一种影响萝卜花青素合成的myb1同源基因及其鉴定方法
技术领域
1.本发明涉及影响萝卜花青素合成的myb1同源基因;特别涉及一种影响萝卜花青素合成的myb1同源基因及其鉴定方法。
背景技术:
2.萝卜(raphanus sativusl.)为十字花科萝卜属蔬菜,在中国常年栽培面积仅次于大白菜,在农业生产中占有重要地位。萝卜的种植区域广泛,种植品种繁多。云南红萝卜是以通海乌萝卜和兰坪紫萝卜为代表的一类富含花青素(anthocyanidin)的特异种质资源。花青素(anthocyanidin)是植物中最大的一类水溶性色素,属于黄酮类代谢物质,具有抑制细胞突变增殖、抗炎、抗菌、抗氧化及降血压等功效(he and giusti,2010)。目前在自然界中分离鉴定的600多种花青素主要由6种花青素苷元衍生而来,分别是矢车菊素苷元、飞燕草素苷元、天竺葵素苷元、芍药花素苷元、矮牵牛素苷元和锦葵素苷元(kong et al.,2003;戴思兰&洪艳,2016)。萝卜中的花青素主要由呈紫色的矢车菊素苷元和呈红色的天竺葵素苷衍生而来(kato et al.,2013)。以云南红萝卜为原料提取的萝卜红色素因色泽红艳、稳定性好,已广泛用于食品、医药和化妆品等行业。云南省自20世纪90年代开始在通海县、兰坪县和东川市大力发展萝卜红色素加工产业,其产品获得了原产地标记认证。截至2019年,云南省年产萝卜红色素100吨,直接产值过亿元,90%以上初加工产品销往美国、日本、欧洲、韩国等地,国际市场占有率在80%以上。因其种植加工技术简单、效益稳定,红萝卜已发展成为云南蔬菜产业中促进山区农民持续增收的一项特色优势产业。但是,生产上使用的红萝卜品种种质混杂,根形差异大,色素含量不均,加工性状参差不齐,致使萝卜红色素提取工艺难度增大,阻碍了产品品质的提升,制约了萝卜红色素产业的进一步发展。因此挖掘云南红萝卜肉质根花青素合成关键调控基因、解析其调控机制,并以此为基础开发相应的分子标记进行辅助育种,将为云南红萝卜的遗传改良提供理论基础和技术支撑。
3.目前国内外的研究者也围绕萝卜肉质色这一重要的商品性状开展了相关研究,遗传多样性及核型分析的结果表明不同肉色萝卜品种间的亲缘关系较近(许江等,2011;方平等,2012);红色肉质萝卜在萌发2
‑
3d后色素就出现,说明花青素的合成在萌发早期就已经启动,分布格局已经固定(吕发生等,2006)。转录组分析表明,红色肉质萝卜中rsdfr、rsans、rsufgt、rsf3h、rschs3和rsf3’h1的表达量要高于其它类型的材料,其中rsufgt的表达是花青素时空积累的关键控制点(park et al,2011;muleke et al.2017;sun et al,2018)。目前已发现外源茉莉酮酸甲酯、赤霉素和uv
‑
a可诱导白色肉质萝卜产生并积累花青素(su et al.2014;al
‑
dhabi et al.2015),表明不同肉色类型萝卜都具备合成花青素的相关结构基因,只是调控其表达的机制不同。最近的研究表明,转录因子rsmyb1/90能够直接调控花青素合成结构基因的表达,是决定萝卜皮色的关键基因(lim et al.,2016,2017;luo et al.,2019),在转基因拟南芥中rsmyb1能直接调控bhlh转录因子的表达;bhlh转录因子rstt8能与rsmyb1/90组成myb
‑
bhlh
‑
wdr(mbw)复合体,共同调节萝卜色素合成(yi et al.2018)。lai等(2019)通过同源克隆的方法鉴定得到了两个rsmyb1/90同源基因,其中位
于第二连锁群的myb1a是涪陵胭脂萝卜花青素生成的关键基因;表观遗传学研究发现,在位于第7连锁群的myb1/90基因启动子区域,由一个cacta转座子插入引起甲基化可导致myb1/90表达量增高,这与心里美萝卜红肉性状密切相关(wang et al.,2020)。萝卜根肉色性状遗传复杂,上述研究结果并不能完全揭示花青素合成调控机制。加之,云南红萝卜多分布于海拔1800m以上的低纬高原,长期处于高紫外线辐射和昼夜温差大的生态环境下,与目前研究较多的北方“心里美”萝卜相比,花青素含量高,叶形、根形及皮色等性状均有较大差别。
