或基因。
[0085]“载体”是指设计用于将核酸引入一种或多种细胞类型的多核苷酸序列。载体包括克隆载体、表达载体、穿梭载体、质粒、噬菌体颗粒、盒等。
[0086]“表达载体”是指包含编码目的多肽的dna序列的dna构建体,所述编码序列与能够在适合的宿主中影响dna表达的适合控制序列有效地连接。此类控制序列可以包括影响转录的启动子,控制转录的任选的操纵子序列,编码mrna上适合的核糖体结合位点的序列,增强子以及控制转录和翻译终止的序列。
[0087]
术语“有效地连接”意指:指定组分处于允许它们以预期方式起作用的关系(包括但不限于并列)。例如,调控序列与编码序列有效地连接,使得编码序列的表达受调控序列的控制。
[0088]“信号序列”是与蛋白质的n
‑
末端部分附接的氨基酸序列,所述氨基酸序列有利于蛋白质在细胞外的分泌。细胞外的蛋白质的成熟形式缺乏在分泌过程中被切除的信号序列。
[0089]“生物学活性的”是指具有指定生物活性,例如酶活性的序列。
[0090]
术语“比活性”是指在特定条件下每单位时间通过酶或酶制剂可转化为产物的底物的摩尔数。比活性通常表示为单元(u)/mg蛋白质。
[0091]
如本文所使用的,“序列同一性百分比”意指当使用具有默认参数的clustal w算法比对时,特定序列具有与指定的参考序列中的氨基酸残基相同的至少一定百分比的氨基酸残基。参见thompson等人(1994)nucleic acids res.[核酸研究]22:4673
‑
4680。clustal w算法的默认参数是:
[0092]
空位开放罚分:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
10.0
[0093]
空位延伸罚分:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
0.05
[0094]
蛋白质权重矩阵:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀ
blosum系列
[0095]
dna权重矩阵:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
iub
[0096]
延迟发散序列%:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀ
40
[0097]
空位分隔距离:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ8[0098]
dna转换权重:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
0.50
[0099]
列表亲水残基:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
gpsndqekr
[0100]
使用负性矩阵:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
关
[0101]
切换特殊残基罚分:
ꢀꢀꢀꢀꢀꢀꢀ
开
[0102]
切换亲水罚分:
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
开
[0103]
切换结束空位分隔罚分
ꢀꢀꢀꢀ
关。
[0104]
与参考序列相比,缺失被视为不同一的残基。包括在任一末端发生的缺失。例如,相对于成熟多肽,具有成熟617残基多肽的c
‑
末端的五个氨基酸缺失的变体将具有99%序列同一性百分比(612/617相同的残基
×
100,四舍五入到最接近的整数)。此类变体将被与成熟多肽具有“至少99%序列同一性”的变体所涵盖。
[0105]“融合”多肽序列经由两个受试多肽序列之间的肽键连接,即有效地连接。
[0106]
术语“丝状真菌”是指所有丝状形式的真菌亚门(eumycotina),特别是子囊菌亚门(pezizomycotina)物种。
[0107]
术语“约”是指参考值的
±
5%。
[0108]
术语“肽酶”或“蛋白酶”是指水解多肽或寡肽中肽键的酶。如本文所使用的,术语肽酶或蛋白酶包括分配到ec 3.4亚类的酶。
[0109]
术语“外肽酶”或“外切蛋白酶”是指作用于水解多肽或寡肽末端(氨基或羧基)的肽键的肽酶。作用于多肽的氨基末端的外肽酶在本文中被称为氨肽酶。氨肽酶可以根据其特异性起到从氨基末端切割或释放单个氨基酸、二肽和三肽的作用。作用于羧基末端的外肽酶在本文中被称为羧肽酶。羧肽酶可以根据其特异性起到从羧基末端切割或释放单个氨基酸、二肽和三肽的作用。
[0110]
术语“内肽酶”或“内切蛋白酶”是指水解蛋白质或寡肽中的内部肽键的肽酶或蛋白酶。
[0111]“水解产物”是其中化合物被水分解的反应的产物。蛋白质的水解产物或“蛋白质水解产物”在用水将蛋白质键水解时出现。可通过加热或酶来增加蛋白质的水解。水解期间,蛋白质被分解成更小的蛋白质、多肽和游离氨基酸。
[0112]
其他定义如下文示出。
[0113]
额外的突变
[0114]
在一些实施例中,本发明的蛋白酶进一步包括可提供另外的性能或稳定性益处的一个或多个突变。示例性的性能益处包括但不限于:增加的热稳定性、增加的储存稳定性、增加的溶解度、改变的ph曲线、增加的比活性、经修饰的底物特异性、经修饰的底物结合、经修饰的ph依赖性活性、经修饰的ph依赖性稳定性、增加的氧化稳定性、和增加的表达。在一些情况下,性能益处是在相对较低的温度下实现的。在一些情况下,性能益处是在相对较高的温度下实现的。
[0115]
此外,本发明的蛋白酶可以包含任何数量的保守氨基酸取代。下表中列出了示例性保守氨基酸取代。
[0116]
表1保守氨基酸取代
[0117]
[0118]
读者将理解,一些前述保守突变可以通过遗传操作产生,而其他通过用遗传或其他方式将合成的氨基酸引入多肽中产生。
[0119]
本发明的蛋白酶可以是“前体”、“未成熟”或“全长”的,在这种情况下,它们包含信号序列;或“成熟”的,在这种情况下,它们缺乏信号序列。成熟形式的多肽通常是最有用的。除非另有说明,否则本文使用的氨基酸残基编号是指相应蛋白酶多肽的成熟形式。只要所得多肽保留蛋白酶活性,本发明的蛋白酶多肽也可被截短以去除n
‑
末端或c
‑
末端。此外,蛋白酶可以是来源于更长的氨基酸序列的活性片段。活性片段的特征在于保留全长酶的一些或全部活性,但具有来自n
‑
末端、来自c
‑
末端或其内部或组合的缺失。
[0120]
