对btn2具有特异性的抗体及其用途
技术领域
1.本发明涉及结合btn2a1、将巨噬细胞群向抗肿瘤m1巨噬细胞转移并直接激活nk细胞以及对癌细胞具有细胞毒性的抗btn2a1激活抗体。或者,或组合地,所述抗体可激活vγ9/vδ2t细胞。此类抗体特别适用于治疗癌症。
背景技术:
2.巨噬细胞表现出从经典激活的m1到替代的m2类型的不同表型。m1巨噬细胞在迁移后迅速从单核细胞分化,被细菌衍生产物(如lps(脂多糖))以及与感染相关的信号(如ifnγ)激活。它们是高度炎症的,具有高吞噬和杀菌潜力。它们分泌重要的促炎细胞因子,如tnfα、il
‑
1、il
‑
6和il
‑
12以及活性氧。相反,当肉芽组织形成发生时,m2巨噬细胞存在于愈合过程的后期;它们拮抗炎症反应,从而启动愈合。这些抗炎细胞募集成纤维细胞并激活它们以分化成肌成纤维细胞,该肌成纤维细胞释放促血管生成因子以募集内皮祖细胞并使新血管形成,这一过程通过关键抗炎细胞因子il
‑
4、il
‑
10和il
‑
13的分泌而发生,并且还与ros、一氧化氮(no)和tnfα的产生减少有关。
3.肿瘤微环境(tme)将巨噬细胞强烈极化为m2样表型,特别是在肿瘤从癌症治疗中恢复的情况下。这种极化不仅具有高度的促血管生成作用,而且是免疫抑制的。因此,在大多数实体癌中,肿瘤相关巨噬细胞(tam)浸润的增加长期以来一直与患者预后不良有关,突出了它们作为癌症潜在诊断和预后生物标志物的价值。
4.因此,重编程和选择性杀死m2巨噬细胞被认为是一种有前途的治疗策略(zhu y et al.cancer res 2014)。
5.此外,作为先天免疫细胞,自然杀伤(nk)细胞在癌症免疫监督中发挥关键作用。nk细胞无需事先致敏即可清除多种异常或应激细胞,甚至优先杀死干细胞样细胞或癌症干细胞。一旦与靶细胞形成免疫突触,nk细胞释放预先形成的溶细胞颗粒,包括穿孔素和颗粒酶,其功能是诱导细胞裂解。
6.基于该假设,若干研究已经成功利用nk细胞的过继转移对抗各种肿瘤,尤其是血液系统恶性肿瘤。但是,癌症采用各种策略来延迟、改变甚至停止抗肿瘤免疫,导致对肿瘤生长的控制失败。nk细胞的抗肿瘤反应也面临很多限制。特别地,肿瘤微环境(tme)仍然是nk细胞,尤其是过继转移的nk细胞有效性的主要障碍。例如,肿瘤浸润免疫细胞如树突状细胞(dc)、抑制性或致耐受性巨噬细胞和调节性t(treg)细胞以及嵌入细胞外基质中的癌症相关成纤维细胞,可能通过分泌免疫抑制细胞因子或干扰受体表达而干预nk细胞的活化。例如,在tme中,tgf
‑
β(特别是由m2极化巨噬细胞分泌)被认为是nk细胞的主要抑制细胞因子,它限制了nk细胞的数量和抗转移功能。
7.因此,从治疗的角度,具有可以通过i)抑制肿瘤环境的免疫抑制作用和ii)直接触发nk细胞活化和介导的细胞毒性来刺激抗肿瘤活性的工具将是非常有价值的。
8.嗜乳脂蛋白构成跨膜蛋白家族,其包含嗜乳脂蛋白(btn)、btn样(btnl)以及上皮内t细胞(skint)蛋白的选择和维持(arnett and viney,nat rev immunol 2014)。它们的
胞外部分含有与b7共刺激分子的相应结构域表现出同源性的igv样和igc样结构域(arnett and viney,2014),因此嗜乳脂蛋白被认为是扩展的b7或ig超家族成员。
9.嗜乳脂蛋白(btn)基因家族由人的13种基因组成,形成8个不同的组(abeler
‑
dorner et al.trends immunol 2012;afrache et al.immunogenetics 2012)。七种人btn基因聚集在6号染色体的mhc i类区域,并分为三个亚家族,其形成系统发育相关的组:btn1、btn2和btn3。btn1亚家族仅包含原型单拷贝btn1a1基因,而btn2和btn3亚家族各自包含三种基因btn2a1、btn2a2和btn2a3(其为假基因),分别为btn3a1、btn3a2和btn3a3。已经描述了btn基因家族成员的若干基因多态性,这些基因与不同的疾病有关,包括高血压、慢性肾功能衰竭、包涵体肌炎、1型和2型糖尿病或hcv感染(chen et al.int j clin exp pathol,2015;horibe et al.am j hypertens,2011&2014;milman et al.clin respir,2011;murakata et al.biomed rep,2014;oguri et al.j med genet,2013;pacheco et al.orphanet j rare dis,2016)。已经描述了btnl2、btn2a1、btn3a2和btn3a3的单核苷酸变体,以及涉及btnl3和btnl8的有害拷贝数变异(aigner et al.bmc genet,2013)。
10.btn1a1作为最早鉴定的嗜乳脂蛋白,是乳脂球的形成、分泌和稳定所需要的(ogg et al.proc natl acad sci,2004)。随后,有人提出b7基因和mhc i类和ii类基因可能具有共同的祖先基因,并且可以编码参与类似功能(诸如t细胞活化)的蛋白质(rhodes et al.genomics,2001;harly c et al.blood 2012)。与btn3a1和btn3a3类似,btn2a1和btn2a2蛋白亚型显示igv和igc胞外结构域、跨膜结构域和特征性胞内b30.2结构域,但btn3a2不显示。在小鼠中,btn2a2是人btn2a2基因的单拷贝基因和直系同源物。重组人btn2a1
‑
fc蛋白显示,btn2a1的特定糖型结合树突细胞(dc)上发现的凝集素分子dc
‑
sign。btn2a1与dc
‑
sign的结合取决于通过肿瘤细胞中表达时蛋白质的高甘露糖糖基化作用(malcherek et al.j immunol,2007)。
11.越来越多的证据随后表明,嗜酪蛋白在免疫系统中发挥不同的作用。来自基因型组织表达项目(the gtex consortium,2013)的53个人组织样品的rna
‑
seq显示正常组织中普遍存在btn2a1转录物表达。使用基因表达谱交互分析(tang,z.et al.nucleic acids res,2017)比较来自gtex的rna
‑
seq数据和来自癌症基因组图谱(tcga)数据库的数据表明btn2a1转录物表达在包括以下的若干种癌症中的调节:宫颈鳞状细胞癌和宫颈腺癌、肺小细胞癌、卵巢癌、胰腺癌和子宫内膜癌。
12.之前已经报道了识别btn2a的两种亚型的抗体(wo2019057933),但是所述抗体抑制活化的vγ9/vδ2t细胞产生ifn
‑
γ和/或tnf
‑
α,和/或抑制活化的vγ9/vδ2t细胞的溶细胞功能,和/或抑制活化的vγ9/vδ2t细胞的增殖。
13.vγ9/vδ2t细胞是免疫防御的重要效应子。它们直接裂解感染的病原体或异常细胞。此外,它们通过诱导树突状细胞(dc)成熟以及同型转换和免疫球蛋白生成来调节免疫应答。免疫系统的这一重要细胞平台由表面受体、趋化因子和细胞因子严格调控。vγ9/vδ2t细胞由非肽磷酸化类异戊二烯途径代谢物(称为磷酸激动剂(pag))激活。
14.开发靶向btn2a1和激活免疫细胞,尤其是一种以上的免疫细胞区室,例如巨噬细胞、nk和/或γδt细胞,尤其是vγ9/vδ2t细胞的抗体,可能特别适用于癌症治疗和感染性疾病的治疗。
15.发明概述
16.本公开提供了结合btn2a(特别是结合btn2a1亚型—例如,人btn2a1多肽)的首次抗体,其表现出至少一个以下特性:
17.i.抑制单核细胞向m2巨噬细胞极化,
18.ii.诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
19.iii.直接触发nk细胞激活,
20.iv.增强nk细胞介导的细胞毒性。
21.特别地,本公开的抗体表现出特性i)和ii)中的至少一个,以及特性iii)和iv)中的至少一个,更特别地,本公开的抗体表现出特性i)
‑
iv)。
22.因此,根据本公开的此类抗体可以
23.‑
通过将巨噬细胞表型和功能转向促炎m1巨噬细胞,导致促炎细胞因子的分泌而有利于抗肿瘤微环境,和/或
24.‑
直接触发nk细胞激活,从而进一步增强其溶细胞活性。
25.因此,根据本公开的此类抗体代表可用于癌症治疗的各种策略中的强大工具。
26.在具体实施方案中,本公开的抗btn2a抗体与以下任一竞争结合btn2a:
27.‑
参考鼠抗体mab 101g5,其包含(i)包含seq id no:19的氨基酸序列的重链可变区和(ii)包含seq id no:20的氨基酸序列的轻链可变区,或
28.‑
参考鼠抗体mab 107g3,其包含(i)包含seq id no:1的氨基酸序列的重链可变区和包含seq id no:2的氨基酸序列的轻链可变区。
29.在一些实施方案中,本公开的抗btn2a抗体结合包含位于以下位置的氨基酸残基的表位:
30.‑
seq id n
°
17的第65、68、69、72、78、84、85、95、97、100位,或
31.‑
seq id n
°
17的第212、213、218、220、224、229位。
32.在具体实施方案中,本公开的抗btn2a抗体包含:
33.‑
包含seq id no:3的重链可变区cdr1、包含seq id no:4的重链可变区cdr2、包含seq id no:5的重链可变区cdr3、包含seq id no:6的轻链可变区cdr1、包含seq id no:7的轻链可变区cdr2和包含seq id no:8的轻链可变区cdr3,或