4.为阐明云南红萝卜肉质根花青素特异积累的分子机制,本发明人所在课题组利用选育自云南红萝卜地方材料的高代自交系rr1(p1)与加工型萝卜(白皮白肉)高代自交系wr1(p2)进行杂交,创制了bc1p1、bc1p2、f1、f2遗传群体,并对两个亲本材料及200个f2代材料分别进行重测序及slaf简化基因组测序,利用f2遗传群体构建了高密度遗传图谱,对肉质根花青素性状关联qtl定位进行了研究。
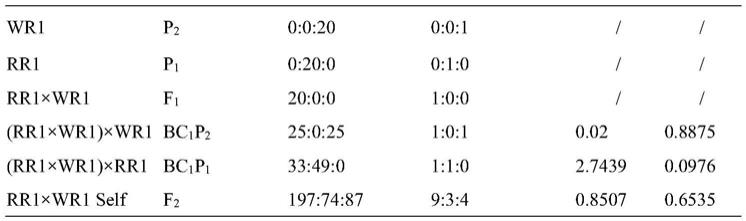
5.表型观测结果发现,当不考虑各个体间的着色部位及程度的情况下,在f2遗传群体中,其具有颜色性状(pigment,pi)个体数和具有无色性状(none
‑
pigment,np)个体数的比例符合3:1的孟德尔分离比,而在bc1p1群体中所有个体均具有颜色,而在bc1p2群体中pi个体数和np个体数的比例为1:1,这表明在已构建的遗传群体中,萝卜肉质根色素合成由主效单基因控制。
6.结合f2个体的花青素含量及已构建的遗传图谱,lod阈值对应于0.99置信水平上,在lg07上检测到一个qtl。利用已发表的萝卜基因组数据,将该位点映射到萝卜基因组物理图谱上,通过对附近位点候选基因的功能注释,初步推断控制萝卜花青素积累的候选基因为rsmyb1.3。半定量pcr实验表明,rsmyb1.3在rr1中表达,而在wr1中不表达;对rsmyb1.3
‑
rr1和rsmyb1.3
‑
wr1的结构进行分析,发现在rsmyb1.3
‑
wr1的第一外显子中具有一个4bp的碱基插入,这一插入导致了rsmyb1.3
‑
wr1提前形成了终止密码子而失去功能。针对这一插入及rsmyb1.3
‑
rr1和rsmyb1.3
‑
wr1的结构特征,设计了两对引物rsmyb1.3
‑
a f/r和rsmyb1.3
‑
b f/r,其中,rsmyb1.3
‑
a f/r可特异性地从rsmyb1.3
‑
wr1扩增得到511bp的片段,而rsmyb1.3
‑
b f/r可特异性地从rsmyb1.3
‑
rr1中扩增得到467bp的片段。利用这两对引物对f1代植株材料进行扩增,发现f1代材料中均可扩增得到511bp和467bp条带,表明rsmyb1.3在f1代材料中为杂合形态。在f2代群体中,在np性状个体中只能扩增得到511bp片段,而在pi性状个体中可得到467bp和511bp/467bp两种形态,可见,利用这两对引物的扩增结果可判断遗传材料中rsmyb1.3的基因型,这两对引物的扩增片段可作为功能标记筛选具有花青素合成功能的遗传材料。
7.在针对云南红萝卜品质、产量等方面的育种和品种改良过程工作中,需要对育种材料进行杂交或回交,创制出大量的育种中间材料,对这些材料肉质根颜色的传统鉴定方法是将萝卜从地里拔出后,切开观察肉质根颜色,该方法费时、费力,还会导致肉质根受到损伤,移栽后成活率低,造成优良育种中间材料的损失。
8.现有的利用分子标记进行萝卜肉质根红肉色,紫皮色的鉴别方案有:
[0009]“一种鉴别萝卜肉质根红肉色的分子标记”[0010]“一种鉴定萝卜肉质根紫皮性状的特异分子标记”[0011]
在针对云南红萝卜品质、产量等方面的育种和品种改良过程工作中,需要对育种材料进行杂交或回交,创制出大量的育种中间材料,对这些材料肉质根颜色的传统鉴定方
法是将萝卜从地里拔出,切开观察肉质根颜色,之后再移栽到土里等待授粉、采种。该方法费时、费力,还会导致肉质根受到损伤,移栽后成活率低,造成优良育种中间材料损失。本发明通过本标记对萝卜种质资源、育种材料和分离后代植株进行早期鉴定,可减少待鉴定材料植株的种植与鉴定规模,大大降低了鉴定和栽培管理过程中的人力、物力和财力的投入,解决了传统根肉色鉴定方法将肉质根切开观察,导致受损种根贮藏困难、成活率低的难题,提高选择效率;此外,该标记可用于进行萝卜呈色性状的分子标记辅助育种,提高复杂性状的遗传改良效率。