本发明的蛋白酶可以是“嵌合”或“杂合”多肽,因为其包括第一蛋白酶多肽的至少一部分和第二蛋白酶多肽的至少一部分。本发明的蛋白酶可进一步包含异源信号序列,即允许跟踪或纯化等的表位。示例性异源信号序列来自地衣芽孢杆菌(b.licheniformis)淀粉酶(lat)、枯草芽孢杆菌(b.subtilis)(amye或apre)、和链霉菌属(streptomyces)cela。
[0121]
变体蛋白酶的生产
[0122]
本发明的蛋白酶可以在宿主细胞中产生,例如通过分泌或细胞内表达。在将蛋白酶分泌到细胞培养基中后,可以获得包含蛋白酶的培养细胞材料(例如,全细胞培养液)。任选地,蛋白酶可以从宿主细胞中分离,或甚至从细胞培养液中分离,这取决于最终蛋白酶所希望的纯度。可以根据本领域熟知的方法克隆和表达编码蛋白酶的基因。合适的宿主细胞包括细菌、真菌(包括酵母和丝状真菌)和植物细胞(包括藻类)。特别有用的宿主细胞包括黑曲霉(aspergillus niger)、米曲霉(aspergillus oryzae)或里氏木霉(trichoderma reesei)。其他宿主细胞包括细菌细胞,例如枯草芽孢杆菌(bacillus subtilis)或地衣芽孢杆菌(b.licheniformis),以及链霉菌属(streptomyces)、大肠杆菌(e.coli)。
[0123]
宿主细胞还可以表达编码同源或异源蛋白酶(即,与宿主细胞不同物种的蛋白酶)或一种或多种其他酶的核酸。蛋白酶可以是变体蛋白酶。另外,宿主可以表达一种或多种辅酶、蛋白质、肽。
[0124]
载体
[0125]
可以构建包含编码蛋白酶的核酸的dna构建体以在宿主细胞中表达。由于遗传密码中熟知的简并性,编码相同氨基酸序列的变体多核苷酸可以用常规技术进行设计和制备。优化用于特定宿主细胞的密码子也是本领域熟知的。可以将编码蛋白酶的核酸掺入载体。可以使用熟知的转化技术(例如以下公开的那些)将载体转移至宿主细胞中。
[0126]
载体可以是可以转化到宿主细胞中并在宿主细胞内复制的任何载体。例如,包含编码蛋白酶的核酸的载体可以在作为繁殖和扩增载体的手段的细菌宿主细胞中转化并复制。载体也可以被转化到表达宿主中,使得编码核酸可以被表达为功能性蛋白酶。用作表达宿主的宿主细胞可以包括,例如,丝状真菌。美国真菌遗传学库存中心(fgsc)的菌株目录列出了适合在真菌宿主细胞中表达的载体。参见fgsc,菌株目录,密苏里大学,网址为www.fgsc.net(最新修改时间为2007年1月17日)。代表性的载体是pjg153,其是可在细菌宿主中复制的无启动子cre表达载体。参见harrison等人,(2011年6月)applied environ.microbiol[应用和环境微生物学]77:3916
‑
22。pjg153可以用常规技术进行修饰以包含并表达编码蛋白酶的核酸。
[0127]
可以将编码蛋白酶的核酸有效地连接到适合的启动子,其允许在宿主细胞中转
录。启动子可以是在选择的宿主细胞中显示转录活性的任何dna序列,并且可以来源于编码与宿主细胞同源抑或异源的蛋白质的基因。用于指导编码蛋白酶的dna序列的转录(尤其是在细菌宿主中)的示例性启动子为大肠杆菌lac操纵子的启动子、天蓝色链霉菌(streptomyces coelicolor)琼脂酶基因daga或cela启动子、地衣芽孢杆菌(bacillus licheniformis)α
‑
淀粉酶基因(amyl)的启动子、嗜热脂肪芽孢杆菌(bacillus stearothermophilus)生麦芽糖淀粉酶基因(amym)的启动子、解淀粉芽孢杆菌(bacillus amyloliquefaciens)α
‑
淀粉酶(amyq)的启动子、枯草芽孢杆菌xyla和xylb基因的启动子等。对于在真菌宿主中的转录,有用的启动子的实例是衍生自编码米曲霉(aspergillus oryzae)taka淀粉酶、米氏根瘤菌(rhizomucor miehei)天冬氨酸蛋白酶、黑曲霉(aspergillus niger)中性α
‑
淀粉酶、黑曲霉酸稳定性α
‑
淀粉酶、黑曲霉葡糖淀粉酶、米氏根瘤菌脂肪酶、米曲霉碱性蛋白酶、米曲霉三糖磷酸异构酶或构巢曲霉(a.nidulans)乙酰胺酶的基因的启动子。当编码蛋白酶的基因在细菌物种(如大肠杆菌)中表达时,可以从,例如,包括t7启动子和噬菌体λ启动子的噬菌体启动子中选择适合的启动子。用于在酵母物种中表达的适合的启动子的实例包括但不限于酿酒酵母(saccharomyces cerevisiae)的gal 1和gal 10启动子以及毕赤酵母(pichia pastoris)aox1或aox2启动子。cbh1是来自里氏木霉(t.reesei)的内源诱导型启动子。参见liu等人(2008)“improved heterologous gene expression in trichoderma reesei by cellobiohydrolase i gene(cbh1)promoter optimization[纤维二糖水解酶i基因(cbh1)启动子优化改善里氏木霉中的异源基因表达],”acta biochim.biophys.sin(shanghai)[生物化学与生物物理学报(上海)]40(2):158
‑
65。
[0128]
编码序列可以有效地连接到信号序列。编码信号序列的dna可以是与待表达的蛋白酶基因天然相关或来自不同属或物种的dna序列。可以将包含dna构建体或载体的信号序列和启动子序列引入真菌宿主细胞,并且可以来源于相同的来源。例如,信号序列是有效地连接到cbh1启动子的cbh1信号序列。
[0129]
表达载体还可以包含合适的转录终止子,以及在真核生物中,包含有效地连接到编码变体蛋白酶的dna序列的聚腺苷酸化序列。终止序列和聚腺苷酸化序列可以适当地来源于与启动子相同的来源。
[0130]
所述载体可以进一步包含使得所述载体能够在宿主细胞中复制的dna序列。此类序列的实例为质粒puc19、pacyc177、pub110、pe194、pamb1、和pij702的复制起点。
[0131]
载体还可以包含可选择标记,例如其产物弥补分离的宿主细胞中的缺陷的基因,例如来自枯草芽孢杆菌或地衣芽孢杆菌的dal基因,或者赋予抗生素抗性(例如氨苄青霉素、卡那霉素、氯霉素或四环素抗性)的基因。此外,载体可以包含曲霉属选择标记,例如amds、argb、niad和xxsc,引起潮霉素抗性的标记,或者选择可以通过共转化(如本领域已知的)实现。参见,例如,国际pct申请wo 91/17243。
[0132]
细胞内表达在某些方面可能是有利的,例如,当使用某些细菌或真菌作为宿主细胞产生大量蛋白酶用于随后的富集或纯化时。蛋白酶的细胞外分泌至培养基中还可用于制备包含分离的蛋白酶的培养的细胞材料。
[0133]