34.‑
包含seq id no:21的重链可变区cdr1、包含seq id no:22的重链可变区cdr2、包含seq id no:23的重链可变区cdr3、包含seq id no:24的轻链可变区cdr1、包含seq id no:25的轻链可变区cdr2和包含seq id no:26的轻链可变区cdr3。
35.在具体实施方案中,本公开的抗btn2a抗体包含:
36.‑
包含与seq id no:1的氨基酸序列具有至少90%同一性的序列的重链可变区和包含与seq id no:2的氨基酸序列具有至少90%同一性的序列的轻链可变区,或
37.‑
包含与seq id no:19的氨基酸序列具有至少90%同一性的序列的重链可变区和包含与seq id no:20的氨基酸序列具有至少90%同一性的序列的轻链可变区。
38.在一个具体实施方案中,本公开的抗体进一步表现出至少一种以下特性:
39.‑
激活来自vγ9vδ2t细胞的溶细胞分子的分泌,
40.‑
激活vγ9vδ2t细胞的溶细胞功能,和/或
41.‑
激活vγ9vδ2t细胞的增殖。
42.最特别地,根据此类实施方案的抗btn2a抗体典型地可以与参考鼠抗体mab 107g3
竞争结合btn2a,所述参考鼠抗体mab 107g3包含(i)包含seq id no:1的氨基酸序列的重链可变区和(ii)包含seq id no:2的氨基酸序列的轻链可变区。
43.更具体地,此类抗btn2a抗体可包含:
44.‑
包含seq id no:3的重链可变区cdr1、包含seq id no:4的重链可变区cdr2、包含seq id no:5的重链可变区cdr3、包含seq id no:6的轻链可变区cdr1、包含seq id no:7的轻链可变区cdr2和包含seq id no:8的轻链可变区cdr3,或
45.‑
包含与seq id no:1的氨基酸序列具有至少90%同一性的序列的重链可变区和包含与seq id no:2的氨基酸序列具有至少90%同一性的序列的轻链可变区,或
46.‑
包含与seq id no:1的氨基酸序列具有至少90%同一性的序列的重链可变区和包含与seq id no:2的氨基酸序列具有至少90%同一性的序列的轻链可变区。
47.在本公开的一些实施方案中,本公开的抗体对btn2a1具有特异性。
48.在具体实施方案中,本公开的抗btn2a抗体是人抗体、嵌合抗体或人源化抗体。
49.本公开还涵盖编码上述抗btn2a抗体的重链和/或轻链的核酸分子。
50.本公开还涉及包含此类核酸的宿主细胞,特别用于制备上述任一种抗btn2a抗体。
51.本公开的另一方面涉及用于治疗,特别是用于治疗癌症或感染性疾病的如上所述的抗btn2a。
52.典型地,根据本公开的癌症包括血癌和实体癌,包括宫颈癌(如鳞状细胞癌和宫颈腺癌)、卵巢癌;皮肤癌,包括鳞状细胞癌和黑色素瘤;肺癌,包括肺小细胞癌;前列腺癌;结肠癌;胰腺癌和子宫内膜癌,更具体地:鳞状细胞癌和宫颈腺癌、鳞状细胞癌、卵巢癌、肺小细胞癌、前列腺癌、结肠癌、胰腺癌和子宫内膜癌。
53.本公开还涉及药物组合物,其包含如上所述的抗btn2a抗体和至少药学上可接受的载体。
54.本公开还提供用于在受试者中激活免疫应答的方法,包括向受试者施用有效量的本文公开的抗btn2a。
55.发明详述
56.定义
57.如本文所用,术语“btn2”具有本领域的一般含义,并且是指包括seq id no:17的btn2a1或seq id no:18的btn2a2的人btn2多肽。
58.seq id no:17:btn2a亚型1前体(智人):
59.mesaaalhfsrpasllllllslcalvsaqfivvgptdpilatvgenttlrchlspeknaedmevrwfrsqfspavfvykggrerteeqmeeyrgrttfvskdisrgsvalvihnitaqengtyrcyfqegrsydeailhlvvaglgskplismrghedggirlecisrgwypkpltvwrdpyggvapalkevsmpdadglfmvttaviirdksvrnmscsinntllgqkkesvifipesfmpsvspcavalpiivvilmipiavciywinklqkekkilsgekeferetreialkelekervqkeeelqvkeklqeelrwrrtflhavdvvldpdtahpdlflsedrrsvrrcpfrhlgesvpdnperfdsqpcvlgresfasgkhywevevenviewtvgvcrdsverkgevllipqngfwtlemhkgqyravsspdrilplkeslcrvgvfldyeagdvsfynmrdrshiytcprsafsvpvrpffrlgcedspificpaltgangvtvpeegltlhrvgthqsl
60.seq id no:18:btn2a亚型2前体(智人):
61.mepaaalhfslpasllllllllllslcalvsaqftvvgpanpilamvgenttlrchlspeknaedmevrwfrsqfspavfvykggrerteeqmeeyrgritfvskdinrgsvalvihnvtaqengiyrcyfqegrsydeailrlvv
aglgskplieikaqedgsiwlecisggwypepltvwrdpygevvpalkevsiadadglfmvttaviirdkyvrnvscsvnntllgqeketvifipesfmpsaspwmvalaviltaspwmvsmtvilavfiifmavsiccikklqrekkilsgekkveqeekeiaqqlqeelrwrrtflhaadvvldpdtahpelflsedrrsvrrgpyrqrvpdnperfdsqpcvlgwesfasgkhywevevenvmvwtvgvcrhsverkgevllipqngfwtlemfgnqyralssperilplkeslcrvgvfldyeagdvsfynmrdrshiytcprsaftvpvrpffrlgsddspificpaltgasgvmvpeeglklhrvgthqsl
62.如本文所用,术语“抗体”或“免疫球蛋白”具有相同的含义,并且将在本公开中同等地使用。
63.本文所用的术语“抗体”是指免疫球蛋白分子和免疫球蛋白分子的免疫活性部分,即含有免疫特异性结合抗原的抗原结合位点的分子。如此,术语抗体不仅涵盖完整的抗体分子,还涵盖抗体片段以及抗体和抗体片段的变体(包括衍生物)。
64.如本文所用的术语“抗体”还包括双特异性或多特异性分子。可以将抗体衍生化或连接到另一个功能分子,例如另一个肽或蛋白质(例如,另一个抗体或受体的配体)以产生结合至少两个不同结合位点或靶分子的双特异性分子。实际上,可以将抗体衍生化或连接至一种以上的其他功能分子以产生结合两种以上不同结合位点和/或靶分子的多特异性分子;此类多特异性分子也旨在涵盖于本文所用的术语“双特异性分子”中。为了产生双特异性分子,本发明的抗体可以功能性连接(例如,通过化学偶联、基因融合、非共价结合或其他方式)至一个或多个其他结合分子,例如另一抗体、抗体片段、肽或结合模拟物,从而产生双特异性分子。此外,对于双特异性分子是多特异性的实施方案,除了第一和第二靶标表位之外,该分子还可以包括第三结合特异性。在一个实施方案中,本文公开的双特异性分子包含作为结合特异性的至少一种抗体或其抗体片段,包括例如fab、fab'、f(ab')2、fv、unibody或单链fv。抗体也可以是轻链或重链二聚体,或其任何最小片段,例如如ladner等人美国专利号4,946,778所述的fv或单链构建体。
65.可用于本文公开的双特异性分子的其他抗体是鼠、嵌合和人源化单克隆抗体(mab)。
66.本公开的双特异性分子可以通过使用本领域已知的方法结合组分结合特异性来制备。例如,每一个彼此结合。当结合特异性是蛋白质或肽时,多种偶联剂或交联剂可用于共价缀合。交联剂的实例包括蛋白质a、碳二亚胺、n
‑
琥珀酰亚胺
‑
s
‑
乙酰硫代乙酸酯(sata)、5,5'
‑
二硫代双(2
‑
硝基苯甲酸)(dtnb)、邻苯二马来酰亚胺(opdm)、n
‑
琥珀酰亚胺
‑3‑
(2吡啶基二硫代)丙酸酯(spdp)和4
‑
(n马来酰亚胺基甲基)环己烷
‑1‑
羧酸磺基琥珀酰亚胺酯(磺基
‑
smcc)(karpovsky et al.,1984;liu et al.,1985)。其他方法包括brennan et al.,1985;glennie et al.,1987;paulus,1985中描述的方法。或者,两种结合特异性都可以在同一载体中编码,并在同一宿主细胞中表达和组装。当双特异性分子是mab x mab、mab x fab、fab x f(ab')2或配体x fab融合蛋白时,该方法特别有用。本公开的双特异性分子可以是包含一个单链抗体和结合决定簇的单链分子,或包含两个结合决定簇的单链双特异性分子。双特异性分子与其特异性靶标的结合可以通过例如酶联免疫吸附测定(elisa)、放射免疫测定(rea)、facs分析、生物测定(例如生长抑制和细胞凋亡)或蛋白质印迹测定来确认。这些测定中的每一个通常通过使用对目标复合物特异的标记试剂(例如,抗体)来检测特定的目标蛋白质
‑
抗体复合物的存在。
67.在天然抗体中,两条重链通过二硫键相互连接,并且每条重链通过二硫键与轻链