[0012]
现有分子标记鉴定萝卜红色肉质根的方案的缺点:(1)所定位到的功能基因与本研究的基因不同,由于萝卜花青素产生的遗传机制在不同萝卜种质中存在基因型差异,导致rsmyb1.3失活的4
‑
bp碱基插入为首次发现。(2)检测方法不同,本方法特异地对导致rsmyb1.3失活的4
‑
bp碱基插入位点进行检测,而目前方法则是对位于不同染色体位置的myb基因内含子大小进行检测。
技术实现要素:
[0013]
为解决上述现有技术存在的问题,本发明的目的在于利用设计标记对苗期萝卜植株叶片组织的dna进行pcr扩增,依照其电泳结果可对rsmyb1.3在无花青素合成萝卜及云南红萝卜中的基因型进行鉴定。本发明获得的标记可对萝卜种质资源、育种材料和分离后代植株进行早期鉴定,减少待鉴定材料植株的种植与鉴定规模,大大降低鉴定过程中的工作量和物力财力的投入,提高选择效率;同时也可以解决传统肉质根颜色鉴定方法将肉质根切开观察,导致种根贮藏困难、成活率低的问题。
[0014]
为达到上述目的,本发明的技术方案为:
[0015]
一种影响萝卜花青素合成的myb1同源基因rsmyb1.3,其特征在于,该基因为rsmyb1(loc_108814812)同源基因,是影响萝卜花青素合成的关键基因,无色素类型萝卜中该基因第197位碱基处由于一个4
‑
bp的片段(5
’‑
aatt
‑3’
)插入而提前形成终止密码子,从而导致植株中无花青素合成;云南红萝卜中rsmyb1.3第197位碱基处则无插入,可正常表达,植株内有花青素产生。
[0016]
基于权利要求1基因的分子标记,其特征在于,基于rsmybl.3基因启动子区域内的indel位点差异开发,rsmyb1.3
‑
a f/r标记可特异性扩增“aatt”碱基插入的rsmyb1.3单倍型基因,适用于无色素类型萝卜的鉴定;rsmyb1.3
‑
b f/r可特异性扩增云南红萝卜中rsmyb1.3单倍型基因。
[0017]
所述分子标记如下:
[0018]
rsmyb1.3
‑
a f:5'
‑
gggaaatggcaccaagttaa
‑
3';
[0019]
rsmyb1.3
‑
a r:5'
‑
tcccaaaagtttatgaaggcga
‑
3'
[0020]
rsmyb1.3
‑
b f:5'
‑
atgcacgtcactaccctcatt
‑
3'
[0021]
rsmyb1.3
‑
b r:5'
‑
tttaataaacgacaatccaacac
‑
3'
[0022]
进一步的,所述分子标记在鉴别萝卜肉质根呈色方面的应用。
[0023]
进一步的,基于上述分子标记的鉴别萝卜植株是否呈色的方法,该方法以待鉴别萝卜品种或分离群体中单株的基因组dna为模板,通过pcr扩增,对扩增产物进行鉴定,若扩增结果有且仅有511bp条带,说明该待鉴定材料肉质根皮、肉色均为白色,植株叶脉为绿色;
若待鉴定品种或材料扩增出该467bp条带或者同时扩增出511bp和467bp条带,说明该鉴定品种或材料潜在为肉质根根皮、根肉或者叶脉带色的萝卜品种或单株。
[0024]
进一步的,所述方法中的待鉴别萝卜材料是来自育种亲本材料、分离世代群体或市场上常见的萝卜品种,实验中需要用到待测材料的dna,该dna可以从待测材料的种子、苗期叶片或者生长中后期的萝卜植株中提取获得。
[0025]
进一步的,所述的分子标记在具有白皮,白肉,绿色茎秆或肉质根根皮、根肉或者叶脉带色萝卜分子辅助选择育种中的应用。
[0026]
相对于现有技术,本发明的有益效果为:
[0027]
