1.本发明总体上涉及感染和炎性疾病以及这些感染性疾病的长期后遗症的治疗,并且特别涉及硫酸葡聚糖在治疗这种冠状病毒感染和感染性疾病以及可能由冠状病毒感染引起的炎性疾病中的用途。
背景技术:
2.冠状病毒(cov)是一组引起哺乳动物和鸟类疾病的相关包膜病毒。在人类中,冠状病毒感染可能导致呼吸道感染,所述感染可能是轻微的,诸如一些普通感冒病例,以及其他可能是致命的,诸如严重急性呼吸综合征(sars)、中东呼吸综合征(mers)和冠状病毒疾病2019(covid-19)。
3.冠状病毒的危险系数差异很大。冠状病毒可能会引起具有诸如发烧和腺样体肿大引起的喉咙痛的主要症状的感冒。冠状病毒也可能引起肺炎,无论是直接病毒性肺炎还是继发性细菌性肺炎,以及支气管炎,无论是直接病毒性支气管炎还是继发性细菌性支气管炎。冠状病毒也可能与长期器官疾病和功能性衰弱有关,有时称为“长期covid”。
4.2019年12月,中国武汉报道了肺炎疫情。2019年12月31日,疫情被追溯到一种新的冠状病毒株,世界卫生组织(who)将其临时命名为2019-ncov,后来国际病毒分类委员会(international committee on taxonomy of viruses)将其更名为严重急性呼吸综合征冠状病毒2(sars-cov-2)。武汉毒株已被鉴定为来自2b组的β冠状病毒(β-cov)的新毒株,与sars-cov具有约70%的遗传相似性。
5.肺部通常是受sars-cov-2影响最大的器官,因为病毒通过在肺部的ii型肺泡细胞中最丰富的酶血管紧张素转换酶2(ace2)进入宿主细胞。所述病毒使用一种称为“刺突”(膜粒)的特殊表面糖蛋白来接合ace2并进入宿主细胞。存在于宿主细胞表面上的硫酸乙酰肝素有助于刺突蛋白与ace2之间的这种结合。随着肺泡疾病的发展,呼吸衰竭可能会发展,并且死亡可能随之而来。
6.冠状病毒也可能影响胃肠器官,因为ace2在胃、十二指肠和直肠上皮的腺细胞以及小肠的内皮细胞和肠上皮细胞中大量表达。
7.由sars-cov-2(即covid-19)引起的感染性疾病具有发热、咳嗽和气短形式的常见症状。肌肉疼痛、咳痰、腹泻和喉咙痛不太常见。虽然大多数病例会导致轻微症状,但有些会发展为肺炎,并且在那些受影响最严重的病例中,covid-19可能会迅速发展为急性呼吸窘迫综合征(ards),从而引起呼吸衰竭、感染性休克、过度炎症、氧化应激、神经损害、微血栓形成、纤维化和/或多器官衰竭,特别是在长期covid患者中。
8.全世界都在努力研发针对sars-cov-2的疫苗。目前,缺乏针对covid-19和sars-cov-2感染以及受损组织的长期有害后果的有效治疗。因此,普遍需要一种能有效治疗冠状病毒感染(包括covid-19)的治疗。
技术实现要素:
9.总的目的是提供冠状病毒感染或感染性疾病的治疗。
10.一个特别的目的是提供sars-cov-2感染和covid-19的治疗。
11.这些目标和其他目标通过如本文公开的实施方案来满足。
12.本发明的一个方面涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制和/或治疗冠状病毒感染或感染性疾病。
13.本发明的另一方面涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制或治疗选自急性呼吸窘迫综合征(ards)和全身炎性响应综合征(sirs)组成的组的炎性疾病。
14.本发明的其他方面涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制和/或治疗由能够结合细胞表面硫酸乙酰肝素蛋白聚糖(hspg)的病原体引起的感染或感染性疾病。
15.本文提供的实验数据指示,硫酸葡聚糖或其药学上可接受的盐可用于治疗冠状病毒感染和冠状病毒感染性疾病以及由能够结合hspg的病原体引起的其他感染及其长期后果。硫酸葡聚糖或其药学上可接受的盐也能够刺激修复性生长因子从组织储备中释放,并且压制选定免疫细胞中的促炎细胞因子。这意味着硫酸葡聚糖或其药学上可接受的盐预防或至少显著抑制循环血液中这些细胞因子水平的增加,所述细胞因子水平的增加引起冠状病毒感染后或ards或sirs中的炎性疾病。实施方案的硫酸葡聚糖还具有消退由炎性病况引起的炎性疤痕的潜力,并且促进功能性组织再生。这些作用与硫酸葡聚糖在诱导代谢正常化和改善肌肉和肝脏方面的作用一起对长期covid-19患者很重要。
附图说明
16.通过结合附图参考以下描述,可最好地理解实施方案及其进一步的目的和优点,其中:
17.图1示出淀粉样蛋白-β和prpc之间的蛋白质-蛋白质相互作用的硫酸葡聚糖竞争。
18.图2示出在施用低分子量硫酸葡聚糖(lmw-ds)之前和之后2小时,肌萎缩侧索硬化(als)患者的血浆肝细胞生长因子(hgf)水平。
19.图3是示出lmw-ds诱导的大脑谷氨酸水平变化的图。
20.图4a-4d是示出lmw-ds改变的腺嘌呤核苷酸(atp、adp、amp)水平和atp/adp比率作为线粒体磷酸化能力的测量的图。
21.图5a-5d是示出lmw-ds改变的氧化型和还原型烟碱型辅酶水平的图。
22.图6a-6c是示出lmw-ds改变的代表氧化应激的生物标志物水平的图。
23.图7是示出lmw-ds改变的硝酸盐水平作为no介导的硝化应激的测量的图。
24.图8a-8c是示出lmw-ds改变的n-乙酰天冬氨酸(naa)及其底物水平的图。
25.图9示出在创伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg体重)的情况下,在严重创伤性脑损伤(stbi)后2天处死的大鼠的去蛋白脑匀浆中测量的naa浓度。对照组以假手术动物为代表。数值是12只动物的平均值。标准偏差用竖线表示。*与对照组显著不同,p《0.01。**与stbi 2天显著不同,p《0.01。
26.图10示出在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg体重和重复施用15mg/kg体重)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量
的atp浓度。对照组以假手术动物为代表。数值是12只动物的平均值。标准偏差用竖线表示。*与对照组显著不同,p《0.01。**与stbi 2天显著不同,p《0.01。
27.图11示出在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg体重和重复施用15mg/kg体重)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的抗坏血酸浓度。对照组以假手术动物为代表。数值是12只动物的平均值。标准偏差用竖线表示。*与对照组显著不同,p《0.01。**与stbi 2天显著不同,p《0.01。
28.图12示出在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg体重和重复施用15mg/kg体重)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的谷胱甘肽(gsh)浓度。对照组以假手术动物为代表。数值是12只动物的平均值。标准偏差用竖线表示。*与对照组显著不同,p《0.01。**与stbi 2天显著不同,p《0.01。
29.图13示出在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg体重和重复施用15mg/kg体重)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的naa浓度。对照组以假手术动物为代表。数值是12只动物的平均值。标准偏差用竖线表示。*与对照组显著不同,p《0.01。**与stbi 2天显著不同,p《0.01。
30.图14示出在lmw-ds施用后血液中活化的部分凝血活酶时间(aptt)的变化。
31.图15示出在不存在(盐水)或存在lmw-ds(4.0μm)的情况下,用转化生长因子β2(tgfβ2)(1.0ng/ml)刺激人小梁网细胞72小时,随后进行纤连蛋白(fibronectin)的免疫荧光标记(绿色)和hoecsht细胞核染色(青色)。(15a)代表性图像是来自共聚焦z堆叠的最大强度投影,以及(15b)直方图显示纤连蛋白染色的定量(平均值
±
sem,n=6),p《0.05(威尔科克森检验(wilcoxon test))。
32.图16示出(16a)显示用于诱导前段纤维化模型的方法的示意图。每周两次前房内注射tgfβ1诱导小梁网(tm)纤维化,阻断aqh流出并升高iop。(16b)显示在ic tgfβ1治疗的第一个14天,接着ic tgfβ1与每日皮下盐水媒介物对照或lmw-ds治疗的另外14天期间的iop测量的线图。灰色阴影区域指示正常的iop水平,**p《0.01,****p《0.0001(双向anova)。(16c 16d)包括前段的角度的眼组织切片的代表性图像以及显示了盐水和lmw-ds治疗组中tm中免疫反应性层粘连蛋白和纤连蛋白染色的水平的相关直方图,**p《0.01***p《0.001(t检验)。(16e)显示来自盐水和lmw-ds治疗组的视网膜切片中的rgc标志物brn3a的代表性图像和直方图,**p《0.01(曼-惠特尼检验(mann-whitney test)),gcl-神经节细胞层。(16f)显示盐水和lmw-ds治疗组中的分段rnfl(箭头)的代表性光学相干断层扫描图像和相关直方图,****p《0.0001(t检验)。盐水组n=5,lmw-ds组n=7。(16g)显示lmw-ds在poag中的潜在机制的示意图。
33.图17将pbmc在不存在(培养基,未刺激)或存在以下刺激的情况下培养24小时:(17a)lps(0.01ng/ml),(17b)肽聚糖(30ng/ml),(17c)商陆有丝分裂原(1.0μg/ml)(17d)pha-l(1.0μg/ml),(17e、17f)cpg(0.2μm或1.0μm) il-15(15ng/ml),或(17g、17h)cytostim(10μl/ml或30μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
tm
,呈60μg/m、200μg/m或600μg/ml)。通过elisa定量上清液中的il-6水平。数据表示为来自六个或10-11个(lps)供体的平均值
±
sem。数据绘制为刺激物 媒介物百分比。(-)指示至少一个供体低于检测限。**媒介物 lps和600μg/ml ilb
tm
lps之间的曼-惠特尼u检验比较,p=0.005。*媒介物 pha-l和600μg/ml ilb
tm
pha-l之间的曼-惠特尼u检验比较,p=0.048。
34.图18将从pbmc纯化的单核细胞在不存在刺激(培养基)的情况下培养或用lps(0.01ng/ml)或肽聚糖(30ng/ml)在不存在(媒介物)或存在(18a、18d)lmw-ds(ilb
tm
;60μg/ml、200μg/ml或600μg/ml)、(18b、18e)地塞米松(dexamethasone)(3.0μm)或(18c、18f)肝素(2.0μg/ml、6.0μg/ml或20μg/ml)的情况下刺激24小时。通过elisa定量细胞培养物上清液中的il-6水平。数据表示为平均值
±
sem,n=10。*指示低于检测限(5pg/ml)。 p《0.05, p《0.001对刺激的显著性差异(曼-惠特尼u检验)。
35.图19将pbmc在不存在刺激(培养基,未刺激)或存在以下刺激的情况下培养24小时:lps(0.01ng/ml)、肽聚糖(30ng/ml)、pha-l(1.0μg/ml)、cpg(0.2μm) il-15(15ng/ml)、商陆有丝分裂原(1.0μg/ml)或cytostim(10μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
tm
,呈60μg/ml、200μg/ml或600μg/ml)。通过luminex定量上清液中的干扰素γ(ifnγ)水平。除非另有指示,否则数据表示为刺激物 媒介物百分比和来自12个供体的平均值
±
sem。(-)指示至少一个重复低于定量限,( )指示至少一个重复高于定量限,(^)指示来自11个供体的数据,(*)指示来自6个供体的数据。与用媒介物刺激进行比较:#p《0.05,##p《0.01,###p《0.001,并且n.s指示不显著(曼-惠特尼检验,双尾)。
36.图20将pbmc在不存在刺激(培养基,未刺激)或存在以下刺激的情况下培养24小时:lps(0.01ng/ml)、肽聚糖(30ng/ml)、pha-l(1.0μg/ml)、cpg(0.2μm) il-15(15ng/ml)、商陆有丝分裂原(1.0μg/ml)或cytostim(10μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
tm
,呈60μg/ml、200μg/ml或600μg/ml)。通过luminex定量上清液中的白细胞介素8/趋化因子(c-x-c基序)配体8(il-8/cxcl8)水平。除非另有指示,否则数据表示为刺激物 媒介物百分比和来自12个供体的平均值
±
sem。(-)指示至少一个重复低于定量限,( )指示至少一个重复高于定量限,(^)指示来自11个供体的数据,(*)指示来自6个供体的数据。与用媒介物刺激进行比较:#p《0.05,##p《0.01,###p《0.001,并且n.s指示不显著(曼-惠特尼检验,双尾)。
37.图21将pbmc在不存在刺激(培养基,未刺激)或存在以下刺激的情况下培养24小时:lps(0.01ng/ml)、肽聚糖(30ng/ml)、pha-l(1.0μg/ml)、cpg(0.2μm) il-15(15ng/ml)、商陆有丝分裂原(1.0μg/ml)或cytostim(10μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
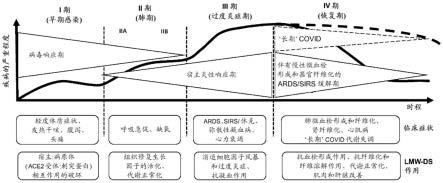
tm
,呈60μg/ml、200μg/ml或600μg/ml)。通过luminex定量上清液中的肿瘤坏死因子α(tnfα)水平。除非另有指示,否则数据表示为刺激物 媒介物百分比和来自12个供体的平均值
±
sem。(-)指示至少一个重复低于定量限,( )指示至少一个重复高于定量限,(^)指示来自11个供体的数据,(*)指示来自6个供体的数据。与用媒介物刺激进行比较:#p《0.05,##p《0.01,###p《0.001,并且n.s指示不显著(曼-惠特尼检验,双尾)。
38.图22将pbmc在不存在刺激(培养基,未刺激)或存在以下刺激的情况下培养24小时:lps(0.01ng/ml)、肽聚糖(30ng/ml)、pha-l(1.0μg/ml)、cpg(0.2μm) il-15(15ng/ml)、商陆有丝分裂原(1.0μg/ml)或cytostim(10μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
tm
,呈60μg/ml、200μg/ml或600μg/ml)。通过luminex定量上清液中的il-1β水平。除非另有指示,否则数据表示为刺激物 媒介物百分比和来自12个供体的平均值
±
sem。(-)指示至少一个重复低于定量限,( )指示至少一个重复高于定量限,(^)指示来自11个供体的数据,(*)指示来自6个供体的数据。与用媒介物刺激进行比较:#p《0.05,##p《0.01,###p《0.001,并且n.s指示不显著(曼-惠特尼检验,双尾)。
39.图23将pbmc在不存在刺激(培养基,未刺激)或存在以下刺激的情况下培养24小时:lps(0.01ng/ml)、肽聚糖(30ng/ml)、pha-l(1.0μg/ml)、cpg(0.2μm) il-15(15ng/ml)、商陆有丝分裂原(1.0μg/ml)或cytostim(10μl/ml)加媒介物(0.027%盐水)或lmw-ds(ilb
tm
,呈60μg/ml、200μg/ml或600μg/ml)。通过luminex定量上清液中的il-10水平。除非另有指示,否则数据表示为刺激物 媒介物百分比和来自12个供体的平均值
±
sem。(-)指示至少一个重复低于定量限,( )指示至少一个重复高于定量限,(^)指示来自11个供体的数据,(*)指示来自6个供体的数据。与用媒介物刺激进行比较:#p《0.05,##p《0.01,###p《0.001,并且n.s指示不显著(曼-惠特尼检验,双尾)。
40.图24(24a、24b)sars-cov-2附着于细胞表面的模型和(24c)用lmw-ds抑制刺突蛋白与ace2之间的相互作用。
41.图25示意性地示出lmw-ds结合covid-19的有益效果和作用。
42.图26在lmw-ds治疗后患者中的血清naa水平。
43.图27在lmw-ds治疗后患者中的血清尿酸水平。
44.图28在lmw-ds治疗后患者血清中的氧嘌呤总量。
45.图29在lmw-ds治疗后患者中的血清硝酸盐水平。
46.图30在lmw-ds治疗后患者中的血清硝酸盐 亚硝酸盐水平。
47.图31在lmw-ds治疗后患者中的血清mda水平。
48.图32在lmw-ds治疗后患者中的血清ala水平。
49.图33在lmw-ds治疗后患者中的血清citr水平。
50.图34在lmw-ds治疗后患者中的血清orn/citr水平。
51.图35在lmw-ds治疗后患者中的血清α-生育酚水平。
52.图36在lmw-ds治疗后患者中的血清γ-生育酚水平。
53.图37使用life elisa测定评估lmw-ds对sars-cov-2刺突蛋白与ace-2相互作用的影响。数据代表平均值
±
sem(n=3)。
54.图38示意性示出在硫酸葡聚糖治疗后als患者中的血清乳酸盐水平。
55.图39示意性示出在硫酸葡聚糖治疗后als患者中的alsaq-40adl评分。
56.图40示意性地示出在硫酸葡聚糖治疗后als患者中的血清肌红蛋白水平。
57.图41示意性地示出在硫酸葡聚糖治疗后als患者中的血清肌酸激酶水平。
58.图42示意性地示出在硫酸葡聚糖治疗后als患者中的血清肝细胞生长因子(hgf)水平。
59.图43示出在硫酸葡聚糖治疗后als患者血清中的总胆红素水平。
具体实施方式
60.本发明总体上涉及感染和炎性疾病以及这些感染性疾病的长期后遗症的治疗,并且特别涉及硫酸葡聚糖在治疗这种冠状病毒感染和感染性疾病以及可能由冠状病毒感染引起的炎性疾病中的用途。
61.冠状病毒(cov)在冠状病毒科、巢状病毒目和核病毒域中构成了正冠状病毒亚科。其是包膜病毒,具有正义单链rna基因组和螺旋对称的核衣壳。已知六种人冠状病毒,其中一种分为两种不同的毒株,总共有七种人冠状病毒毒株。这些毒株中的四种通常产生普通
感冒的轻微症状;β-cov属的人冠状病毒oc43(hcov-oc43)、β-cov属的人冠状病毒hku1(hcov-hku1)、α-cov属的人冠状病毒229e(hcov-229e)以及α-cov属的人冠状病毒nl63(hcov-nl63)。三种毒株产生潜在严重的症状;所有这三种都是β-cov毒株;中东呼吸综合征相关冠状病毒(mers-cov)、严重急性呼吸综合征冠状病毒(sars-cov或sars-cov-1)和严重急性呼吸综合征冠状病毒2(sars-cov-2)。
62.如本文中所使用的冠状病毒和冠状病毒感染或感染性疾病是指哺乳动物,优选人、受试者或患者中的任何冠状病毒和由这种冠状病毒引起的感染或感染性疾病。在一个实施方案中,冠状病毒选自由mers-cov、sars-cov和sars-cov-2组成的组,并且冠状病毒可引起选自由mers-cov的mers、sars-cov的sars和sars-cov-2的covid-19组成的组的冠状病毒感染性疾病。在一特定的实施方案中,冠状病毒是sars-cov-2,并且冠状病毒感染性疾病是covid-19。
63.本发明涉及硫酸葡聚糖在预防、抑制和/或治疗冠状病毒感染和冠状病毒感染性疾病及其后果中的用途。如本文所示,实施方案的硫酸葡聚糖具有与sars-cov-2感染者的初始感染、疾病进展和不良组织响应相关的多模态作用机制(图25)。实施方案的硫酸葡聚糖对患有covid-19症状长期效应的患者也具有积极作用,所述长期效应也称为长期covid和后covid病况。
64.这些多模式作用机制包括在早期感染阶段破坏宿主-病原体蛋白质-蛋白质相互作用,从而抑制或压制冠状病毒与靶分子(诸如sars-cov-2的ace2)的结合,从而防止或至少抑制病毒进入宿主细胞。
65.如图24a和24b所示,像其他β冠状病毒一样,sars-cov-2的附着和细胞进入由刺突糖蛋白(spg)介导。spg不仅与其受体血管紧张素转换酶2(ace2)相互作用,还与糖胺聚糖结合,诸如硫酸乙酰肝素(hs),其以硫酸乙酰肝素蛋白聚糖(hspg)的形式存在于大多数哺乳动物细胞的表面。spg与细胞表面上系栓的hs的结合增加了病毒颗粒在细胞表面的局部浓度,并且促进了spg与ace2的结合(图24b)。实施方案的可溶性未系栓硫酸葡聚糖能够在受体呈递之前以hs形式与spg结合,从而抑制spg与细胞表面上的hspg和ace2的结合,并且防止或至少显著减少冠状病毒(诸如sars-cov-2)在受感染受试者中的局部浓度、附着和细胞进入(图24c)。因此,实施方案的硫酸葡聚糖因此不仅可用于预防或至少抑制冠状病毒感染,而且一旦受试者已经被冠状病毒感染,还将通过干扰冠状病毒与细胞表面上的ace2和hspg之间的相互作用来限制冠状病毒在受试者体内的传播和复制。
66.实施方案的硫酸葡聚糖还可以减少冠状病毒的传播,即冠状病毒在群体中的传播。更详细地,实施方案的硫酸葡聚糖抑制冠状病毒接近人类细胞,从而抑制病毒复制和任何随后的从感染细胞的病毒脱落。这种减少的病毒脱落负荷不仅通过限制受试者体内的传播而有益于受试者,而且有益于群体,因为受感染的受试者因此不太可能将冠状病毒传播给其他受试者。
67.因此,硫酸葡聚糖通过与病毒颗粒上的spg结合来破坏ace2受体与spg之间的相互作用而在阶段i(早期感染,图25)具有如图24a-24c、37中所示的医疗效果。
68.实施方案的硫酸葡聚糖还具有代谢正常化和组织修复生长因子(诸如肝细胞生长因子(hgf))的活化的作用(图2、42)。实施方案的硫酸葡聚糖的这些作用对于在肺期(阶段ii,图25)期间压制冠状病毒在受感染受试者中的负面作用是重要的,在肺期,所述病毒可
引起肺炎并通过影响生长因子介导的无疤痕组织修复而负面影响受试者的呼吸组织。实施方案的硫酸葡聚糖还具有消退和逆转已形成的疤痕的能力,从而允许受损组织愈合。此外,实施方案的硫酸葡聚糖能够诱导细胞中的代谢正常化,诸如在改善的线粒体功能中所见(图4a-4d,27-28;表5-8、15-18)。
69.更详细地说,实施方案的硫酸葡聚糖保护线粒体功能并减少氧化应激,这在改善抗氧化状态的恢复、通过保持atp产生和代谢保护线粒体atp能量供应、线粒体磷酸化能力的正常化(均由硫酸葡聚糖诱导)中可见。因此,硫酸葡聚糖能够正常化、保护和保存暴露于损害或疾病的细胞中的线粒体功能,这对于获得能够对抗感染性疾病的最佳功能细胞是重要的。
70.因此,实施方案的硫酸葡聚糖通过活化组织修复生长因子和诱导代谢正常化而在感染的阶段ii(肺期)具有医疗效果(图25)。
71.实施方案的硫酸葡聚糖还对任何随后的与临床症状相关的过度炎症期有效,包括急性呼吸窘迫综合征(ards)、全身炎性响应综合征(sirs)、脓毒性休克、弥散性血管内凝血(dic)和甚至器官衰竭。事实上,严重受影响的患者显示出显著的炎性响应,这可能另外引起dic并引起外周微血栓,包括在肺泡毛细血管中,这导致受影响血管中的微血栓。实施方案的硫酸葡聚糖已经显示出抗炎性,并且因此可以引起受试者中任何细胞因子风暴和过度炎症以及这种过度炎症的有害后果的消退或至少减少(图17a、17d、18a、19-23)。硫酸葡聚糖还具有抗凝血效果,这将有助于对抗dic和在严重受影响受试者中看到的微血栓(图14)。
72.就作用于免疫系统中的特定细胞和选择性减少这些细胞释放的促炎细胞因子而言,硫酸葡聚糖的抗炎作用是选择性的。更详细地说,本文提供的实验数据显示,硫酸葡聚糖特别靶向单核细胞和t淋巴细胞,并且引起活化单核细胞的il-6、il-10、il-1β、il-8、tnfα和ifnγ以及活化t淋巴细胞的il-6、il-10、tnfα和ifnγ的浓度依赖性和显著减少(图17-23)。与例如地塞米松和其他类固醇相比,硫酸葡聚糖选择性减少促炎细胞因子的一个显著优点是硫酸葡聚糖不会影响免疫系统产生的所有细胞因子,并且不会将细胞因子产生减少到非常低的水平。在感染性疾病,诸如冠状病毒感染中,为了对抗感染,需要免疫系统的受控活化。实施方案的硫酸葡聚糖可通过减少(与关闭形成鲜明对比)来自免疫系统的选定细胞的选定促炎性细胞因子来实现这种受控活化,从而减少发展过度炎性病况(诸如ards、sirs和脓毒性休克)的风险,同时仍允许免疫系统对抗冠状病毒感染。
73.硫酸葡聚糖相对于基于抗体的炎性治疗的优点在于,硫酸葡聚糖不像这种抗体那样靶向单一促炎细胞因子,而是降低几种关键促炎细胞因子的水平。另一优点是与抗体相比,硫酸葡聚糖在体内的半衰期相对较短(在人体中,c
max
为2至3小时),从而能够在与冠状病毒感染后的过度炎症风险期同步的明确限定的时间段期间实现受控的抗炎效果。
74.从covid-19恢复的受试者可能患有由ards、sirs和微血栓形成引起的器官纤维化。这种器官纤维化不仅包括肺纤维化,而且还可能包括其他器官,诸如肾纤维化和心肌病。实施方案的硫酸葡聚糖具有抗血栓形成作用,这在这种器官纤维化中是有用的(图15-16)。此外,实施方案的硫酸葡聚糖已经显示出不仅具有抗纤维化作用,而且具有纤维溶解作用,即能够去除器官纤维化后的现有疤痕并促进无疤痕组织重塑。
75.因此,实施方案的硫酸葡聚糖通过消退任何细胞因子风暴和过度炎症并通过具有抗纤维化和抗凝血作用,在感染的阶段iii(过度炎症期)具有医疗效果(图25)。
76.有新的证据表明,长期covid效应(诸如慢性疲劳综合征或肌痛性脑脊髓炎)由代谢失调引起。更详细地说,线粒体功能障碍和氧化/硝化应激见于长期covid患者中,包括atp产生改变和氧化/硝化应激增加。实施方案的硫酸葡聚糖能够通过使代谢和线粒体功能正常化来抵消这种代谢失调和氧化/硝化应激(图4a-4d,27-28;表5-8、15-18)。