manual,cold spring harbor laboratory press,cold spring harbor,n.y.,1988),coligan et al.,eds.,current protocols in immunology,greene publishing assoc.and wiley interscience,n.y.,(1992,1993),and muller,meth.enzymol.92:589
‑
601(1983),这些参考文献通过引用整体并入本文。用于测定抗体的k
d
的方法是通过使用表面等离振子共振,或使用诸如系统的生物传感器系统(还参见有关亲和力评估的详细信息rich rl,day ys,morton ta,myszka dg.high
‑
resolution and high
‑
throughput protocols for measuring drug/human serum albumin interactions using.anal biochem.2001)或系统。平台基于生物层干涉测量(bli)技术。bli技术的原理基于从两个表面反射的白光的光学干涉图—一层固定蛋白和一个内部参考层。固定在生物传感器尖端表面上的配体与溶液中的分析物之间的结合会增加生物传感器尖端的光学厚度,这导致以纳米为单位测量的干涉图案的偏移。波长偏移(δλ)是生物层光学厚度变化的直接度量,当在一段时间内测量这种偏移并将其幅度绘制为时间的函数时,可以获得经典的结合/解离曲线。这种相互作用是实时测量的,允许监测结合特异性、结合速率和解离速率以及浓度(参见abdiche et al.2008以及结果中的详细信息)。亲和力测量通常在25℃下进行。
74.如本文所用的术语“k
assoc”或“k
a”或“k
on”旨在是指特定抗体
‑
抗原相互作用的结合速率,而如本文所用的术语“k
dis”或“k
d”或k
off
旨在是指特定抗体
‑
抗原相互作用的解离速率。
75.如本文所用,术语“单克隆抗体”或“单克隆抗体组合物”是指单一分子组成的抗体分子的制剂。单克隆抗体组合物显示出对特定表位的单一结合特异性和亲和力。
76.如本文所用,术语“特异性”是指抗体可检测地结合抗原(诸如本公开中的btn2a亚型,包括btn2a1和btn2a2)上呈递的表位的能力。它典型地旨在是指以10nm或更小、5nm、1nm或更小、100pm或更小或10pm或更小的k
d
结合人btn2a,特别是btn2a1的抗体或蛋白质。典型地,k
d
为10
‑3pm
‑
10nm,特别是0.1pm
‑
10nm,特别是0.1pm
‑
5nm,或1pm
‑
10nm,特别是1pm
‑
5nm或10pm
‑
5nm。典型地,本公开的抗体对btn2a1或btn2a1和btn2a2两者具有特异性,并且具有如上定义的k
d
。在一些实施方案中,还可以通过在细胞系(例如hek
‑
293t细胞系)中表达btn2a1并通过用增加浓度(例如从5ng/ml至75μg/ml)的如本文公开的纯化的抗
‑
btn2a1 mab,或用阴性对照如其对照同种型对转染细胞进行染色来评估特异性。平均荧光强度数据的非线性回归分析允许将ec
50
测定为观察到最大荧光的50%的mab浓度。典型地,根据本公开,btn2a1特异性抗体与btn2a1结合的ec
50
小于50μg/ml,特别是小于40μg/ml(参见实施例部分),特别是0.1μg/ml
‑
50μg/ml或0.5μg/ml
‑
20μg/ml。
77.短语“识别抗原的抗体”和“对抗原具有特异性的抗体”在本文中与术语“特异性结合抗原的抗体”可互换使用。
[0078]“选择性结合”典型地是指抗体与靶标(例如表位)的结合更强,与另一个靶标的结合相比,它是特异性的。如果抗体对第一靶标的亲和力高于其对第二靶标的亲和力,则抗体与第一靶标的结合强于第二靶标。典型地,如果抗体以低于平衡解离常数或ec
50
的平衡解离常数(k
d
)或ec
50
与第一靶标结合,则抗体与第一靶标的结合强于第二靶标。最具体地,试剂在相关程度上根本不结合第二靶标。在本技术的一些实施方案中,抗体对btn2a并且最特别
地对btn2a1具有选择性。特异性还可以进一步通过例如在抗原的特异性结合相对于与其他无关分子(例如特异性抗原是btn2a多肽,尤其是btn2a1,并且其他“无关分子”例如可以是btn3亚型)的非特异性结合中约10:1、约20:1、约50:1、约100:1、10000:1或更高的亲和力之比来表现。
[0079]
可以使用对密切相关的其他蛋白质(例如btn3亚型)与预期靶蛋白(btn2a亚型)进行比较的交叉反应性测定来测试本文公开的抗体的选择性。当无法检测这种交叉反应时,同时在相同的抗体稀释度下给出预期靶标的强信号时,典型地认为该抗体具有选择性(参见对应于图3的实施例中详述的结果)。“与抗原交叉反应”的抗体旨在是指以10nm或更小、1nm或更小或100pm或更小的k
d
结合该抗原的抗体。“不与特定抗原交叉反应”的抗体旨在是指以100nm或更大的k
d
、或1μm或更大的k
d
或10μm或更大的k
d
结合该抗原的抗体。在某些实施方案中,不与抗原交叉反应的此类抗体在标准结合测定中表现出对这些蛋白质基本上不可检测的结合(典型地参见表3或图3a)。
[0080]
在具体实施方案中,本公开的抗btn2a1抗体与如下定义的序列seq id n
°
35的食蟹猴btn2a1直系同源物(cynobtn2a1;ncbi参考号xp_015304392.1)交叉反应:
[0081]
mqrqfskasrpclpwvlmepaaalhfslpaslillllllrlcalvsaqftvvgptdpilamvgenttlrchlspeknaedmevrwfrsqfspavfvykggrerteeqmeeyrgrttfvskdisrgsvaliihnvtaqengtyrcyfqegrsydeailhlmvaglgskplvemrghedggirlecisrgwypkpltvwrdpygrvvpalkevfppdtdglfmvttaviirdksmrnmscsisdtllgqkkesvifipesfmpsvspcvvalpiivvflmiiiavciywinrlqketkilsgekeserktreiavkelkkervqkekelqvkeqlqeelrwrrtvlhavdvvldpdtahpdlllsedrrsvrrcplghlgesvpdnperfnsepcvlgresfasgkhywevevenviewtvgvcrdsverkeevllrprngfwtlemckgqyralsspkrilplkeslcrvgvfldyeagdvsfynmrdrshiytcprlafsvpvrpffrigsddspificpaltgasgitvpeeglilhrvgtnqslmpvgtrcyghgmrptgfirmreergihrttreerepdmqnfdlgahwsnnlpsarsreflnsdlvpdhslespvtpglanktgepqaevtclcfslpsselrafpstatnhnhkatalgsdlhievkgyedggihlecrstgwypqpqiqwsntkgqhipavkapvvadgvglyavaasvimrgssgegvsciirnsllglektasisitdpffrnaqpwiaalagtlpisllllagasyflwrqqkekialsretereremkemgyaatkqeislrggekslayhgthisylaaperwemavfpnsglprclltlillqlpkldsapfdvigppepilavvgedaelpcrlspnasaehlelrwfrkkvspavlvhrdgreqeaeqmpeyrgratlvqdgiaegrvalrirgvrvsddgeytcffredgsyeealvhlkvaalgsdphismqvqengeiwlectsvgwypepqvqwrtskgekfpstsesrnpdeeglftvaasviirdtsvknvscyiqnlllgqekeveifipg。
[0082]
在这些实施方案的一些中,本文公开的抗体以与hubtn2a上获得的相应ec
50
相当(大于或小于10%)的ec
50
结合cynobtn2a1胞外域。
[0083]
术语“同一性”是指两个多肽分子之间或两个核酸分子之间的序列相似性。当两个比较序列中的一个位置被相同的碱基或相同的氨基酸残基占据时,则相应的分子在该位置是相同的。两个序列之间的同一性百分比对应于两个序列共享的匹配位置数除以比较的位置数再乘以100。通常,当将两个序列比对以获得最大同一性时进行比较。可以通过使用例如gcg(genetics computer group,program manual for the gcg package,第7版,madison,wisconsin)堆积程序或任何序列比较算法(如blast、fasta或clustalw)进行比对来计算同一性。
[0084]
根据本公开的参考分子的功能性变体表现出基本上等于或优于参考分子(例如:
107g3 mab)的相应功能特性的功能特性。基本上等于在本文中意指所述功能性变体保留参考分子的相应功能特性的至少约50%、60%、70%、80%、90%、95%或100%。
[0085]
一方面,本公开涉及对如上定义的btn2a1具有特异性的抗体。典型地,这种抗体的特征在于它以如上定义的10nm或更小的k
d
结合人btn2a1。
[0086]
在一些实施方案中,本公开的抗体不与btn3亚型交叉反应。
[0087]
根据本发明,对btn2a,特别是btn2a1具有特异性的抗体的特征还典型地进一步在于其具有至少一个以下特性:
[0088]
i.其抑制单核细胞向m2巨噬细胞极化,
[0089]