表达载体典型地包括克隆载体的组分,例如像允许载体在选择的宿主生物体中自主复制的元件和用于选择目的一个或多个表型可检测的标记。表达载体通常包含控制核苷
酸序列,例如启动子、操纵子、核糖体结合位点、翻译起始信号以及任选地抑制子基因或一个或多个激活子基因。另外,表达载体可包含编码氨基酸序列的序列,所述氨基酸序列能够将蛋白酶靶向宿主细胞细胞器(如过氧化物酶体)或靶向具体宿主细胞区室。这样的靶向序列包括但不限于序列skl。对于在控制序列的指导下进行表达,将蛋白酶的核酸序列以关于表达的适当方式与控制序列有效地连接。
[0134]
用于分别连接编码蛋白酶的dna构建体、启动子、终止子和其他元件,并将它们插入到含有复制所需信息的适合的载体中的程序是本领域技术人员熟知的(参见,例如,sambrook等人,molecular cloning:a laboratory manual[分子克隆:实验室手册],第2版,cold spring harbor[冷泉港实验室],1989,和第3版,2001)。
[0135]
宿主细胞的转化和培养
[0136]
包含dna构建体或表达载体的分离的细胞有利地用作重组产生蛋白酶的宿主细胞。通过将dna构建体(以一或多个拷贝)整合到宿主染色体中,可以方便地用编码酶的dna构建体转化细胞。通常认为这种整合是有利的,因为dna序列更可能在细胞中稳定保持。可以根据常规方法,例如,通过同源或异源重组,将dna构建体整合到宿主染色体中。可替代地,可以用如上所述的与不同类型的宿主细胞相关的表达载体转化细胞。
[0137]
适合的细菌宿主生物的实例是革兰氏阳性细菌物种,如芽孢杆菌属(bacillaceae),包括枯草芽孢杆菌(bacillus subtilis)、地衣芽孢杆菌(bacillus licheniformis)、迟缓芽孢杆菌(bacillus lentus)、短芽孢杆菌(bacillus brevis)、嗜热脂肪地芽孢杆菌(geobacillus stearothermophilus)(原嗜热脂肪芽孢杆菌(bacillus stearothermophilus)、嗜碱芽孢杆菌(bacillus alkalophilus)、解淀粉芽孢杆菌(bacillus amyloliquefaciens)、凝结芽孢杆菌(bacillus coagulans)、灿烂芽孢杆菌(bacillus lautus)、巨大芽孢杆菌(bacillus megaterium)和苏云金芽孢杆菌(bacillus thuringiensis);链霉菌属(streptomyces)物种,如鼠灰链霉菌(streptomyces murinus);乳酸细菌物种,包括乳球菌属物种(lactococcus sp.),如乳酸乳球菌(lactococcus lactis);乳杆菌属物种(lactobacillus sp.),包括罗伊氏乳杆菌(lactobacillus reuteri);明串珠菌属物种(leuconostoc sp.);片球菌属物种(pediococcus sp.);和链球菌属物种(streptococcus sp.)。可替代地,可以选择属于肠杆菌科(enterobacteriaceae)(包括大肠杆菌)或假单胞菌科(pseudomonadaceae)的革兰氏阴性细菌物种的菌株作为宿主生物。
[0138]
适合的酵母宿主生物可以选自生物技术相关的酵母物种,例如但不限于酵母物种,如毕赤酵母属物种(pichia sp.)、汉逊酵母属物种(hansenula sp.)或克鲁维酵母属(kluyveromyces)、耶氏酵母属(yarrowinia)、裂殖酵母属(schizosaccharomyces)物种或者酵母属(saccharomyces)物种,包括酿酒酵母(saccharomyces cerevisiae),或者属于裂殖酵母属的物种,例如,粟酒裂殖酵母(s.pombe)。甲基营养型酵母物种菌株毕赤酵母可以用作宿主生物。可替代地,宿主生物可以是汉逊酵母属物种。丝状真菌中适合的宿主生物包括曲霉属(aspergillus)的物种,例如,黑曲霉、米曲霉、塔宾曲霉(aspergillus tubigensis)、泡盛曲霉(aspergillus awamori)或构巢曲霉(aspergillus nidulans)。可替代地,镰孢菌属(fusarium)物种(例如,尖孢镰孢菌(fusarium oxysporum))或根瘤菌属(rhizomucor)物种(如米氏根瘤菌)的菌株可以用作宿主生物。其他适合的菌株包括嗜热菌
属(thermomyces)和毛霉菌属(mucor)物种。此外,木霉属物种(trichoderma sp.)可以用作宿主。转化曲霉属宿主细胞的适合的程序包括,例如,ep 238023中描述的程序。真菌宿主细胞表达的蛋白酶可以被糖基化,即,将包含糖基部分。糖基化模式可以与野生型蛋白酶中存在的相同或不同。糖基化的类型和/或程度可能改变酶和/或生化特性。
[0139]
从表达宿主缺失基因是有利的,其中可以通过转化的表达载体治愈基因缺陷。已知方法可用于获得具有一种或多种失活基因的真菌宿主细胞。可以通过完全或部分缺失,通过插入失活或通过使基因对其预期目的不起作用的任何其他方式使得所述基因被阻止表达功能性蛋白来完成基因灭活。已克隆的、来自木霉属物种或其他丝状真菌宿主的任何基因均可以缺失,例如,cbh1、cbh2、egl1和egl2基因。基因缺失可通过本领域已知方法通过将待失活的所希望的基因的形式插入质粒中来完成。
[0140]
将dna构建体或载体引入宿主细胞中包括以下技术,例如转化;电穿孔;核显微注射;转导;转染,例如脂质转染介导的和deae
‑
糊精介导的转染;与磷酸钙dna沉淀一起孵育;用dna包被的微粒高速轰击;以及原生质体融合。通用转化技术是本领域已知的。参见,例如sambrook等人(2001),同上。异源蛋白质在木霉属中的表达描述于例如美国专利号6,022,725。对于曲霉属菌株的转化,还参考了cao等人(2000)science[科学]9:991
‑
1001。可以用载体系统构建遗传稳定的转化体,由此编码蛋白酶的核酸被稳定整合到宿主细胞染色体中。然后通过已知技术选择并纯化转化体。
[0141]
用于转化的木霉属物种的制备例如可以涉及从真菌菌丝体制备原生质体。参见campbell等人(1989)curr.genet.[当代遗传学]16:53
‑
56。菌丝体可以从发芽的营养孢子获得。使用消化细胞壁的酶处理菌丝体,产生原生质体。通过悬浮培养基中渗透稳定剂的存在来保护原生质体。这些稳定剂包括山梨糖醇、甘露糖醇、氯化钾、硫酸镁等。通常这些稳定剂的浓度在0.8m与1.2m之间变化,例如山梨糖醇的1.2m溶液可以用于悬浮培养基中。