77.实施方案的硫酸葡聚糖在恢复正常的线粒体相关能量代谢中是有效的,对三磷酸嘌呤和嘧啶核苷酸的浓度具有积极的作用。事实上,与健康对照相比,硫酸葡聚糖治疗几乎可以使受损组织中的atp水平和naa浓度正常化。
78.实施方案的硫酸葡聚糖引起氧化/硝化应激的显著降低。特别是,作为主要水溶性脑抗氧化剂的抗坏血酸和作为主要细胞内巯基(sh)供体的谷胱甘肽(gsh)的水平显著提高。此外,作为膜磷脂的多不饱和脂肪酸的终产物的丙二醛(mda)水平并因此作为活性氧物质(reactive oxygen species;ros)介导的脂质过氧化的标志物,在硫酸葡聚糖施用后显示出显著的降低。此外,受损组织中的亚硝酸盐/硝酸盐的总和也显著减少(图30)。上述氧化/硝化应激标志物都指示在硫酸葡聚糖治疗后,抗氧化/抗硝化状态的恢复有所改善(图29-31,33-36)。
79.通过具有抗血栓形成、抗纤维变性和纤维溶解作用,实施方案的硫酸葡聚糖在本发明的阶段iv(恢复期,图25)中也具有医疗效果。实施方案的硫酸葡聚糖还通过改善患者细胞的线粒体功能来诱导代谢正常化。
80.通过减少肌肉退化和改善肌肉功能(图38至42),实施方案的硫酸葡聚糖在阶段iv(恢复期,图25)中具有进一步的医疗效果。
81.有新的数据显示,covid-19,特别是长期covid-19,可能会引起肝脏损害。本文提供的实验数据显示,实施方案的硫酸葡聚糖对肝功能具有积极作用,并且能够使人类患者中受干扰的肝功能正常化(图43),这在阶段iv是有益的(图25)。
82.通过诱导肝细胞生长因子(hgf)的显著释放,实施方案的硫酸葡聚糖在恢复期(图25)也具有有益效果,所述肝细胞生长因子在器官再生和伤口愈合中起重要作用。通过诱导受sars-cov-2感染和sars-cov-2感染诱导的炎性响应负面影响的器官和组织的组织修复和再生以及伤口愈合,由实施方案的硫酸葡聚糖诱导的血浆hgf水平升高(图2、42)对covid-19患者有益。
83.本发明的一个方面涉及用于预防、抑制和/或治疗冠状病毒感染或感染性疾病的硫酸葡聚糖或其药学上可接受的盐。
84.在一个实施方案中,冠状病毒感染性疾病选自mers、sars和covid-19。在一个特定实施方案中,冠状病毒感染性疾病是covid-19。
85.在一个实施方案中,冠状病毒感染选自由冠状病毒引起的冠状病毒感染组成的组,所述冠状病毒选自由mers-cov、sars-cov和sars-cov-2组成的组。在一个特定实施方案中,冠状病毒感染由sars-cov-2引起。
86.本发明还涉及硫酸葡聚糖或其药学上可接受的盐在制造用于预防、抑制和/或治疗冠状病毒感染或感染性疾病的药物中的用途。
87.本发明进一步涉及预防、抑制和/或治疗冠状病毒感染或感染性疾病的方法。所述方法包括向患有冠状病毒感染或感染性疾病或具有患冠状病毒感染或感染性疾病风险的受试者施用有效量的硫酸葡聚糖或其药学上可接受的盐。
88.如本文中所使用的冠状病毒感染或感染性疾病的治疗不一定意味着冠状病毒感染或感染性疾病的治愈性治疗,还包括抑制或减轻冠状病毒感染或感染性疾病的短期和长期症状。因此,治疗还包括延迟冠状病毒感染或感染性疾病的发作,包括延迟、预防症状发作或消退与冠状病毒感染或感染性疾病相关的既定病理。
89.冠状病毒不是唯一使用细胞表面糖胺聚糖硫酸乙酰肝素(hspg)作为受体的病原体。形成鲜明对比的是,hspg充当几种病毒以及细菌和寄生虫病原体的受体,如下表1中所示(bartlett和woo park,heparan sulfate proteoglycans in infection,glycans in diseases and therapeutics 2011:第2章:31-62,m.s.g(编))。
90.表1-hspg-病原体相互作用
91.92.93.94.95.[0096][0097]
因此,实施方案的硫酸葡聚糖也可用于预防、抑制和/或治疗由能够结合细胞表面硫酸乙酰肝素蛋白聚糖(hspg),特别是能够结合hspg以促进初始细胞附着和/或后续细胞进入的病原体引起的病原体感染或感染性疾病。
[0098]
本发明的另一方面涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制和/或治疗由能够结合细胞表面硫酸乙酰肝素蛋白聚糖(hspg)的病原体引起的感染或感染性疾病。
[0099]
在一个特定实施方案中,病原体能够结合hspg以促进初始细胞附着和/或随后的细胞进入。
[0100]
在另一个特定的实施方案中,病原体能够结合hspg的硫酸乙酰肝素部分。
[0101]
在一个实施方案中,病原体是选自表1的细菌、病毒、朊病毒或寄生虫。在一个特定实施方案中,病原体不是人类免疫缺陷病毒(hiv),诸如不是hiv-1和hiv-2。
[0102]
在一个实施方案中,所述病原体选自由以下组成的组:炭疽杆菌(bacillus anthracis)、蜡样芽胞杆菌(bacillus cereus)、伯氏疏螺旋体(borrelia burgdorferi)、百日咳博德特氏菌(bordetella pertussis)、肺炎衣原体(chlamydia pneumoniae)、沙眼衣原体(chlamydia trachomatis)、流感嗜血杆菌(haemophilus influenzae)、幽门螺杆菌(helicobacter pylori)、单核细胞增多性李斯特菌(listeria monocytogenes)、结核分枝杆菌(mycobacterium tuberculosis)、淋病奈瑟氏菌(neisseria gonorrhoeae)、脑膜炎奈瑟氏菌(neisseria meningitides)、脑膜炎奈瑟氏菌(neisseria meningitides)、恙虫病东方体(orientia tsutsugamushi)、牙龈卟啉单胞菌(porphyromonas gingivalis)、绿脓假单胞菌(pseudomonas aeruginosa)、金黄色葡萄球菌(staphylococcus aureus)、无乳链球菌(streptococcus agalactiae)、化脓性链球菌(streptococcus pyogenes)、肺炎链球菌(streptococcus pneumoniae)和小肠结肠炎耶尔森氏菌(yersinia enterocolitica)。
[0103]
炭疽杆菌引起炭疽;蜡样芽胞杆菌引起恶心、呕吐和腹泻;伯氏疏螺旋体引起莱姆病(lyme disease);百日咳博德特氏菌引起百日咳;肺炎衣原体引起肺炎;沙眼衣原体引起衣原体;流感嗜血杆菌引起菌血症、肺炎、会厌炎、急性细菌性脑膜炎、蜂窝组织炎、骨髓炎和感染性关节炎;幽门螺杆菌引起胃炎和溃疡,但也与多种其他疾病有关,例如,特发性血小板减少性紫癜、缺铁性贫血、动脉粥样硬化、阿尔茨海默病(alzheimer's disease)、多发性硬化、冠状动脉疾病、牙周炎、帕金森病(parkinson's disease)、吉兰-巴雷综合征(guillain-barr
é
syndrome)、红斑痤疮、牛皮癣、慢性荨麻疹、斑秃、各种自身免疫性皮肤病、过敏性紫癜(henoch
‑‑
)、维生素b12低血浓度、自身免疫性中性粒细胞减少症、抗磷脂综合征、浆细胞恶血质、反应性关节炎、中心性浆液性脉络膜视网膜炎、开角型青光眼、睑炎、糖尿病、代谢综合征、各种类型的过敏、非酒精性脂肪性肝病、非酒精性脂肪性肝炎、肝纤维化和肝癌;单核细胞增多性李斯特菌引起李斯特菌病;结核分枝杆菌引起结核病;淋病奈瑟氏菌引起淋病;脑膜炎奈瑟氏菌引起脑膜炎和其他形式的脑膜炎球
菌疾病,诸如脑膜炎球菌血症,一种危及生命的败血症;恙虫病东方体引起恙虫病;牙龈卟啉单胞菌引起牙周疾病,以及上胃肠道、呼吸道和结肠中的牙周疾病,并且还与阿尔茨海默病和类风湿性关节炎有关;绿脓假单胞菌引起肺炎和各种败血病况;金黄色葡萄球菌引起皮肤感染,包括脓肿、呼吸道感染(诸如鼻窦炎)和食物中毒;无乳链球菌引起新生儿感染,包括新生儿感染败血症、肺炎和脑膜炎;化脓性链球菌引起皮肤感染、新生儿感染,但也可能导致风湿热、急性感染后肾小球肾炎和pandas;肺炎链球菌引起肺炎;并且小肠结肠炎耶尔森氏菌引起耶尔森氏菌病。
[0104]
在一个实施方案中,所述病原体是选自以下组成的组的病毒:腺相关病毒2型(aav2)、腺病毒、冠状病毒、柯萨奇病毒(coxsackievirus)、巨细胞病毒、登革热病毒(denv)、口蹄疫病毒(fmdv)、单纯疱疹病毒1(hsv-1)和hsv-2、乙型肝炎病毒、丙型肝炎病毒、人γ疱疹病毒8(hhv-8)(卡波济氏肉瘤相关疱疹病毒(kaposi's sarcoma-associated herpesvirus;kshv))、人免疫缺陷病毒1(hiv-1)、人乳头瘤病毒(hpv)、人类t细胞嗜淋巴病毒1型(htlv1)、日本脑炎病毒(jev)、伪狂犬病病毒、呼吸道合胞体病毒(rsv)、鼻病毒、辛德比斯病毒(sindbis virus;sinv)、痘苗病毒(vacv)、西尼罗病毒(west nile virus;wnv)和黄热病病毒(yellow fever virus)。
[0105]
在另一个实施方案中,病原体是选自由以下组成的组的病毒:aav2、腺病毒、冠状病毒、柯萨奇病毒、巨细胞病毒、denv、fmdv、hsv-1和hsv-2、乙型肝炎病毒、丙型肝炎病毒、hhv-8、hpv、htlv1、jev、伪狂犬病病毒、rsv、鼻病毒、sinv、vacv、wnv和黄热病毒。
[0106]
柯萨奇病毒引起无菌性脑膜炎;巨细胞病毒引起单核细胞增多症和肺炎;denv引起登革热;fmdv引起口蹄疫;hsv引起唇疱疹、生殖器疱疹和传染性;乙型肝炎病毒引起乙型肝炎;丙型肝炎病毒引起丙型肝炎、肝细胞癌和淋巴瘤;hhv-8引起卡普西肉瘤(kapsi’ssarcoma)、原发性渗出性淋巴瘤、hhv-8相关的多中心卡斯尔曼病和kshv炎性细胞因子综合征;hpv引起癌前病变、宫颈癌、hpv阳性口咽癌、生殖器疣和喉乳头状瘤病;htlv1引起成人t细胞淋巴瘤(atl)、htlv-i相关脊髓病、葡萄膜炎和粪类圆线虫高度感染;jev引起日本脑炎;伪狂犬病病毒引起奥耶斯基氏病(aujeszky’s disease);rsv引起呼吸道感染,包括细支气管炎和肺炎;鼻病毒引起普通感冒;sinv引起辛德比斯热(sindbis fever);wnv引起西尼罗河热(west nile fever);并且黄热病病毒引起黄热病。
[0107]
在一个实施方案中,病原体是选自由以下组成的组的寄生虫:蓝氏贾第鞭毛虫(giardia lamblia)、利什曼原虫属某些种(leishmania spp.)、脑胞内原虫属某些种(encephalitozoon spp.)、犬新孢子虫(neospora caninum)、疟原虫属某些种(plasmodium spp.)、刚地弓形虫(toxoplasma gondii)和克氏锥虫(trypanosoma cruzi)。
[0108]
蓝氏贾第鞭毛虫引起贾第鞭毛虫病;利什曼原虫属引起利什曼病;脑胞内原虫属引起微孢子虫病;犬新孢子虫引起受感染畜的自然流产;疟原虫属引起疟疾;刚地弓形虫引起弓形虫病;并且克氏锥虫引起人类的恰加斯病(chagas disease)、马的马媾疫(dourine)和苏拉病(surra),以及牛的布鲁氏菌病样疾病(brucellosis-like disease)。
[0109]
本发明还涉及硫酸葡聚糖或其药学上可接受的盐在制造用于预防、抑制和/或治疗由能够结合细胞表面hspg的病原体引起的感染或感染性疾病的药物中的用途。
[0110]
本发明进一步涉及预防、抑制和/或治疗由能够结合细胞表面hspg的病原体引起的感染或感染性疾病的方法。所述方法包括向患有由能够结合细胞表面hspg的病原体引起
的感染或感染性疾病或具有患有由能够结合细胞表面hspg的病原体引起的感染或感染性疾病的风险的受试者施用有效量的硫酸葡聚糖或其药学上可接受的盐。
[0111]
如本文中所使用,由能够结合细胞表面hspg的病原体引起的感染或感染性疾病的治疗不一定意味着对感染或感染性疾病的治愈性治疗,还包括抑制或减轻症状或消退与感染或感染性疾病相关的既定病理。因此,治疗还包括延迟感染或感染性疾病的发作,包括延迟与感染或感染性疾病相关的症状的发作。
[0112]
ards是一种呼吸衰竭,其特征是肺部中广泛炎症的快速发作。ards可能由败血症、胰腺炎、外伤、肺炎和误吸引起。潜在的机制包括对形成肺的微小气囊屏障的细胞的弥漫性损伤、表面活性剂功能障碍、免疫系统的活化和身体调节凝血的功能障碍。实际上,ards损害了肺交换氧气和二氧化碳的能力。成人诊断的依据是pao2/fio2比率(动脉氧分压与吸入氧分数的比率)小于300mm hg,尽管呼气末正压(peep)大于5cm h2o。
[0113]
sirs是一种影响全身的炎性状态。这是身体对感染性或非感染性损伤的响应。sirs经常因一个或多个器官或器官系统的衰竭而变得复杂,并且可能导致急性肾损伤、休克和多器官功能障碍综合征。
[0114]
sirs是一种与全身炎症、器官功能障碍和器官衰竭相关的严重病况。其是细胞因子风暴的子集,其中存在各种细胞因子的异常调节。sirs还与败血症密切相关,在败血症中,患者满足sirs的标准,并且具有疑似或证实的感染。sirs的表现包括体温低于36℃或高于38℃,心率高于每分钟90次,呼吸急促(呼吸频率高),每分钟呼吸20次以上;或者,动脉二氧化碳分压小于4.3kpa(32mmhg),而白细胞计数小于4000细胞/立方毫米(4
×
109细胞/升)或大于12,000细胞/立方毫米(12
×
109细胞/升);或存在超过10%的未成熟中性粒细胞(带状)。当这些标准中的两个或更多个在有或没有感染迹象的情况下得到满足时,患者可能被诊断患有sirs。患有sirs和急性器官功能障碍的患者可被称为严重sirs。
[0115]
实施方案的硫酸葡聚糖已经显示出抗炎性,并且因此可引起受试者中与ards和sirs相关的任何细胞因子风暴和炎症的消退或至少减少(图17a、17d、18a、19-23)。
[0116]
就作用于免疫系统中的特定细胞和选择性减少这些细胞释放的促炎细胞因子而言,硫酸葡聚糖的抗炎作用是选择性的。更详细地说,本文提供的实验数据显示,硫酸葡聚糖特别靶向单核细胞和t淋巴细胞,并且引起活化单核细胞的il-6、il-10、il-1β、il-8、tnfα和ifnγ以及活化t淋巴细胞的il-6、il-10、tnfα和ifnγ的浓度依赖性和显著减少(图17-23)。与例如地塞米松和其他类固醇相比,硫酸葡聚糖减少促炎细胞因子的一个显著优点是硫酸葡聚糖不完全关闭细胞因子,并且不影响免疫系统产生的所有细胞因子。在感染性疾病,诸如冠状病毒感染中,为了对抗感染,需要免疫系统的受控活化。实施方案的硫酸葡聚糖可通过减少(与关闭来自免疫系统的选定细胞的选定促炎性细胞因子形成鲜明对比)降低发展ards和sirs的风险来实现这种受控活化。
[0117]
本发明的另一方面涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制或治疗选自由ards和sirs组成的组的炎性疾病。
[0118]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐用于预防、抑制或治疗ards。
[0119]
在另一个实施方案中,硫酸葡聚糖或其药学上可接受的盐用于预防、抑制或治疗sirs。
[0120]
在一个特定实施方案中,炎性疾病由感染引起,诸如冠状病毒感染或由另一种hspg结合病原体引起的感染。
[0121]
本发明还涉及硫酸葡聚糖或其药学上可接受的盐在制造用于预防、抑制和/或治疗选自由ards和sirs组成的组的炎性疾病的药物中的用途。
[0122]
本发明进一步涉及用于预防、抑制和/或治疗选自由ards和sirs组成的组的炎性疾病的方法。所述方法包括向患有选自由ards和sirs组成的组的炎性疾病或具有患有炎性疾病风险的受试者施用有效量的硫酸葡聚糖或其药学上可接受的盐。
[0123]
如本文中所使用的选自由ards和sirs组成的组的炎性疾病的治疗不一定意味着炎性疾病的治愈性治疗,还包括抑制或减轻炎性疾病的短期和长期症状。因此,治疗还包括延迟感染或感染性疾病的发作,包括延迟与炎性疾病相关的症状的发作,以及消退长期的病理学问题,诸如纤维增生、肌病和在这种疾病之后出现的慢性疲劳病况。
[0124]
本发明还涉及硫酸葡聚糖或其药学上可接受的盐,其用于预防、抑制或治疗由sars-cov-2感染引起的长期效应covid-19症状。
[0125]
长期效应covid-19症状包括器官损害,特别是对受试者的心脏、肝脏和肺部的损害。从covid-19康复后几个月进行的成像测试显示,即使在只经历了轻微covid-19症状的人身上,心肌也受到了持久的损害。此外,covid-19可能对肺部中的小气囊(肺泡)造成长期损害。由此产生的疤痕组织可能导致长期的呼吸问题。很大一部分住院的covid-19患者肝功能异常。常见的长期症状主要是乏力、气短、咳嗽、心悸和嗅觉受损。其他症状包括胸痛、肌肉和关节痛、体重减轻和胃肠道病症。已报道的长期效应covid-19患者的临床指标包括肺功能受损和肺部损害、心血管效应(诸如心肌炎症)、大脑变化以及嗅觉和味觉受损(嗅觉缺失和嗅觉减退)。
[0126]
本发明还涉及硫酸葡聚糖或其药学上可接受的盐在制造用于预防、抑制和/或治疗由sars-cov-2感染引起的长期效应covid-19症状的药物中的用途。
[0127]
本发明进一步涉及用于预防、抑制和/或治疗由sars-cov-2感染引起的长期效应covid-19症状的方法。所述方法包括向患有由sars-cov-2感染引起的长期效应covid-19症状或具有患有由sars-cov-2感染引起的长期效应covid-19症状的风险的受试者施用有效量的硫酸葡聚糖或其药学上可接受的盐。
[0128]
在下文中,对硫酸葡聚糖的(平均)分子量和硫含量的提及也适用于硫酸葡聚糖的任何药学上可接受的盐。因此,硫酸葡聚糖的药学上可接受的盐优选具有如以下实施方案中讨论的平均分子量和硫含量。
[0129]
在实施方案的优选范围之外的硫酸葡聚糖被认为具有较差的效果且/或对细胞或受试者造成负面的副作用。
[0130]
例如,与具有较低平均分子量的硫酸葡聚糖相比,分子量超过10,000da(10kda)的硫酸葡聚糖通常具有较低的作用与副作用曲线。这意味着与平均分子量在优选范围内的硫酸葡聚糖分子相比,对于较大的硫酸葡聚糖分子(》10,000da),可安全地施用于受试者的硫酸葡聚糖的最大剂量较低。因此,当要将硫酸葡聚糖在体内施用于受试者时,此类较大的硫酸葡聚糖分子不太适合临床应用。
[0131]
硫酸葡聚糖是硫酸化多糖,并且特别地硫酸化葡聚糖,即由许多葡萄糖分子组成的多糖。如本文所定义的平均分子量表示单个硫酸化多糖可具有不同于所述平均分子量的
分子量,但平均分子量表示硫酸化多糖的平均分子量。这进一步意味着硫酸葡聚糖样品的所述平均分子量周围将存在分子量的自然分布。
[0132]
通常使用间接方法(诸如凝胶排斥/渗透色谱、光散射或粘度法)确定硫酸葡聚糖的平均分子量,或更准确地重均分子量(mw)。使用此类间接方法确定平均分子量将取决于许多因素,包括柱和洗脱液的选择、流速、校准程序等。
[0133]
重均分子量(mw):通常用于对分子大小而不是数值敏感的方法,例如光散射和尺寸排阻色谱(sec)方法。如果假设为正态分布,则mw每侧的重量相同,即分子量低于mw的样品中硫酸葡聚糖分子的总重量等于分子量高于mw的样品中硫酸葡聚糖分子的总重量。参数ni表示样品或批次中分子量为mi的硫酸葡聚糖分子的数量。
[0134]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw等于或低于10,000da。在一个特定实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw在2,000da至10,000da的区间内。
[0135]
在另一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw在2,500da至10,000da的区间内,优选在3,000da至10,000da的区间内。在一个特定实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw在3,500da至9,500da的区间内,诸如在3,500da至8,000da的区间内。
[0136]
在另一个特定的实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw在4,500da至7,500da的区间内,例如在4,500da至6,500da的区间内或在4,500da至5,500da的区间内。
[0137]
因此,在一些实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw等于或低于10,000da、等于或低于9,500da、等于或低于9,000da、等于或低于8,500da、等于或低于8,000da、等于或低于7,500da、等于或低于7,000da、等于或低于6,500da、等于或低于6,000da或等于或低于5,500da。
[0138]
在一些实施方案中,硫酸葡聚糖或其药学上可接受的盐的mw等于或高于1,000da、等于或高于1,500da、等于或高于2,000da、等于或高于2,500da、等于或高于3,000da、等于或高于3,500da、等于或高于4,000da或等于或高于4,500da。这些实施方案中的任一个可与定义mw上限的上述实施方案中的任一个组合,诸如与等于或低于10,000da的上限组合。
[0139]
在一个特定实施方案中,如上所述的硫酸葡聚糖或其药学上可接受的盐的mw是平均mw,并且优选地通过凝胶排阻/渗透色谱、尺寸排阻色谱、光散射或基于粘度的方法确定。
[0140]
数均分子量(mn):通常通过端基测定(例如核磁共振(nmr)光谱或色谱法)得出。如果假设为正态分布,则可发现mn每侧的硫酸葡聚糖分子的数量相同,即分子量低于mn的样品中硫酸葡聚糖分子的数量等于分子量高于mn的样品中硫酸葡聚糖分子的数量。
[0141]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的如通过nmr光谱测量的mn在1,850da至3,500da的区间内。
[0142]
在一个特定实施方案中,硫酸葡聚糖或其药学上可接受的盐的如通过nmr光谱测量的mn在1,850da至2,500da的区间内,优选在1,850da至2,300da的区间内,诸如在1,850da至2,000da的区间内。
[0143]
因此,在一些实施方案中,硫酸葡聚糖或其药学上可接受的盐的mn等于或低于3,500da、等于或低于3,250da、等于或低于3,000da、等于或低于2,750da、等于或低于2,500da、等于或低于2,250da或等于或低于2,000da。另外,硫酸葡聚糖或其药学上可接受的盐的mn等于或高于1,850da。
[0144]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的每个葡萄糖单元的平均磷酸根数在2.5至3.0的区间内。
[0145]
在一个特定实施方案中,硫酸葡聚糖或其药学上可接受的盐的每个葡萄糖单元的平均磷酸根数在2.5至2.8的区间内,优选在2.6至2.7的区间内。
[0146]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的平均葡萄糖单元数在4.0至6.0的区间内。
[0147]
在一个特定实施方案中,硫酸葡聚糖或其药学上可接受的盐的平均葡萄糖单元数在4.5至5.5的区间内,优选在5.0至5.2的区间内。
[0148]
在一个实施方案中,硫酸葡聚糖或其药学上可接受的盐的如通过nmr光谱测量的mn在1,850da至3,500da的区间内,每个葡萄糖单元的平均硫酸根数在2.5至3.0的区间内,并且硫酸葡聚糖的葡萄糖单元中c2位置的平均硫酸化为至少90%。
[0149]
在一个实施方案中,硫酸葡聚糖的平均葡萄糖单元数为约5.1,每个葡萄糖单元的平均硫酸根数在2.6至2.7的区间内,并且mn在1,850da至2,000da的区间内。
[0150]
在一个实施方案中,硫酸葡聚糖的药学上可接受的盐是硫酸葡聚糖的钠盐。在一个特定实施方案中,硫酸葡聚糖的钠盐的平均葡萄糖单元数为约5.1,每个葡萄糖单元的平均硫酸根数在2.6至2.7的区间内,并且包括na
抗衡离子的mn在2,100da至2,300da的区间内。
[0151]
在一个实施方案中,硫酸葡聚糖的平均葡萄糖单元数为5.1、每个葡萄糖单元的平均硫酸根数为2.7、如通过nmr光谱测量的不含na
的平均mn为约1,900da至1,950da,并且如通过nmr光谱测量的含有na
的平均mn为约2,200da至2,250da。
[0152]
根据实施方案的硫酸葡聚糖可作为硫酸葡聚糖的药学上可接受的盐诸如钠盐或钾盐提供。
[0153]
wo 2016/076780中公开了根据实施方案的当前优选的硫酸葡聚糖。
[0154]
受试者优选地是哺乳动物受试者,更优选灵长类动物,并且特别地人受试者。然而,硫酸葡聚糖或其药学上可接受的盐也可用于兽医应用。动物受试者的非限制性实例包括灵长类动物、猫、狗、猪、马、小鼠、大鼠。
[0155]
硫酸葡聚糖或其药学上可接受的盐优选通过注射施用于受试者,并且特别地通过静脉内(i.v.)注射、皮下(s.c.)注射或(i.p.)腹膜内注射,优选i.v.或s.c.注射。可使用的其他胃肠外施用途径包括肌肉内和关节内注射。硫酸葡聚糖或其药学上可接受的衍生物的注射可替代地或另外地直接发生在例如受试者体内将发生靶向效应的组织或器官或其他部位。
[0156]
实施方案的硫酸葡聚糖或其药学上可接受的盐优选地与选定的溶剂或赋形剂配制为水性注射溶液。溶剂有利地为水性溶剂,并且特别地缓冲溶液。这种缓冲溶液的非限制性实例是柠檬酸缓冲液,诸如柠檬酸水合物(cam)缓冲液,或磷酸盐缓冲液。例如,实施方案的硫酸葡聚糖可溶解于盐水诸如0.9%nacl盐水中,然后任选地用75mm cam缓冲并使用氢
氧化钠将ph调节至约5.9。非缓冲溶液也是可能的,包括水性注射溶液,诸如盐水,即nacl(水溶液)。此外,如果需要缓冲溶液,可使用除cam之外的其他缓冲系统。
[0157]
实施方案不限于注射,并且可替代地使用其他施用途径,包括经口、经鼻、经口、经直肠、经皮、经气管、经支气管或局部。然后用基于特定施用途径选择的合适赋形剂或媒介物配制活性化合物硫酸葡聚糖。
[0158]