ii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
[0090]
iii.其直接触发nk细胞激活,
[0091]
iv.其增强nk细胞介导的细胞毒性。
[0092]
优选地,本公开的抗体表现出特性i和ii的至少一个,以及特性iii和iv的至少一个,最可能地,它表现出特性i)
‑
iv)。
[0093]
可以使用例如实施例部分中详述的测定在抗btn2a抗体中筛选具有这种有利特性的本公开的抗btn2a抗体。下文简要描述所述测定及其实施。
[0094]
在巨噬细胞极化测定中,如在本领域所经典进行的,在抗btn2a抗体,特别是之前定义的抗btn2a1抗体存在下,或阴性对照(如其对照同种型)存在下,m2巨噬细胞从巨噬细胞中产生。在m
‑
csf(巨噬细胞集落刺激因子)诱导的m2极化期间,可以将gm
‑
csf(粒细胞
‑
巨噬细胞集落刺激因子)和ifn
‑
γ添加到单核细胞中,作为m2分化抑制的对照。相反,在gm
‑
csf存在下极化的m1巨噬细胞也可以典型地用作表型对照。极化后,m1和m2相关标志物在质膜上的表达可以通过流式细胞术在所得巨噬细胞中评估。
[0095]
根据本公开,可以方便地检测的m1标志物非限制性地包括pdl1、cd86、cd40、cd80和/或socs3。典型地,检测pdl1和/或cd86。根据本公开,可以方便地检测的m2标志物非限制性地包括cd14、cd163、cd206和cd209,典型地,检测cd14和/或cd163。
[0096]
典型地,当:
[0097]
‑
与其对照同种型相比,当在本公开的抗btn2a抗体存在下产生m2巨噬细胞时,观察到至少一种m1标志物的显著增加;和/或
[0098]
‑
与其阴性对照(如其对照同种型)相比,当在抗btn2a抗体存在下产生m2巨噬细胞时,观察到至少一种m2标志物的显著减少时,
[0099]
本公开的抗btn2a诱导对单核细胞向m2巨噬细胞极化的抑制。
[0100]
此外,典型地通过使用适当的商业试剂盒(例如,elisa样试剂盒),可以在培养上清液中定量作为m1和m2巨噬细胞之间特征性区分功能的细胞因子分泌谱(尤其包括m2相关抗炎il
‑
10细胞因子和促炎m1相关tnfα细胞因子)。也可以容易检测以表征m1或m2巨噬细胞表型的其他细胞因子,包括m1相关细胞因子的il
‑
1、il
‑
6、il
‑
12和il
‑
23,以及m2相关细胞因子的tgfβ和il
‑
10。
[0101]
因此,在一些实施方案中,当:
[0102]
‑
与阴性对照(如其对照同种型)相比,当在本公开的抗btn2a抗体存在下培养m2巨噬细胞时,观察到至少一种m2相关细胞因子的分泌显著减少,和/或
[0103]
‑
与阴性对照(如其对照同种型)相比,当在本公开的抗btn2a抗体存在下培养m2巨
噬细胞时,观察到至少一种m1相关细胞因子的分泌显著增加时,
[0104]
单核细胞向m2巨噬细胞极化的抑制也可以被认为是由本公开的抗btn2a抗体的作用的结果。
[0105]
典型地,根据本公开的抗btn2a抗体可以剂量依赖性地抑制单核细胞向m2巨噬细胞的极化,如通过m2相关标志物(例如cd14和/或cd163)的表达减少和/或m2相关细胞因子(如il
‑
10)的分泌减少所评估。此类抗体关于所述至少一种m2相关细胞因子的分泌或所述至少一种m2相关标志物的表达的半数最大抑制浓度(ic
50
)可以在剂量
‑
反应曲线中测定,如实施例部分所详述。在一些具体实施方案中,本发明的抗btn2a抗体表现出:
[0106]
‑
m2相关标志物表达(典型地为cd14和/或cd163)的ic
50
为0.05μg/ml,特别是0.1μg/ml
‑
100μg/ml,特别是50μg/ml,和/或
‑
m2相关细胞因子分泌(典型地为il
‑
10)的ic
50
为0.01μg/ml,特别是0.05μg/ml
‑
100μg/ml,特别是50μg/ml,最特别是0.1
‑
20μg/ml。
[0107]
此外,或可选地,尽管存在促进m2的刺激,根据本公开的抗btn2a抗体可以剂量依赖性地扭转单核细胞向m1巨噬细胞的极化,如通过m1相关标志物(例如cd86和/或pdl1)的表达增加和/或m2相关细胞因子(如il
‑
10)的分泌减少所评估。此类抗体关于所述至少一种m1相关细胞因子的分泌或所述至少一种m1相关标志物的表达的半数最大有效浓度(ec
50
)可以在剂量
‑
反应曲线中测定,如实施例部分所详述。在一些具体实施方案中,本发明的抗btn2a抗体表现出:
[0108]
‑
m1相关标志物表达(典型地为cd86和/或pdl1)的ec
50
为0.01μg/ml,特别是0.1μg/ml
‑
100μg/ml,特别是50μg/ml,最特别是1
‑
50μg/ml,和/或
[0109]
‑
m1相关细胞因子分泌(典型地为tnfα)的ec
50
为0.01μg/ml,特别是0.05μg/ml
‑
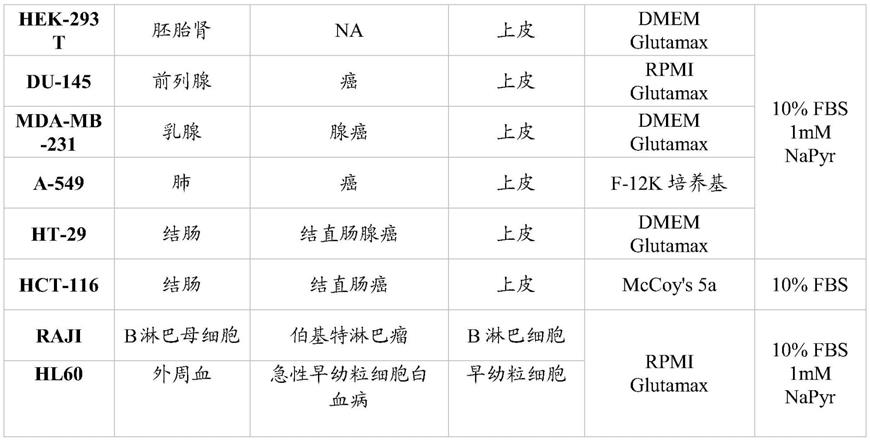
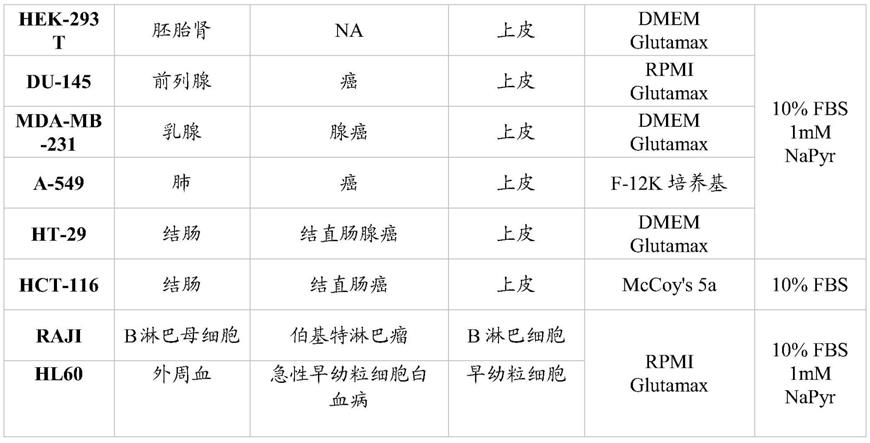
100μg/ml,特别是50μg/ml,最特别是0.1
‑
10μg/ml。
[0110]
在m2巨噬细胞逆转测定中,典型地在m
‑
csf存在下从单核细胞产生的m2巨噬细胞可以在抗btn2a抗体(最特别是之前定义的抗btn2a1抗体)存在下或与阴性对照(如其对照同种型)一起与或不与脂多糖(lps)一起培养。可以将gm
‑
csf(粒细胞
‑
巨噬细胞集落刺激因子)和ifnγ添加到m2培养物中作为m2逆转的阳性对照。在gm
‑
csf存在下极化的m1巨噬细胞典型地可用作表型对照。在逆转实验之后,在不存在lps的情况下逆转的巨噬细胞可以通过流式细胞术如上所述分析m1或m2相关标志物的表达。细胞因子分泌也可以如上所述定量。
[0111]
典型地,当:
[0112]
‑
与其对照同种型相比,当在所述抗btn2a抗体存在下培养m2巨噬细胞时,观察到m1标志物的显著增加;和/或
[0113]
‑
与其阴性对照(如其对照同种型)相比,当在所述抗btn2a抗体存在下培养m2巨噬细胞时,观察到m2标志物的显著减少时,
[0114]
本公开的抗btn2a抗体诱导m2巨噬细胞向m1巨噬细胞的逆转。
[0115]
可以通过用本文定义的抗btn2a抗体或用于阴性对照的对照同种型,添加或不添加il
‑
2和/或il
‑
15培养nk细胞(典型地来自健康供体)来评估自然杀伤(nk)细胞的活化。至少48小时后,尤其是至少4天后,然后可以针对其活化标记物如cd69和/或cd25对nk细胞进行胞外表型分析。典型地,与阴性对照(例如其同种型对照)相比,当在所述抗btn2a存在下(用或不用il
‑
2和/或il
‑
15进一步活化)观察到nk细胞活化标志物cd69和/或cd25的显著增加时,nk细胞活化被认为是由本公开的抗btn2a抗体诱导。本文提供的结果清楚证实本发明
的抗体触发nk细胞的直接活化。
[0116]
nk细胞的细胞毒性的增强可以通过评估nk细胞脱粒在体外进一步评估。在这种功能测定中,将癌细胞系(如白血病细胞系(骨髓性白血病)或癌(例如结肠癌、乳腺癌或肺腺癌)细胞系)在上述nk细胞(如上文所述预先用抗btn2a抗体或其阴性对照活化)存在下并且存在或不存在il
‑
2和/或il
‑
15的情况下共同培养。nk细胞脱粒典型地可以通过流式细胞术评估为cd107阳性nk细胞的百分比(存在或不存在il
‑
2和/或il
‑
15)。
[0117]
典型地,当用所述抗btn2a抗体活化nk细胞后,nk细胞脱粒(例如cd107阳性nk细胞的百分比)与其阴性对照(如其对照同种型)相比显著增加时,nk细胞的细胞毒性被认为是由抗btn2a抗体诱导。典型地,本公开的抗btn2a抗体剂量依赖性地诱导nk细胞脱粒(特别是针对如前所述的各种癌细胞系)。此类抗体在nk细胞脱粒方面的半数有效浓度(ec
50
)可以在如实施例部分详述的剂量
‑
反应曲线中测定。
[0118]
参考抗101g5和/或107g3可用作上述功能测定中的阳性对照。
[0119]
在一些实施方案中,如下文详述,本公开的抗btn2a抗体抑制单核细胞向m2巨噬细胞的极化,诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,触发直接nk细胞活化,和/或将nk细胞介导的细胞毒性增强至基本上等于或优于参考抗体mab 101g5或mab 107g3的水平。“抑制单核细胞向m2巨噬细胞的极化,诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,触发直接nk细胞活化,和/或将nk细胞介导的细胞毒性增强至基本上等于或优于参考抗体的水平”在本文中意指与参考抗体mab 107g3或101g5中的任一种相比,观察到所测试的抗btn2a1抗体的小于20%,特别是小于15%,特别是小于10%,典型地小于5%的测试功能活性的变化。
[0120]
本公开还涵盖以下抗体:
[0121]
1)对之前定义的btn2a,特别是btn2a1具有特异性,特别是具有至少一种以下特性的抗体:
[0122]
‑
它结合人btn2a1,典型地在细胞系,例如用编码人btn2a1的质粒转染的hek293t细胞系中表达,如实施例中所述,更具体地以低于50μg/ml,更具体地低于40μg/ml的ec
50
或它以10nm或更小的k
d
结合btn2a1和/或它在结合btn2a1上对比非特异性结合具有约10:1、约20:1、约50:1、约100:1、10000:1或更高的亲和力之比;