[0142]
取决于钙离子浓度,将dna摄取到宿主木霉属物种菌株中。一般情况下,在摄取溶液中使用在约10mm至50mm之间的cacl2。额外的合适的化合物包括缓冲系统,例如te缓冲液(10mm tris,ph 7.4;1mm edta)或10mm mops(ph 6.0)和聚乙二醇。据信聚乙二醇使细胞膜融合,从而允许培养基的内容物被递送至木霉属物种菌株的细胞质中。该融合时常留下整合到宿主染色体中的多个拷贝的质粒dna。
[0143]
通常,使用已经进行了渗透性处理的原生质体或细胞转化木霉属物种,通常以105至107/ml、特别是2x 106/ml的密度进行。可以将100μl体积的在合适的溶液(例如1.2m山梨糖醇和50mm cacl2)中的这些原生质体或细胞与所希望的dna混合。通常,向摄取溶液中添加高浓度的peg。可以将从0.1至1体积的25%peg 4000添加到原生质体悬浮液中;然而,向原生质体悬浮液中添加约0.25体积是有用的。也可以将添加剂例如二甲亚砜、肝素、亚精胺、氯化钾等添加到摄取溶液中以促进转化。类似的程序可用于其他真菌宿主细胞。参见,例如美国专利号6,022,725。
[0144]
表达
[0145]
产生蛋白酶的方法可以包括在有利于产生所述酶的条件下如上所述培养宿主细胞,并从细胞和/或培养基中回收所述酶。
[0146]
用于培养细胞的培养基可以是适合于所考虑的宿主细胞生长并获得蛋白酶表达的任何常规培养基。合适的培养基和培养基组分可从商业供应商获得或可以根据公开的配
方(例如,如在美国典型培养物保藏中心(american type culture collection)的目录中所述)来制备。
[0147]
从宿主细胞中分泌的酶可以用于全培养液制剂中。在本发明的方法中,可以使用本领域已知的任何培养方法来实现重组微生物的用过的全发酵液的制备,从而导致蛋白酶的表达。因此,发酵可以理解为包括在实验室或工业发酵罐中在合适的培养基和允许蛋白酶被表达或分离的条件下进行的摇瓶培养、小规模或大规模发酵(包括连续发酵、分批发酵、补料分批发酵或固态发酵)。术语“用过的全发酵液”在本文中定义为包括培养基、胞外蛋白质(例如,酶)和细胞生物质的发酵材料的未分级内容物。应当理解,术语“用过的全发酵液”还涵盖已经使用本领域熟知的方法裂解或透化的细胞生物质。
[0148]
从宿主细胞中分泌的酶可方便地通过熟知的程序从培养基中回收,所述程序包括通过离心或过滤从培养基中分离细胞,并借助于盐(例如硫酸铵)沉淀培养基的蛋白质组分,然后使用色谱程序,例如离子交换色谱法、亲和色谱法等。
[0149]
可以将在载体中编码蛋白酶的多核苷酸有效地连接到控制序列,所述控制序列能够通过宿主细胞提供编码序列的表达,即所述载体是表达载体。可以例如通过添加其他转录调控元件来修饰控制序列,以使由控制序列指导的转录水平对转录调节因子更有应答。控制序列尤其可以包含启动子。
[0150]
宿主细胞可以在允许表达蛋白酶的合适条件下培养。这些酶的表达可以是组成型的,使得它们能被连续产生,或诱导型的,需要刺激来启动表达。在诱导型表达的情况下,当需要时可通过例如向培养基中添加诱导物质,例如地塞米松或iptg或槐糖来启动蛋白质生产。多肽也可以在体外无细胞系统(例如tnt
tm
(普洛麦格公司(promega))兔网织红细胞系统)中重组生产。
[0151]
表达宿主也可以在有氧条件下在对于宿主来说适当的培养基中培养。可以提供摇动或搅动和通气的组合,其中生产在对所述宿主来说适当的温度下(例如从约25℃至约75℃(例如30℃至45℃),这取决于宿主的需要和所希望的蛋白酶的生产)发生。培养可以发生从约12至约100小时或更长时间(以及其间的任何小时值,例如从24至72小时)。典型地,培养液的ph为约4.0至约8.0,同样取决于宿主相对于蛋白酶的生产所需的培养条件。
[0152]
富集和纯化蛋白酶的方法
[0153]
发酵、分离和浓缩技术是本领域熟知的,并且可使用常规方法来制备含蛋白酶多肽的溶液。
[0154]
发酵后,获得发酵液,通过常规分离技术去除微生物细胞和各种悬浮固体(包括残余的粗发酵材料),以便获得蛋白酶溶液。通常使用过滤、离心、微滤、旋转真空鼓式过滤、超滤、离心然后超滤、提取或色谱法等。
[0155]
期望浓缩含蛋白酶多肽的溶液以优化回收。使用未浓缩的溶液需要增加孵育时间以收集富集或纯化的酶沉淀物。
[0156]
使用常规浓缩技术浓缩含酶溶液,直至获得所希望的酶水平。含酶溶液的浓缩可以通过在本文中所讨论的技术中的任一种来实现。富集和纯化的示例性方法包括但不限于旋转真空过滤和/或超滤。
[0157]
将酶溶液浓缩成浓缩的酶溶液,直至浓缩的含蛋白酶多肽的溶液的酶活性达到所希望的水平。
[0158]
富集或纯化的酶可以制成为液体(溶液,浆料)或固体(颗粒,粉末)的最终产品。
[0159]
本发明的优选实施例
[0160]
根据本发明的一方面,发现了一些氨肽酶停滞或仅缓慢消化在次末n
‑
末端位置具有脯氨酸的肽或蛋白质。特别地,发现了这些氨肽酶不消化具有n
‑
末端序列x
‑
pro
‑
gln
‑
gln
‑
pro
‑
de(其中x为任一氨基酸)的肽的蛋白质。在生产蛋白质水解产物中使用此类氨肽酶将产生具有低量的x氨基酸的水解产物,这是由于这种肽对消化具有抗性。
[0161]
谷氨酸单钠(msg)形式的谷氨酸是常用的风味增强剂。它负责咸味或鲜味。msg可通过酶水解蛋白质来产生。在这方面,谷蛋白在谷氨酰胺中含量很高并且可以是msg的来源(可以使用谷氨酰胺酶将谷氨酰胺转化为谷氨酸)。根据本发明的一方面,发现了谷蛋白含有显著量的序列x
‑
pro
‑
gln
‑
gln
‑
pro
‑
,其充分限制了可从谷蛋白中释放出的谷氨酰胺的量。
[0162]
根据本发明的一方面,提供了一种用于从蛋白质材料制备蛋白质水解产物的方法,其中在水性条件下将蛋白质材料与蛋白水解酶组合接触,所述蛋白水解酶组合具有对在次末n
‑
末端具有脯氨酸的肽具有特异性的外肽酶。在优选实施例中,外肽酶对具有五个氨基酸序列x
‑
pro
‑
gln
‑
gln
‑
pro
‑
作为n
‑
末端的肽具有特异性,其中x为氨基末端氨基酸并且可以是任一天然存在的氨基酸,pro为脯氨酸并且gln为谷氨酰胺。
[0163]
优选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少70%序列同一性的序列或其活性片段。更优选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少80%序列同一性的序列或其活性片段。仍更优选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少85%序列同一性的序列或其活性片段。在又更优选实施例中,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少90%序列同一性的序列或其活性片段。