硫酸葡聚糖或其药学上可接受的盐的合适剂量范围可以根据应用而变化,诸如体外相比于体内、受试者的大小和体重、受试者治疗的病况以及其他考虑因素。特别是对于人受试者,可能的剂量范围可为1μg/kg至100mg/kg体重,优选10μg/kg至50mg/kg体重。
[0159]
在优选的实施方案中,将硫酸葡聚糖或其药学上可接受的盐配制成以0.05mg/kg至50mg/kg受试者体重、优选0.05mg/kg或0.1mg/kg至40mg/kg受试者体重、并且更优选0.05mg/kg或0.1mg/kg至30mg/kg、或0.1mg/kg至25mg/kg、或0.1mg/kg至15mg/kg、或0.1mg/kg至10mg/kg受试者体重范围内的剂量施用。优选的剂量在0.25mg/kg至5mg/kg,优选0.5mg/kg至2.5mg/kg,更优选0.75mg/kg至2mg/kg受试者体重的范围内选择。
[0160]
硫酸葡聚糖或其药学上可接受的衍生物可以单次施用来施用,诸如以单次推注注射的形式施用。此推注剂量可相当快地注射到受试者,但有利地随着时间的推移而输注,使得硫酸葡聚糖溶液在几分钟内输注到患者,诸如在5至10分钟期间。
[0161]
替代地,硫酸葡聚糖或其药学上可接受的盐可在治疗期间多次(即至少两次)施用。
[0162]
硫酸葡聚糖或其药学上可接受的盐可与其他活性剂一起依次、同时或以包含硫酸葡聚糖或其药学上可接受的盐和至少一种其他活性剂的组合物的形式施用。至少一种活性剂可选自可用于上述疾病、病症或病况中的任一种的任何药剂。至少一种活性剂还可以是细胞疗法中的细胞形式,诸如干细胞,包括但不限于胚胎干细胞(esc)和间充质基质细胞(msc)。
[0163]
实施例
[0164]
在以下实施例中,使用了硫酸葡聚糖的钠盐,在本文中称为低分子量硫酸葡聚糖(lmw-ds)(tikomed ab,viken,sweden,wo 2016/076780)。
[0165]
实施例1
[0166]
附着于宿主组织是冠状病毒入侵和传播的关键步骤。因此,破坏宿主-病原体蛋白质-蛋白质相互作用可能是抑制冠状病毒入侵的有效方式。
[0167]
当前的研究调查了lmw-ds(tikomed ab,viken,sweden,wo 2016/076780)抑制寡聚β-淀粉样蛋白与病原体蛋白prpc之间的蛋白质-蛋白质相互作用的能力,试图揭示其抑制蛋白质-蛋白质相互作用的潜力。
[0168]
材料与方法
[0169]
化学品和抗体
[0170]
链霉亲和素hrp来自biolegend;β-淀粉样蛋白-(1-42)-生物素来自innovagen;正常人细胞朊病毒蛋白(prpc)来自merck;tmb来自ebioscience;1,1,1,3,3,3-六氟-2-丙醇(hfip)来自sigma;抗淀粉样蛋白β抗体克隆6e10来自biolegend;抗小鼠hrp来自cellsignaling;硫酸葡聚糖钠盐(dsss)(m.w.》500,000da)来自sigma;葡聚糖(m.w.450,000-650,000da)来自sigma;maxisorp板来自sigma。
[0171]
淀粉样蛋白-β寡聚体的制备
[0172]
β-淀粉样蛋白的寡聚化是基于以前的方法(stine等人,methods mol.biol.2011,670:13-32;aimi等人,jneurochem.2015,134:611-617)优化的。简言之,将淀粉样蛋白-β溶解在hfip中至1.0mm的最终浓度,使其经受保护性超声处理,并且小心蒸发hfip。将产生的肽膜储存在-20℃℃的密封容器中。在使用前,将肽膜缓慢溶解在dmso中至5.0mm的最终浓度并使其经受10分钟的保护性超声处理。为了制备寡聚体,将dmso溶液在冰冷的dmem培养基中稀释至100μm的最终浓度,并且在37℃(β-淀粉样蛋白-生物素)下孵育16小时。为了制备单体,将dmso溶液在冰冷的18mohm水中稀释至100μm的最终浓度并立即使用。
[0173]
淀粉样蛋白-β单体和寡聚体的鉴定
[0174]
将优化用于产生淀粉样蛋白-β的单体或寡聚体的制剂溶解在含有5%sds的非还原性凝胶样品缓冲液中。使用非还原mes运行缓冲液在15%bis-tris凝胶上运行蛋白质。将凝胶转移至pvdf,在10%脱脂牛奶中阻断,然后与抗淀粉样蛋白-β抗体在4℃下一起孵育过夜,并且用抗小鼠hrp显影,随后用ecl显影,并且暴露于胶片。
[0175]
定量寡聚淀粉样蛋白-β与prpc之间的蛋白质-蛋白质相互作用的elisa方法
[0176]
将prpc在碳酸盐包被缓冲液中稀释至10
×
的包被量(在100μl中;每孔500ng prpc的最终量)并施加至maxisorp板。然后将板密封并在4℃下放置过夜。在pbs-tween中仔细洗涤包被的板并用pbs中的2%bsa阻断。洗涤板,并且在添加到每个孔之前,将100μl寡聚淀粉样蛋白-β-生物素肽制剂(终浓度200nm)与测试化合物小心混合。将板在室温下孵育60分钟,洗涤并用链霉亲和素-hrp处理,并在进一步洗涤后使用tmb显色(用2n h2so4终止反应)。在30分钟内在450nm处读取吸光度。
[0177]
所有条件均一式三份进行。淀粉样蛋白-β-生物素与prpc的结合如aimi等人,j neurochem.2015,134:611-617所述进行计算。
[0178]
曲线拟合
[0179]
通过对浮动四参数逻辑方程的迭代曲线拟合来进行定量药理学分析。
[0180]
结果
[0181]
dsss像lmw-ds一样以浓度依赖性方式竞争寡聚淀粉样蛋白-β和prpc之间的蛋白质-蛋白质相互作用(图1;表2)。定量药理学分析指示,lmw-ds显示出与dsss相当的总体亲和力,然而在并列的可竞争结合水平和希尔系数方面的明显差异表明了两种化合物之间的不同相互作用(图1;表2)。与dsss和lmw-ds相反,葡聚糖未能明显竞争寡聚淀粉样蛋白-β与prpc之间的蛋白质-蛋白质相互作用。
[0182]
表2-竞争淀粉样蛋白-β与prpc之间的蛋白质-蛋白质相互作用能力的定量药理学分析
[0183]
化合物可竞争结合(%)ic
50
(μg/ml)希尔系数dsss101
±
20.62
±
0.071.51
±
0.06lmw-ds85
±
40.42
±
0.161.00
±
0.21
[0184]
讨论
[0185]
以前曾报道过高分子量硫酸葡聚糖(dsss)在低μg/ml范围内的有效浓度下与寡聚淀粉样蛋白-β和prpc之间的蛋白质-蛋白质相互作用竞争(aimi等人,j neurochem.2015,134:611-617)。在本研究中,相对于aimi等人的研究,方法的优化导致产生了明显更大比例
的寡聚淀粉样蛋白-β。蛋白质-蛋白质相互作用elisa的优化导致更大程度的特异性蛋白质-蛋白质相互作用;更大的竞争动态范围有助于对竞争化合物的相互作用进行定量药理学分析。因此,本研究比aimi等人报道的研究有所改进。
[0186]
dsss和lmw-ds显示出相当的亲和力来竞争寡聚淀粉样蛋白-β与prpc之间的蛋白质-蛋白质相互作用,分别产生0.62
±
0.07和0.42
±
0.16μg/ml的ic
50
值。竞争性质的希尔分析指示,与dsss相关的相对较高的希尔系数相比,lmw-ds显示出较浅的竞争曲线,这为dsss与lmw-ds之间不同的药理作用提供了证据。
[0187]
因此,lmw-ds竞争寡聚淀粉样蛋白-β与prpc之间的蛋白质-蛋白质相互作用,并从而可用于预防或至少抑制蛋白质-蛋白质相互作用。如用用lmw-ds观察到的这种效应在涉及蛋白质-蛋白质相互作用的冠状病毒感染和冠状病毒感染性疾病中具有潜力。因此,lmw-ds具有破坏sars-cov-2三聚体刺突蛋白与ace2相互作用的潜力,这在实施例13中得到进一步证实。
[0188]
实施例2
[0189]
本实施例研究了lmw-ds在增强als患者血液中组织修复生长因子的释放和活化方面的潜力。
[0190]
材料与方法
[0191]
这是一项单中心、开放单组研究,其中评价了als患者皮下(s.c.)施用lmw-ds的安全性、耐受性和功效。临床共有10次计划就诊:1次筛查就诊分为2个部分(就诊1a和就诊1b),5次研究医药产品(imp)施用就诊(就诊2至就诊6)和3次随访就诊(就诊7至就诊9)。每位单独患者的研究参与计划为约4个月,包括筛查和随访就诊。
[0192]
imp的活性药物成分是低分子量硫酸葡聚糖(lmw-ds)(tikomed ab,viken,sweden,wo 2016/076780)。所述药物用作s.c.注射溶液并由20mg/ml lmw-ds和9mg/ml nacl组成。每个玻璃小瓶含有10ml。施用剂量取决于第一次lmw-ds施用前就诊2时患者的体重。在腹部、大腿或臀部的交替侧(按优先顺序)皮下注射lmw-ds。施用1mg/kg的五次注射,给药间隔1周。通过商业elisa(r&d
·
hgf elisa)从每次就诊时采集的血样中测量免疫反应性hgf的变化。
[0193]
结果
[0194]
lmw-ds诱导als患者体内的血浆hgf水平显著升高,在lmw-ds施用后约2小时达到最大峰值(图2,表3),在lmw-ds施用后6小时血浆水平降低但仍显著升高(表3)。
[0195]
表3-lmw-ds指数增加als患者中的血浆hgf
[0196][0197]
相较于lmw-ds施用之前,*p《0.01
[0198]
讨论
[0199]
hgf(有时被称为散射因子(sf))是一种旁分泌细胞生长、运动和形态发生因子。已经证明其在胚胎器官发育以及还有成人器官再生和伤口愈合中具有主要作用。
[0200]
由lmw-ds诱导的血浆hgf水平的升高将通过诱导受sars-cov-2感染和由sars-cov-2感染诱导的炎性响应(包括ards、sirs和器官纤维化)负面影响的器官和组织的组织修复和再生以及伤口愈合而有益于covid-19患者。
[0201]
实施例3
[0202]
通过冷冻脑样品的高效液相色谱(hplc)分析,评估了在大鼠严重创伤性脑损伤(stbi)后每日皮下注射lmw-ds对谷氨酸兴奋性毒性和线粒体功能的影响。结果表明,lmw-ds干扰线粒体功能,以改善能量代谢,并且也降低了谷氨酸兴奋毒性。
[0203]
材料和方法
[0204]
stbi的诱导和药物施用方案
[0205]
根据动物护理的国际标准和指南,本研究中使用的实验方案由罗马天主教大学伦理委员会(ethical committee of the catholic university of rome)批准。300-350g体重(b.w.)的雄性wistar大鼠在受控环境中随意喂食标准实验室饮食和水。
[0206]
其被分成三组:
[0207]
1)n=6只经受stbi的动物,30分钟后进行药物施用并在tbi后2天处死(急性期1)
[0208]
2)n=6只经受stbi的动物,30分钟后进行药物施用并在tbi后7天处死(急性期2)。
[0209]
3)n=6只经受stbi的动物,3天后进行药物施用并在tbi后7天处死(慢性期)。
[0210]
作为麻醉混合物,动物通过i.p.注射接受35mg/kg b.w.氯胺酮(ketamine)和0.25mg/kg b.w.咪达唑仑(midazolam)。根据“重量下降”撞击加速模型,通过将450g重量从2m高度下降到已经由预先固定在颅骨上的金属盘保护的大鼠头部来诱导stbi(marmarou等人,j neurosurg.1994;80:291-300)。患有颅骨骨折、癫痫发作、鼻出血或未能幸免于撞击的大鼠被排除在研究之外。在每个治疗期结束时,再次麻醉大鼠,然后立即处死。
[0211]
药物治疗是皮下注射0.5ml的lmw-ds(tikomed ab,viken,sweden,wo 2016/076780;15mg/kg)并根据前述示意性方案施用。
[0212]
脑组织处理
[0213]
在麻醉期间,在所有动物中进行体内颅骨切除术,在小心地取出大鼠的颅骨后,暴
露脑并用外科刮刀取出脑,并且迅速投入液氮中。在湿重(w.w.)测定后,组织制备如先前公开的那样受到影响(tavazzi等人,neurosurgery.2005;56:582-589;vagnozzi等人,neurosurgery.2007;61:379-388;tavazzi等人,neurosurgery.2007;61:390-395;amorini等人,,j cell mol med.2017;21:530-542.)。简而言之,用7ml冰冷的氮饱和沉淀溶液进行全脑匀化,所述沉淀溶液由ch3cn 10mm kh2po4,ph 7.40(3:1;v:v)构成,并且使用设置为24,000rpm/min的ultra-turrax(janke&kunkel,staufen,germany)。在4℃下以20,690
×
g离心10分钟后,保存澄清的上清液,用3ml沉淀溶液补充沉淀,并且如上所述再次均化。进行第二次离心(20,690
×
g,4℃下10分钟),保存沉淀,将上清液与先前获得的上清液合并,通过剧烈搅拌用两倍体积的hplc级chcl3提取并如上离心。收集含有水溶性低分子量化合物的上层水相,使其经受氯仿洗涤两次以上(这个程序允许从缓冲的组织提取物中去除所有的有机溶剂和任何脂溶性化合物),用10mm kh2po4,ph 7.40调节体积,最终得到含水的10%组织匀浆并保存在-80℃下直至分析。
[0214]
嘌呤-嘧啶代谢物的hplc分析
[0215]
将每个去蛋白组织样品的等分试样通过0.45μm hv millipore过滤器过滤,并且装载(200μl)到hypersil c-18,250
×
4.6mm,5μm粒径柱上,所述柱配备有其自身的保护柱(thermo fisher scientific,rodano,milan,italy)并连接到由具有高度灵敏的二极管阵列检测器(配备有5cm光路流通池)并设置在200nm波长和300nm波长之间的surveyor系统(thermo fisher scientific,rodano,milan,italy)组成的hplc装置。数据采集和分析由pc使用hplc制造商提供的软件包进行。
[0216]
根据对现有离子配对hplc方法的轻微修改,在单次色谱运行中分离了属于嘌呤-嘧啶曲线(如下所列)并与组织能量状态、线粒体功能和氧化-硝化应激相关的代谢物(lazzarino等人,anal biochem.2003;322:51-59;tavazzi等人,clin biochem.2005;38:997-1008)。通过将保留时间、吸收光谱和峰面积与新制备的具有已知浓度的超纯标准混合物的色谱运行峰面积进行比较,在适当的波长(206nm、234nm和260nm)下对组织提取物的色谱运行中的感兴趣化合物进行分配和计算。
[0217]
化合物的列表:胞嘧啶、肌酸酐、尿嘧啶、β-假尿苷、胞苷、次黄嘌呤、鸟嘌呤、黄嘌呤、胞苷二磷酸胆碱(cdp-胆碱)、抗坏血酸、尿苷、腺嘌呤、亚硝酸盐(-no
2-)、还原型谷胱甘肽(gsh)、肌苷、尿酸、鸟苷、胞苷一磷酸(cmp)、丙二醛(mda)、胸苷、乳清酸、硝酸盐(-no
3-)、尿苷一磷酸(ump)、烟酰胺腺嘌呤二核苷酸、氧化型(nad
)、腺苷(ado)、肌苷一磷酸(imp)、鸟苷一磷酸(gmp)、尿苷二磷酸-葡萄糖(udp-glc)、udp-半乳糖(udp-gal)、氧化型谷胱甘肽(gssg)、udp-n-乙酰基-葡糖胺(udp-glcnac)、udp-n-乙酰基-半乳糖胺(udp-galnac)、腺苷一磷酸(amp)、鸟苷二磷酸-葡萄糖(gdp-葡萄糖)、胞苷二磷酸(cdp)、udp、gdp、烟酰胺腺嘌呤二核苷酸磷酸、氧化型(nadp
)、腺苷二磷酸-核糖(adp-核糖)、胞苷三磷酸(ctp)、adp、尿苷三磷酸(utp)、鸟苷三磷酸(gtp)、烟酰胺腺嘌呤二核苷酸、还原型(nadh)、腺苷三磷酸(atp)、烟酰胺腺嘌呤二核苷酸磷酸、还原型(nadph)、丙二酰基-coa、辅酶a(coa-sh)、乙酰基-coa、n-乙酰基天冬氨酸(naa)。
[0218]
游离氨基酸和含氨基化合物的hplc分析
[0219]
使用邻苯二甲醛(opa)和3-巯基丙酸(mpa)的混合物对样品进行柱前衍生来执行主要游离氨基酸(faa)和含氨基的化合物(agcc)(如下所列)的同步测定,如其他地方所详
细描述(amorini等人,j cell mol med.2017;21:530-542;amorini等人,mol cell biochem.2012;359:205-216)。简而言之,每天制备由25mmol/l opa、1%mpa、237.5mmol/l硼酸钠(ph 9.8)构成的衍生混合物并置于自动进样器中。在24℃下用opa-mpa对样品(15μl)进行自动柱前衍生,并且将25μl衍生混合物装载到hplc柱(hypersil c-18,250
×
4.6mm,5μm粒径,恒温在21℃)上以用于随后的色谱分离。在谷氨酸的情况下,在衍生程序和随后的注射之前,用hplc级h2o将去蛋白的脑提取物稀释20倍。opa-aa和opa-agcc的分离以1.2ml/min的流动速率使用两种流动相(流动相a=24mmol/l ch3coona 24mmol/l na2hpo4 1%四氢呋喃 0.1%三氟乙酸,ph 6.5;流动相b=40%ch3oh 30%ch3cn 30%h2o),使用适当的阶梯梯度进行(morini等人,j cell mol med.2017;21:530-542;amorini等人,mol cell biochem.2012;359:205-216)。
[0220]
通过将保留时间和峰面积与新鲜制备的具有已知浓度的超纯标准混合物的色谱运行的峰面积进行比较,在338nm波长下对全脑提取物的色谱运行中的opa-aa和opa-agcc进行分配和计算。
[0221]
faa和acgc化合物的列表:天冬氨酸(asp)、谷氨酸(glu)、天冬酰胺(asn)、丝氨酸(ser)、谷氨酰胺(gln)、组氨酸(his)、甘氨酸(gly)、苏氨酸(thr)、氨瓜氨酸(citr)、精酸(arg)、丙氨酸(ala)、牛磺酸(tau)、γ-氨基丁酸(gaba)、酪氨酸(tyr)、s-腺苷高半胱氨酸(sah)、l-胱硫醚(l-cysat)、缬氨酸(val)、蛋氨酸(met)、色氨酸(trp)、苯丙氨酸(phe)、异亮氨酸(ile)、亮氨酸(leu)、鸟氨酸(orn)、赖氨酸(lys)。
[0222]
统计学分析
[0223]
使用科尔莫戈罗夫-斯米尔诺夫检验(kolmogorov-smirnov test)对正态数据分布进行检验。对于重复测量,通过双向anova估计组间差异。fisher保护的最小二乘法被用作事后检验。只有小于0.05的双尾p值被认为统计上显著的。
[0224]
结果
[0225]
在24种标准和非标准氨基酸和含伯氨基的化合物的脑值中,最明显的结果是lmw-ds治疗对stbi诱导的谷氨酸(glu)的增加具有显著的抑制作用(图3),因此这种化合物过量必然导致兴奋性降低。
[0226]
然而,这种效果只有在损伤后早期(stbi后30分钟)施用时才可见,当在stbi后3天注射lmw-ds时,这种兴奋毒性标志物无效。还值得强调的是,lmw-ds对参与所谓的甲基循环(met、l-cysat、sah)的化合物具有显著的有益作用,见表4。
[0227]
表4-大脑化合物的浓度
[0228]
[0229]
[0230][0231]
a p《0.01(与对照相比),
b p《0.05(与对照相比),
c p《0.01(与tbi 2天相比),
d p《0.05(与tbi 2天相比),
e p《0.01(与tbi 5天相比),
f p《0.05(与tbi 5天相比),
g p《0.01(与急性期2相比),
h p《0.05(与急性期2相比),
i p《0.01(与慢性期相比),
j p《0.05(与慢性期相比)
[0232]
表3列出了以μmol/g(w.w.)计的化合物
[0233]
如表5中所示,lmw-ds积极影响与能量代谢和线粒体功能相关的各种化合物。特别令人感兴趣的是作为线粒体磷酸化能力测量的腺嘌呤核苷酸浓度和atp/adp比率(图4a-4d)。
[0234]
表5-能量代谢物的浓度
[0235]
[0236]
[0237]
[0238][0239]
a p《0.01(与对照相比),
b p《0.05(与对照相比),
c p《0.01(与tbi 2天相比),
d p《0.05(与tbi 2天相比),
e p《0.01(与tbi 5天相比),
f p《0.05(与tbi 5天相比),
g p《0.01(与急性期2相比),
h p《0.05(与急性期2相比),ip《0.01(与慢性期相比),
j p《0.05(与慢性期相比)
[0240]
表4列出了以nmol/g(w.w.)计的化合物
[0241]
还观察到氧化型和还原型烟碱辅酶的显著变化(图5a-5d)。
[0242]
还测量了与氧化应激相关的参数,并且在施用lmw-ds后检测到氧化应激的显著降低。特别是,对作为主要水溶性脑抗氧化剂的抗坏血酸和作为主要细胞内sh供体的gsh进行了测量。结果显示,如表5和图6a-6c中所示,在施用lmw-ds后,其水平显著提高。
[0243]
此外,还测量了mda,其作为膜磷脂的多不饱和脂肪酸的终产物,并且因此作为ros介导的脂质过氧化的标志物。在施用lmw-ds后,mda水平显示显著降低。上述氧化应激标志物都指示用lmw-ds治疗后抗氧化状态的恢复的改善(图6a-6c)。
[0244]
还分析了no介导的硝化应激(亚硝酸盐和硝酸盐)的代表性指标。lmw-ds施用显著降低了stbi急性期和慢性期的硝酸盐浓度(图7)。
[0245]
naa是一种脑特异性代谢物,并且是监测tbi后恶化或恢复的一种有价值的生化标志物。naa在神经元中由天冬氨酸和乙酰基-coa通过天冬氨酸n-乙酰转移酶合成。为了确保naa周转,分子必须在细胞区室之间移动以到达少突胶质细胞,在那里被天冬酰化酶(aspa)降解为乙酸盐和天冬氨酸盐。分解代谢酶aspa的上调和为提供底物天冬氨酸和乙酰基-coa的有效性而出现的naa下降是代谢障碍状态的标志。在这个研究中,在stbi后测量了naa和其底物,并且显示了在lmw-ds施用后水平的显著改善(图8a-8c)。
[0246]
当动物在损伤后早期(30分钟)接受lmw-ds施用时,这些对能量代谢物的影响特别明显。值得注意的是,无论是在stbi后2天处死动物,还是在stbi后7天进行处死,都可以观察到lmw-ds的总体有益效果。在这些动物组中,与agcc和能量代谢物相关的代谢的总体改善更加明显,从而表明lmw-ds施用对脑代谢具有长期的积极作用。
[0247]
讨论
[0248]
本文提供的数据表明,早期施用lmw-ds降低了谷氨酸兴奋毒性水平,并且通过保护线粒体功能改善了代谢体内稳态的不利变化。
[0249]
更详细地说,lmw-ds保护线粒体功能并减少氧化应激,如尤其在改善抗氧化状态的恢复、通过保持atp产生和代谢保护线粒体atp能量供应、线粒体磷酸化能力的正常化(均由lmw-ds诱导)中所见。因此,lmw-ds能够保护和保持暴露于损害或疾病的细胞中的线粒体功能,这对于具有可对抗感染性疾病的功能细胞是重要的。lmw-ds诱导的代谢正常化在肺期(阶段ii)和恢复期(阶段iv)对covid-19患者都是有益的(图25)。
[0250]
实施例4
[0251]
本研究的目的是评价lmw-ds对大鼠闭合性弥漫性重度tbi(stbi)实验模型产生的生化、分子和组织解剖学损害的潜在神经保护作用。在本研究中,通过对低分子量代谢物(代表能量代谢、氧化/硝化应激、抗氧化剂和治疗动物脑组织提取物中的游离氨基酸)的hplc分析获得结果。
[0252]
材料和方法
[0253]
stbi的诱导和药物施用方案
[0254]
在本研究中使用300-350g体重的雄性wis tar大鼠(n=160)。其在受控的环境中随意喂食标准的实验室饮食和水。
[0255]
作为可接受的麻醉混合物,动物通过肌肉注射接受35mg/kg体重的氯胺酮和0.25mg/kg体重的咪达唑仑。根据marmarou等人j.neurosurg.1994,80:291-300建立的“重量下降”撞击加速度模型诱导弥漫性stbi。这个模型导致弥漫性轴索损伤,并且能够再现人类弥漫性tbi的物理和机械特征。
[0256]
通过将450g的重物从2米高处掉落到由头盔(预先使用牙科粘固剂固定在颅骨上的金属盘)保护的大鼠头上以将机械力均匀地分布到大脑来诱导严重的tbi。使老鼠俯卧在一个特殊容器中的特殊聚氨酯泡沫床上;这种泡沫耗散了大部分势能(来自机械力),并且防止动物在撞击后的任何反弹,这种反弹可能导致脊椎损害。
[0257]
患有颅骨骨折、癫痫发作、鼻出血或未能幸免于撞击的大鼠被排除在研究之外。tbi诱导后2天或7天,再次麻醉大鼠,然后立即处死。这些时间点与最严重的生化紊乱(2天)一致,或在轻度脑损伤的情况下,与完全代谢恢复(7天)一致。
[0258]
药物治疗包括皮下注射0.5ml的lmw-ds(tikomed,viken,sweden,wo 2016/076780),并且根据下述方案以3种不同浓度(1mg/kg、5mg/kg和15mg/kg体重)施用。假手术动物进行相同的麻醉程序,但tbi和用作对照组。
[0259]
实验设计
[0260]
将本研究中使用的大鼠分成4组,以便在tbi后的两个不同时间进行三种不同浓度的lmw-ds的功效研究。如随后所述,根据下述程序,每组中均有经受特定代谢分析处理的动物和旨在进行组织形态学研究的其他动物。
[0261]
组-1
[0262]
专用于生化评价的对照(n=12)。另外四只动物用于组织形态学研究。本组大鼠总数:n=16
[0263]
组-2
[0264]
将经受stbi的不进行药物治疗的大鼠分为以下亚组:
[0265]
1. 12只动物经受stbi并在tbi后2天处死
[0266]
2. 12只动物经受stbi并在tbi后7天处死
[0267]
每个亚组再用四只大鼠进行组织形态学研究。本组大鼠总数:n=32。
[0268]
组-3
[0269]
使大鼠经受stbi并在tbi后30分钟后接受lmw-ds的单次施用,在tbi后2天处死。将动物分为以下亚组:
[0270]
1. 12只动物经受stbi并以1mg/kg b.w.治疗lmw-ds
[0271]
2. 12只动物经受stbi并以5mg/kg b.w.治疗lmw-ds
[0272]
3. 12只动物经受stbi并以15mg/kg b.w.治疗lmw-ds
[0273]
每个亚组再用四只大鼠进行组织形态学研究。本组大鼠总数:n=48。
[0274]
组-4
[0275]
使大鼠经受stbi并在tbi后30分钟后接受lmw-ds的单次施用,在tbi后7天处死。将动物分为以下亚组:
[0276]