[0123]
‑
不与btn3交叉反应;
[0124]
和,
[0125]
2)进一步或替代地,其表现出一种或多种功能特性:
[0126]
–
其通过vγ9/vδ2t细胞激活溶细胞分子(特别是ifn
‑
γ)的产生,和/或
[0127]
‑
其激活vγ9/vδ2t细胞的溶细胞功能,和/或
[0128]
‑
其激活vγ9/vδ2t细胞的增殖。
[0129]
值得注意的是,vγ9/vδ2t细胞的增殖、溶细胞功能和溶细胞分子(特别是ifn
‑
γ)的产生通过活化的vγ9/vδ2t细胞实现,这典型地通过本文公开的抗体实现。
[0130]
可以使用实施例中描述的细胞测定法,特别是通过基于elisa评估vγ9/vδ2t细胞分泌的ifnγ,和/或对各种癌细胞系(如daudi细胞系、jurkat细胞系、l
‑
ipc细胞系或mda
‑
mb
‑
134细胞系)的cd107脱粒测定,从抗btn2a1抗体中筛选出具有这种有利特性的本公开的抗btn2a1抗体。
[0131]
根据本公开的溶细胞分子典型地在于ifnγ或tnfα细胞因子。
[0132]
如本文所用,“活化溶细胞分子(典型地在ifnγ和/或tnfα中)的产生”是指当与对照活化的vγ9/vδ2t细胞(以igg1或杂交瘤培养基为对照)相比时,观察到由活化的vγ9/vδ2t细胞产生至少ifnγ或tnfα的显著增加,所述vγ9/vδ2t细胞通过与靶细胞系(daudi、jurkat、l
‑
ipc或mda
‑
mb
‑
134细胞系)共培养或通过磷酸激动剂(pag)而被活化。典型地,由活化的vγ9/vδ2t细胞产生ifnγ或tnfα的激活可在细胞测定中通过用流式细胞术评估的抗ifnγ或tnfα的抗体进行细胞内标记来测量,或通过基于elisa剂量的vγ9/vδ2t细胞在其培养基中分泌的ifnγ或tnfα来测量。在下文实施例(参见材料和方法部分)中更详细地描述了这种测定。
[0133]
如本文所用,“激活活化的vγ9/vδ2t细胞的溶细胞功能”是指当与对照活化的人vγ9/vδ2t细胞(以igg1或杂交瘤培养基为对照)相比时,观察到活化的人vγ9/vδ2t细胞的溶细胞功能的显著增加,所述人vγ9/vδ2t细胞通过与靶细胞系(例如daudi、jurkat、l
‑
ipc或mda
‑
mb
‑
134细胞系)共培养或通过磷酸激动剂(pag)而被活化。典型地,活化的vγ9/vδ2t细胞的溶细胞功能的激活可根据针对标准细胞系诱导vγ9/vδ2t细胞脱粒的激活的测量来测定,共同使用cd107a和cd107b作为用于检测阳性脱粒vγ9/vδ2t细胞的脱粒标志物。12
‑
豆蔻酸
‑
13
‑
乙酸佛波醇(pma)与离子霉素处理vγ9/vδ2t细胞典型地可用作vγ9/vδ2t细胞活化的阳性对照。在下文实施例中更详细地描述了这种测定。
[0134]
如本文所用,“激活活化的vγ9/vδ2t细胞的增殖”是指当与用igg1作为对照活化的vγ9/vδ2t细胞的增殖相比时,观察到活化的vγ9/vδ2t细胞的增殖的显著增加,所述vγ9/vδ2t细胞通过与靶细胞系(例如daudi、jurkat、l
‑
ipc或mda
‑
mb
‑
134细胞系)共培养或通过磷酸激动剂(pag)而被活化。典型地,活化的vγ9/vδ2t细胞的增殖可在细胞测定中通过cfse或cell trace紫染色和从外周血和流式细胞术中提取纯化的vγ9/vδ2t细胞的流式细胞仪来测量,或在有或没有刺激的情况下通过监测外周血单核细胞内vγ9/vδ2t细胞区室的扩增来测量。
[0135]
在一些实施方案中,如下文所述,本公开的抗btn2a1抗体将活化的vγ9/vδ2t细胞的溶细胞功能激活至基本上等于或优于参考抗体mab 107g3的水平。“将活化的vγ9/vδ2t细胞的溶细胞功能激活至基本上等于参考抗体的水平”在本文中意指与参考mab107g3相比,观察到所测试的抗btn2a1抗体的活化的vγ9/vδ2t细胞的溶细胞功能的变化小于15%,特别是小于10%,典型地小于5%。
[0136]
在一些实施方案中,如下文所述,本公开的抗btn2a1抗体将活化的vγ9/vδ2t细胞的溶细胞分子(即:至少ifnγ或tnfα)的产生激活至基本上等于或优于参考抗体mab 107g3的水平。“将通过vγ9/vδ2t细胞活化的至少ifnγ或tnfα激活至基本上等于参考抗体的水平”在本文中意指与参考mab 107g3相比,观察到所测试的抗btn2a1抗体的活化的vγ9/vδ2t细胞的溶细胞分子产生的变化小于15%,特别是小于10%,典型地小于5%。
[0137]
在一些实施方案中,如下文所述,本公开的抗btn2a1抗体将活化的vγ9/vδ2t细胞的溶细胞功能激活至基本上等于或优于参考抗体mab 107g3的水平。“将活化的vγ9/vδ2t细胞的溶细胞功能激活至基本上等于参考抗体的水平”在本文中意指与参考mab107g3相比,观察到所测试的抗btn2a1抗体的活化的vγ9/vδ2t细胞的溶细胞功能的变化小于15%,特别是小于10%,典型地小于5%。
[0138]
参考抗体mab 101g5、107g3及其变体
[0139]
本文公开的抗体包括参考单克隆抗体mab 101g5,其包含分别如seq id no:19和20中定义的相应vh和vl区,以及mab 107g3,其包含分别如seq id no:1和2中定义的相应vh和vl区。
[0140]
本公开的其他抗体包括与分别如seq id no:1和2中定义或分别如seq id no:19和20中定义的vh和vl区具有至少90%,特别是至少95、96、97、98、99或100%同一性的那些。
[0141]
在特定实施方案中,根据本公开的抗btn2a抗体,典型地是人源化抗btn2a1,含有包含seq id no:3的重链可变区cdr1、包含seq id no:4的重链可变区cdr2、包含seq id no:5的重链可变区cdr3、包含seq id no:6的轻链可变区cdr1、包含seq id no:7的轻链可变区cdr2和包含seq id no:8的轻链可变区cdr3。在本文公开的抗体的具体实施方案中,6个cdr区与seq id no:3
‑
8中定义的参考mab 107g3的6个cdr区100%相同。
[0142]
在其他具体实施方案中,根据本公开的抗btn2a1抗体,典型地是人源化抗btn2a1,含有包含seq id no:21的重链可变区cdr1、包含seq id no:22的重链可变区cdr2、包含seq id no:23的重链可变区cdr3、包含seq id no:24的轻链可变区cdr1、包含seq id no:25的轻链可变区cdr2和包含seq id no:26的轻链可变区cdr3。
[0143]
在本文公开的抗体的具体实施方案中,6个cdr区与seq id no:21
‑
26中定义的参考mab 101g5的6个cdr区100%相同。本文公开的其他抗体包括具有通过氨基酸缺失、插入或取代而突变的氨基酸,但在cdr区与参考mab 101g5的cdr区具有至少60、70、80、90、95、96、97、98、99或100%同一性的那些抗体。典型地,根据本公开,与seq id no:21
‑
26中定义的参考抗体mab 107g3的cdr序列相比,抗体可以在一个或多个cdr中具有1、2、3或4个氨基酸变异(包括缺失、插入或取代)。
[0144]
在本文公开的抗体的其他具体实施方案中,6个cdr区与seq id no:3
‑
8中定义的参考mab 107g3的6个cdr区100%相同。本文公开的其他抗体包括具有通过氨基酸缺失、插入或取代而突变的氨基酸,但在cdr区与参考mab 107g3的cdr区具有至少60、70、80、90、95、96、97、98、99或100%同一性的那些抗体。典型地,根据本公开,与seq id no:3
‑
8中定义的参考抗体mab 107g3的cdr序列相比,抗体可以在一个或多个cdr中具有1、2、3或4个氨基酸变异(包括缺失、插入或取代)。
[0145]
在一些实施方案中,本公开的抗体是mab 101g5或mab 107g3的突变变体,其6个cdr区分别与参考mab 101g5或107g3的相应6个cdr区100%相同,并且其中与相应的参考抗体的相应框架区相比,所述突变变体抗体包括其中不超过1、2、3、4或5个氨基酸通过在fr1、fr2、fr3和fr4区中的氨基酸缺失、插入或取代而突变的突变氨基酸序列。
[0146]
功能性变体抗体
[0147]
如实施例中提供的实验数据所示,参考mab 107g3结合人btn2a1的第65、68、69、72、78、84、85、95、97、100位上的残基。因此,本公开涵盖结合包含位于seq id n
°
17的第60
‑
100位氨基酸残基的构象表位,并且最特别地包含位于seq id n
°
17的第65、68、69、72、78、84、85、95、97、100位上的氨基酸残基的构象表位的mab,并且其具有一种或多种如先前定义的和如下文进一步提醒的功能特性,特别是具有参考mab 107g3的一种或多种功能特性。
[0148]
还如实施例中提供的实验数据所示,参考mab 101g5结合btn2a1的第212、213、218、220、224、229位上的残基。因此,本公开涵盖结合包含位于seq id n
°
17的第210
‑
230位氨基酸残基的构象表位,并且最特别地包含位于seq id n
°
17的第212、213、218、220、224、