利用设计标记对苗期萝卜植株叶片组织的dna进行pcr扩增,依照其电泳结果可对rsmyb1.3在无花青素合成萝卜及云南红萝卜中的基因型进行鉴定。本发明获得的标记可对萝卜种质资源、育种材料和分离后代植株进行早期鉴定,减少待鉴定材料植株的种植与鉴定规模,大大降低鉴定过程中的工作量和物力财力的投入,提高选择效率;同时也可以解决传统肉质根颜色鉴定方法将肉质根切开观察,导致种根贮藏困难、成活率低的问题。
[0028]
现有分子标记鉴定萝卜红色肉质根的方案的缺点:所定位到的功能基因与本研究得到的基因不同,由于萝卜花青素产生的遗传机制在不同萝卜种质中存在基因型差异,所以已有标记无法在云南红萝卜这一特异资源的遗传育种中进行利用。
附图说明
[0029]
图1.a.亲本自交系和f2个体的表型。(a)母系
‘
yaas
‑
rr1’。(b)父系
‘
yaas
‑
wr1’。(d
‑
l)不同颜色的f2个体的表型。b.f2个体色素皮肤性状的频率分布。c.f2个体色素肉性状的频率分布。
[0030]
图2.与萝卜呈色有关qtl定位。横坐标表示连锁群中标记的顺序。上面的纵坐标和下面的纵坐标表示lod值,中间纵坐标表示表型贡献率。上图和下图的曲线表示所检测qtl的遗传坐标和lod评分(top)。蓝线为标记所对应的lod值;红线为该标记物对应的表型贡献率;灰线是阈值线。
[0031]
图3.基于半定量pcr分析的rsmyb1.3在亲本材料间的基因表达谱。
[0032]
图4.白萝卜和红萝卜rsmyb1.3序列比对。
“‑”
表示与参考基因组中的等位基因loc202036相同的碱基。“.”表示缺失的碱基。提前终止密码子用红框表示。
[0033]
图5.开发引物pcr标记f1、f2个体的结果。(a代表引物rsmyb1.3
‑
af/r;b代表引物rsmyb1.3
‑
bf/r;编号3,4,17,106,184,196分别代表肉质根根皮、根肉或者叶脉带红色的f2单株;编号为6,9,25,102,163,175分别代表肉质根根皮、根肉或者叶脉带紫色的f2单株;编号为5,36,47,91,93分别代表者白皮,白肉,绿色茎秆的f2单株)
具体实施方式
[0034]
下面结合附图和具体实施方式对本发明技术方案做进一步详细描述:
[0035]
如图1
‑
5所示,
[0036]
实施例1:一种影响萝卜花青素合成的myb1同源基因及其分子标记开发
[0037]
为阐明云南红萝卜肉质根花青素特异积累的分子机制,发明人利用选育自云南红萝卜地方材料的高代自交系rr1(p1)与加工型萝卜(白皮白肉)高代自交系wr1(p2)进行杂
交,创制了bc1p1、bc1p2、f1、f2遗传群体,并对两个亲本材料及200个f2代材料分别进行重测序及slaf简化基因组测序,利用f2遗传群体构建了高密度遗传图谱,对肉质根花青素性状关联qtl定位进行了研究。
[0038]
表型观测结果发现,当不考虑各个体间的着色部位及程度的情况下,在f2遗传群体中,其具有颜色性状(pigment,pi)个体数和具有无色性状(none
‑
pigment,np)个体数的比例符合3:1的孟德尔分离比(表1),而在bc1p1群体中所有个体均具有颜色,而在bc1p2群体中pi个体数和np个体数的比例为1:1(表1),表明在已构建的遗传群体中,萝卜肉质根色素合成由主效单基因控制。
[0039]
结合f2个体的花青素含量及已构建的遗传图谱,lod阈值对应于0.99置信水平上,在lg07上检测到一个qtl(图2)。利用已发表的萝卜基因组数据,将该位点映射到萝卜基因组物理图谱上,通过对附近位点候选基因的功能注释,初步推断控制萝卜花青素积累的候选基因为rsmyb1.3。半定量pcr实验表明,rsmyb1.3在rr1中表达,而在wr1中不表达(图3);对rsmyb1.3
‑
rr1和rsmyb1.3
‑
wr1的结构进行分析,发现在rsmyb1.3
‑
wr1的第一外显子中具有一个4bp的碱基插入(图4),这一插入导致了rsmyb1.3
‑
wr1提前形成了终止密码子而失去功能。针对这一插入及rsmyb1.3
‑
rr1和rsmyb1.3
‑