1. 12只动物经受stbi并以1mg/kg b.w.治疗lmw-ds
[0277]
2. 12只动物经受stbi并以5mg/kg b.w.治疗lmw-ds
[0278]
3. 12只动物经受stbi并以15mg/kg b.w.治疗lmw-ds
[0279]
每个亚组再用四只大鼠进行组织形态学研究。本组大鼠总数:n=48。
[0280]
组-5
[0281]
使大鼠(n=12)经受stbi并在tbi后30分钟、3天和5天重复施用最大剂量的lmw-ds(15mg/kg b.w.),在tbi后7天处死。另外四只大鼠用于组织形态学研究。本组大鼠总数:n=16
[0282]
用于生化和基因表达分析的脑组织处理
[0283]
为了最小化代谢物损失,在麻醉期间对所有动物进行了体内颅骨切除术。小心地取出大鼠颅骨,暴露大脑,沿矢状裂急剧切开,分离两个半球。专用于生化分析的半球由在液氮中预冷的铝舌冷冻夹紧,然后浸入液氮中。引入冷冻夹紧程序是为了加速组织的冷冻,从而最大限度地减少潜在的代谢物损失。
[0284]
将用于分子生物学分析的其余半球置于5-10体积的溶液(invitrogen life technologies)中,这是一种可稳定和保护rna免于降解的rna稳定溶液。脑样品在4℃下储存过夜,以使溶液完全渗透组织。
[0285]
用于代谢物分析的组织均化如下所述进行。在湿重(w.w.)测定后,将冷冻半球放入7ml冰冷的氮饱和沉淀溶液(1:10w/v)中,所述沉淀溶液由ch3cn 10mm kh2po4,ph 7.40(3:1;v:v)构成,并且使用设置为24,000rpm/min的ultra-turrax均化器(janke&kunkel,staufen,germany)进行均化。在4℃下以20,690
×
g离心10分钟后,保存澄清的上清液,用10mm kh2po4的等分试样补充沉淀,并且如上所述再次均化并在-20℃下保存过夜,以便从组
织中完全回收水相。进行第二次离心(20,690
×
g,在4℃下10min),通过用两倍体积的hplc级chcl3剧烈搅拌提取与先前获得的上清液合并的上清液并如上离心。收集上层水相(含有水溶性低分子量化合物),使其经受氯仿洗涤两次以上(这个程序允许从缓冲的组织提取物中去除所有的有机溶剂和任何脂溶性化合物),用10mm kh2po4,ph 7.40调节体积,最终得到含水的10%组织匀浆并保存在-80℃下直至分析。
[0286]
能量代谢物、抗氧化剂和氧化/硝化应激生物标志物的hplc分析
[0287]
将每个去蛋白组织样品的等分试样通过0.45μm hv millipore过滤器过滤,并且装载(200μl)到hypersil c-18,250
×
4.6mm,5μm粒径柱上,所述柱配备有其自身的保护柱(thermo fisher scientific,rodano,milan,italy)并连接到由具有高度灵敏的二极管阵列检测器(配备有5cm光路流通池)并设置在200nm波长和300nm波长之间的surveyor系统(thermo fisher scientific,rodano,milan,italy)组成的hplc装置。数据采集和分析由pc使用hplc制造商提供的软件包进行。
[0288]
根据先前对现有离子对hplc方法的轻微修改,在单次色谱运行中分离与组织能量状态、线粒体功能抗氧化剂和氧化/硝化应激的代表相关的代谢物(如下所列)(lazzarino等人,anal biochem.2003,322:51-59;tavazzi等人,clin biochem.2005,38:997-1008)。通过将保留时间、吸收光谱和峰面积与新制备的具有已知浓度的超纯标准混合物的色谱运行峰面积进行比较,在适当的波长(206nm、234nm和260nm)下对组织提取物的色谱运行中的感兴趣化合物进行分配和计算。
[0289]
化合物的列表:胞嘧啶、肌酸酐、尿嘧啶、β-假尿苷、胞苷、次黄嘌呤、鸟嘌呤、黄嘌呤、cdp-胆碱、抗坏血酸、尿苷、亚硝酸盐(no2)、还原型谷胱甘肽(gsh)、肌苷、尿酸、鸟苷、cmp、丙二醛(mda)、硝酸盐(no3)、ump、nad
、ado、imp、gmp、udp-葡萄糖(udp-glc)、udp-半乳糖(udp-gal)、udp-n-乙酰基-葡糖胺(udp-glcnac)、udp-n-乙酰基-半乳糖胺(udp-galnac)、amp、gdp-葡萄糖、udp、gdp、nadp
、adp-核糖、ctp、adp、utp、gtp、nadh、atp、nadph、丙二酰基-coa、辅酶a(coa-sh)、乙酰基-coa、n-乙酰基天冬氨酸(naa)。
[0290]
游离氨基酸和含氨基化合物的hplc分析
[0291]
使用邻苯二甲醛(opa)和巯基丙酸(mpa)的混合物对样品进行柱前衍生来执行主要游离氨基酸(faa)和含氨基的化合物(agcc)(如下所列)的同步测定,如其他地方所详细描述(amorini等人,j cell mol med.2017,21(3):530-542;amorini等人,mol cell biochem.2012;359:205-216)。简而言之,每天制备由25mmol/lopa、1%mpa、237.5mmol/l硼酸钠(ph 9.8)构成的衍生混合物并置于自动进样器中。在24℃下用opa-mpa对样品(15μl)进行自动柱前衍生,并且将25μl衍生混合物装载到hplc柱(hypersil c-18,250
×
4.6mm,5μm粒径,恒温在21℃)上以用于随后的色谱分离。为了正确定量谷氨酸,在衍生程序和随后的注射之前,用hplc级h2o将去蛋白的脑提取物稀释20倍。opa-aa和opa-agcc的分离以1.2ml/min的流动速率使用两种流动相(流动相a=24mmol/l ch3coona 24mmol/l na2hpo4 1%四氢呋喃 0.1%三氟乙酸,ph 6.5;流动相b=40%ch3oh 30ch3cn 30%h2o),使用适当的阶梯梯度进行。
[0292]
通过将保留时间和峰面积与新鲜制备的具有已知浓度的超纯标准混合物的色谱运行的峰面积进行比较,在338nm波长下对全脑提取物的色谱运行中的opa-aa和opa-agcc进行分配和计算。
[0293]
faa和agcc化合物的列表:天冬氨酸(asp)、谷氨酸(glu)、天冬酰胺(asn)、丝氨酸(ser)、谷氨酰胺(gln)、组氨酸(his)、甘氨酸(gly)、苏氨酸(thr)、瓜氨酸(citr)、精氨酸(arg)、丙氨酸(ala)、牛磺酸(tau)、γ-氨基丁酸(gaba)、酪氨酸(tyr)、s-腺苷高半胱氨酸(sah)、l-胱硫醚(l-cystat)、缬氨酸(val)、蛋氨酸(met)、色氨酸(trp)、苯丙氨酸(phe)、异亮氨酸(ile)、亮氨酸(leu)、鸟氨酸(orn)、赖氨酸(lys)。
[0294]
用于组织形态学分析的脑组织处理
[0295]
充分麻醉后,如(di pietro等人,sci rep.2017,7(1):9189)所述对大鼠进行经心脏灌注。简而言之,进行了胸廓切开术,并且将肝素溶液注入门静脉,以避免在整个手术过程中血液凝固。之后,进行右心房切口,并且将灌注针推进升主动脉。用100ml ph 7.4的磷酸盐缓冲溶液(pbs)进行灌注,以便在用ph 7.4的pbs溶液中的100ml 4%多聚甲醛(pfa)进一步灌注之前洗去血液。从颅骨快速取出后,通过在4℃下在pbs溶液中的4%pfa中浸泡2小时来后固定每个脑。通过在4℃下将全脑浸入富含增加的蔗糖溶液(10%、20%和30%)的pbs中24小时,然后在剥离型模具容器(agar scientific,essex,uk)中植入最佳切割温度包埋介质(oct)(thermo shandon,runcorn,uk)中获得冷冻保护。浸泡在oct中的大脑在-80℃下储存前在干冰碎冰中快速冷冻。
[0296]
统计学分析
[0297]
通过学生t检验(student’s t-test)来估计组间的差异。只有小于0.05的双尾p值被认为统计上显著的。
[0298]
结果
[0299]
stbi后2天记录的生化数据总结
[0300]
增加剂量的lmw-ds对所测量的脑能量代谢的影响
[0301]
表6总结了涉及磷酸化高能嘌呤和嘧啶化合物的值。特别明显的是由stbi引起的三磷酸核苷酸(atp、gtp、utp和ctp)的消耗,伴随着adp和udp-葡萄糖(udp-glcnac)和udp-半乳糖(udp-galnac)的n-乙酰化衍生物的增加。
[0302]
在损伤后的这个时候,用lmw-ds治疗在改善细胞能量代谢方面仅部分有效:在所有三种剂量的测试药物中,记录到明显更高的高能磷酸盐(atp、gtp和ctp)值。对utp和adp的浓度没有影响。值得回顾的是,大鼠tbi后48小时代表了脑代谢的关键时间点,与线粒体功能的最大变化一致,包括线粒体质量控制的变化。在tbi的这个实验模型中,这个时间点可以被认为是一种“转折点”,在这个点处定义大脑代谢的恢复或不恢复。
[0303]
表6-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在stbi后2天处死的大鼠的去蛋白脑匀浆中测量的能量代谢物(磷酸化嘌呤和嘧啶)的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0304][0305][0306]
在表6-25中,粗体指示与对照组有显著差异(p《0.05);粗体下划线指示与tbi有显著差异(p《0.05);并且粗斜体指示与对照组和tbi有显著差异(p《0.05)。
[0307]
增加剂量的lmw-ds对烟碱辅酶的影响
[0308]
表7总结了氧化型(nad
和nadp
)和还原型(nadh和nadph)烟碱辅酶的值。表7还报
道了nad
/nadh比率的计算的一维值,所述值适用于评价代谢在多大程度上依赖于糖酵解或线粒体氧化磷酸化。
[0309]
如本文中先前所观测,stbi引起nad
、nadp
和nad
/nadh比率的降低。在这个时间点,用lmw-ds治疗仅在最大测试剂量(15mg/kg b.w.)下有效,所述剂量产生了对烟碱辅酶库的显著保护,并且避免了向糖酵解的代谢转换,从而间接暗示了整体更好的线粒体功能。
[0310]
表7-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在stbi后2天处死的大鼠的去蛋白脑匀浆中测量的烟碱辅酶的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。nad
/nadh比率是一维的。
[0311][0312][0313]
增加剂量的lmw-ds对coa-sh衍生物的影响
[0314]
表8报道了涉及游离coa-sh和coa-sh衍生物的数据。特别是乙酰基-coa是线粒体代谢的关键化合物,从而允许三羧酸循环(tca循环)的正确功能,由此确保电子传递链(etc)的连续电子供应。tca是产生还原型辅酶(nadh和fadh2)的主要细胞周期,通过将其的电子分别转移到线粒体复合物i和ii,并且是etc和氧化代谢的燃料。所有的化合物(尤其是乙酰基-coa)都受stbi显著影响。当剂量为5mg/kg或15mg/kg b.w.时,观察到这种化合物的部分拯救作用。损伤后30分钟向动物施用lwm-ds。
[0315]
表8-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的游离coa-sh和coa-sh衍生物(乙酰基-coa和丙二酰基-coa)的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0316][0317]
增加剂量的lmw-ds对抗氧化剂和氧化/硝化应激生物标志物的影响
[0318]
表9显示了主要水溶性脑抗氧化剂(抗坏血酸和gsh)以及氧化(mda)和硝化应激的生物标志物(-no
2-和-no
3-)的浓度。丙二醛(mda)来源于膜磷脂的不饱和脂肪酸的分解,是ros介导的脂质过氧化的结果。亚硝酸盐(-no
2-)和硝酸盐(-no
3-)是一氧化氮(no)代谢的稳定终产物,在病理条件下,由一氧化氮合酶(inos)的可诱导形式过量产生并通过与ros反应产生活性氮物质(reactive nitrogen species;rns):
[0319]
在撞击后两天,在经历stbi的大鼠中,两种水溶性抗氧化剂均出现25%至45%的下降。还记录了氧化/硝化应激标签的相应增加。施用lwm-ds显著改善抗坏血酸和还原型谷胱甘肽(gsh)的浓度,并且显著降低脑组织亚硝酸盐和硝酸盐。当使用15mg kg/b.w.时,这些影响更加显著。
[0320]
表9-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的抗氧化剂和氧化/硝化应激生物标志物的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0321][0322]
增加剂量的lmw-ds对去磷酸化嘌呤和嘧啶的影响
[0323]
表10中报道的大多数化合物来源于嘌呤和嘧啶核苷酸的降解途径并与细胞能量代谢间接相关。接受stbi的大鼠大脑中所有这些化合物的浓度较高,但cdp-胆碱除外,其中大部分受到药物施用的积极影响。
[0324]
表10-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的去磷酸化的嘌呤和嘧啶的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0325][0326]
增加剂量的lmw-ds对n-乙酰基天冬氨酸(naa)的影响
[0327]
naa是哺乳动物大脑中最丰富的n-乙酰化氨基酸,其浓度几乎等于人类神经递质谷氨酸的浓度。尽管尚未完全阐明naa的生物学作用。先前在临床前和临床研究中已经显示,tbi降低naa浓度,并且其在头部损伤后的时程变化反映了atp的时程变化。特别是,stbi导致naa体内稳态的不可逆改变,naa是脑能量代谢的良好替代标志物,naa水平的降低和恢复比脑震荡后运动员的症状清除慢得多。因此,naa在tbi研究中具有特殊的意义。
[0328]
在撞击后两天,在stbi大鼠中观察到全脑naa降低40%(图9)。当以5mg/kg或15mg/kg b.w.施用时,lmw-ds对naa浓度产生有益的影响。尽管显著低于对照组,但施用两种药物剂量中任一种的大鼠的naa显著高于在stbi大鼠中发现的值,在接受最高剂量的lmw-ds的大鼠中发现最高的naa水平。
[0329]
增加剂量的lmw-ds对参与神经传递的游离氨基酸的影响
[0330]
表11中列出的化合物是直接(glu、gaba)或间接(gln、asp、asn、gly、ser、thr、ala)参与神经传递的氨基酸。特别是,glu是主要的兴奋性氨基酸,其作用被gaba抵消。谷氨酸的兴奋毒性由ser、gly、thr和ala调节,并且与涉及神经元和星形胶质细胞的glu-gln循环的功能有关。如先前的研究所示,我们在此发现,在stbi大鼠中,这些氨基酸中的大多数在损伤后两天增加。当药物以5mg/kg或15mg/kg b.w.皮下输注时,用单次施用lmw-ds治疗动物是部分有效的在大多数情况下,不同化合物的值明显好于在未治疗的stbi动物组中发现的值,但不优于对照组的值。
[0331]
表11-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的具有神经递质功能的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0332][0333][0334]
增加剂量的lmw-ds对参与甲基循环的游离氨基酸的影响
[0335]
表12中报道的游离氨基酸参与所谓的甲基循环,调节在细胞代谢中作为甲基供体的化合物的体内稳态,或参与半胱氨酸的形成,半胱氨酸是唯一具有游离-sh基团的氨基酸。严重的头部创伤引起这一重要代谢途径的主要参与者的显著变化。在任何测试剂量下,通过lwm-ds完成甲硫氨酸的恢复。药物治疗在使其他氨基酸正常化方面部分有效。在撞击后7天,将在相应的表中给出对l-胱硫醚(l-cystat)变化的评论。
[0336]
表12-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1、5和15mg/kg体重)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的参与甲基循环和-sh基团体内稳态的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动
物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0337][0338]
增加剂量的lmw-ds对参与一氧化氮(no)生成的游离氨基酸的影响
[0339]
表13说明了在由一氧化氮合酶(nos)催化的反应中,直接参与no生成的游离氨基酸的浓度,一氧化氮合酶是一个以三种同种型存在的酶家族:内皮nos(enos)、神经元nos(nnos)、诱导型nos(inos)。最后一种同种型(inos)是参与硝化应激的同种型。一氧化氮是通过复杂的反应生成,其中精氨酸(arg)提供进行部分氧化并形成瓜氨酸(citr)和no的氮原子。stbi后2天的动物显示arg的同时减少和citr的增加,与显示稳定的no终产物亚硝酸盐和硝酸盐增加的数据一致(表8)。当使用15mg/kg b.w.剂量时,lmw-ds的施用特别有效。
[0340]
表13-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/k、5mg/k和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的参与一氧化氮形成的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0341][0342]
增加剂量的lmw-ds对长链游离氨基酸的影响
[0343]
表14中报道的游离氨基酸代表可用于产生细胞用来补充tca循环的α-酮酸的碳骨架的来源。在这些化合物中,只有异亮氨酸(ile)受到stbi的显著影响并在接受药物治疗的大鼠中恢复。
[0344]
表14-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的长链游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0345][0346][0347]
增加剂量的lmw-ds对作为渗透物的游离氨基酸和芳香族游离氨基酸的影响
[0348]
表15中总结的结果清楚地显示,stbi引起所有这些游离氨基酸浓度的增加。特别是,牛磺酸(tau)的增加可能表明试图通过增加最重要的脑渗透物中的一种的水平来抵消细胞水肿。不同的是,芳香族游离氨基酸的增加可能表明神经递质血清素(由色氨酸形成)和多巴胺(首先由苯丙氨酸然后由酪氨酸的生物转化产生)的生物合成减少。在撞击后的这个时间点没有观察到lmw-ds施用的显著效果。
[0349]
表15-在脑外伤诱导后30分钟进行的不施用和单次施用增加剂量的lwm-ds(1mg/kg、5mg/kg和15mg/kg b.w.)的情况下,在tbi后2天处死的大鼠的去蛋白脑匀浆中测量的作为渗透物的游离氨基酸和芳香族游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0350][0351]
stbi后7天记录的生化数据总结
[0352]
增加剂量的lmw-ds对所测量的脑能量代谢的影响
[0353]
表16总结了涉及磷酸化高能嘌呤和嘧啶化合物的值。特别明显的是在stbi后7天没有观察到三磷酸核苷酸(atp、gtp、utp和ctp)消耗的改善。伴随amp和adp增加的是udp衍生物(udp-glc、udp-gal、udp-glcnac和udp-galnac)浓度的显著变化。总的来说,应强调的是,损伤后较长时间的特征通常是由stbi诱导的生化、代谢和分子改变的恶化。
[0354]
在损伤后的这个时间,用lmw-ds治疗产生了脑能量代谢的普遍改善,当药物施用剂量高于1mg/kg b.w.时更明显。尽管即使在接受15mg/kg b.w.lwm-ds重复施用的大鼠中
也记录到了与对照组的差异,但在药物治疗的动物中发现了显著更高的三磷酸核苷酸值。特别相关的是atp/adp比率的计算的一维值(其被认为是线粒体磷酸化能力的良好指标)的逐渐恢复,所述值通过增加施用于stbi动物的药物剂量而逐渐增加。
[0355]
表16-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的能量代谢物(磷酸化嘌呤和嘧啶)的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0356]
[0357][0358]
为了更好地显示药物效果与药物剂量相关,我们在图10中用图表报道了关于atp的结果。有可能观察到atp增加与施用剂量有某种关系,并且在任何测试剂量下,药物施用产生了最重要的高能磷酸盐的显著增加。
[0359]
增加剂量的lmw-ds对烟碱辅酶的影响
[0360]
表17总结了氧化型(nad
和nadp
)和还原型(nadh和nadph)烟碱辅酶的值。表17还报道了nad
/nadh比率的计算的一维值,所述值适用于评估代谢在多大程度上依赖于糖酵解或线粒体氧化磷酸化。
[0361]
如前所述,在损伤后7天,在stbi大鼠中记录了烟碱辅酶和nad
/nadh比率的显著下降。除最低剂量外,用lmw-ds治疗显著提高了烟碱辅酶的浓度。特别是,单次和重复施用15mg/kg b.w.lmw-ds能够使nad 水平正常化,并且恢复在对照动物中测定的正确的nad
/nadh比率。
[0362]
表17-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中
测量的烟碱辅酶的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0363][0364]
增加剂量的lmw-ds对coa-sh衍生物的影响
[0365]
表18报道了涉及游离coa-sh和coa-sh衍生物的数据。针对coa-sh和乙酰基-coa检测到5mg/kg或15mg/kg b.w.的施用(此剂量作为单次施用和重复施用)的显著积极效果,从而表明对于tca循环的功能更有利的代谢条件。
[0366]
表18-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的游离coa-sh和coa-sh衍生物(乙酰基-coa和丙二酰基-coa)的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0367][0368][0369]
增加剂量的lmw-ds对抗氧化剂和氧化/硝化应激生物标志物的影响
[0370]
表19显示了主要水溶性脑抗氧化剂(抗坏血酸和gsh)以及氧化(mda)和硝化应激的生物标志物(-no
2-和-no
3-)的浓度。在撞击后7天,在经历stbi的大鼠中,两种水溶性抗氧化剂的浓度都没有恢复。还记录了显著高水平的氧化/硝化应激标签。施用最高单次和重复
剂量的lwm-ds的效果特别有利于拯救抗坏血酸和还原型谷胱甘肽(gsh)的浓度,伴随脑组织亚硝酸盐和硝酸盐的明显降低。当使用5mg kg/b.w.时,这些影响也很显著。
[0371]
表19-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的抗氧化剂和氧化/硝化应激生物标志物的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0372][0373][0374]
为了更好地理解药物作用与药物剂量相关,我们在图11和12中用图表报道了关于抗坏血酸和gsh的结果。
[0375]
增加剂量的lmw-ds对去磷酸化嘌呤和嘧啶的影响
[0376]
在损伤后7天接受stbi的大鼠中,观察到表20中报道的大多数化合物的进一步恶化,这些化合物源于嘌呤和嘧啶核苷酸的降解途径,并且与细胞能量代谢间接相关。这些化合物中的大部分受到药物施用的积极影响。
[0377]
表20-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的去磷酸化嘌呤和嘧啶的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0378][0379][0380]
增加剂量的lmw-ds对n-乙酰基天冬氨酸(naa)的影响
[0381]
如前所述,stbi导致naa体内稳态的不可逆的改变。甚至在这个研究中,我们发现在stbi后7天,全脑naa比在对照大鼠中测量的低约50%,见图13。有趣的是,在接受增加剂量的单次lmw-ds或重复施用最大测试剂量的大鼠中,检测到naa的剂量依赖性增加。
[0382]
增加剂量的lmw-ds对参与神经传递的游离氨基酸的影响
[0383]
表21中列出的化合物是直接(glu、gaba)或间接(gln、asp、aasn、gly、ser、thr、ala)参与神经传递的氨基酸。与对照组相比,这些氨基酸中的大多数在损伤后7天在stbi大鼠中仍然较高。从这个表中可以明显看出,lmw-ds的施用是有效的,特别是当药物以15mg/kg b.w.皮下输注时,无论是单次施用还是重复施用。特别相关的是glu的正常化,因此表明
lmw-ds能够消除stbi后过量glu释放引起的兴奋性毒性。
[0384]
表21-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的具有神经递质功能的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0385][0386][0387]
增加剂量的lmw-ds对参与甲基循环的游离氨基酸的影响
[0388]
如表22中所示,当与对照组的相应值相比时,在撞击后7天,stbi大鼠中参与所谓甲基循环或半胱氨酸形成的游离氨基酸水平仍然不同。在接受最高剂量lwm-ds(单次施用或重复施用)的动物中观察到met增加。如在损伤后2天已经观察到,这些药物水平产生了l-胱硫醚(l-cysat)的显著增加。由于这种化合物是产生半胱氨酸(cys)的中间物,可以设想假设l-半胱氨酸的增加会导致cys的增加。值得一提的是,cys的测定需要特殊的额外hplc测定,并用f-moc进行衍生,f-moc是一种与仲胺和cys反应的荧光化合物。