229位上的残基的构象表位的mab,并且其具有一种或多种如先前定义的和如下文进一步提醒的功能特性,特别是具有参考mab 101g5的一种或多种功能特性。
[0149]
在其他实施方案中,本公开的功能性变体抗体具有全长重链和轻链氨基酸序列;或可变区重链和轻链氨基酸序列,或与上述参考抗体mab 101g5或mab 107g3中任一个的相应氨基酸序列同源或更具体地相同的所有6个cdr区氨基酸序列,并且其中此类功能性变体抗体保留所述参考抗体的所需功能特性。
[0150]
参考mab 101g5抗体或参考抗体mab 107g3的功能性变体,特别是在本公开单克隆抗体的上下文中所用的vl、vh或cdr的功能性变体仍然允许抗体保留亲本抗体(例如mab 101g5或mab 107g3)的至少相当大比例(至少约50%、60%、70%、80%、90%、95%或100%)的亲和力(典型地通过表面等离子体共振(spr)测量的k
d
评估,或使用基于生物层干涉测量(bli)技术的平台,典型地在25℃下进行)和/或亲本抗体(例如mab 101g5或mab 107g3)的选择性,并且在某些情况下,本公开的这种单克隆抗体可能涉及比亲本ab(例如mab 101g5或mab 107g3)更高的亲和力、选择性和/或特异性。
[0151]
如本文公开的参考mab 101g5或107g3或所述参考抗体的变体的所需功能特性可以选自下组:
[0152]
i.对btn2a1的特异性,特别是与在细胞系中表达的人btn2a1结合的特性,例如用编码人btn2a1的质粒转染的hek
‑
293t btn2ko细胞,如实施例中所述,更具体地以低于50μg/ml,更具体地低于40μg/ml的ec
50
,或以如通过表面等离子体共振(spr)(通常在25℃)或luminex测定(如实施例中所示)或(abdiche et al.2008)测量的10nm或更小的k
d
和/或它结合btn2a1相比非特异性结合具有约10:1、约20:1、约50:1、约100:1、10000:1或更高的亲和力之比;和/或
[0153]
ii.抑制单核细胞向m2巨噬细胞的极化,典型地如实施例部分所示评估,和/或
[0154]
iii.诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,典型地如实施例部分所示评估,和/或
[0155]
iv.直接触发nk细胞活化,典型地如实施例部分所示评估,和/或
[0156]
v.增强nk细胞介导的细胞毒性,典型地如实施例部分所示评估。
[0157]
在一些更具体实施方案中,本文公开的参考mab 107g3或所述参考抗体的变体的所需功能特性还可选自下组:
[0158]
vi.激活vγ9/vδ2t细胞产生的溶细胞分子(例如:ifnγ或tnfα),典型地如实施例所示评估,
[0159]
vii.激活vγ9/vδ2t细胞的溶细胞功能,典型地如实施例所示评估;和/或
[0160]
viii.激活vγ9/vδ2t细胞增殖,典型地如实施例所示评估。
[0161]
典型地,参考mab 101g5或107g3的功能性变体的根据以上(ii)至(v)点的功能特性基本上等于或优于如上所述的相应参考抗体mab101g5或107g3的相应功能特性。基本上等于在本文中意指所述功能性变体保留参考mab 107g3的相应功能特性的至少约50%、60%、70%、80%、90%、95%、96%、97%、98%、99%或100%。
[0162]
典型地,参考mab 107g3的功能性变体的根据以上(vi)至(viii)点的功能特性基本上等于或优于如上所述的相应参考抗体mab107g3的相应功能特性。基本上等于在本文中意指所述功能性变体保留参考mab 107g3的相应功能特性的至少约50%、60%、70%、80%、
90%、95%、96%、97%、98%、99%或100%。
[0163]
例如,本公开涉及参考mab 101g5的功能性变体抗体,其包含可变重链(v
h
)和可变轻链(v
l
)序列,其中cdr序列,即6个cdr区;hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3与seq id no:21
‑
26中所定义的mab 101g5参考抗体的相应cdr序列共享至少60、70、90、95或100序列同一性百分比,其中所述功能性变体抗体特异性结合btn2a,并且所述抗体表现出至少一种以下功能特性i)
‑
iv):
[0164]
i.其抑制单核细胞向m2巨噬细胞极化,
[0165]
ii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
[0166]
iii.其直接触发nk细胞激活,
[0167]
iv.其增强nk细胞介导的细胞毒性。
[0168]
优选地,它表现出特性i)和ii)中的至少一种以及特性iii和iv)中的至少一种,最优选地表现出特性i)
‑
iv)。
[0169]
本公开还涉及参考mab 107g3的功能性变体抗体,其包含可变重链(v
h
)和可变轻链(v
l
)序列,其中cdr序列,即6个cdr区;hcdr1、hcdr2、hcdr3、lcdr1、lcdr2、lcdr3与seq id no:3
‑
8中所定义的mab 107g3参考抗体的相应cdr序列共享至少60、70、90、95或100序列同一性百分比,其中所述功能性变体抗体特异性结合btn2a,并且所述抗体表现出至少一种以下功能特性i)
‑
iv):
[0170]
i.其抑制单核细胞向m2巨噬细胞极化,
[0171]
ii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
[0172]
iii.其直接触发nk细胞激活,
[0173]
iv.其增强nk细胞介导的细胞毒性,
[0174]
v.其激活vγ9/vδ2t细胞产生ifnγ或tnfα,
[0175]
vi.其激活vγ9/vδ2t细胞的溶细胞功能,
[0176]
vii.其激活vγ9/vδ2t细胞的增殖。
[0177]
优选地,所述功能性变体表现出功能活性i)至iv)中的至少一种(优选至少活性i和/或ii,和iii和/或iv)和功能活性v)至vii)中的至少一种。
[0178]
它还涉及mab 101g5参考抗体的功能性变体抗体,其包含与分别在seq id no:19和20中定义的所述mab 101g5参考抗体的相应重链和轻链可变区至少80%、90%或至少95、96%、97%、98%、99%或100%相同的重链可变区和轻链可变区;功能性变异抗体特异性结合btn2,并表现出至少一种以下功能特性:
[0179]
i.其抑制单核细胞向m2巨噬细胞极化,
[0180]
ii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
[0181]
iii.其直接触发nk细胞激活,
[0182]
iv.其增强nk细胞介导的细胞毒性。
[0183]
优选地,它表现出特性i)和ii)中的至少一种以及特性iii和iv)中的至少一种,最优选地表现出特性i)
‑
iv)。
[0184]
它还涉及mab 107g3参考抗体的功能性变体抗体,其包含与分别在seq id no:1和2中定义的所述mab 107g3参考抗体的相应重链和轻链可变区至少80%、90%或至少95、96%、97%、98%、99%或100%相同的重链可变区和轻链可变区;功能性变异抗体特异性结
合btn2,并表现出至少一种以下功能特性:
[0185]
i.其抑制单核细胞向m2巨噬细胞极化,
[0186]
ii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转,
[0187]
iii.其直接触发nk细胞激活,
[0188]
iv.其增强nk细胞介导的细胞毒性,
[0189]
v.其激活通过vγ9/vδ2t细胞产生溶细胞分子(ifnγ或tnfα),vi.其激活vγ9/vδ2t细胞的溶细胞功能,和/或
[0190]
vii.其激活vγ9/vδ2t细胞的增殖。
[0191]
在一些实施方案中,所述功能变体表现出功能活性i)至iv)中的至少一种(特别是至少活性i和/或ii,和iii和/或iv)和功能活性v)至vii)中的至少一种。
[0192]
在一些实施方案中,所述功能变体表现出一种或多种功能活性i至iv或一种或多种功能活性v至vii。
[0193]
典型地,参考mab 101g5或107g3的功能性变体的根据以上(i)至(iv)点的功能特性基本上等于或优于如上所述的相应参考抗体mab 101g5或107g3的相应功能特性。基本上等于在本文中意指所述功能性变体保留参考mab 107g3的相应功能特性的至少约50%、60%、70%、80%、90%、95%、96%、97%、98%、99%或100%。
[0194]
在各种实施方案中,抗体可表现出上文讨论的一种或多种所需功能特性。
[0195]
例如,所述抗体可以是人抗体、人源化抗体或嵌合抗体。典型地,抗体或蛋白质是人源化抗体,更特别地人源化沉默抗体。
[0196]
如本文所用,术语“沉默”抗体是指在测量靶细胞的细胞裂解的体外adcc活性测定中所测量的无或低adcc活性的抗体。
[0197]
在一个实施方案中,术语“无或低adcc活性”是指沉默抗体表现出的adcc活性低于相应野生型(非沉默)抗体(例如野生型人igg1抗体)所观察到的adcc活性的50%,例如低于10%。典型地,与对照fab抗体相比,在沉默抗体的体外adcc活性测定中未观察到可检测的adcc活性。
[0198]