[0164]
仍更优选地,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少95%序列同一性的序列或其活性片段。在仍更优选实施例中,外肽酶具有与malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一具有至少99%序列同一性的序列或其活性片段。在最优选实施例中,外肽酶具有根据malpro11(seq id no:1)、mcipro4(seq id no:2)、tcipro1(seq id no:3)、fvepro4(seq id no:4)、和ssppro2(seq id no:5)之一的序列或其活性片段。
[0165]
在本发明的优选实施例中,蛋白水解酶混合物具有第二外肽酶。优选地,第二外肽酶为氨肽酶。更优选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少70%序列同一性的序列或其氨肽酶活性片段。仍更优选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少80%序列同一性的序列或其氨肽酶活
性片段。又更优选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少85%序列同一性的序列或其氨肽酶活性片段。仍更优选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少90%序列同一性的序列或其氨肽酶活性片段。
[0166]
在仍更优选实施例中,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少95%序列同一性的序列或其氨肽酶活性片段。又更优选地,氨肽酶具有与seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一具有至少99%序列同一性的序列或其氨肽酶活性片段。仍更优选地,氨肽酶具有根据seq id no:10、seq id no:11、seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17和seq id no:28之一的序列或其氨肽酶活性片段。在最优选实施例中,氨肽酶具有根据seq id no:10的序列或其氨肽酶活性片段。
[0167]
在本发明的其他优选实施例中,蛋白水解酶混合物还具有内肽酶。优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少70%序列同一性的序列或其内肽酶活性片段。更优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少80%序列同一性的序列或其内肽酶活性片段。仍更优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少85%序列同一性的序列或其内肽酶活性片段。又更优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少90%序列同一性的序列或其内肽酶活性片段。在仍更优选实施例中,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少95%序列同一性的序列或其内肽酶活性片段。又更优选地,内肽酶具有与seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一具有至少99%序列同一性的序列或其内肽酶活性片段。在最优选实施例中,内肽酶具有根据seq id no:18、seq id no:19、seq id no:20、seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26和seq id no:27之一的序列或其内肽酶活性片段。
[0168]
在本发明的优选实施例中,蛋白质材料为植物来源蛋白质、动物来源蛋白质、鱼类来源蛋白质、昆虫来源蛋白质或微生物来源蛋白质。优选地,蛋白质材料包含谷蛋白、大豆蛋白、乳蛋白、卵蛋白、乳清蛋白、酪蛋白、肉、血红蛋白或肌球蛋白。
[0169]
在其他优选实施例中,蛋白水解酶混合物至少具有如上所述的对在次末n
‑
末端具
有脯氨酸的肽具有特异性的外肽酶、第二外肽酶和内肽酶。优选地,这些酶用于同时处理蛋白质材料。在其他优选实施例中,在不同时间使用这些酶。
[0170]
在本发明的优选实施例中,用于生产蛋白质水解产物的方法是用于生产具有升高水平的谷氨酸的水解产物。根据本发明的这一方面,蛋白水解酶混合物具有谷氨酰胺酶。优选地,谷氨酰胺酶具有与seq id no:29具有至少70%序列同一性的序列或其谷氨酰胺酶活性片段。更优选地,谷氨酰胺酶具有与seq id no:29具有至少80%序列同一性的序列或其谷氨酰胺酶活性片段。仍更优选地,谷氨酰胺酶具有与seq id no:29具有至少85%序列同一性的序列或其谷氨酰胺酶活性片段。在又更优选实施例中,谷氨酰胺酶具有与seq id no:29具有至少90%序列同一性的序列或其谷氨酰胺酶活性片段。仍更优选地,谷氨酰胺酶具有与seq id no:29具有至少95%序列同一性的序列或其谷氨酰胺酶活性片段。在又更优选实施例中,谷氨酰胺酶具有与seq id no:29具有至少99%序列同一性的序列或其谷氨酰胺酶活性片段。在最优选实施例中,谷氨酰胺酶具有根据seq id no:29的序列或其谷氨酰胺酶活性片段。
[0171]
根据本发明的这一方面,蛋白质材料为谷蛋白。
[0172]
在其他优选实施例中,用于生产蛋白质水解产物的方法是用于生产具有升高水平的脯氨酸的水解产物。
[0173]
在本发明的另一方面,提供了一种蛋白质水解产物,所述蛋白质水解产物是根据以上披露的任一方法生产的。
[0174]
在本发明的另一方面,提供了一种食物产品,所述食物产品具有如上所述的蛋白质水解产物。
[0175]
实例
[0176]
实例1真菌x
‑
pro蛋白酶的克隆
[0177]
选择两个真菌菌株,热白丝菌(melanocarpus albomyces)cbs177.67(gicc#2522192)和樟绒枝霉(malbrancheae cinamonea)cbs 343.55(gicc#2518670),作为可用于多种工业应用的酶的潜在来源。热白丝菌cbs177.67和樟绒枝霉cbs 343.55购自cbs