[0389]
表22-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的参与甲基循环和-sh基团体内稳态的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0390][0391]
增加剂量的lmw-ds对参与一氧化氮(no)生成的游离氨基酸的影响
[0392]
表23显示了直接参与no生成的游离氨基酸的浓度。stbi后7天的动物仍显示arg减少,citr增加,与显示稳定的no终产物亚硝酸盐和硝酸盐增加的数据一致(表8)。当使用5mg/kg或15mg/kg b.w.(单次和重复)时,lmw-ds的施用特别有效。
[0393]
表23-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的参与一氧化氮形成的游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0394][0395]
增加剂量的lmw-ds对长链游离氨基酸的影响
[0396]
代表用于产生细胞用来补充tca循环的α-酮酸的碳骨架来源的表24中报道的游离氨基酸在stbi和用感兴趣的药物治疗的任何其他动物组后7天实际上是正常的。
[0397]
表24-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的长链游离氨基酸的浓度。对照组以假手术动物为代表。数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0398][0399][0400]
增加剂量的lmw-ds对作为渗透物的游离氨基酸和芳香族游离氨基酸的影响
[0401]
表25中总结的结果清楚地显示,在损伤后7天,stbi引起牛磺酸(tau)浓度的增加。lmw-ds施用使tau浓度正常化,并且引起芳香族氨基酸的增加。
[0402]
表25-在不施用和施用增加剂量的lwm-ds(单次施用1mg/kg、5mg/kg和15mg/kg b.w.,并且重复施用15mg/kg b.w.)的情况下,在stbi后7天处死的大鼠的去蛋白脑匀浆中测量的作为渗透物的游离氨基酸和芳香族游离氨基酸的浓度。对照组以假手术动物为代表。
[0403]
数值为每组12只动物的平均值
±
s.d.,并且以nmol/g w.w.表示。
[0404][0405]
讨论
[0406]
为评价增加剂量的lmw-ds对在损伤后不同时间经历stbi的大鼠的大量脑代谢物的影响而进行的研究证明,施用这种化合物产生了脑代谢的总体改善。
[0407]
lmw-ds在恢复线粒体相关的能量代谢方面是有效的,在未经治疗的stbi动物中严重失衡,对三磷酸嘌呤和嘧啶核苷酸的浓度具有积极的作用。特别地,在撞击后7天,atp水平仅比对照组的值低16%,而在stbi大鼠中发现降低了35%(表16和图10)。值得注意的是,在同一时间点用lmw-ds治疗的动物中的naa浓度仅比对照组的值低16%,而stbi动物显示比这种化合物的值低48%。这一发现再次有力地证实了naa体内稳态与正确的线粒体能量代谢之间的严格联系,并且强调了能够对线粒体功能起积极作用的药物干预的重要性。
[0408]
由lmw-ds施用产生的脑代谢的总体改善还涉及烟碱辅酶和游离coa-sh和coa-sh衍生物的代谢。这意味着药物治疗的动物(尽管接受stbi)具有准正常的辅酶,以确保正确的氧化还原反应并允许tca循环的良好功能。
[0409]
上述脑代谢的改善肯定有助于另一个显著的药物作用,即消除glu兴奋毒性。此外,药物影响含硫氨基酸。可能地,这种效应可能与含有s原子的药物分子有关。增加此原子的生物利用率可能会导致这些氨基酸生物合成的净增加,其中一个氨基酸(met)在甲基化反应和所谓的甲基循环中是至关重要的。
[0410]
在这项研究中记录的进一步的积极作用是在接受lmw-ds施用的stbi大鼠中抗氧化剂的增加和氧化/硝化应激的生化标签的减少。甚至这种现象也可能与线粒体功能的正常化有关,因为功能障碍的线粒体是ros和rns的主要细胞内来源。相关的是,lmw-ds的效果在stbi后7天比2天更明显。这有力地表明,由药物施用引起的脑代谢的总体改善不是暂时现象。
[0411]
这个实验证实了lmw-ds在保护线粒体功能和减少氧化应激中的积极作用,如尤其在改善抗氧化状态的恢复、通过保持atp产生和代谢保护线粒体atp能量供应、线粒体磷酸化能力的正常化(均由lmw-ds诱导)中所见。因此,lmw-ds能够保护和保持暴露于损害或疾病的细胞中的线粒体功能,这对于具有可对抗感染性疾病的功能细胞是重要的。lmw-ds诱导的代谢正常化在肺期(阶段ii)和恢复期(阶段iv)对covid-19患者都是有益的(图25)。
[0412]
实施例5
[0413]
本实施例研究了lmw-ds作为als患者的抗凝剂和抗血栓剂的潜力。
[0414]
材料与方法
[0415]
这是一项单中心、开放单组研究,其中评价了als患者皮下(s.c.)施用lmw-ds的安全性、耐受性和功效。临床共有10次计划就诊:1次筛查就诊分为2个部分(就诊1a和就诊1b),5次imp施用就诊(就诊2至就诊6)和3次随访就诊(就诊7至就诊9)。每位单独患者的研究参与计划为约4个月,包括筛查和随访就诊。
[0416]
imp的活性药物成分是低分子量硫酸葡聚糖(lmw-ds)(tikomed ab,viken,sweden,wo 2016/076780)。将lmw-ds制备成s.c.注射用溶液,并且由20mg/ml lmw-ds和9mg/ml nacl组成。每个玻璃小瓶含有10ml。施用剂量取决于第一次lmw-ds施用前就诊2时患者的体重。在腹部、大腿或臀部的交替侧(按优先顺序)皮下注射lmw-ds。施用1mg/kg的五次注射,给药间隔1周。从每次就诊采集的血样中记录活化部分凝血活酶时间(aptt)的变化。
[0417]
结果
[0418]
在注射lmw-ds之前,基线时的平均aptt为26.5s。在每次lmw-ds治疗后,观察到短暂的增加,最大值出现在给药后2至2.5小时,相对于基线的相对变化分别为 33.5%和 26.5%。给药后6小时,aptt值恢复到给药前水平。
[0419]
讨论
[0420]
急性组织炎症升高凝血因子viii,导致血液中的aptt降低和组织中的微血栓形成。本实施例中提供的实验数据显示,lmw-ds增加了血液中的aptt,并从而用作抗凝血剂和抗血栓剂。这种抗凝血和抗血栓作用对处于过度炎症期(阶段iii)的covid-19受试者是有益的(图25)。
[0421]
实施例6
[0422]
在这项研究中,lmw-ds通过在diversity plus组中进行分析来表征。组由基于人类原代细胞的系统组成,旨在以体外格式模拟人体的不同方面。diversity plus组中的12个系统(表26)允许在模拟各种人类疾病状态的广泛系统中以公正的方式表征测试试剂。p系统由来自健康人类供体的一种或多种原代细胞类型构建而成,并添加刺激物,诸如细胞因子或生长因子,以捕获人体组织或病理条件下天然存在的相关信号网络。在th1(3c系统)和th2(4h系统)炎性环境中,以及在对动脉平滑肌细胞具有特异性的th1炎性状态(casm3c系统)中,对血管生物学进行建模。其他系统概括了系统免疫响应的各个方面,包括单核细胞驱动的th1炎症(lps系统)或t细胞刺激(sag系统)、巨噬细胞活化驱动的慢性th1炎症(lmphg系统)和生发中心发生的b细胞的t细胞依赖性活化(bt系统)。be3c系统(th1)和bf4t系统(th2)代表肺部的气道炎症,而myof系统模拟肌成纤维细胞-肺组织重塑。最后,皮肤生物学在kf3ct系统建模th1皮肤炎症和hdf3cgf系统建模伤口愈合中进行了阐述。
[0423]
每种测试试剂都生成根据单独系统环境中蛋白质生物标志物读数的变化而创建的标签曲线。生物标志物读数(每个系统7-17个)针对治疗和生物学相关性进行选择,可预测疾病结果或特定药物效果,并且使用具有已知作用机制(moa)的试剂进行验证。通过检测蛋白质的基于免疫的方法(例如elisa)或测量增殖和活力的功能性测定来定量测量每个读数。读数是多样的,并且包括细胞表面受体、细胞因子、趋化因子、基质分子和酶。总的来说,diversity plus组含有148个生物标志物读数,这些读数捕获在特定系统的生理环境中发生的生物变化。
[0424]
材料和方法
[0425]
四种浓度的lmw-ds(tikomed ab,viken,sweden,wo 2016/076780;150nm、440nm、1.3μm、4μm)在diversity plus组中由eurofins进行研究。
[0426]
用于多样性plus的方法
[0427]
biomap系统中的人原代细胞用于早期传代(第4代或更早),以最小化对细胞培养条件的适应并保持生理信号传导响应。所有细胞均来自多个供体(n=2-6)的集合,根据制造商的建议进行商业购买和处理。人血液衍生的cd14
单核细胞在添加到mphg系统之前在体外分化成巨噬细胞。使用的缩写如下:人脐静脉内皮细胞(huvec)、外周血单核细胞(pbmc)、人新生儿真皮成纤维细胞(hdfn)、b细胞受体(bcr)、t细胞受体(tcr)和toll样受体(tlr)。
[0428]
每个系统中使用的细胞类型和刺激物如下:3c系统[huvec (il-1β、tnfα和ifnγ)]、4h系统[huvec (il-4和组胺)]、lps系统[pbmc和huvec lps(tlr4配体)]、sag系统[pbmc和huvec tcr配体]、bt系统[cd19
b细胞和pbmc (α-igm和tcr配体)]、bf4t系统[支气管上皮细胞和hdfn (tnfα和il-4)]、be3c系统[支气管上皮细胞 (il-1β、tnfα和ifnγ)]、casm3c系统[冠状动脉平滑肌细胞 (il-1β、tnfα和ifnγ)]、hdf3cgf系统[hdfn (il-1β、tnfα、ifnγ、egf、bfgf和pdgf-bb)]、kf3ct系统[角质细胞和hdfn (il-1β、tnfα、ifnγ和tgfβ)]、myof系统[分化的肺肌成纤维细胞 (tnfα和tgfβ)]和mphg系统[huvec和m1巨噬细胞 酶原(tlr2配体)]。
[0429]
系统来源于单细胞类型或共培养系统。贴壁细胞类型在96或384孔板中培养直至汇合,然后添加pbmc(sag和lps系统)。bt系统由与pbmc共培养的cd19 b细胞组成,并且用bcr活化因子和低水平tcr刺激进行刺激。在dmso(小分子;最终浓度≤0.1%)或pbs(生物制剂)中制备的测试试剂在刺激前1小时以指定的浓度添加,并且保持在培养物中持续24小时或如另外所指示(48小时,myof系统;72小时,bt系统(可溶性读数);168小时,bt系统(分泌型igg))。每个板含有适用于每个系统的药物对照(例如,1.1μm的传统对照测试试剂秋水仙碱)、阴性对照(例如,非刺激条件)和媒介物对照(例如,0.1%dmso)。直接elisa用于测量细胞相关和细胞膜靶的生物标志物水平。使用检测、基于珠的多重免疫测定或捕获elisa对来自上清液的可溶性因子进行定量。通过对贴壁细胞进行磺基罗丹明b(srb)染色以及对悬浮液中的细胞进行还原来检测测试试剂对细胞增殖和活力的明显不利影响(细胞毒性)。对于增殖测定,在亚影响力下培养单独的细胞类型,并且在针对每个系统优化的时间点进行测量(48小时:3c和casm3c系统;72小时:bt和hdf3cgf系统;96小时:sag系统)。通过srb(24小时:3c、4h、lps、sag、bf4t、be3c、casm3c、hdf3cgf、kf3ct和mphg系统;48小时:myof系统),并且通过对悬浮液中的细胞进行alamarblue染色(24小时:sag系统;42小时:bt系统)来测量贴壁细胞的细胞毒性。
[0430]
数据分析
[0431]
将测试试剂处理的样品中的生物标志物测量值除以对照样品(来自同一板的至少6个媒介物对照)的平均值,以产生然后被log
10
变换的比率。在95%的置信区间,使用历史媒介物对照数据计算显著性预测包络线。
[0432]
曲线分析
[0433]
当2个或更多个连续浓度相对于媒介物对照在相同方向上变化、在显著性包络线之外并具有至少一个效应大小》20%的浓度(|log
10
比|》0.1)时,注释生物标志物活性。如果生物标志物的关键活性在一些系统中增加,但在其他系统中减少,则这些活性被描述为受调节的。当总蛋白水平下降超过50%(srb或alamarblue水平的log
10
比《-0.3)时,表示细胞毒性状况并用x轴上方的细黑箭头指示。当在3个或更多个系统中检测到细胞毒性时,认为化合物具有广泛的细胞毒性。具有可检测的广谱细胞毒性的测试试剂的浓度从生物标志物活性注释和下游基准、相似性搜索和聚类分析中排除。抗增殖作用由来自以较低密度铺板的细胞的srb或alamarblue log
10
比《-0.1来定义并由x轴上方的灰色箭头指示。细胞毒性和抗增殖箭头只需要一个浓度就能满足曲线注释的指示阈值。
[0434]
基准分析
[0435]
当两个曲线的读数在显著性包络线之外且在相同方向上的效应大小》20%时,注
释共同的生物标志物读数。当一个曲线的读数在效应大小》20%的显著性包络线之外,而另一个曲线的读数在包络线之内或在相反方向上时,注释分化生物标志物。除非特别说明,否则测试试剂和基准试剂的最高非细胞毒性浓度都包括在基准叠加分析中。
[0436]
相似性分析
[0437]
当两个曲线的读数在显著性包络线之外且在相同方向上的效应大小》20%时,注释共同的生物标志物读数。具有3个或更多个可检测细胞毒性系统的测试试剂浓度从相似性分析中排除。具有1-2个可检测细胞毒性系统的测试试剂浓度将连同与测试试剂最高浓度匹配的数据库覆盖一起包括在相似性搜索分析中。随后是下一个最高浓度的测试试剂的额外叠加,所述测试试剂不含具有可检测细胞毒性的系统,并且各自的数据库匹配。为了确定在diversity plus组中运行的化合物的曲线之间的相似程度,我们开发了一种定制的相似性度量(biomap z标准),这是一种组合方法,与其他测试方法(包括皮尔逊(pearson)和斯皮尔曼(spearman)的相关系数)相比,所述方法在参考试剂的机制分类中具有改进的性能。这种方法更有效地说明了数据点、系统、活性生物标志物读数的数量变化以及生物标志物读数变化的幅度,这些都是曲线的特征。首先产生皮尔逊相关系数(r)来测量两个曲线之间的线性关联,所述线性关联基于关系的方向和大小的相似性。由于皮尔逊相关性会受到任何生物标志物活性大小的影响,因此每个系统的加权平均tanimoto度量被用作过滤器,以解释不太稳固的系统的代表性不足。tanimoto度量不考虑生物标志物活性的幅度,但在每个系统的加权基础上解决了读数的属性和数量是否相同的问题。首先通过将每个曲线归一化为单位向量(例如,)来计算实值tanimoto度量,然后应用以下公式:其中a和b是2个曲线向量。然后,在此计算中将其并入系统加权平均实值tanimoto度量:所述计算使用每个i
th
系统(ti)的实值tanimoto评分和每个i
th
系统(wi)的权重。按以下公式计算每个系统的wi:其中lr是比较的2个曲线的比率的最大绝对值。基于参考化合物的最佳性能,如果皮尔逊相关系数(r)≥0.7,则将曲线鉴定为具有机制相关相似性。最后,使用fisher的r-to-z变换来计算z评分,以将短尾分布转换为正态分布,如下所示:然后,根据以下公式生成z标准,其针对公共读数(cr)的数量进行调整:较大的z标准值对应于较高的置信水平,并且这是用于对相似性结果进行分级的度量。
[0438]
聚类分析
[0439]
聚类分析(功能相似度图)使用成对相关分析的结果,以将试剂曲线的“接近度”从多维空间投影到二维空间。在这个分析过程中产生的试剂曲线的功能性聚类使用皮尔逊相关值,用于每种浓度下每种试剂的曲线的成对比较,然后使成对相关数据经受多维标度。皮尔逊相关系数(r)≥0.7的相似曲线由线连接。不相互聚类的试剂被解释为机制上不同的。针对测试了3种或更多种试剂的项目进行这个分析。从聚类分析中排除细胞毒性浓度。
[0440]
机制热图分析
[0441]
机制热图分析提供了测试化合物和19种共有机制的可视化,从而允许比较所有化合物浓度和共有机制的生物标志物活性。机理热图分析中使用的合成共有曲线是来自结构不同的化学类别的多种化合物的平均代表性曲线。通过对所有选择的曲线(不同浓度的多种试剂)的每个生物标志物终点的值进行平均来计算曲线,以建立共有机制曲线。当生物标志物活性相对于媒介物对照在显著性包络线之外表达时,其在共有机制和化合物的热图中被着色。红色代表增加的蛋白质表达,蓝色代表减少的表达,并且白色表示水平不变或在过滤条件内。相对于媒介物对照,较暗的颜色代表生物标志物活性的较大变化。使用r和r的gplots包来制备机制热图。
[0442]
测定接受标准
[0443]
测定包括由平台为组成diversity plus组的系统中测试的试剂生成的多参数数据集。测定含有适用于每个系统的药物对照(例如,传统对照测试试剂秋水仙碱)、阴性对照(例如,非刺激条件)和媒介物对照(例如,dmso)。测定是基板的,并且数据接受标准取决于板性能(媒介物对照孔的cv%)和所述系统历史对照的系统性能。qa/qc皮尔逊检验首先从历史阳性对照的参考数据集中建立1%的假阴性皮尔逊临界值。所述过程遍历阳性对照参考数据集中系统生物标志物读数的每个曲线,从而计算每个曲线与数据集中剩余曲线的平均值之间的皮尔逊值。用于确定1%假阴性临界值的皮尔逊值总数是参考数据集中存在的曲线总数。所有计算值的百分之一处的皮尔逊值是1%的假阴性皮尔逊临界值。如果实验板的阴性对照或药物对照曲线与参考数据集中的历史对照曲线的平均值之间的皮尔逊值超过1%的假阴性皮尔逊临界值,则系统通过。当每个单独的系统通过皮尔逊测试,并且所有项目板中有95%的cv%《20%时,接受整体测定。
[0444]
结果
[0445]
如表26中所示,diversity plus组包含12个单独的基于biomap人原代细胞的共培养系统。
[0446]
表26-diversity plus组
[0447]
[0448]
[0449][0450]
当两个或多个连续浓度相对于媒介物对照在相同方向上变化,在95%显著性包络线之外,并且具有至少一个效应大小》20%的浓度时,注释生物标志物活性(|log
10
比率|》0.1)。如果生物标志物关键活性在一些系统中增加,但在其他系统中减少,则这些活性被描述为受调节的。
[0451]
lmw-ds是活性的,具有25个带注释的读数。在本研究中测试的浓度下,lmw-ds对任何人原代细胞都没有细胞毒性。lmw-ds介导的关键生物标志物活性的变化包括炎症相关活性,表现为血管细胞粘附分子1(vcam-1)、单核细胞趋化蛋白-1(mcp-1)、可溶性肿瘤坏死因子α(stnfα)、干扰素诱导的t细胞α趋化因子(i-tac)、γ干扰素诱导的单核因子(mig)和γ干扰素诱导的蛋白10(ip-10)的减少以及嗜酸细胞活化趋化因子3(eot3)以及白细胞介素8(il-8)的增加。lmw-ds还具有免疫调节活性,表现为分泌型免疫球蛋白g(sigg)和巨噬细胞集落刺激因子(m-csf)的减少,以及可溶性il-17a(sil-17a)和分化簇69(cd69)的增加。lmw-ds还显示出组织重塑活性,表现为基质金属蛋白酶-1(mmp-1)、纤溶酶原活化因子抑制剂-1(pai-1)、尿激酶纤溶酶原活化因子受体(upar)和表皮生长因子受体(egfr)的增加,以及止血相关活性,表现为血栓调节蛋白(tm)的增加。表27总结了lmw-ds对diversity plus组中12种不同人原代细胞的影响。
[0452]
表27
–
diversity plus结果的总结
[0453][0454]
参考数据库含有》4,500种活性剂(生物制剂、批准的药物、化学品和实验试剂)的曲线,并且可用于分类和鉴定最相似的曲线。
[0455]
在从参考数据库中对数学上相似的化合物曲线进行的无监督搜索中,lmw-ds(4m)与克赛(clexane)(30μg/ml)最相似(皮尔逊相关系数,r=0.701)。克赛(依诺肝素钠(enoxaparin sodium))是一种低分子量肝素,其是用于治疗深静脉血栓形成(dvt)的抗凝剂。以下系统中注释了五种常见活性:bt(sigg、sil-17a)、casm3c(mig)和hdf3cgf(vcam-1、ip-10)。
[0456]
讨论
[0457]
在研究中,lmw-ds的特征在于在diversity plus组中对基于人原代细胞的测定进行了分析,所述测定对器官(脉管系统、免疫系统、皮肤、肺)的复杂组织和疾病生物学以及一般组织生物学进行了建模。diversity plus组评估了lmw-ds在保留与体内结果相关的复杂串扰和反馈机制的条件下的生物学影响。
[0458]
lmw-ds在本研究中测试的浓度下具有活性且无细胞毒性。lmw-ds仅在最高浓度(4μm)时对人原代内皮细胞具有适度和选择性的抗增殖作用。lmw-ds曲线具有25个带注释的读数,从而指示免疫和炎症相关读数以及基质相关生物标志物的调节。特定的活性包括减少的炎症相关stnfα、vcam-1、ip-10(cxcl10)、mig(cxcl9)、i-tac(cxcl11)和mcp-1,以及增加的il-8。在bf4t系统中仅在较低浓度下观察到适度增加的嗜酸细胞活化趋化因子-3。免疫调节活性包括bt系统中降低的sigg以及il-17a和il-17f,但对b细胞没有任何抗增殖作用。还鉴定出减少的m-csf,以及增加的cd69、sil-17a和sil-17f。lmw-ds也调节组织重塑生物标志物,包括增加的mmp-1、pai-1、upar、egfr和止血相关的tm。在casm3c和hdf3cgf系统中,包括mig、vcam、ip-10和itac的关键炎症生物标志物在所有测试浓度下均有所下降,而在多个系统中发现趋化因子il-8的增加。这些数据共同指示,lmw-ds在炎症和伤口愈合生物学背景下在调节免疫活化和/或免疫消退响应中起作用。
[0459]
炎症标志物的调节指示lmw-ds在治疗多种慢性和急性炎症和包括炎症组分的疾病中的用途。
[0460]
损伤后的最初阶段,先天/促炎响应和获得性免疫响应的选定成分被上调,以维持对外来病原体的防御,清除损伤部位处存在的组织碎片,并且协调与创伤响应相关的组织重塑、细胞增殖和血管生成过程。然而,为了进行适当的伤口愈合,必须调节或关闭这种初始炎性响应,以允许基质的重建、再细胞化和组织重塑。这种免疫消退活性由lmw-ds诱导,包括mmp-1、par-1和upar的活化,从而指示诱导的免疫消退可用于治疗在covid-19受损的组织,否则会导致有害的纤维化形成。
[0461]
lmw-ds调节了hdf3cgf系统中许多生物标志物的活性,但仅调节了myof系统中的il-8。两个系统都包括成纤维细胞,但hdf3cgf模拟伤口愈合和与伤口愈合相关的基质重塑,而myof更像是胶原沉积的纤维化模型。因此,结果指示lmw-ds具有免疫调节和组织重塑活性,但没有诱导不希望的胶原纤维变性,所述胶原纤维变性会导致有害的纤维变性沉积。
[0462]
总之,lmw-ds似乎能使创伤或疾病后组织中存在的炎症正常化和消退,因此这些结果与前述实施例中lmw-ds的作用一致。
[0463]
因此,lmw-ds调节促炎细胞因子和趋化因子的分泌,从而抑制疾病受损组织中的炎性信号传导。这些发现与抑制有害的炎性响应有关,包括ards和sirs以及感染性休克,如在受影响最严重的covid-19受试者中所见。此外,lmw-ds的组织重塑作用将有利于靶向covid-19受试者的肺纤维化、肾纤维化和心肌病。
[0464]
实施例7
[0465]
在细胞系中研究了由lmw-ds诱导的基因表达变化的分析。
[0466]
材料与方法
[0467]
实验设计
[0468]
对于每个细胞系,设置n=8
×
25cm2培养烧瓶。在处理当天(接种后24小时),对每种细胞类型收获两个烧瓶。这代表第0天的时间点。从剩余的烧瓶中,用对照培养基处理三个烧瓶,并用含有lmw-ds的培养基(cm)处理三个烧瓶,以得到0.01mg/ml的最终浓度。48小时后从处理过的烧瓶中收集细胞。因此,收集的数据代表(a)未处理的细胞(第0天对照和第2天对照)和(b)用lmw-ds(tikomed ab,viken,sweden,wo 2016/076780)处理48小时的细胞(第2天lmw-ds处理)。
[0469]
包被用于所有细胞的组织培养板
[0470]
通过在每个烧瓶中添加2ml的50μg/ml聚-d-赖氨酸于汉克氏平衡盐溶液(hank's balanced salt solution;hbss)中的溶液并在37℃下在黑暗中孵育过夜来包被25cm2烧瓶。用细胞培养水洗涤烧瓶,并且在黑暗中风干30min。通过向每个烧瓶中添加1ml的25μg/ml层粘连蛋白于磷酸盐缓冲盐水(pbs)中的溶液并在37℃下在黑暗中孵育2小时来包被烧瓶。在铺板细胞前,用pbs洗涤层粘连蛋白瓶三次。
[0471]
人脐静脉内皮细胞(huvec)
[0472]
制备培养基200 大血管内皮补充剂(m200 lves)添加剂(1:50)并预热至37℃。将细胞在37℃水浴中解冻不超过2分钟并轻轻转移到含有20ml杜尔贝克氏改良伊格尔培养基(dulbecco's modified eagle medium)、营养混合物f-12(dmem-f12)的50ml管中。通过小心翻转管两次来混合细胞悬浮液。将细胞以400
×
g旋转10分钟。去除上清液并将细胞重新
悬浮在10ml培养基(m200 lves添加剂)中。
[0473]
用细胞计数器计数细胞。将1,000,000个细胞/瓶接种在25cm2烧瓶中(n=8)并将培养基加满至每烧瓶总共5ml。将细胞在37℃下与5%co2一起孵育。在lmw-ds处理之前,使细胞沉降24小时。
[0474]
人施万细胞(human schwann cell)
[0475]
通过向高葡萄糖dmem中添加10%胎牛血清(fbs)来制备施万细胞生长培养基并将其预热至37℃。将细胞在37℃水浴中解冻不超过2分钟。
[0476]
将来自12个小瓶的细胞分别轻轻转移到含有10ml高葡萄糖dmem培养基的管中并在400相对离心场(rcf)下离心10分钟。将沉淀重新悬浮在培养基中。将来自12个小瓶的细胞混合并平均分配到预先包被的25cm2烧瓶中(n=8)。将细胞在37℃下与5%co2一起孵育。在lmw-ds处理之前,使细胞沉降24小时。
[0477]