沉默的效应子功能可以通过抗体的fc恒定部分的突变获得,并且已经描述于本领域中:strohl 2009(lala&n297a);baudino 2008,d265a(baudino et al.,j.immunol.2008,strohl,co biotechnology 20 2009)。沉默的igg1抗体的实例包含在igg1 fc氨基酸序列(eu编号)的第234、235和/或331位降低adcc的突变。另一种沉默的igg1抗体包含n297a突变,其导致糖基化或非糖基化抗体。
[0199]
cdr变体的序列可能通过大多数保守性替换与亲本抗体序列的cdr序列不同,例如变体中至少10个,诸如至少9个、8个、7个、6个、5个、4个、3个、2个或1个替换是保守性氨基酸残基替代。在本公开上下文中,保守性替换可以定义为如下所反映的氨基酸类别内的替换:
[0200]
脂族残基i、l、v和m。
[0201]
环烯基相关残基f、h、w和y。
[0202]
疏水性残基a、c、f、g、h、i、l、m、r、t、v、w和y。
[0203]
带负电荷的残基d和e。
[0204]
极性残基c、d、e、h、k、n、q、r、s和t。
[0205]
带正电荷的残基h、k和r。
[0206]
小残基a、c、d、g、n、p、s、t和v。
[0207]
非常小的残基a、g和s。
[0208]
涉及转角的残基a、c、d、e、g、h、k、n,q、r、s、p和涉及形成的残基t。
[0209]
柔性残基q、t、k、s、g、p、d、e和r。
[0210]
更多的保守性替换分组包括:缬氨酸
‑
亮氨酸
‑
异亮氨酸、苯丙氨酸
‑
酪氨酸、赖氨酸
‑
精氨酸、丙氨酸
‑
缬氨酸和天冬酰胺
‑
谷氨酰胺。与任何一个mab1
‑
6的cdr相比,在变体cdr中基本上保留了亲水(hydropathic)/亲水特性和残基重量/大小方面的保守性。本领域通常理解亲水氨基酸指数在赋予蛋白质相互作用的生物学功能方面的重要性。已经接受的是,氨基酸的相对亲水特性有助于所得蛋白质的二级结构,这反而限定了蛋白质与其他分子(例如酶、底物、受体、dna、抗体、抗原等)的相互作用。根据它们的疏水性和电荷特征,为每种氨基酸指定了亲水指数,它们是:异亮氨酸( 4.5);缬氨酸( 4.2);亮氨酸( 3.8);苯丙氨酸( 2.8);半胱氨酸/胱氨酸( 2.5);蛋氨酸( 1.9);丙氨酸( 1.8);甘氨酸(
‑
0.4);苏氨酸(
‑
0.7);丝氨酸(
‑
0.8);色氨酸(
‑
0.9);酪氨酸(
‑
1.3);脯氨酸(
‑
1.6);组氨酸(
‑
3.2);谷氨酸(
‑
3.5);谷氨酰胺(
‑
3.5);天冬氨酸(
‑
3.5);天冬酰胺(
‑
3.5);赖氨酸(
‑
3.9)和精氨酸(
‑
4.5)。类似残基的保留也可以或替代地通过相似性得分来测量,如使用blast程序来测定(例如,可通过ncbi获得的blast 2.2.8,其使用标准设置blosum62,open gap=11和extended gap=1)。合适的变体典型地表现出与亲本肽至少约80%的同一性。根据本公开,第一氨基酸序列与第二氨基酸序列具有至少70%同一性是指第一序列与第二氨基酸序列具有70;71;72;73;74;75;76;77;78;79;80;81;82;83;84;85;86;87;88;89;90;91;92;93;94;95;96;97;98;99或100%同一性。根据本公开,第一氨基酸序列与第二氨基酸序列具有至少50%同一性是指第一序列与第二氨基酸序列具有50;51;52;53;54;55;56;57;58;59;60;61;62;63;64;65;66;67;68;69;70;71;72;73;74;75;76;77;78;79;80;81;82;83;84;85;86;87;88;89;90;91;92;93;94;95;96;97;98;99或100%同一性。
[0211]
在一些实施方案中,本公开的抗体是嵌合抗体,典型地是嵌合的小鼠/人抗体。术语“嵌合抗体”是指单克隆抗体,其包含衍生自非人动物的抗体的vh结构域和vl结构域和人抗体的ch结构域和cl结构域。作为非人动物,可以使用任何动物,诸如小鼠、大鼠、仓鼠、兔等。特别地,所述小鼠/人嵌合抗体可包含任一种mab 101g5或mab 107g3参考抗体的vh和vl结构域。
[0212]
在一些实施方案中,本公开的抗体是人源化抗体。在具体实施方案中,本公开的抗体是包含任一种mab 101g5或mab 107g3参考抗体的6个cdr的人源化抗体。如本文所用,术语“人源化抗体”是指其中框架区(fr)已经被修饰以包含来自与亲本免疫球蛋白(例如鼠cdrs)相比具有不同物种(例如人种)的供体免疫球蛋白的fr的抗体。
[0213]
在一些实施方案中,本公开的抗体选自下组:fab、f(ab')2、fab'和scfv。如本文所用,术语“fab”表示具有约50,000的分子量和抗原结合活性的抗体片段,其中在用蛋白酶(木瓜蛋白酶)处理igg获得的片段中,约一半h链的n末端侧和整个l链通过二硫键结合在一起。术语“f(ab')2”是指具有约100,000的分子量和抗原结合活性的抗体片段,在用蛋白酶(胃蛋白酶)处理igg获得的片段中,其比通过铰链区的二硫键结合的fab略大。术语“fab'”是指具有约50,000的分子量和抗原结合活性的抗体片段,其通过切割f(ab')2的铰链区的二硫键获得。单链fv(“scfv”)多肽是共价连接的vh:vl异二聚体,其通常由包括通过肽编码
接头连接的vh和vl编码基因的基因融合体表达。本公开的人scfv片段包括通常通过使用基因重组技术保持适当构象的cdr。
[0214]
具有突变体氨基酸序列的功能性变体抗体可通过编码核酸分子的诱变(例如,定点诱变或pcr介导的诱变),然后使用本文所述的功能测定法测试编码的改变抗体的保留功能(即上述功能)来获得。
[0215]
与参考101g5或107g3 mab交叉竞争的抗体
[0216]
可以基于它们以统计学显著的方式与上述所述参考mab 107g3交叉竞争(例如,竞争性抑制结合)的能力,在标准btn2a1结合测定中鉴定具有与本文公开的参考mab 101g5或参考mab 107g3的类似有利特性的其他抗体。
[0217]
例如,首先可以从人重组抗体文库中使用例如噬菌体展示技术或从表达用btn2a1抗原免疫的人可变区抗体的转基因小鼠中筛选测试抗体与btn2a1的结合亲和力,如典型地在实施例中评估(参考材料和方法部分)。
[0218]
在另一个实施方案中,本公开提供与至少上述参考mab 107g3或参考mab 101g5结合相同表位的抗体。如实施例部分所示,参考mab 101g5和107g3不结合btn2a1上的相同表位。
[0219]
测试抗体交叉竞争或抑制本公开的抗体与人btn2a1结合的能力证实了所述测试抗体可以与该抗体竞争与人btn2a1的结合。根据非限制性理论,该抗体可以如与其竞争的抗体结合人btn2a1上相同或相关的(例如,结构上相似或空间上接近)表位。
[0220]
例如,以下测试可用于筛选抗btn2a1抗体与mab 107g3参考抗体交叉竞争的能力和/或筛选抗btn2a1抗体与所述参考抗体结合相同表位的能力抗体:用人btn2a1(典型地为实施例中描述的hek293t)转染的btn2ko细胞可以用饱和浓度(例如10μg/ml)的参考抗体mab 107g3染色。然后,可以测试不同剂量的测试抗btn2a1 mab与mab 107g3参考抗体的竞争潜力。在此类参考抗体存在的情况下,确实与参考抗体竞争的mab将不能识别btn2a1。数据可以表示为平均荧光强度。或者,竞争测定可以在如实施例部分所述的分箱测定(binning assay)中进行。典型地,分箱实验可以通过将重组人btn2a1固定在生物传感器上并提供参考抗体,然后提供竞争抗体来进行。
[0221]
可以进一步针对mab 101g5或mab 107g3的有利特性测试所选抗体,特别是如先前详述的。
[0222]
因此,在一个实施方案中,本公开提供分离的抗体,其竞争结合参考mab 101g5或参考mab 107g3而结合btn2a1,其中
[0223]
所述抗体:
[0224]
i.具有对btn2a1的特异性,特别是与在细胞系中表达的人btn2a1结合的特性,例如用编码人btn2a1的质粒转染的hek
‑
293t btn2 ko细胞,如实施例中所述,更具体地以低于50μg/ml,更具体地低于40μg/ml的ec
50
,或如通过表面等离子体共振(spr)或luminex测定(如实施例中所示)或(abdiche et al.2008)测量的10nm或更小的k
d
和/或它结合btn2a1相比非特异性结合具有约10:1、约20:1、约50:1、约100:1、10000:1或更高的亲和力之比(更多细节还参见上文);和/或
[0225]
ii.其抑制单核细胞向m2巨噬细胞的极化
[0226]
iii.其诱导m2巨噬细胞向抗肿瘤m1巨噬细胞的逆转
[0227]
iv.其直接触发nk细胞活化,和/或
[0228]
v.其增强nk细胞介导的细胞毒性。
[0229]
或者或进一步地,所述抗体:
[0230]
vi.激活通过vγ9/vδ2t细胞产生溶细胞分子(例如:ifnγ或tnfα),典型地如实施例所示评估,和/或
[0231]
vii.激活vγ9/vδ2t细胞的溶细胞功能,典型地如实施例所示评估;和/或
[0232]
viii.激活vγ9/vδ2t细胞增殖,典型地如实施例所示评估。
[0233]
更具体地,在此类实施方案中,所述抗体不与btn3交叉反应。
[0234]