wr1的结构特征,设计了两对引物rsmyb1.3
‑
a f/r和rsmyb1.3
‑
b f/r,其中,rsmyb1.3
‑
af/r可特异性地从rsmyb1.3
‑
wr1扩增得到511bp的片段,而rsmyb1.3
‑
bf/r可特异性地从rsmyb1.3
‑
rr1中扩增得到467bp的片段。
[0040]
表1.6个遗传群体萝卜色素表型分离鉴定
[0041][0042][0043]
实施例2:影响萝卜花青素合成的myb1同源基因分子标记在后代群体中的验证
[0044]
利用这两对引物对f1代植株材料进行扩增,发现f1代材料均可扩增得到511bp和467bp条带(图5),表明rsmyb1.3在f1代材料中为杂合形态。在f2代群体中具有np性状个体中只能扩增得到511bp片段,而在pi性状个体中可得到467bp和511bp/467bp两种形态(图5),说明f2代群体材料中的rsmyb1.3位点既有杂合个体也有纯和个体。可见,利用这两对引物进行扩增,其结果可判断遗传材料中rsmyb1.3的基因型,这两对引物的扩增片段可作为功能标记筛选具有花青素合成功能的遗传材料。
[0045]
无花青素积累性状萝卜的rsmyb1.3单倍型基因(rsmyb1.3
‑
wr1)由于4
‑
bp的碱基插入而失去功能,从而导致萝卜植株无法合成花青素。rsmyb1.3
‑
wr1是首次鉴定得到,在萝卜自然群体中分布较广,对于育种工作具有较大的利用价值。
[0046]
提供一组鉴定萝卜色素合成决定基因rsmyb1.3基因型的pcr引物序列:
[0047]
rsmyb1.3
‑
a f:5'
‑
gggaaatggcaccaagttaa
‑
3';
[0048]
rsmyb1.3
‑
a r:5'
‑
tcccaaaagtttatgaaggcga
‑
3'
[0049]
rsmyb1.3
‑
b f:5'
‑
atgcacgtcactaccctcatt
‑
3'
[0050]
rsmyb1.3
‑
b r:5'
‑
tttaataaacgacaatccaacac
‑
3'
[0051]
图1.a.亲本自交系和f2个体的表型。(a)母系
‘
yaas
‑
rr1’。(b)父系
‘
yaas
‑
wr1’。(d
‑
l)不同颜色的f2个体的表型。b.f2个体色素皮肤性状的频率分布。c.f2个体色素肉性状的频率分布。
[0052]
图2.与萝卜呈色有关qtl定位。横坐标表示连锁群中标记的顺序。上面的纵坐标和下面的纵坐标表示lod值,中间纵坐标表示表型贡献率。上图和下图的曲线表示所检测qtl的遗传坐标和lod评分(top)。蓝线为标记所对应的lod值;红线为该标记物对应的表型贡献率;灰线是阈值线。
[0053]
图3.基于半定量pcr分析的rsmyb1.3在亲本材料间的基因表达谱。
[0054]
图4.白萝卜和红萝卜rsmyb1.3序列比对。
“‑”
表示与参考基因组中的等位基因loc202036相同的碱基。“.”表示与其他等位基因相比缺失的碱基。提前终止密码子用红框表示。
[0055]
图5.开发引物pcr标记f1、f2个体的结果。(a代表引物rsmyb1.3
‑
a f/r;b代表引物rsmyb1.3
‑
b f/r;编号3,4,17,106,184,196分别代表肉质根根皮、根肉或者叶脉带红色的f2单株;编号为6,9,25,102,163,175分别代表肉质根根皮、根肉或者叶脉带紫色的f2单株;编号为5,36,47,91,93分别代表者白皮,白肉,绿色茎秆的f2单株)
[0056]
以上所述,仅为本发明的具体实施方式,但本发明的保护范围并不局限于此,任何不经过创造性劳动想到的变化或替换,都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应该以权利要求书所限定的保护范围为准。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。