小鼠皮质神经元(lonza)
[0478]
通过向500ml神经基础培养基中添加10ml b-27无血清补充剂和2.5ml glutamax
tm-i补充剂来制备培养基。将培养基预热至37℃。将来自12个小瓶的细胞在37℃水浴中依次解冻不超过2分钟并轻轻转移至15ml管中。向每个中缓慢逐滴添加9ml培养基。通过小心翻转管两次来混合细胞悬浮液。
[0479]
将细胞以200
×
g离心5分钟。去除上清液(至最后0.5ml),并且通过研磨将细胞缓慢重新悬浮。将来自12个小瓶的细胞混合并平均分配到预先包被的25cm2烧瓶中(n=8)。将细胞在37℃下与5%co2一起孵育24小时。
[0480]
小鼠运动神经元(aruna)
[0481]
根据表28制备培养基。
[0482]
表28-培养基的制备
[0483][0484]
使培养基(见表28)预热至37℃。将细胞在37℃水浴中解冻不超过2分钟。缓慢逐滴添加9ml培养基。通过小心翻转管两次来混合细胞悬浮液。用细胞计数器对细胞进行计数。将细胞以200
×
g离心5分钟。去除上清液(至最后0.5ml),并且通过研磨将细胞缓慢重新悬浮。将来自8个小瓶的细胞混合并平均分配到预先包被的25cm2烧瓶中(n=8)。在处理之前,将细胞在37℃下与5%co2一起孵育24小时。
[0485]
药物治疗
[0486]
lmw-ds以20mg/ml的储备浓度提供并保存在4℃的温度监控冰箱中。在无菌dmem-f12中制备新鲜的100
×
lmw-ds储备液(1.0mg/ml)。将浓缩的药物储备液无菌过滤并添加到各自的培养基中(19.6ml cm和0.4ml lmw-ds储备溶液)。使用19.6ml cm和0.4ml dmem-f12进行对照。将lmw-ds和cm添加到各自的烧瓶(每个烧瓶5ml)中,以使每个培养皿中lmw-ds的浓度达到0.01mg/ml,每个培养皿中总共有10ml cm。
[0487]
培养物收集和细胞裂解
[0488]
将cm吸到干净且有标签的15ml falcon管中。将烧瓶(无培养基)放入-80℃冰箱中持续30分钟。falcon管中的cm以3000
×
g旋转5分钟。去除上清液,并且在室温(rt,
ˉ
22℃)下,将小沉淀重新悬浮在2.5ml trizol:水(4:1)溶液中。
[0489]
将冷冻的烧瓶一个接一个地从冰箱中取出并将合适管中的trizol-水转移到烧瓶中。将烧瓶在室温下放置5分钟,然后将内容物吸回到15ml falcon管中(用溶液彻底清洗烧瓶底部后)。在显微镜下检查烧瓶以确保完全去除细胞。将15ml falcon管中的收集的裂解物放入-80℃冰箱中。
[0490]
rna提取
[0491]
从冰箱中取出含有匀浆的falcon管并在室温下储存5分钟,以使核蛋白复合物完
全解离。
[0492]
从每个样品中取出两等份的1ml裂解物,并且向每份中添加200μl氯仿(在细胞裂解步骤期间使用0.2ml氯仿/1ml trizol试剂)并剧烈摇动管。将样品在室温下储存2-3分钟,随后在4℃下以12,000
×
g离心15分钟。
[0493]
混合物分成三层:下层的红色苯酚-氯仿相、中间相和无色的上层水相。rna保留在顶部水相中,dna保留在白色中间(间期)相中,并且蛋白质保留在粉红色底部(有机)相中。水相的顶部3/4被转移到新的干净的eppendorf管中。
[0494]
通过添加等量的100%乙醇,rna从水相中沉淀出来。将沉淀的rna固定在旋转筒(spin cartridge)上,洗涤两次并干燥。在50μl不含rna酶的温水中洗脱rna。通过nanodrop测量纯化的rna的量和质量。在转移到source bioscience进行阵列分析之前,将rna储存在-80℃下。
[0495]
表达数据的分析计划
[0496]
将表达数据下载到每个细胞系的单独文件中。
‘
背景校正’表达是来自阵列的“gprocessedsignal”的数据,所述数据是从相关探针的实际信号中提取的背景信号的结果。这是阵列分析中最常用的。对所有样品的背景校正信号进行log2变换以用于统计分析。为了降低样品中的错误发现率,去除了低于
‘
表达水平’的信号。对于log2变换表达值,
‘
低于表达’水平设置为5。
[0497]
统计学分析
[0498]
基于每个阵列上对照探针的表达模式,决定在分析前对所有阵列进行中值居中(median centering),以减少结果的可变性。数据按细胞类型分组并使用以下算法分析每种细胞类型:
[0499]
·
d0对照与d2对照样品的比较-在正常培养的细胞中看到的表达变化
[0500]
·
d0对照与d2
·
lmw-ds处理的样品的比较-在lmd-ds处理的培养物的细胞中看到的表达变化
[0501]
·
d2对照与d2 lmd-ds处理的样品的比较-在培养物中由lmw-ds诱导的差异表达。
[0502]
进行初步分析以筛选出在三个数据集的任何组合之间没有差异表达的基因。进行简单、非严格的anova(p《0.05)来寻找表达模式。排除在三个数据集上没有变化的探针。使用火山图(volcano plot)分析剩余探针集的倍数变化和显著性。探针表达超过20%的变化(倍数变化(fc)≥1.2或fc≤0.84)首先被认为是显著的,以允许检测表达模式。
[0503]
质量参数
[0504]
根据从施万细胞的细胞储备液中获得的细胞计数计算接种密度。以其最佳密度接种huvecs。
[0505]
来自阵列服务提供商的额外质量控制指示rna是高质量的(无降解),并且量在来自agilent的低输入rna微阵列的参数内。
[0506]
对原始数据的分析指示,如所预期,阵列之间存在显著差异。然而,很容易通过归一化技术消除这些差异(由包括于所有阵列中的相同对照样品的差异反映)。所选择的消除阵列间差异的数据中值居中并不影响代表不同rna浓度的对照之间预期看到的总体差异。
[0507]
施万细胞的表达分析
[0508]
如前所述,在数据分析之前,去除在施万细胞中不表达的基因。对于log2变换表达
值,
‘
低于表达’水平设置为5。这留下了15,842个独特的探针以在施万细胞培养物中进行分析。在分析的下一步中,对三组数据(d0对照与d2对照样品的比较;d0对照与d2 lmw-ds处理的样品的比较;d2对照与d2
·
lmd-ds处理的样品的比较)进行分析,以确定cm对细胞的作用以及由lmw-ds诱导的相对变化。
[0509]
当比较d0对照与d2对照样品时,585个基因在施万细胞培养物中差异表达。受这些基因影响的分子功能涉及以下:细胞运动(1.14e-07-2.49e-03);细胞形态(5.56e-07-2.36e-03);细胞发育(7.3e-06-2.48e-03);细胞生长和增殖(7.3e-06-2.48e-03);细胞装配和组织(1.23e-05-2.36e-03);细胞功能和维持(1.23e-05-2.47e-03);细胞死亡和存活(1.53e-05-2.51e-03);脂质代谢(8.14e-05-1.6e-03);小分子生物化学(8.14e-05-1.6e-03);分子转运(1.18e-04-2.29e-03);蛋白质运输(1.62e-04-1.6e-03);碳水化合物代谢(3.22e-04-1.78e-03);基因表达(3.98e-04-2.2e-03);细胞信号传导(4.39e-04-2.25e-03);细胞间信号传导和相互作用(5.05e-04-2.48e-03);细胞妥协(7.69e-04-1.58e-03);细胞周期(1.12e-03-1.8e-03);氨基酸代谢(1.6e-03-1.6e-03);以及核酸代谢(1.6e-03-1.6e-03)。
[0510]
上面给出的值是代表这些基因与不同途径关联的统计学显著性的p值。两个p值代表观察到的统计显著性的下限和上限(p《0.05是显著的)。
[0511]
如当比较d0对照与d2
·
lmw-ds处理的样品时所评估,lmw-ds在施万细胞培养物中诱导1244个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞形态(1.43e-08-8.39e-04);细胞运动(1.4e-07-9.6e-04);翻译后修饰(3.93e-07-6.71e-05);蛋白质合成(3.93e-07-1.08e-04);蛋白质运输(3.93e-07-1.26e-06);细胞死亡和存活(2.13e-06-8.65e-04);细胞装配和组织(7.46e-06-8.24e-04);dna复制、重组和修复(7.46e-06-7.46e-06);细胞功能和维持(9.53e-06-6.46e-04);基因表达(1.27e-05-4.92e-04);细胞发育(1.29e-05-9.06e-04);细胞生长和增殖(1.29e-05-9.06e-04);细胞间信号传导和相互作用(1.97e-05-8.81e-04);氨基酸代谢(4.22e-05-8.24e-04);小分子生物化学(4.22e-05-8.24e-04);脂质代谢(4.81e-05-3.64e-04);分子转运(3.64e-04-3.64e-04);和细胞周期(4.53e-04-4.86e-04)。
[0512]
如当比较d2对照与d2
·
lmw-ds处理的样品时所评估,lmw-ds在施万细胞培养物中诱导700个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞形态(1.49e-07-5.62e-03);细胞装配和组织(1.49e-07-5.95e-03);细胞运动(7.24e-07-6.06e-03);细胞死亡和存活(9.41e-06-5.95e-03);氨基酸代谢(2.56e-05-3.7e-03);翻译后修饰(2.56e-05-1.05e-03);小分子生物化学(2.56e-05-3.7e-03);细胞间信号传导和相互作用(5.05e-05-5.76e-03);基因表达(7.18e-05-4.94e-03);细胞周期(1.06e-04-5.95e-03);细胞发育(1.06e-04-5.95e-03);细胞功能和维持(1.96e-04-5.95e-03);细胞生长和增殖(2.35e-04-5.95e-03);dna复制、重组和修复(2.75e-04-5.95e-03);细胞信号传导(5.92e-04-2.54e-03);细胞妥协(6.26e-04-6.26e-04);脂质代谢(6.26e-04-1.85e-03);分子转运(6.26e-04-5.95e-03);蛋白质合成(1.05e-03-1.93e-03);对治疗的细胞响应(1.85e-03-1.85e-03);蛋白质运输(2.66e-03-5.95e-03);以及rna转录后修饰(4.32e-03-4.32e-03)。
[0513]
机械分子网络模型模拟了lmw-ds差异调控分子的作用,使得这些变化的功能结果能够被评价。计算机(in silico)模型指示lmw-ds抑制神经元细胞死亡;细胞凋亡;和蛋白
质合成,并活化血管生成;细胞迁移;细胞活力;细胞存活;细胞运动;细胞增殖;细胞分化;细胞内稳态;细胞周期进程;细胞转化;和rna表达。
[0514]
表29总结了培养的施万细胞中基因表达变化的结果。
[0515]
表29-施万细胞中的基因表达变化的总体模式
[0516][0517][0518]
在对照培养物中在两天中表达改变的21个基因在lmw-ds处理的培养物中在相同的两天期间根本没有显示出任何变化。在对照培养物中表达增加的1个基因在lmw-ds处理的培养物中在相同的两天期间下调。在对照培养物中下调的13个基因在lmw-ds处理的培养物中在两天期间上调。122个基因在培养基中被生长因子显著下调,并且这种下调在lmw-ds处理的培养物中甚至更强。441个基因在对照培养物中上调,并且lmw-ds的添加使这种上调明显更强。
[0519]
huvec的表达分析
[0520]
如前所述,在尝试任何分析之前,已经去除了huvec中不表达的基因。对于log2变换表达值,
‘
低于表达’水平设置为5。这留下了15,239个独特的探针以在huvec培养物中进行分析。在下一步中,分析三组数据以确定cm对细胞中基因表达的影响以及由lmw-ds诱导的差异。进行初步分析以筛选出在三个数据集的任何组合之间没有差异表达的基因。进行简单、非严格的anova(p《0.05)来寻找表达模式。在三个数据集中没有变化的基因被消除,留下总共12,313个探针(10,368个基因)进行分析。
[0521]
当比较d0对照与d2对照样品时,1551个基因在huvec培养物中差异表达。受这些基因影响的分子功能涉及以下:细胞装配和组织(2.55e-15-1.29e-03);细胞功能和维持(2.55e-15-1.29e-03);细胞周期(1.98e-11-1.32e-03);细胞形态(3.18e-10-1.29e-03);基因表达(1.05e-08-2.01e-04);细胞发育(1.66e-07-1.37e-03);细胞生长和增殖(1.66e-07-1.37e-03);dna复制、重组和修复(2.04e-07-9.84e-04);细胞死亡和存活(2.09e-07-1.3e-03);rna转录后修饰(4.86e-06-6.53e-04);细胞运动(9.9e-06-1.18e-03);翻译后修饰(1.92e-05-1.34e-03);细胞间信号传导和相互作用(2.19e-05-9.1e-04);蛋白质合成(5.49e-05-1.14e-03);细胞妥协(8.16e-05-8.16e-05);分子转运(6.27e-04-6.27e-04);蛋白质运输(6.27e-04-6.27e-04);细胞信号传导(8.86e-04-8.86e-04);对治疗的细胞响应(9.84e-04-9.84e-04);和蛋白质降解(1.14e-03-1.14e-03)。
[0522]
如当比较d0对照与d2 lmw-ds处理的样品时所评估,lmw-ds在huvec培养物中诱导了1779个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞装配和组织(4.14e-17-9.7e-04);细胞功能和维持(4.14e-17-8.05e-04);细胞周期(5.83e-14-9.85e-04);细
胞形态(1.69e-10-7.48e-04);基因表达(7.99e-09-8.62e-04);细胞死亡和存活(2e-08-8.4e-04);细胞发育(1.28e-07-8.88e-04);细胞生长和增殖(1.28e-07-8.88e-04);dna复制、重组和修复(3.07e-07-9.7e-04);rna转录后修饰(1.13e-06-6.31e-04);细胞运动(1.42e-06-8.34e-04);翻译后修饰(3.4e-05-9.17e-04);细胞间信号传导和相互作用(6.97e-05-9.56e-04);分子转运(7.43e-05-9.7e-04);蛋白质运输(7.43e-05-7.43e-05);rna运输(1.57e-04-5.72e-04);蛋白质合成(1.92e-04-9.02e-04);细胞妥协(2.47e-04-6.28e-04);和细胞信号传导(4.64e-04-9.02e-04)。
[0523]
如当比较d2对照与d2 lmw-ds处理的样品时所评估,lmw-ds在huvec培养物中诱导了76个基因的差异表达。受这些基因影响的分子功能涉及以下:dna复制、重组和修复(9.62e-05-2.57e-02);细胞周期(1.22e-04-2.4e-02);细胞发育(1.59e-04-2.67e-02);细胞形态(4.64e-04-2.42e-02);细胞功能和维持(4.64e-04-2.57e-02);脂质代谢(9.49e-04-1.07e-02);分子转运(9.49e-04-1.61e-02);小分子生物化学(9.49e-04-1.87e-02);细胞妥协(1.6e-03-2.62e-02);细胞死亡和存活(2.06e-03-2.67e-02);氨基酸代谢(2.7e-03-2.7e-03);碳水化合物代谢(2.7e-03-1.07e-02);细胞间信号传导和相互作用(2.7e-03-2.4e-02);细胞装配和组织(2.7e-03-2.57e-02);细胞生长和增殖(2.7e-03-2.4e-02);细胞运动(2.7e-03-2.4e-02);能量产生(2.7e-03-2.7e-03);核酸代谢(2.7e-03-1.07e-02);翻译后修饰(2.7e-03-1.61e-02);基因表达(5.39e-03-2.36e-02);rna转录后修饰(5.39e-03-2.4e-02);药物代谢(8.07e-03-1.61e-02);维生素和矿物质代谢(8.07e-03-8.07e-03);蛋白质合成(1.07e-02-1.07e-02);rna运输(1.07e-02-1.07e-02);对治疗的细胞响应(1.24e-02-1.24e-02);和自由基清除(1.43e-02-1.43e-02)。
[0524]
虽然在处理2天后,对照与lmw-ds处理的培养物之间的总体差异乍一看并不大,但是lmw-ds对基因表达变化的影响是显著的,特别是当考虑到lmw-ds对生长因子诱导的基因表达的调节时。
[0525]
使用机械分子网络模型,有可能模拟由lmw-ds差异调控的基因的效果,以寻找这些变化的功能结果。计算机模型指示lmw-ds抑制神经元细胞死亡;细胞凋亡;和蛋白质合成,并活化血管生成;细胞迁移;细胞活力;细胞存活;细胞运动;细胞增殖;细胞分化;细胞内稳态;细胞周期进程;细胞转化;和rna表达。
[0526]
huvec对照培养物包含生长因子。在处理的培养物中,将lmw-ds添加到已经含有生长因子的培养基中。
[0527]
表30总结了培养的huvec中的基因表达变化的结果。在对照培养物中在两天中(在生长因子的作用下)表达改变的67个基因在lmw-ds处理的培养物中在相同的两天期间根本没有显示出任何变化。在含有生长因子的对照培养物中表达增加的4个基因在lmw-ds处理的培养物中在相同的两天期间下调。对照培养物中被生长因子下调的11个基因在lmw-ds处理的培养物中在两天期间上调。120个基因被生长因子显著下调,并且这种下调在lmw-ds处理的培养物中甚至更强。229个基因在对照培养物中上调,并且lmw-ds的添加使这种上调明显更强。
[0528]
表30-huvec中的基因表达变化的总体模式
9.01e-03);分子转运(1.05e-03-9.01e-03);细胞信号传导(1.13e-03-5.09e-03);和碳水化合物代谢(4.71e-03-4.71e-03)。
[0536]
如当比较d0对照与d2
·
lmw-ds处理的样品时所评估,lmw-ds在运动神经元中诱导425个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞死亡和存活(2.87e-08-6.27e-03);细胞运动(4.73e-07-6.47e-03);细胞形态(4.95e-07-7.47e-03);细胞发育(1.02e-06-7.13e-03);细胞生长和增殖(1.02e-06-7.48e-03);细胞装配和组织(7.03e-06-7.47e-03);细胞功能和维持(7.03e-06-7.47e-03);基因表达(1.95e-05-6.18e-03);细胞周期(2.88e-05-7.48e-03);dna复制、重组和修复(3.39e-05-5.16e-03);氨基酸代谢(7.75e-05-4.68e-03);小分子生物化学(7.75e-05-4.68e-03);细胞妥协(8.23e-05-4.61e-03);细胞间信号传导和相互作用(3.27e-04-7.48e-03);维生素和矿物质代谢(3.27e-04-3.27e-04);蛋白质合成(8.94e-04-5.29e-03);翻译后修饰(9.67e-04-9.67e-04);分子转运(9.7e-04-4.68e-03);蛋白质运输(9.7e-04-9.7e-04);碳水化合物代谢(1.44e-03-1.92e-03);对治疗的细胞响应(1.92e-03-1.92e-03);和脂质代谢(4.68e-03-4.68e-03)。
[0537]
表31-运动神经元中的基因表达变化的总体模式
[0538][0539][0540]
皮质神经元的表达分析
[0541]
如前所述,在尝试任何分析之前,已经去除了运动神经元中不表达的基因。对于log2变换表达值,
‘
低于表达’水平设置为5。这留下了10,653个独特的探针,其中系列中至少三个样品达到了表达阈值。在下一步中,分析三组数据以确定cm对细胞的作用和由lmw-ds诱导的差异。
[0542]
当从一组分离的细胞中发育出皮质神经元表型时,正常培养条件下基因表达的变化模拟了皮质神经元的正常发育过程。正常培养基中的生长因子是这些细胞分化所必需的那些。这些培养物中存在的应激因子是氧化应激(对于组织培养条件来说是正常的)。
[0543]
当比较d0对照与d2对照样品时,1101个基因在运动神经元培养物中差异表达。受这些基因影响的分子功能涉及以下:细胞装配和组织(3.57e-25-6.65e-04);细胞功能和维持(3.57e-25-6.65e-04);细胞形态(4.28e-22-6.36e-04);细胞发育(4.28e-22-6.53e-04);细胞生长和增殖(4.28e-22-6.6e-04);细胞间信号传导和相互作用(2.16e-13-6.65e-04);分子转运(5.18e-12-4.95e-04);细胞运动(1.86e-11-6.65e-04);细胞死亡和存活(3.37e-11-6.41e-04);基因表达(1.27e-08-8.96e-05);蛋白质合成(3.84e-07-8.69e-05);小分子生物化学(6.65e-07-5.18e-04);细胞妥协(7.12e-06-4.54e-04);蛋白质降解
(1.62e-05-1.62e-05);氨基酸代谢(2.11e-05-4.25e-04);蛋白质运输(3.4e-05-3.4e-05);细胞信号传导(8.69e-05-3e-04);翻译后修饰(8.69e-05-2.15e-04);蛋白质折叠(2.15e-04-2.15e-04);细胞周期(2.69e-04-3.07e-04);dna复制、重组和修复(2.69e-04-4.77e-04);核酸代谢(2.69e-04-2.69e-04);脂质代谢(3.12e-04-5.18e-04);和碳水化合物代谢(5.18e-04-5.18e-04)。
[0544]
如当比较d0对照与d2
·
lmw-ds处理的样品时所评估,lmw-ds在运动神经元中诱导609个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞装配和组织(3.91e-15-1.83e-03);细胞功能和维持(3.91e-15-1.83e-03);细胞形态(2.53e-13-1.43e-03);细胞发育(2.53e-13-1.81e-03);细胞生长和增殖(2.53e-13-1.83e-03);细胞运动(4.95e-09-1.2e-03);细胞间信号传导和相互作用(5.96e-09-1.47e-03);细胞死亡和存活(2.25e-08-1.77e-03);分子转运(7.08e-08-1.79e-03);dna复制、重组和修复(3.03e-06-1.71e-03);细胞妥协(9.23e-06-7.65e-04);氨基酸代谢(1.75e-05-1.64e-03);细胞周期(1.75e-05-1.77e-03);小分子生物化学(1.75e-05-1.79e-03);蛋白质合成(2.77e-05-1.5e-03);蛋白质运输(2.77e-05-1.9e-04);细胞信号传导(7.65e-05-1.73e-03);翻译后修饰(3.01e-04-1.4e-03);基因表达(3.65e-04-1.15e-03);药物代谢(6.49e-04-6.49e-04);碳水化合物代谢(6.95e-04-7.69e-04);维生素和矿物质代谢(1.09e-03-1.09e-03);和核酸代谢(1.44e-03-1.73e-03)。
[0545]
如当比较d0对照与d2
·
lmw-ds处理的样品时所评估,lmw-ds在运动神经元中诱导247个基因的差异表达。受这些基因影响的分子功能涉及以下:细胞形态(6.01e-08-1.01e-02);细胞发育(7.46e-08-1.01e-02);细胞生长和增殖(7.46e-08-1.01e-02);细胞死亡和存活(4.23e-07-1.01e-02);细胞运动(2.69e-06-9.91e-03);细胞装配和组织(1.57e-05-1.01e-02);细胞功能和维持(1.57e-05-1.01e-02);细胞周期(1.01e-04-1.01e-02);细胞间信号传导和相互作用(1.01e-04-1.01e-02);脂质代谢(1.56e-04-1.01e-02);小分子生物化学(1.56e-04-1.01e-02);基因表达(2.28e-04-3.38e-03);rna损害和修复(2.28e-04-2.28e-04);rna转录后修饰(2.28e-04-2.28e-04);分子转运(4.18e-04-8.32e-03);细胞妥协(4.47e-04-2.2e-03);蛋白质合成(2.66e-03-7.29e-03);蛋白质运输(4.11e-03-8.32e-03);蛋白质降解(5.64e-03-7.29e-03);以及dna复制、重组和修复(7.31e-03-1.01e-02)。
[0546]
表32-皮质神经元中的基因表达变化的总体模式
[0547][0548]
lmw-ds对线粒体中的氧化应激途径的影响
[0549]
线粒体中发生的氧化应激途径对于各种感染性疾病以及还有衰老和年龄相关退行性疾病非常重要。正常生长条件会在细胞中触发一定量的氧化应激,这有助于体内和体
外老化过程。
[0550]
在正常条件下培养的施万细胞中,复合物i(nadh脱氢酶)被抑制,而复合物iv(细胞色素c氧化酶)被活化。当将lmw-ds添加至培养物中时,复合物iii(细胞色素bc1)被抑制。复合物iii的抑制作用抑制了参与癌症和神经疾病的发病机理的氧化应激现象。
[0551]
复合物iii(有时被称为辅酶q:细胞色素c-氧化还原酶或细胞色素bc1复合物)是电子转运链(ec 1.10.2.2)中的第三种复合物,在atp的生物化学生成(氧化磷酸化)中起着关键作用。复合物iii是一种由线粒体(细胞色素b)和核基因组(所有其他亚基)编码的多亚基跨膜蛋白。复合物iii存在于所有动物和所有需氧真核生物的线粒体和大多数真细菌的内膜中。复合物iii的突变引起运动不耐受以及多系统病症。bc1复合物包含11个亚基、3个呼吸亚基(细胞色素b、细胞色素c1、里斯克蛋白(rieske protein))、2个核心蛋白和6个低分子量蛋白。