‑
knaw真菌生物多样性中心(cbs
‑
knaw fungal biodiversity centre)(uppsalalaan 8,3584ct乌特勒支市(utrecht),荷兰)。使用illumina的下一代测序技术对染色体dna进行测序,并且在注释后鉴定两种真菌x
‑
pro蛋白酶:从热白丝菌cbs177.67中鉴定malpro11,以及从樟绒枝霉cbs 343.55中鉴定mcipro4。malpro11和mcipro4的全长蛋白质序列分别示出于seq id no:1和seq id no:2。
[0178]
选择jgi数据库(https://genome.jgi.doe.gov/portal/)中列出的三个真菌菌株(桔绿木霉(trichoderma citrinoviride)tucim 6016、轮枝样镰孢菌(fusarium verticillioides)7600和壳多胞属物种(stagonospora sp.)src1lsm3a)作为可用于多种工业应用的酶的潜在来源。blast检索(altschul等人,j mol biol[分子生物学杂志],215:403
‑
410,1990)能够鉴定出三种蛋白酶:从桔绿木霉tucim 6016中鉴定tcipro1、从轮枝样镰孢菌7600中鉴定fvepro4、和从壳多胞属物种src1lsm3a中鉴定ssppro2。tcipro1(jgi菌株id:trici4,蛋白质id:1136694)、fvepro4(jgi菌株id:fusve2,蛋白质id:4472)、和ssppro2(jgi菌株id:stasp1,蛋白质id:303285)的全长蛋白质序列分别列出为seq id no:3、seq id no:4和seq id no:5。
sciences),#3641),并在恒温混匀仪(thermomixer)中在50℃和600rpm下孵育5min,随后添加10μl的稀释酶样品(或仅添加水作为空白对照)。在恒温混匀仪中在50℃和600rpm下孵育20min后,通过在95℃加热10min来终止蛋白酶反应。
[0186]
如通过茚三酮反应所检测的,从二肽(phe
‑
pro或ser
‑
pro)水解产生的游离pro用于示出蛋白水解活性。反应前,将茚三酮(西格玛公司,#151173)溶解于100%乙醇中至最终浓度为5%(w/v)。为了开始茚三酮反应,首先将40μl的1m醋酸钠(ph 2.8)与10μl的5%茚三酮溶液在96
‑
mtp pcr板(爱思进公司(axygen),pcr
‑
96m2
‑
hs
‑
c)中混合,之后添加50μl上述蛋白酶反应溶液。然后将整体混合物在热循环仪(伯乐公司(biorad))中在95℃下孵育15min。添加100μl的75%乙醇后,使用spectramax 190在440nm(a
440
)处测量所得溶液的吸光度。通过从酶样品的a
440
中减去空白对照的a
440
来计算净a
440
,并且然后绘制出相对于不同蛋白质浓度(从0.3125ppm到20ppm)的图。结果示出于图3a和图3b。每个值为一式两份测定的平均值,方差小于5%。因此蛋白水解活性显示为净a
440
。用phe
‑
pro(图3a)或ser
‑
pro(图3b)作为底物的蛋白水解测定表明了malpro11、mcipro4、tcipro1、fvepro4和ssppro2均为活性蛋白酶。
[0187]
实例4纯化的真菌x
‑
pro蛋白酶的ph曲线
[0188]
用phe
‑
pro二肽作为底物,在不同ph值(范围从ph 6至10)的25mm bis
‑
tris丙烷缓冲液中研究纯化的蛋白酶(malpro11、mcipro4、tcipro1、fvepro4和ssppro2)的ph曲线。测定前,首先将45μl具有特定ph值的50mm bis
‑
tris丙烷缓冲液(补充有0.1mm cocl2)与45μl的20mm phe
‑
pro(溶于水)在96
‑
mtp中混合,然后添加10μl用水稀释的酶(12.5ppm的malpro11、25ppm的mcipro4、12.5ppm的tcipro1、12.5ppm的fvepro4、6.25ppm的ssppro2,或仅添加水作为空白对照)。反应的进行和分析如实例3所述。各ph值下的酶活性报告为相对活性,其中最佳ph值下的酶活性为100%。测试的ph值为6、6.5、7、7.5、8、8.5、9.5和10。每个值为一式两份测定的平均值,方差小于5%。如图4中所示,malpro11、mcipro4、tcipro1、fvepro4或ssppro2的最佳ph分别为8、8.5、8.5、8、或8。
[0189]
实例5纯化的真菌x
‑
pro蛋白酶的温度曲线
[0190]
在50mm tris
‑
hcl缓冲液(ph 7.5)中使用phe
‑
pro二肽作为底物来分析纯化的蛋白酶(malpro11、mcipro4、tcipro1、fvepro4和ssppro2)的温度曲线。反应前,将溶解于50mm tris
‑
hcl缓冲液(ph 7.5,补充有0.05mm cocl2)中的90μl 10mm phe
‑
pro二肽添加到200μl pcr管中,然后将其在热循环仪(伯乐公司(biorad))中在所希望的温度(即30℃至80℃)下孵育5min。孵育后,将10μl用水稀释的酶(12.5ppm的malpro11、25ppm的mcipro4、12.5ppm的tcipro1、12.5ppm的fvepro4、6.25ppm的ssppro2或仅添加水作为空白对照)添加至底物溶液中以开始反应。在不同温度的热循环仪中孵育20min后,淬灭反应并分析,如实例3所示。报告的活性为相对活性,其中最佳温度下的活性被设为100%。测试的温度为30℃、35℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、和80℃。每个值为一式两份测定的平均值,方差小于5%。如图5中所示,malpro11、mcipro4、tcipro1、fvepro4或ssppro2的最佳温度分别为55℃、50℃、50℃、45℃或50℃。
[0191]
实例6纯化的真菌x
‑
pro蛋白酶的热稳定性
[0192]
在热稳定性测试前,将phe
‑
pro二肽底物溶解于50mm tris
‑
hcl缓冲液(ph 7.5,补充有0.05mm cocl2)中至最终浓度为10mm。将纯化的蛋白酶(malpro11、mcipro4、tcipro1、
fvepro4和ssppro2)稀释于0.2ml水中至最终浓度为200ppm,随后在不同温度(4℃、55℃、60℃、65℃、70℃、75℃、80℃)下孵育5min。孵育后,将每种酶溶液进一步用水稀释至特定浓度(12.5ppm的malpro11、25ppm的mcipro4、12.5ppm的tcipro1、12.5ppm的fvepro4、6.25ppm的ssppro2或仅添加水作为空白对照)。为测量蛋白水解活性,将10μl所得酶溶液与90μl底物溶液混合;并且如实例3中所述进行反应并分析。将活性报告为残基活性,其中在4℃下孵育的酶样品的活性被设为100%。每个值为一式两份测定的平均值,方差小于5%。如图6中所示,所有蛋白酶在70℃、75℃和80℃下孵育5min后丧失了其活性;并且除了mcipro4,所有其他四种蛋白酶在65℃下孵育5min后也丧失了其活性。
[0193]
实例7纯化的真菌x
‑
pro蛋白酶的五肽水解分析
[0194]
在50mm tris
‑
hcl缓冲液(ph 7.5)中进行纯化的蛋白酶(malpro11、mcipro4、tcipro1、fvepro4和ssppro2)对于五肽gln
‑
pro
‑
gln
‑
gln
‑
pro(吉尔生化公司,上海)(seq id no:9)的蛋白水解活性。反应前,将酶用水稀释到200ppm。将五肽底物溶解于50mm tris
‑
hcl缓冲液(ph7.5,补充有0.05mm cocl2)中至最终浓度为10mm。为了开始反应,将90μl的10mm五肽溶液添加到非结合96
‑
mtp(康宁生命科学公司,#3641),并在恒温混匀仪中在50℃和600rpm下孵育5min,随后添加10μl的稀释酶样品(或仅添加水作为空白对照)。在恒温混匀仪中在50℃和600rpm下孵育1小时后,通过在95℃加热10min来终止蛋白酶反应。
[0195]
应用检测伯胺的茚三酮反应来证明五肽水解。反应前,制备茚三酮溶液,其含有2%茚三酮(w/v)、0.5m醋酸钠、40%乙醇和0.2%果糖(w/v)。为了开始反应,将90μl茚三酮溶液与10μl上述蛋白酶反应溶液在96
‑
mtp pcr板中混合。然后将整体混合物在热循环仪中在95℃下孵育15min。添加100μl的75%乙醇后,使用spectramax 190在570nm(a
570
)处测量所得溶液的吸光度。结果示出于图7中。每个值为一式两份测定的平均值,方差小于5%。那些蛋白酶样品相比于空白对照的a
570
增量表明所有纯化的蛋白酶均能够水解五肽gln
‑
pro
‑
gln
‑
gln
‑
pro。
[0196]
实例8:谷蛋白预水解产物的制备和分析
[0197]
通过schlichtherle
‑
cerny和amad
ò
(2002)中所述方法的修改版本来制备含有水溶性谷蛋白肽和氨基酸的底物。将以下成分在100ml螺盖瓶中混合:6.4g谷蛋白(西格玛
‑
奥德里奇(sigma
‑
aldrich)公司,丹麦哥本哈根)、0.123g acpepn2、0.6g谷氨酰胺酶sd
‑
c100s(天野公司(amano),日本名古屋)、63mg碱性蛋白酶(工业生物科学公司,丹麦布拉布兰)、1.73g nacl(分析级,飞世尔科技公司(fischer scientific),丹麦罗斯基勒)、和24.3g水。将瓶在热块中孵育,同时在600rpm和55℃下磁力搅拌18小时。随后通过加热至95℃并持续10min来使酶失活,将其在4600rpm下离心5min,并且将上清液通过0,45μm注射器过滤器过滤。
[0198]
对于残余肽的n
‑
末端序列测定,将谷蛋白预水解产物通过0,2μm注射器过滤器过滤,并且将其中2μl加载到来自日本岛津公司(shimadzu)的ppsq
‑
31b蛋白质序列分析仪上。制备了25pmol所有20种常见氨基酸的混合物并且将其用作标准品。这些氨基酸在标准品中的保留时间和峰面积被用于鉴定和量化在埃德曼(edman)循环仪的每一步骤后释放的氨基酸。从这些结果中可以衍生出残余肽n
‑
末端的共有序列。此共有序列为:xpqqp,其中x为任一氨基酸,p为脯氨酸并且q为谷氨酰胺。此外,结果示出了73%的残余肽在次末端位置具有脯氨酸。
[0199]
使用对接到orbitrap fusion质谱仪(赛默科技公司)的dionex3000rslcnano lc(赛默科技公司)进行纳米lc
‑
ms/ms分析。用牵引发射器将1μl每种样品加载到2cm阱柱(100μm i.d.,375μm o.d.,c18,5μm反相颗粒)上,所述阱柱连接至15cm分析柱(75μm i.d.,375μm o.d.,填充有reprosil c18,3μm反相颗粒(梅斯奇博士股份有限公司(dr.maisch gmbh),德国阿梅尔布(ammerbuch)
‑
entringen))。使用5%
‑
53%溶剂b(h2o/ch3cn/tfe/hcooh(100/800/100/1)v/v/v/v)的37分钟梯度,以300nl/min的流速进行分离,以进入纳米电喷雾离子源(赛默科技公司)。orbitrap fusion仪器以数据依赖性ms/ms模式运行。通过orbitrap(在m/z 200下以120.000的分辨率获得的ms扫描)测量肽质量,并且从最强的肽m/z中选择尽可能多的离子,并使用线性离子阱(ltq)中的(高能碰撞解离)hcd对其在1.6秒内进行片段化。启动动态排除,列表大小为500个质量,持续时间为40秒,并且相对于列表上的质量的排除质量宽度为
±
10ppm。
[0200]
使用proteome discoverer 2.0和本地mascot服务器处理原始(raw)文件并且针对uniprot green plants进行搜索。使用proteome discoverer2.0中的内置区域检测模块估算出所有所鉴定肽的面积。
[0201]
评估来自谷蛋白水解的残余肽中结合的gln量的重要工具是q
‑
面积。q
‑
面积=q
n
*面积,其中q
n
为gln残基在肽中的数量,并且面积为特定肽产生的色谱峰曲线下面积。
[0202]
结果示出了氨基酸的一个特异性序列或“基序”,xpqqp,其是大部分所检测的肽中所共有的。基于q面积,据估计在n
‑
末端携带此序列基序的肽含有大约60%的残余谷氨酰胺。
[0203]
结论:两个独立的分析技术示出了谷蛋白预水解产物中残余肽的n
‑
末端具有共有序列xpqqp。
[0204]
实例9:x
‑
proap对谷蛋白预水解产物的测试
[0205]
通用程序:反应混合物由250μl谷蛋白预水解产物、11.8μl 50mg/ml谷氨酰胺酶、10.2μl acpepn2和98μg x
‑
proap组成。添加milliq水至310μl或415μl的总体积。总体积在每次实验中都是恒定的,但在实验间会随着所用x