[0552]
在huvec中,用lmw-ds处理后,没有检测到氧化应激对线粒体的影响的显著调节。
[0553]
在正常培养条件下,运动神经元似乎遭受显著的氧化应激。这导致一些细胞凋亡机制的活化并涉及细胞色素c、aif、半胱天冬酶3、8和9的活化。此外,运动神经元的特征在于细胞中淀粉样蛋白-β的产生,其进一步加剧氧化应激和线粒体断裂(通过fias1)以及脂肪酸的氧化。此外,复合物v被活化。
[0554]
向培养物中添加lmw-ds通过经由防止淀粉样蛋白-β产生及其对线粒体断裂和功能障碍以及随后的损害的负面影响,以及经由抑制脂肪酸氧化来防止和抑制细胞凋亡,从而改善了这些负面影响。lmd-ds也抑制涉及trak1和pink1的反应途径,从而有助于改善线粒体功能。lmw-ds进一步降低了h2o2的水平。另一个效应是抑制htra2有助于抑制细胞凋亡。
[0555]
在正常培养条件下,皮质神经元暴露于显著的氧化应激,导致淀粉样蛋白-β的产生和路易体(lewy body)形成,并且涉及突触核蛋白α的活化和ros水平的增加;细胞凋亡;线粒体断裂;和涉及c161的线粒体功能的降低。向培养物中添加lmw-ds能够预防和逆转大多数这些有害作用,例如淀粉样蛋白-β的累积和路易体病理、线粒体功能障碍。一些细胞凋亡诱导机制保持活性,可能是由于在培养物中的强活化。
[0556]
lmw-ds对谷氨酸兴奋毒性的影响
[0557]
谷氨酸是参与长时程增强(ltp),即学习和记忆功能的必需兴奋性氨基酸。然而,过多的谷氨酸也与兴奋毒性有关,导致神经元死亡。后一种现象被假设参与慢性神经退行性病况以及还有tbi中触发的神经元死亡。参与谷氨酸信号传导的基因在huvec中不表达,但在本研究中使用的施万细胞和神经元细胞系中存在。
[0558]
运动神经元培养中的基线条件抑制了谷氨酸产生。这种抑制不受lmw-ds的影响。在基线处,皮质神经元中的谷氨酸产量增加。添加lmw-ds并没有改变这些细胞中谷氨酸的产生。
[0559]
向施万细胞的cm中添加lmw-ds诱导了蛋白质复合物的表达(calm、gβγ、grm7、pick1)。更重要的是,lmw-ds增加了施万细胞中谷氨酸转运蛋白并且特别是slc1a2/3的活性和/或水平,从而导致突触前神经元产生和释放的谷氨酸的清除。因此,lmw-ds诱导施万细胞从突触间隙中清除有毒的谷氨酸,从而阻止其发挥兴奋毒性。
[0560]
slc1a3,溶质载体家族1(神经胶质高亲和力谷氨酸转运蛋白),成员3,是一种在人类中由slc1a3基因编码的蛋白质。slc1a3通常也被称为谷氨酸天冬氨酸转运蛋白(glast)
或兴奋性氨基酸转运蛋白1(eaat1)。slc1a3主要在质膜中表达,使其能够从细胞外空间清除谷氨酸。也定位于线粒体内膜,作为苹果酸-天冬氨酸穿梭的部分。slc1a3在体内作为同源三聚体起作用。slc1a3通过三个na
和一个h
阳离子的共转运和一个k
阳离子的反向转运介导谷氨酸和天冬氨酸的转运。这种共转运偶联(或同向转运)允许谷氨酸逆着浓度梯度转运到细胞中。slc1a3在整个cns中表达,并且在小脑的星形胶质细胞和博格曼胶质细胞(bergmann glia)中高度表达。在视网膜中,slc1a3在米勒细胞(muller cell)中表达。slc1a3也在许多其他组织中表达,包括心肌细胞。
[0561]
slc1a2,溶质载体家族1成员2,也被称为兴奋性氨基酸转运蛋白2(eaat2)和谷氨酸转运蛋白1(glt-1),是一种人类中由slc1a2基因编码的蛋白质。slc1a2是蛋白质溶质载体家族的一员。膜结合蛋白是从cns内突触处的细胞外空间清除兴奋性神经递质谷氨酸的主要转运蛋白。谷氨酸清除对于正确的突触活化和防止谷氨酸受体过度活化造成的神经元损害是必要的。slc1a2负责大脑中90%以上的谷氨酸重摄取。
[0562]
这些发现指示,lmw-ds可能可用于在其细胞外高水平有害的情况下,如tbi后,预防谷氨酸兴奋性毒性。
[0563]
lmw-ds对细胞粘附的影响
[0564]
lmw-ds的一个显著的表型强效应是对细胞粘附的影响,这是细胞类型特异性的。细胞粘附在神经元中受影响最大,其次是施万细胞,而huvec不受影响。
[0565]
基因表达的分析指示,这是由于lmw-ds对调控细胞附着的酶的表达的影响,所述酶包括金属肽酶,也称为基质金属蛋白酶(mmp),见表33。
[0566]
这些分子对施万细胞中调控细胞运动和附着的途径的综合作用(17个分子,见表33)是这样的,即细胞粘附将被抑制,而细胞运动将被活化,而在huvec(1个分子,adam11)中,粘附将不受影响,但血管生成将被活化。
[0567]
表33-施万细胞中调控细胞运动和附着的途径的分子
[0568][0569]
分析神经元中由lmw-ds诱导的差异基因表达的效果。在运动神经元中,相同的金属肽酶依赖性途径可能负责在施万细胞中看到的细胞脱离,见表34。
[0570]
表34-运动神经元中调控细胞运动和附着的途径的分子
[0571]
[0572][0573]
然而,没有发现mmp相关基因在皮质神经元中有差异表达。
[0574]
这一发现导致了对影响细胞附着和附着相关分子的所有分子相互作用及其在四种不同培养物中对细胞附着的影响的重新评估。217种附着相关分子(197种基因和20种药物)的完整列表如下所示:
[0575]
ace2、acp1、adam15、adgrb1、adgre2、adipoq、ag490、ambn、angpt1、antxr1、arap3、arms2、巴马司他(batimastat)、bcam、bcap31、bcar1、苯甲氧基羰基-leu-leu-leu-醛、
bmp2、bmp4、btc、c1qbp、ca
2
、ca9、cadm1、calr、花萼海绵诱癌素(calyculin)a、半胱天冬酶、cbl、cd209、cd36、cd44、cd46、cdh13、西伐他汀(cerivastatin)、氯霉素(chloramphenicol)、硫酸软骨素、clec4m、秋水仙碱、i型胶原蛋白、胶原、comp、crk、crp、csf1、csf2rb、ctgf、姜黄素(curcumin)、cxcl12、环状amp、dab2、dag1、dcn、ddr1、去铁螯素(desferriexochelin)772sm、dock2、dsg2、dsg4、多晶型磷灰石(durapatite)、efna、efna1、efnb、efnb1、egf、egfr、egr1、eln、eng、ep300、eph受体、epha8、ephb1、依替巴肽(eptifibatide)、乙二胺四乙酸、ets1、f11r、f3、fbln5、fbn1、fc受体、fcn2、fermt2、fes、fgf2、fgfr1、纤维蛋白(fibrin)、fn1、病灶粘附激酶、fsh、fut3、fut6、fut7、fyn、hacd1、肝素、组蛋白h3、组蛋白h4、hras、hspg2、htn1、透明质酸、皮质醇、过氧化氢、icam1、icam2、igf1r、igg、igg3、il1、il1b、il6、ilk、整合素、整合素α4β1、整合素α、ipo9、itga1、itga2、itga3、itga5、itga6、itgb1、itgb2、itgb3、itgb5、jak2、jnk、kp-sd-1、lamc1、层粘连蛋白、层粘连蛋白1、左旋甲状腺素、lgals3、lif、脂多糖、lox、lrp1、lrpap1、mad1l1、甘露糖、mapk7、mbl2、mertk、甲硝唑(metronidazole)、mgat5、mmp2、mn
2
、nck、nedd9、nrg1、冈田酸(okadaic acid)、olr1、p38 mapk、pdgf bb、磷脂酰肌醇(phosphatidylinositol)、pkm、血小板活化因子、pld1、plg、pmp22、podxl、postn、prkcd、ptafr、pten、ptger2、ptk2、ptk2b、ptn、ptpn11、ptprz1、吡咯烷二硫氨基甲酸酯、rac、ralb、ranbp9、rhoa、rhob、rpsa、sdc3、sele、选择素、sell、sema3a、辛伐他汀(simvastatin)、sirpa、sparc、鞘氨醇-1-磷酸、spi1、spp1、spry2、src、stard13、swap70、tek、tfpi、tfpi2、tgfa、tgfb1、tgfbi、tgm2、thbs2、thy1、甲状腺激素、timp2、替罗非班(tirofiban)、tln1、tln2、tnf、tp63、维甲酸(tretinoin)、vav1、vcam1、vcan、vegf、vhl、vtn、vwf和wrr-086。
[0576]
在197个调控细胞附着的基因中,没有一个是由huvec中的lmw-ds差异调控的。在施万细胞培养物中,17个差异表达的分子导致了整体上轻微增加的附着。然而,在神经元中,表达模式导致这些细胞中细胞附着的显著抑制。
[0577]
受lmw-ds影响的上游调控因子途径
[0578]
在施万细胞中,上游调控因子分析显示,lmw-ds通过增加其在系统中的活化或减少其的抑制来调节几种生长因子的作用,如表35中所示。
[0579]
表35-施万细胞中的上游调控因子比较
[0580]
[0581][0582]
在huvec中,lmw-ds增强其作用的生长因子数量相对较少,但仍然非常显著,见表36。
[0583]
表36-huvec中的上游调控因子比较
[0584]
[0585][0586]
如表37中所示,在运动神经元中,上游调控因子分析显示,lmw-ds影响几种生长因子的作用,从而增加其的活化或减少系统中存在的抑制。
[0587]
表37-运动神经元中的上游调控因子比较
[0588][0589]
在皮质神经元中,在正常培养条件下,大多数生长因子依赖途径被正常培养基显著活化。在大多数情况下,这种活化不会被lmw-ds改变。然而,lmw-ds活化作为gdf7下游效应子的分子,从而指示这种生长因子的作用被lmw-ds增强。因为gdf7是神经元的强有力的分化因子,并且这些生长因子的额外活化(除了bdnf和nt3的活化之外)为这些细胞在培养物中的增强分化提供了一个很好的解释。
[0590]
讨论
[0591]
huvec的正常培养条件模拟组织缺氧和再灌注后的环境,其含有高营养含量和生长因子,还补充有肝素。lmw-ds处理的培养物模拟了缺氧和再灌注24小时后添加的lmw-ds的效果。这涉及的真实生活场景是在缺血条件诸如纤维化条件后的血管生成。
[0592]
在施万细胞中,具有高营养含量和葡萄糖的对照培养物再现了施万细胞的活化。lmw-ds处理的培养物模拟了胶质细胞活化24小时后添加的lmw-ds的效果。这概括的现实生活场景是神经系统受损后的胶质细胞活化。
[0593]
具有高营养成分和生长因子的神经元(运动神经元和皮质神经元)的正常培养条
件模拟正常神经元分化期间的环境。在这些培养物中,唯一的负面影响是细胞遭受的氧化应激。与此相关的真实生活场景是在存在大量生长和分化因子的情况下由氧化应激驱动的退行性病况。这对应于其中氧化应激起关键作用的神经退行性疾病或病况的早期阶段。
[0594]
从细胞类型可以清楚地看出,在施万细胞和huvec中观察到的分子效应支持lmw-ds在以下方面中的作用:抗细胞凋亡的保护;血管生成的诱导;增加的细胞迁移和运动;增加的细胞活力和存活;和细胞分化的诱导。对关键分子途径的分析指示,在神经元中,lmw-ds将减少氧化应激对线粒体的影响,并且将减少神经退行相关分子,诸如淀粉样蛋白-β和路易体。
[0595]
因此,来自huvec细胞模型的结果指示,lmw-ds可以防止细胞损害,并且促进受伤或患病组织中新血管的发育,诸如在中风后。来自施万细胞的结果指示,lmw-ds可以防止患病或受损的神经系统中的细胞损失。
[0596]
对关键分子途径的分析指示,在施万细胞中,lmw-ds减少了氧化应激对线粒体的影响并增加了谷氨酸的摄取。在施万细胞中的结果指示,lmw-ds可以防止由于患病或受损的神经系统中的氧化应激和谷氨酸兴奋性毒性而发生的细胞损失。
[0597]
特别重要的是,lmw-ds增加了神经胶质细胞的谷氨酸摄取,如施万细胞所呈现。然而,lmw-ds并没有改变神经元产生的谷氨酸。这很重要,因为谷氨酸是ltp、学习和记忆所需要的。因此,lmw-ds没有改变神经元产生谷氨酸是有益的,因为这种谷氨酸是上述过程中正常神经传递所需要的。然而,由于lmw-ds的作用,受损或死亡细胞释放的谷氨酸水平的增加将被周围的神经胶质细胞有效吸收。因此,由lmw-ds引起的胶质细胞中谷氨酸转运蛋白的活化有效地从神经裂中去除了由受损或死亡的神经元释放的谷氨酸。这继而阻止了谷氨酸发挥其兴奋毒性,并从而进一步损害神经元。因此,lmw-ds诱导神经胶质细胞摄取具有潜在有害神经毒性量的谷氨酸。
[0598]
因此,在神经元中的结果证实了lmw-ds在减少由于氧化应激引起的继发性组织损坏害、促进修复和减少退行相关蛋白积累方面的潜在治疗作用。
[0599]
总的来说,这些结果支持lmw-ds在总体上防止细胞凋亡并且特别是防止神经元细胞死亡、诱导血管生成、增加细胞迁移和运动、增加细胞活力和存活、诱导细胞分化、减少氧化应激的影响、减少谷氨酸兴奋性毒性和减少退行相关蛋白产物(诸如淀粉样蛋白-β和路易体)的产生中的作用。
[0600]
细胞粘附主要在神经元和施万细胞中受到影响,其中lmw-ds促进细胞脱离和运动。在huvec中,细胞粘附不受影响。对细胞粘附的作用主要是由于金属蛋白酶类酶的表达,但其他粘附分子的调节也有助于这种作用。
[0601]
通过tgfβ驱动作为病理反应的瘢痕形成。tgfβ诱导171个分子的大的互连网络,引起免疫细胞的粘附、细胞的活化、细胞运动、细胞的聚集、纤维化和tgfβ的诱导。施用lmw-ds完全消除了tgfβ诱导的免疫细胞粘附、细胞活化、细胞聚集、纤维化和tgfβ自活化的效应。即使当tgfβ被活化时,即,即使存在过量的tgfβ,也可以观察到lmw-ds对施万细胞中由tgfβ驱动的分子网络的失活作用。
[0602]
因此,这些研究证实了lmw-ds在抑制促炎性纤维化细胞因子tgfβ的信号传导方面的潜在治疗作用,所述信号传导反过来抑制组织纤维化,否则组织纤维化在covid-19受试者中是一个严重的组分。
[0603]
实施例8
[0604]
本实施例显示,lmw-ds消退了啮齿动物和人类纤维变性疾病模型中的炎性瘢痕形成和活化的基质重塑,以实现功能性组织再生。
[0605]
结果
[0606]
poag是慢性眼纤维化疾病的实例。在眼睛中,已知tgfβ增加tm中的ecm的沉积,最显著的是纤连蛋白。纤连蛋白存在于连接tm的最内侧区域和施累姆氏管(schlemm’s canal)的鞘物质中并有助于调控组织的收缩特性,并且因此调控aqh引流和iop。在青光眼患者中检测到较高水平的tgfβ和纤连蛋白。在这里,我们证明了lmw-ds显著降低了用tgfβ2处理的培养的人小梁网细胞中的纤连蛋白水平(1136
±
155相对于1579
±
210;p《0.05;图15a、15b)。在此之后,我们在前段纤维化的啮齿动物模型(一种青光眼的实验模型)中评估了皮下(sc)注射lmw-ds的作用,其中纤维化是通过每两周前房内(ic)注射tgfβ1诱导的(图16a);通过建立tm纤维化,每周两次ic tgf-β1注射到第14天使iop显著升高至13mmhg,此时开始每天sc注射lmw-ds或盐水。在接受ic tgfβ1加sc注射盐水的对照大鼠中,iop继续上升,在第28天达到17.6
±
2.7mmhg(图16b)。相比之下,ic tgfβ1联合sc lmw-ds注射到第28天使iop降至11.4
±
0.2mmhg(p《0.0001;图16b),这在正常范围内(10-12mmhg)。与sc盐水处理的对照大鼠中的tm(分别为63.7
±
6.8%和63.6
±
2.9%)相比,tm中的纤维化的免疫组织化学分析显示ecm分子层粘连蛋白(25.6
±
3.5%,p《0.001;图16c)和纤连蛋白(40.2
±
4.0%,p《0.01,图16d)显著减少。这伴随着存活的视网膜神经节细胞(rgc)数量的显著增加(与对照盐水注射的大鼠中为9.0
±
0.3rgc/mm相比,sc lmw-ds注射的大鼠中为12.6
±
0.4rgc/mm;p《0.01;图16e)和保存的视网膜神经纤维层(rnfl)厚度(与在对照sc盐水注射的大鼠中为44.0
±
1.6μm相比,在sc lmw-ds注射的大鼠中为59.9
±
1.2μm;p《0.01;图16f),如通过光学相干断层扫描(oct)所测量。
[0607]
讨论
[0608]
由于在纤维增生条件下炎症先于纤维化,因此了解lmw-ds如何调节人类细胞中的炎症和组织重塑途径是重要的。来自相关培养的人细胞的蛋白质和基因表达数据表明,通过重编程免疫活化、炎症消退和组织重塑(实施例7),lmw-ds调节关键的先天和适应性响应,导致患病和受损人组织的愈合和功能重塑改善。
[0609]
在实施例6中,使用diversity plus测定探索蛋白质水平下的lmw-ds的机制曲线,所述测定包括12种不同的经验证的人原代细胞培养系统,其模拟了不同人组织类型中的炎症和伤口愈合响应。这些数据显示,lmw-ds具有一个独特的有多模式作用的表型曲线;在不同的细胞类型中起作用,并且随后用不同的炎性介体刺激。特别是,lmw-ds降低了3种趋化因子cxcl9、cxcl10和cxcl11的水平,其都是结构相关的、通过cxc受体3(cxcr3)发出信号并且由干扰素和tnfα诱导。cxcr3及其相关趋化因子在免疫细胞的募集和功能中起重要作用,并且与许多炎性和自身免疫性疾病有关。此外,其还被用作疾病的预后和预测生物标志物。cxcr3途径在青光眼的病因学中起重要作用,其表达水平与poag的进展相关。在高眼压模型中,cxcr3的拮抗作用降低了iop,增加了aqh流出,减少了炎症并减少了tm和视网膜细胞的细胞凋亡。
[0610]
基因表达数据(实施例7)和来自青光眼模型的结果都支持lmw-ds独特的多模式影响,因为其作用于tgfβ信号传导途径以及其他ecm重塑信号。在poag中,tgfβ在ecm蛋白在tm
ganglion cell loss in rodents through fibrolysis of the scarred trabecular meshwork.investigative ophthalmology&visual science.2015;56(6):3743-57)。在第0天,使用15
°
的一次性刀片形成通过角膜的自封闭通道,并且该接入点用于所有随后在28天实验中使用无菌玻璃微量移液器(harvard apparatus,kent,uk)进行的每周两次3.5μl ic注射5ng/ml tgfβ1(peprotech,london,uk)。在整个28天的实验中,还每周两次测量iop(在ic注射之前立即)。在第14天,对tgfβ无响应者(即,其的iop在任何时间点都没有增加到基线以上)被排除在进一步分析之外,以产生lmw-ds的n=7只大鼠(14只眼)和盐水媒介物治疗的n=5只大鼠(10只眼)的纳入。在第14天,大鼠接受15mg/kg lmw-ds(tikomed ab)或等量盐水体积的皮下注射,每天持续14天。在第28天进行rfnl的oct测量,作为rgc密度的替代,并且对两组的眼组织进行免疫组织化学处理,以评估tm纤维化和rgc存活的水平。
[0620]
动物和手术
[0621]
根据1986年《动物法(animal act)》(英国)中阐述的内政部指南(home office guideline),遵守根据英国内政部许可证号70/8611在眼科和视觉研究中使用动物的arrive指南和arvo声明,在伯明翰大学(university of birmingham)(英国)的生物医学服务单位(biomedical services unit)进行啮齿动物研究。
[0622]
圈养成年(8-10周龄)雄性斯普拉-道来氏大鼠(sprague dawley rat)(charles river,kent,uk)以在12小时黑暗/光照周期下自由获取食物和水。根据1986年《动物法》(英国)中阐述的内政部指南和在眼科和视觉研究中使用动物的arvo声明,在伯明翰大学(英国)的生物医学服务单位进行外科手术。当大鼠使用2%-5%异氟烷/98%-95%o2(national vet supplies,uk)进行吸入麻醉时,进行眼内注射、光学相干断层扫描和眼内压测量。所有大鼠的健康一直受到密切监控。
[0623]
iop测量
[0624]
使用为大鼠校准的icare tonolab(icare,helsinki,finland)获得iop测量值。在整个28天的实验中,在上午9点与上午11点之间,每周两次记录iop,以避免混淆iop的昼夜节律变化。在麻醉诱导后立即用眼压计从中央角膜进行六次回弹测量并取平均值以给出单一读数(mmhg)。每个图形数据点代表从每只大鼠连续获得的三个读数(6次回弹)的平均值
±
sem,以确保准确的测量。
[0625]
光学相干断层扫描
[0626]
在第28天使用spectralis hra3共聚焦扫描激光检眼镜(heidelberg engineering,heidelberg,germany)进行视网膜神经纤维层(rnfl)测量。图像从处于吸入麻醉下的大鼠获得(如上文
‘
动物和手术’章节所述)。对视神经头周围的视网膜拍摄oct图像,并且使用内置软件分割图像并定量rnfl厚度。
[0627]
免疫组织化学的组织制备
[0628]
通过在第28天腹膜内注射戊巴比妥钠(national veterinary supplies)对大鼠实施安乐死,并且同时在终端麻醉下,用含4%多聚甲醛(pfa;taab,aldermaston,uk)的pbs进行心内灌注。取出眼睛,并在室温下在含4%pfa的pbs中浸泡固定2小时,然后各自在4℃下使用增加浓度(10%、20%和30%蔗糖)冷冻保护24小时。随后,将眼睛包埋在单向剥离模具容器(agar scientific,essex,uk)中的最佳切割温度(oct)包埋介质(thermo shandon,runcorn,uk)中,并且在储存于-80℃之前使用碎干冰快速冷冻。使用bright冷冻切片机
(bright,huntingdon,uk)在-22℃下在通过视神经头(考虑rgc变化)的矢状面中对眼睛进行切片(15μm),并且将其固定在带正电荷的载玻片(superfrost plus,fisher scientific,usa)上并在37℃下干燥2小时并在-20℃下储存。
[0629]
免疫组织化学
[0630]
除非另有说明,否则所有试剂均购自sigma(poole,uk)。将冷冻的组织切片放置30分钟以平衡至室温,然后在pbs 3
×
中水合5分钟,然后浸入0.1%triton x-100中20分钟以渗透切片。再用pbs3
×
5分钟洗涤后,用疏水pap笔(vector laboratories,peterborough,uk)分离眼部切片。使用含0.5%bsa、0.3%tween和15%正常山羊血清的pbs阻断非特异性抗体结合位点。然后将切片放置于一级抗体(层粘连蛋白l9393,纤连蛋白f3648,sigma,brn3a sc-31984,santa cruz)中,并且在4℃下放置过夜,然后在pbs中洗涤3
×
5分钟,并且在室温下与二级抗体(山羊-抗兔alexa fluor 594或山羊-抗小鼠alexa fluor 488)一起孵育1小时。在用含dapi的vectashield(vector laboratories)固定之前,将切片在pbs中洗涤3
×
5分钟。不与一级抗体但与二级抗体一起孵育的对照组织切片全部被阴性染色(未显示)。
[0631]
显微术和分析
[0632]
使用zeiss axioplan 2荧光显微镜(carl zeiss ltd,oberkochen,germany),通过对使用随机数的治疗组进行掩蔽的操作员来分析荧光染色的切片,如前所述(hill,l.j.等人decorin reduces intraocular pressure and retinal ganglion cell loss in rodents through fibrolysis of the scarred trabecular meshwork.investigative ophthalmol.vis.sci.56,3743
–
3757(2015))。简而言之,在相同的曝光(250ms)下捕获每种抗体的图像,以评估ecm水平的像素强度。使用image j软件测量感兴趣区域内阈值以上的免疫荧光像素的百分比。
[0633]
实施例9
[0634]
本实施例研究了lmw-ds在体外影响人pbmc刺激释放il-6的能力。
[0635]
人pbmc可以在体外被多种可以直接或间接地活化各种细胞亚群的剂刺激。监测细胞因子释放允许研究药物的潜在影响,以预测对患者的作用。il-6是典型的促炎细胞因子,其已被证明与多种病理学相关,包括神经退行性病况(如als)、炎性疾病(如关节炎)和病毒感染的严重后果(如covid-19)。
[0636]
材料与方法
[0637]
外周血单核细胞(pbmc)通过ficoll-paque plus(ge healthcare;11778538)密度离心从健康供体中分离。将pbmc在以2
×
105个细胞/孔培养在不存在(未刺激的pbs媒介物对照)或存在刺激(lps、肽聚糖、商陆有丝分裂原、pha-l、cpg il-15或cytostim)的情况下在不存在(媒介物)或存在lmw-ds(tikomed ab,viken,sweden,wo 2016/076780)的情况下在三种浓度60μg/ml、200μg/ml和600μg/ml,在37℃、5%co2下培养24小时。离心后,去除细胞培养物上清液并储存在-20℃,等待通过elisa分析il-6。根据制造商的说明,通过elisa(r&d systems;dy206)对上清液中的il-6水平进行定量。
[0638]
结果
[0639]
历史内部数据指导选择刺激浓度,以使用lps、肽聚糖、商陆有丝分裂原、pha-l、cpg il-15和cytostim的亚最大浓度。用pbmc混合物,所有的刺激增加了il-6释放到细胞培