‑
proap的蛋白质浓度而变化。参考样品含有谷氨酰胺酶但既不含有acpepn2也不含有x
‑
proap。总体积与实验中其他样品相同。
[0206]
所有反应混合物均是在eppendorf管中制备的。将管在eppendorf混合器中在50℃和800rpm下孵育。在特定的时间点,将80μl等分试样取出并且与20μl 2.5m tca(飞世尔科技公司,丹麦罗斯基勒)混合以停止进一步反应。使用酶l
‑
谷氨酸试剂盒(来自拜发公司(r
‑
biopharm),达姆施塔特,德国)对水解产物中的谷氨酸浓度进行量化。将所述方法缩减以用于96孔板,不然根据制造商说明书进行。分析前,将tca/样品混合物在milliq水中进一步稀释400次(总稀释因子=500)。
[0207]
根据nielsen等人(nielsen,petersen等人2001)所述的方法基于邻苯二甲醛(opa;飞世尔科技公司,丹麦罗斯基勒)测定来确定水解度(dh)。通过总氨基酸分析(在欧陆集团(eurofins),瓦伊恩,丹麦进行)来确定氨基酸的平均mw。基于此,将h
tot
计算为7.6mmol/g谷蛋白蛋白质。
[0208]
使用尺寸排阻色谱法(sec)分析氨基酸和肽分布。所使用的系统来自赛默飞世尔科技公司(丹麦)并且是由dionex ultimate3000溶剂支架、泵和具有dionex corona超rs充电的气溶胶检测器(cad)的自动进样器、superdex
tm
肽10/300gl柱(来自默克
公司(merck),哥本哈根,丹麦)组成的。7.2版本用于仪器控制和数据处理。流动相由milliq水中的20%乙腈(acn)和0.1%三氟醋酸(tfa;飞世尔科技公司,丹麦罗斯基勒)组成。注射前,将所有样品在流动相中稀释10次并且使用0.2μm pvdf滤板(材料#3504,康宁公司,肯纳邦克(kennebunk)缅因州,美国)进行过滤。注射体积为10μl并且流速为0.500ml/min,持续55min。
[0209]
所有实验所包括的参考样品均含有谷蛋白预水解产物和谷氨酰胺酶。将其暴露于与所有其他样品相同的处理。为了方便不同运行之间的比较,参考样品被设为含有100%谷氨酸(形成于预水解步骤期间)。所有其他结果均以相对于参考样品的%给出。其他样品含有的成分与参考样品相同,但添加了acpepn2和/或x
‑
proap。
[0210]
图8示出了增加剂量的ssppro2对于谷氨酸产率的作用。测试了两个剂量的ssppro2:131μg/ml和392μg/ml的预水解产物。这导致相对于参考样品,谷氨酸分别增加了16%和34%。在给定条件下,仅acpepn2没有给出任何谷氨酸水平的增加。
[0211]
图9示出了来自与图8相同的样品但进行26h孵育后的结果。这种情况下,相对于参考样品,131μg/ml和392μg/ml的tcipro1导致谷氨酸分别增加了25%和71%。这种情况下,相对于参考样品,单独的acpepn2也给出了谷氨酸16%的增加。
[0212]
图10示出了不同x
‑
proap对于谷氨酸产率的作用。孵育时间为24h。这种情况下,相对于参考样品,单独的acpepn2给出了谷氨酸水平8%的增加。与acpepn2组合,malpro11、mcipro4、tcipro1、pchsec117、和ssppro2分别给出了40%、44%、25%、28%和64%的增加。相比之下,当malpro11、mcipro4和ssppro2单独测试时(不含acpepn2),未观察到谷氨酸水平的增长(不超过实验误差)。结果示出了acpepn2和测试的x
‑
proap协同作用以从预水解产物中的残余肽中释放谷氨酸。因为材料的数量有限,tcipro1和pchsec117未在不含acpepn2的情况下接受测试。
[0213]
图11示出了来自两个额外的经测试x
‑
proap的结果。它们仅在孵育19h和26h后给出了微不足道的反应。图11中示出的为孵育42小时后的结果。这种情况下,单独的acpepn2给出了谷氨酸水平9%的增加。aox
‑
proap和hx
‑
proap分别给出了15%和6%的增加。单独的acpepn2与hx
‑
proap之间的差异在实验误差以内。因为材料有限,在此情况下x
‑
proap的剂量仅为15μg/ml预水解产物。
[0214]
在来自用于图8至图11中谷氨酸结果的相同实验的样品上确定水解曲线。以下给出两个实例。在图12中,比较了在392μg/ml预水解产物中acpepn2样品的水解曲线(实线)和含有acpepn2 ssppro2的样品的曲线(虚线)。用acpepn2 ssppro2产生的水解产物的含有氨基酸的峰的峰面积与仅用acpepn2产生的水解产物相比,高1.5倍。同时,acpepn2 ssppro2水解产物的dp2
‑
5面积与仅acpepn2的水解产物相比,减少了1.3倍。dp2
‑
5面积的减少与氨基酸面积的增加不成正比,因为cad的响应因子不等同于氨基酸和dp2
‑
5肽。图13示出了acpepn2样品和含有hx
‑
proap的样品的水解曲线的相似比较。hx
‑
proap导致的氨基酸的增加非常适度。与观察结果一致,这种处理没有增加gln水平。
[0215]
实例10:x
‑
proap对谷蛋白蛋白浆料的测试
[0216]
从谷蛋白蛋白质生产谷氨酸不需要预水解产物。在设置中测试ssppro2,其中所有组分(包括酶)均在实验开始时混合。
[0217]
使用了schlichtherle
‑
cerny和amad
ò
(2002)所述方法的规模缩减版本。将以下成
分在20ml惠顿瓶(wheaton vial)中混合:2.13g谷蛋白、33mg acpepn2、21mg 碱性蛋白酶、0.2g谷氨酰胺酶、1mg ssppro2、0.58g nacl和约8g水。调节水的量,使得所有成分的总重量等于10.5g。将惠顿瓶在热块中孵育,同时在600rpm和55℃下磁力搅拌长达48小时。在不同时间点取出160μl等分试样并且用40μl 2.5m tca来停止反应。将样品进一步稀释400次,并且如实例9中所述分析谷氨酸(所有化学品和酶的供应商都与实例8和实例9中相同)。
[0218]
孵育24h后,含有ssppro2的样品与不含x
‑
proap的参考样品相比,形成的谷氨酸多22%。需要注意的是这种情况下的参考样品含有活性acpepn2,这与谷蛋白预水解产物实验中的参考样品相反,其中预水解产物是用acpepn2 其他酶(随后失活)制备的。在谷蛋白浆料实验中,不含acpepn2的参考样品是没有意义的。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。