养物上清液中。随着来自前六个供体的细胞在lmw-ds对lps刺激的影响方面产生了有希望的结果,决定在另外六个供体中扩展lps研究。此外,相对于未刺激的pbmc,最初选择的cpg il-15和cytostim浓度使il-6释放增加相对较低,并且因此在来自另外六个供体的细胞中,研究了更高浓度的cpg il-15和cytostim。相对于未刺激的细胞,这些较高的浓度诱发il-6释放的更大增加。
[0640]
lps
[0641]
lps(脂多糖)是一种toll样受体(tlr)4激动剂。在人类pbmc混合物中,由lps直接活化的主要细胞类型是单核细胞,其表达tlr4。单核细胞是先天免疫系统的部分;这些骨髓细胞也可用于模拟对其他骨髓细胞(如巨噬细胞和小胶质细胞)的响应。在本研究中,lps诱发释放到细胞培养物上清液中的il-6显著增加(图17a);lmw-ds引起il-6浓度的浓度依赖性和统计上显著的降低(图17a)。这指示lmw-ds显示出降低单核细胞tlr4活化后il-6的促炎性后果的潜力。
[0642]
肽聚糖
[0643]
肽聚糖是tlr2激动剂,其在pbmc混合物中主要由单核细胞和b淋巴细胞表达;后者是获得性免疫系统的组分,最广为人知的是在产生针对抗原的特异性抗体中发挥不可或缺的作用。肽聚糖诱发释放到细胞培养物上清液中的il-6显著增加,但总体而言,从来自六个供体的细胞的实验来看,几乎没有证据表明lmw-ds(即使在测试的最高浓度下)引起il-6释放的总体减少(图17b)。
[0644]
商陆有丝分裂原
[0645]
商陆有丝分裂原是从美洲商陆(phytolacca americana)中纯化的凝集素。其诱发b淋巴细胞的t淋巴细胞依赖性活化。在本研究中,商陆有丝分裂原通过pbmc混合物诱发il-6释放的大量增加,但是这种释放通常不受lmw-ds影响(图17c)。
[0646]
pha-l
[0647]
pha-l(植物血凝素-l)是来自普通菜豆(phaseolus vulgaris)(红芸豆)的l型亚基凝集素,其与t淋巴细胞表面受体交联,导致其活化。pha-l诱导从pbmc混合物中释放的il-6相对适度增加,并且这以浓度依赖性方式被lmw-ds显著抑制(图17d)。
[0648]
cpg il-15
[0649]
cpg-odn是活化tlr9的短单链dna分子,其在pbmc混合物中主要由单核细胞和b细胞表达。il-15与cpg协同刺激b淋巴细胞。与商陆有丝分裂原不同,cpg il-15直接活化b淋巴细胞,即不依赖于t淋巴细胞。在用来自六个供体的pbmc进行的第一轮实验中,所选择的cpg-odn il-15的浓度仅诱发释放到细胞培养物上清液中的il-6的少量增加。虽然这种相对低水平的il-6释放通常不受lmw-ds影响(图17e),但是用额外供体进行的另外的实验增加了lps刺激的n数,这些相同的额外供体经受了更高浓度的cpg il-15的应用,以试图引起比未刺激细胞可见的明显更稳健的il-6释放;虽然实现了这一点,但仍然没有lmw-ds的一般影响(图17f)。
[0650]
cytostim
[0651]
cytostim是基于抗体的t淋巴细胞活化因子。其与t细胞受体(tcr)结合,并且将其与抗原呈递细胞的主要组织相容性复合物(mhc)分子交联。因此,cytostim刺激cd4和cd8 t淋巴细胞。在本研究中,用来自六个供体的pbmc进行的第一轮实验使用的cytostim浓度仅
6释放到细胞培养物上清液中的浓度依赖性统计上显著的增加(图18d)。肝素在一定程度上反映了这种增强,尽管这种趋势没有达到统计学显著性(图18f)。如所预期,地塞米松的存在导致肽聚糖刺激的il-6释放到细胞培养物上清液中的统计上显著的抑制(图18e)。
[0664]
单核细胞是先天免疫系统的部分;这些骨髓细胞也可用于模拟对其他骨髓细胞(如巨噬细胞和小胶质细胞)的响应。在目前的研究中,lmw-ds抑制lps诱发的纯化单核细胞释放il-6增加的明显和实质性影响提供了这些细胞代表lmw-ds的靶标的强有力的证据。
[0665]
实施例11
[0666]
患病、受损或感染组织中免疫响应的活化通过外周血单核细胞(pbmc)表型平衡的变化来反映。因此,评估研究药物对适应性和先天免疫系统组分的影响可以揭示机制细胞途径,以更好地理解与研究治疗相关的临床变化,以及潜在地鉴定预测不同病理的治疗效果的细胞和/或分子生物标志物。
[0667]
人pbmc可以在体外被多种剂刺激,这些剂将直接和间接活化各种细胞亚群并模拟与受损组织相关的免疫响应。监测pbmc释放的细胞因子可以研究药物的潜在影响并预测药物在特定患者病因中的作用。
[0668]
材料与方法
[0669]
实施例9研究了lmw-ds改变由使用各种刺激物引起的人pbmc分泌il-6的能力。本研究对来自实施例9的细胞培养物上清液进行了更广泛的分析。因此,通过ficoll-paque plus密度离心从健康供体中分离外周血单核细胞(pbmc)。将pbmc以2
×
105个细胞/孔在不存在(未刺激的pbs媒介物对照)或存在刺激(lps、肽聚糖、商陆有丝分裂原、pha-l、cpg il-15或cytostim)的情况下在不存在(媒介物)或存在lmw-ds(tikomed ab,viken,sweden,wo 2016/076780;三种浓度;最高浓度为600μg/ml、200μg/ml和60μg/ml),在37℃、5%co2下培养24小时。
[0670]
因此,对每个pbmc供体的处理如下:
[0671]
i.媒介物
[0672]
ii.刺激
[0673]
iii.刺激 lmw-ds(60μg/ml)
[0674]
iv.刺激 lmw-ds(200μg/ml)
[0675]
v.刺激 lmw-ds(600μg/ml)
[0676]
各自以一式三份进行评估,使得样品总数为:
[0677]
1.lps;165个样品(来自11名供体)
[0678]
2.来自枯草芽孢杆菌(b.subtilis)的肽聚糖;90个样品(来自6名供体)
[0679]
3.pha-l;90个样品(来自6名供体)
[0680]
4. 0.2μm cpg il-15;90个样品(来自6名供体)
[0681]
5. 1.0μm cpg il-15;90个样品(来自6名供体)
[0682]
6.商陆有丝分裂原;90个样品(来自6名供体)
[0683]
7. 10μl/ml cytostim;90个样品(来自6名供体)
[0684]
8. 30μl/ml cytostim;90个样品(来自6名供体)
[0685]
来自所有刺激的上清液样品总数(所有细胞类型)=795
[0686]
处理后,取出细胞培养物上清液,离心并储存于-20℃下,随后解冻以用于使用来
自r&d systems的5-重人磁luminex测定(目录号lxsahm-05)进行多种细胞因子的多重分析。luminex分析完全按照制造商的方案进行。
[0687]
结果
[0688]
历史内部数据指导选择刺激浓度,以使用lps、肽聚糖、商陆有丝分裂原、pha-l、cpg il-15和cytostim的亚最大浓度;使用亚最大浓度的刺激往往允许在存在时所鉴定调制的增加和减少。使用pbmc混合物,所有的刺激都增加了细胞因子向细胞培养物上清液中的释放,尽管一些刺激比其他刺激更有效(图19-23)。
[0689]
lps
[0690]
lps(脂多糖)是一种toll样受体(tlr)4激动剂。在人类pbmc混合物中,由lps直接活化的主要细胞类型是单核细胞,其表达tlr4。单核细胞是先天免疫系统的部分;这些骨髓细胞也可用于模拟对其他骨髓细胞(如巨噬细胞和小胶质细胞)的响应。在本研究中,当评价lps的作用时,ifnγ、tnfα、il-1β和il-10的分泌有适度的增加,但是il-8的分泌有显著的增加(图19-23)。lmw-ds导致ifnγ和il-10的浓度依赖性但较小的减少(图19、23),并且最高浓度的lmw-ds的存在导致il-1β、il-8和tnfα的刺激分泌的适度减少(图20-22)。
[0691]
肽聚糖
[0692]
肽聚糖是tlr2激动剂,其在pbmc混合物中主要由单核细胞和b淋巴细胞表达;后者是获得性免疫系统的组分,最广为人知的是在产生针对抗原的特异性抗体中发挥不可或缺的作用。肽聚糖引起释放到细胞培养物上清液中的il-1β、il-8和tnfα的增加,但是总的来说,从检查来自所有供体的结果来看,几乎没有证据表明lmw-ds(即使在测试的最高浓度下)引起细胞因子释放的普遍减少,尽管存在il-1β和tnfα分泌的相关增加(图19-23)。
[0693]
pha-l
[0694]
pha-l(植物血凝素-l)是来自普通菜豆(phaseolus vulgaris)(红芸豆)的l型亚基凝集素,其与t淋巴细胞表面受体交联,导致其活化。在pbmc混合物中,pha-l诱导ifnγ、il-8、il-10和tnfα的增加,以及il-1β的总体适度增加(图19-23)。lmw-ds导致ifnγ、il-8、tnfα的刺激释放的少量减少,但不导致il-1β的减少(图19-22)。相比之下,lmw-ds导致il-10分泌的大量浓度依赖性减少(图23)。
[0695]
商陆有丝分裂原
[0696]
商陆有丝分裂原是从美洲商陆(phytolacca americana)中纯化的凝集素。其诱发b淋巴细胞的t淋巴细胞依赖性活化。在本研究中,商陆有丝分裂原通过pbmc混合物诱发ifnγ、il-1β、il-8、il-10和tnfα分泌的稳健增加(图19-23),但是这种释放通常不受lmw-ds影响,除了il-10分泌的浓度依赖性减少(图23)。
[0697]
cpg il-15
[0698]
cpg-odn是活化tlr9的短单链dna分子,其在pbmc混合物中主要由单核细胞和b细胞表达。il-15与cpg协同刺激b淋巴细胞。与商陆有丝分裂原不同,cpg il-15直接活化b淋巴细胞,即不依赖于t淋巴细胞。在目前的研究中,只有强有力的证据表明这种刺激增加了il-8的分泌。总的来说,在这些实验中,lmw-ds对这种响应几乎没有影响(图19-23)。
[0699]
cytostim
[0700]
cytostim是基于抗体的t淋巴细胞活化因子。其与t细胞受体(tcr)结合,并且将其与抗原呈递细胞的主要组织相容性复合物(mhc)分子交联。因此,cytostim刺激cd4和cd8 t
one gmbh,kremsmunster,austria)中。在室温下30分钟后,将抽取的血液在1,890
×
g下离心10分钟,并且将得到的血清样品储存在-20℃下直至分析。
[0709]
解冻后,用1ml hplc级乙腈补充每份血清样品的500μl等分试样,涡旋60秒,在台式离心机中以最大速度离心以沉淀蛋白质。用大量hplc级氯仿洗涤上清液以去除有机溶剂,离心,将上层水相转移到不同的管中,清楚地标记以鉴定样品并在-80℃下储存,直到分析以确定不同的水溶性化合物。
[0710]
将每份血清样品的约300μl的第二等分试样避光,然后使用在别处详细描述的方法(lazzarino等人,single-step preparation of selected biological fluids for the high performance liquid chromatographic analysis of fat-soluble vitamins and antioxidants)j chromatogr a.2017;1527:43-52.)进行处理以提取脂溶性抗氧化剂。简而言之,用1ml hplc级乙腈补充样品,剧烈涡旋60秒并在37℃下在水浴中搅拌孵育1小时,以允许完全提取脂溶性化合物。然后将样品在4c下以20,690
×
g离心15分钟,以沉淀蛋白质并将澄清的上清液保存在-80℃下,直到脂溶性维生素和抗氧化剂的hplc分析。
[0711]
在去蛋白血清样品中,通过hplc根据其他地方描述的方法(tavazzi等人,simultaneous high performance liquid chromatographic separation of purines,pyrimidines,n-acetylated amino acids,and dicarboxylic acids for the chemical diagnosis ofinborn errors of metabolism.clin biochem.2005;38:997-1008;romitelli等人,comparison of nitrite/nitrate concentrationin human plasma and serum samples measured by the enzymatic batch griess assay,ion-pairing hplc andion-trap gc-ms:theimportance of a correct removal of proteinsin the griess assay.j chromatogr b analyt technol biomed life sci.2007;851:257-267;amorini等人,metabolic profile of amniotic fluid as a biochemical tool to screen for inborn errors of metabolism and fetal anomalies.mol cell biochem.2012;359:205-216)分离和定量下列水溶性化合物:次黄嘌呤、黄嘌呤、尿酸、丙二醛(mda)、亚硝酸盐、硝酸盐、n-乙酰天冬氨酸(naa)、瓜氨酸(citr)、丙氨酸(ala)和鸟氨酸(orn)。
[0712]
通过hplc根据先前描述的方法(lazzarino等人,cerebrospinal fluid atp metabolites in multiple sclerosis.mult scler j.2010;16:549-554)分离和定量去蛋白血清样品中的以下脂溶性维生素和抗氧化剂:α-生育酚(维生素e)和γ-生育酚。除了上述脂溶性抗氧化剂和维生素之外,还测定血清样品中总胆红素的浓度。
[0713]
统计学
[0714]
通过双尾学生t检验对成对样品进行治疗前和治疗后亚组的比较。每个亚组与对照健康受试者组的比较通过用于不成对观察的双尾非参数曼-惠特尼u检验进行。p《0.05的差异被认为统计上显著的。
[0715]
结果
[0716]
根据统计分析,患者的两个治疗前和治疗后亚组具有显著不同的naa(图26)、尿酸(图27)、mda(图31)、硝酸盐(no3)(图29)、亚硝酸盐 硝酸盐(no3 no2)(图30)、氧嘌呤总量(图28)、ala(图32)、citr(图33)、orn/citr比率(图34)、维生素e(α-生育酚和γ-生育酚)(图35和36)和总胆红素(图43)的血清浓度。
[0717]
在图26-36中,上述化合物的数据显示在箱线图中(报道最小值、最大值、中值、
25%和75%百分位数),其中还包括来自意大利患者队列的历史数据集的对照健康受试者(年龄范围为25-65岁)的值,并且与两组患者(治疗前和治疗后)进行比较。在所有图中,关于对照值的差异由一个星号(*)指示,而两组患者亚组之间的差异由两个星号(**)指示。
[0718]
als患者具有比在健康对照受试者中测得的那些更高的血清naa水平,推测是由于存活神经元的减少(图26)。lmw-ds治疗后测得的naa值显著降低,因此表明药物对细胞存活的保护作用。
[0719]
als患者具有比在对照中测得的那些更高的血清尿酸浓度和氧嘌呤总量(次黄嘌呤 黄嘌呤 尿酸)(图28),这可能是由于能量代谢失衡导致腺嘌呤核苷酸降解途径活化且随后导致循环嘌呤化合物增加。lmw-ds治疗降低了这两个参数,因此表明线粒体功能随着细胞能量状态的增加而改善。
[0720]
als患者具有比在照组中测得的那些更高的硝酸盐(图29)、亚硝酸盐 硝酸盐(图30)和mda(图31)的血清浓度,强烈指示持续的氧化/硝化应激,导致ros介导的脂质过氧化(mda)和一氧化氮代谢(硝酸盐和亚硝酸盐 硝酸盐)的这些稳定终产物的循环水平增加。lmw-ds治疗降低了这些参数的水平,因此指示化合物的直接清除活性,对基因(如bdnf)的积极影响,调控清除酶的水平,和/或对线粒体功能的积极影响,最终导致较低水平的ros产生。
[0721]
als患者具有比在对照中测得的那些更高的血清ala浓度(图32),这可能是因为肌肉蛋白降解速率更高。lmw-ds治疗使ala的循环值正常化(等于对照的值,且显著低于治疗前亚组的值),从而表明药物对肌肉代谢和功能的积极作用。
[0722]
als患者具有比在对照中测得的那些更高的citr血清浓度(图33)和更低的orn/citr比率值(图34),从而证实了由过量的一氧化氮产生和随后的活性氮物质(rns)增加引起的持续硝化应激的存在。lmw-ds治疗改善了这两个参数,可能是通过降低诱导型一氧化氮合酶的表达,所述酶负责触发硝化应激。
[0723]
als患者具有比在对照中测得的那些更高的α-生育酚和γ-生育酚(维生素e的两种主要形式)的血清浓度(图35和36),从而表明主要脂溶性抗氧化剂的显著减少,所述抗氧化剂作为ros介导的膜磷脂的脂肪酸过氧化的阻断物起关键作用。lmw-ds治疗改善了这两个参数。
[0724]
als患者具有比在对照组中测得的那些更高的血清总胆红素浓度,可能反映了肝功能受损。lmw-ds治疗降低了这一参数,再次具有使这种血清代谢物水平正常化的趋势,从而指示肝功能改善。
[0725]
lmw-ds能够改善肝功能,从而抑制肝功能恶化,这在一些严重covid-19患者中可见。因此,lmw-ds治疗的这种效果在恢复期(阶段iv)和长期covid-19受试者中特别有益(图25)。
[0726]
实施例13
[0727]
本实施例研究了使用covid-10刺突-ace2结合测定试剂盒进行评估lmw-ds影响sars-cov-2刺突蛋白与ace2之间的相互作用的能力。
[0728]
covid-19由病毒sars-cov-2的感染引起。现在已经知道,病毒进入人类细胞的机制中的一种是通过sars-cov-2刺突蛋白与血管紧张素i转化酶2(ace2)之间的直接相互作用;后者在各种人类细胞的细胞膜中表达。这种相互作用可以使用covid-19刺
突-ace2结合测定试剂盒进行评估,这是一种基于快速elisa的比色测定。
[0729]
材料与方法
[0730]
covid-19刺突-ace2结合测定试剂盒(目录号cov-sace2)按照制造商的说明(版本1.6)使用,以评估lmw-ds破坏sars-cov-2刺突蛋白与ace2之间的相互作用的能力。
[0731]
以3.0μ/ml至600μg/ml范围内的浓度添加含lmw-ds(tikomed ab,viken,sweden,wo 2016/076780)的盐水。
[0732]
结果
[0733]
有明显的证据表明lmw-ds能够减少sars-cov-2刺突蛋白与ace2之间的相互作用(图37)。更详细地说,lmw-ds引起sars-cov-2刺突蛋白-ace2相互作用的浓度依赖性破坏。预期由lmw-ds引起的sars-cov-2刺突蛋白与ace2之间的相互作用的抑制通过降低sars-cov-2病毒进入人细胞的能力而对患者有益。
[0734]
实施例14
[0735]
使用纵向设计,本研究旨在确定als患者在硫酸葡聚糖施用前和治疗开始后不同时间后选定血清代谢物的变化。所测代谢物的变化指示患者对硫酸葡聚糖的生化响应,这是潜在疾病改变和药物在此als患者群体中的作用机制的基础。
[0736]
材料与方法
[0737]
将硫酸葡聚糖(tikomed ab,viken,sweden,wo 2016/076780)以2mg/kg每周一次通过每日皮下注射施用于8名患有als的人类患者,持续十周。
[0738]
在患者完全休息至少15分钟后,使用标准止血带程序,在硫酸葡聚糖施用之前(第0周)和之后(第5周和第10周)从患者的肘前静脉采集外周静脉血样品,将其放入含有血清分离器和凝块活化剂的单个聚丙烯管(greiner-bio one gmbh,kremsmunster,austria)中。在室温(20-25℃)下30分钟后,使抽取的血液以1,890
×
g离心10分钟以获得血清等分试样。
[0739]
用1ml hplc级乙腈补充500μl的血清等分试样,涡旋60秒,在台式离心机中以最大速度离心以沉淀蛋白质。用大量hplc级氯仿洗涤上清液以去除有机溶剂,离心,并将上层水相转移到不同的管中,清楚地标记以鉴定样品并在-80℃下储存,直到分析以确定不同的水溶性化合物。
[0740]
alsaq-40在治疗前和此后的10周每周和在随访期间问诊时进行评估。
[0741]
结果
[0742]
乳酸由肌肉产生并在正常人和als患者的运动期间积累在血液中。血清乳酸水平升高指示肌肉功能/用途得到改善。
[0743]
图38说明了在硫酸葡聚糖施用之前(第0周)和之后als患者的血清乳酸水平。数据显示,主要来源于肌肉细胞代谢的乳酸的循环水平随着硫酸葡聚糖施用时间的增加而显著升高(*与第0周相比显著不同,p《0.01)。在5周的硫酸葡聚糖治疗后,血清乳酸水平增加了29.8%,从1.78
±
0.59μmol/l增加至2.31
±
1.02μmol/l(p《0.01,威尔科克森符号秩检验(wilcoxon signed-rank test)),而在10周的硫酸葡聚糖治疗后,血清乳酸水平增加了70%,达到3.02
±
1.59μmol/l。因此,硫酸葡聚糖施用导致als患者的肌肉活动增加。
[0744]
标题为
‘
日常生活活动/独立性’(adl)和
‘
身体活动性’(pm)的alsaq-40子评分反映了患者对其身体活动和独立性水平的看法。评分下降反映了身体活动的改善。在10周的硫酸葡聚糖治疗后,adl子评分显著下降18.6%,从58.9
±
21.4降至44.4
±
24.7(p《0.05),见图39,同时pm子评分下降16%,从27.2
±
22.2降至22.7
±
20.2。这些结果指示在治疗期间患者的身体活动得到改善。
[0745]
由硫酸葡聚糖施用诱导的肌肉功能的改善对covid-19患者是有益的,否则这些患者可能会遭受肌肉疼痛和肌肉功能退化。因此,通过硫酸葡聚糖获得的肌肉改善在恢复期(阶段iv)是有益的(图25)。
[0746]
实施例15
[0747]
材料与方法
[0748]
一种呈iia期、单中心、开放标签、单组研究形式的临床试验,其中在中度进展als患者中评价了皮下施用的硫酸葡聚糖的安全性、耐受性和可能的功效。临床试验在瑞典哥德堡萨尔格伦斯卡大学医院(the sahlgrenska university hospital,gothenburg,sweden)进行并由哥德堡大学伦理委员会和瑞典医疗产品局(ethics committee of the university of gothenburg and swedish medical products agency)监督和批准。
[0749]
将硫酸葡聚糖(tikomed ab,viken,sweden,wo 2016/076780)以1mg/kg每周一次通过每日皮下注射施用于13名患有als的人类患者,持续五周。
[0750]
在规定的研究间隔内,通过静脉导管将血样抽取到真空管中。紧接着在采集后由萨尔格伦斯卡大学医院的临床化学实验室(clinical chemistry laboratory at the sahlgrenska university hospital)进行血液血浆的实验室分析。
[0751]
alsfrs-r在治疗前和此后的5个处理周内每周和在随访期间问诊时进行评估。
[0752]
结果
[0753]
肌红蛋白是通常在心脏和骨骼肌组织中发现的蛋白质。当损伤/疾病损害肌肉时,血流中的肌红蛋白水平升高。血清肌红蛋白水平降低指示肌肉退化减少。
[0754]
在4周的硫酸葡聚糖治疗后,来自患者的血清肌红蛋白数据显示肌红蛋白水平的统计上显著的30%降低,从133.92
±
126.28μg/l降低至103.69
±
72.16μg/l(相较于第1天,p=0.021),从而指示在治疗期间患者的肌肉组织退化和肌肉萎缩的速率与药物相关地降低,参见图40。
[0755]
血液中出现肌酶肌酸激酶通常被认为是肌肉损害的生物标志物,并且对于包括als在内的涉及肌肉萎缩的医学病状的诊断特别有用。肌酸激酶水平升高是als患者的常见特征。血清肌酸激酶水平降低指示疾病相关肌病得到缓解。
[0756]
在4周的硫酸葡聚糖治疗后,来自患者的血清肌酸激酶数据显示水平的统计上显著的13.3%降低,从7.15
±
5.74μkat/l降低至6.2
±
5.08μkat/l(p《0.05,威尔科克森符号秩检),从而指示治疗期间患者肌肉萎缩的药物相关减少,参见图41。
[0757]
肝细胞生长因子(hgf)是天然存在的生长因子,其作为一种有效的神经保护剂和生肌剂并在许多动物模型(包括als模型)中显示可用于对抗退行性疾病进展。有趣的是,hauerslev等人(2014,plos one 9:e100594)在肌肉萎缩的小鼠模型中进行了2周的重组hgf治疗后,肌肉质量增加了18%。观察到hgf治疗能够在这种肌萎缩动物模型的骨骼肌中诱导如此快速的再生响应,这与在als患者中观察到的响应于硫酸葡聚糖的快速肌肉响应
相关。
[0758]
在硫酸葡聚糖注射后,als患者的药代动力学数据显示循环hgf的药理学相关水平从820
±
581μg/l至2.5小时的峰值37863
±
14235μg/l的统计上显著的增加(p《0.001),参见图42。这指示在硫酸葡聚糖施用后,对肌肉萎缩的直接肌原性以及间接神经营养性hgf介导的作用的潜力。
[0759]
肌肉退化减少的生化证据支持肌肉功能改善的临床观察。例如,在alsfrs-r中,由颈部、躯干、腰骶部和呼吸肌介导的功能分别通过3个项目进行评估,并且这些类别中的评分与肌肉强度的客观测量值非常一致。值得注意的是,在五周的治疗期间,两名患有严重延髓麻痹的患者经历该症状的几乎完全消退。
[0760]
由硫酸葡聚糖施用诱导的肌肉功能的改善对covid-19患者是有益的,否则这些患者可能会遭受肌肉疼痛和肌肉功能退化。因此,通过硫酸葡聚糖获得的肌肉改善在恢复期(阶段iv)是有益的(图25)。此外,在covid-19患者中,hgf活化的组织修复和伤口愈合的水平增加,这在肺期(阶段ii)是相关的(图25)。
[0761]
上述实施方案应被理解为本发明的几个说明性实施例。本领域技术人员将理解,在不脱离本发明的范围的情况下,可以对实施方案进行各种修改、组合和改变。特别是,在技术上可能的情况下,不同实施方案中的不同部分解决方案可以组合成其他配置。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。