典型地,与参考mab 101g5或107g3竞争结合btn2a1的抗体的根据以上(iii)至(v)点的功能特性基本上等于或优于如上所述的相应参考抗体mab 101g5或107g3的相应功能特性。基本上等于在本文中意指所述功能性变体保留参考mab 107g3的相应功能特性的至少约50%、60%、70%、80%、90%、95%、96%、97%、98%、99%或100%。
[0235]
典型地,与根据本公开的参考mab 107g3竞争结合btn2a1的抗体仍保留参考抗体(例如:mab 107g3)的至少相当大比例(至少约50%、60%、70%、80%、90%、95%或100%)的亲和力和/或选择性,并且在某些情况下可能与比参考抗体(例如:mab 107g3)更大的亲和力、选择性和/或特异性相关。
[0236]
在某个实施方案中,交叉阻断抗体或与参考mab 101g5或107g3竞争结合btn2a1的抗体是嵌合抗体、人源化抗体或人重组抗体。
[0237]
产生单克隆抗体的转染瘤的产生
[0238]
通过本领域已知的任何技术产生本公开的抗体,例如但不限于任何单独或组合的化学、生物学、遗传学或酶学技术。典型地,已知所需序列的氨基酸序列,本领域技术人员可以通过用于生产多肽的标准技术容易地生产所述抗体。例如,它们可以使用熟知的固相方法合成,典型地使用市售的肽合成装置(诸如由applied biosystems,foster city,california制造的)并遵循制造商的说明书。或者,可以通过本领域熟知的重组dna技术合成本公开的抗体。例如,在将编码抗体的dna序列整合入表达载体并将这些载体引入将表达所需抗体的合适的真核或原核宿主中后,可以获得作为dna表达产物的抗体,之后可以使用已知技术分离它们。
[0239]
因此,本公开的另一个目的涉及编码根据本公开的抗体的核酸分子。更特别地,核酸分子编码本文公开的抗体的重链或轻链。更特别地,核酸分子包含与编码任何一种参考抗体mab 107g3的重链可变区(vh区)或轻链可变区(vl)的相应核酸具有至少70%、80%、90%、95%或100%同一性的vh或vl编码区。
[0240]
典型地,所述核酸是dna或rna分子,其可以包括在任何合适的载体中,诸如质粒、粘粒、附加体、人工染色体、噬菌体或病毒载体。如本文所用,术语“载体”、“克隆载体”和“表达载体”是指可以将dna或rna序列(例如外源基因)引入宿主细胞,以转化宿主并促进引入序列的表达(例如转录和翻译)的介质。因此,本公开的另一个目的涉及包含本文公开的核酸的载体。此类载体可包含调节元件,诸如启动子、增强子、终止子等,以在施用于受试者时引起或指导所述抗体的表达。用于动物细胞的表达载体的启动子和增强子的实例包括sv40的早期启动子和增强子、moloney小鼠白血病病毒的ltr启动子和增强子、免疫球蛋白h链的启动子和增强子等。可以使用任何用于动物细胞的表达载体,只要可以插入并表达编码人
抗体c区的基因即可。合适载体的实例包括page107、page103、phsg274、pkcr、psg1βd2
‑
4等。质粒的其他实例包括含有复制起点的复制质粒,或整合质粒,诸如puc、pcdna、pbr等。病毒载体的其他实例包括腺病毒、逆转录病毒、疱疹病毒和aav载体。可通过本领域已知的技术产生此类重组病毒,诸如通过转染包装细胞或通过用辅助质粒或病毒进行瞬时转染。病毒包装细胞的典型实例包括pa317细胞、psicrip细胞、gpenv 细胞、293细胞等。例如,用于产生这种复制缺陷型重组病毒的详细方案可以在wo 95/14785、wo 96/22378、us 5,882,877、us 6,013,516、us 4,861,719、us 5,278,056和wo 94/19478中找到。
[0241]
本公开的进一步的目的涉及通过上述核酸和/或载体转染、感染或转化的宿主细胞。如本文所用,术语“转化”是指将“外源”(即外部或细胞外)基因、dna或rna序列引入宿主细胞,从而宿主细胞将表达引入的基因或序列以产生所需的物质,典型地是由引入的基因或序列编码的蛋白质或酶。接受并表达引入的dna或rna的宿主细胞被“转化”。
[0242]
本文公开的核酸可用于在合适的表达系统中产生本公开的抗体。术语“表达系统”是指在合适条件下的宿主细胞和相容载体,例如,其用于表达由载体携带并引入宿主细胞的外源dna编码的蛋白质。常见的表达系统包括大肠杆菌宿主细胞和质粒载体、昆虫宿主细胞和杆状病毒载体,以及哺乳动物宿主细胞和载体。宿主细胞的其他实例包括但不限于原核细胞(诸如细菌)和真核细胞(诸如酵母细胞、哺乳动物细胞、昆虫细胞、植物细胞等)。具体实例包括大肠杆菌、克鲁维酵母或酵母菌、哺乳动物细胞系(例如hek
‑
293细胞、vero细胞、cho细胞、3t3细胞、cos细胞等)以及原代或已建立的哺乳动物细胞培养物(例如,由淋巴母细胞、成纤维细胞、胚胎细胞、上皮细胞、神经细胞、脂肪细胞等产生)。实例还包括小鼠sp2/0
‑
ag14细胞(atcc crl1581)、小鼠p3x63
‑
ag8.653细胞(atcc crl1580)、其中二氢叶酸还原酶基因(下文称为“dhfr基因”)有缺陷的cho细胞(urlaub g等;1980)、大鼠yb2/3hl.p2.g11.16ag.20细胞(atcc crl1662,以下称为“yb2/0细胞”)等。
[0243]
本公开还涉及用于产生表达本文公开的抗体的重组宿主细胞的方法,所述方法包括以下步骤:(i)将如上所述的重组核酸或载体在体外或离体导入感受态宿主细胞,(ii)体外或离体培养获得的重组宿主细胞,和(iii)任选地,选择表达和/或分泌所述抗体的细胞。这些重组宿主细胞可用于产生本公开的抗体。
[0244]
通过常规免疫球蛋白纯化方法,例如蛋白a
‑
sepharose、羟磷灰石层析、凝胶电泳、透析或亲和层析适当地从培养基分离本公开的抗体。
[0245]
在一些实施方案中,本公开的人嵌合抗体可以通过以下来产生:获得如前所述的编码vl和vh结构域的核酸序列,通过将它们插入具有编码人抗体ch和人抗体cl的基因的动物细胞的表达载体来构建人嵌合抗体表达载体,并通过将表达载体引入动物细胞来表达编码序列。作为人嵌合抗体的ch结构域,可以是属于人免疫球蛋白的任何区域,但igg类的那些区域是合适的,并且也可以使用属于igg类的任何一个亚类,诸如igg1、igg2、igg3和igg4。另外,作为人嵌合抗体的cl,可以是属于ig的任何区域,也可以使用κ类或λ类的区域。用于产生嵌合抗体的方法涉及常规重组dna,且基因转染技术是本领域熟知的(参见morrison sl等(1984)和专利文献us5,202,238;和us5,204,244)。
[0246]
本公开的人源化抗体可以通过以下产生:获得如前所述的编码cdr结构域的核酸序列,通过将它们插入具有编码以下的基因的表达载体来构建人源化抗体表达载体:(i)与人抗体相同的重链恒定区和重链可变框架区,和(ii)与人抗体相同的轻链恒定区轻链可变
框架区,并通过将表达载体引入合适的细胞系来表达基因。人源化抗体表达载体可以是这样的类型:其中编码抗体重链的基因和编码抗体轻链的基因存在于不同的载体上,或者两种基因存在于同一载体上(串联型)。考虑到构建人源化抗体表达载体的容易性,引入细胞系的容易性,以及细胞系中抗体h和l链的表达水平之间的平衡,优选串联型人源化抗体表达载体。串联型人源化抗体表达载体的实例包括pkantex93(wo 97/10354)、pee18等。
[0247]
基于常规重组dna和基因转染技术人源化抗体的方法是本领域熟知的(参见例如,riechmann l.等,1988;neuberger ms等,1985)。可以使用本领域已知的多种技术将抗体人源化,包括例如cdr
‑
移植(ep 239,400、pct公开wo91/09967、美国专利号5,225,539、5,530,101和5,585,089)、镶饰(veneering)或重塑(ep 592,106、ep 519,596、padlan ea(1991)、studnicka gm等(1994)、roguska ma等(1994))和链替换(美国专利号5,565,332)。用于制备此类抗体的一般重组dna技术也是已知的(参见欧洲专利申请ep 125023和国际专利申请wo 96/02576)。
[0248]
可以通过用蛋白酶(木瓜蛋白酶)处理与amh特异性反应的抗体而获得本公开的fab。此外,可以通过以下来产生fab:将编码抗体fab的dna插入用于原核表达系统或用于真核表达系统的载体,并将载体引入原核生物或真核生物(视情况而定)以表达fab。
[0249]
可以通过用蛋白酶(胃蛋白酶)处理与amh特异性反应的抗体而获得本公开的f(ab')2。此外,可以通过用硫醚键或二硫键结合下述fab'来产生f(ab')2。
[0250]
可以通过用还原剂(二硫苏糖醇)处理与amh特异性反应的f(ab')2而获得本公开的fab'。此外,可以通过以下来产生fab':将编码抗体的fab'片段的dna插入原核生物的表达载体或真核生物的表达载体中,并将载体引入原核生物或真核生物(视情况而定)以进行表达。
[0251]
可以通过以下来产生本公开的scfv:获得如前所述编码vh和vl结构域的cdna,构建编码scfv的dna,将dna插入原核生物的表达载体或真核生物的表达载体,然后将表达载体引入原核生物或真核生物(视情况而定)以表达scfv。
[0252]
为制备人源化scfv片段,可以使用称为cdr移植的公知技术,其涉及从供体scfv片段选择互补决定区(cdr),并将它们移植到已知三维结构的人scfv片段框架上(例如,参见wo98/45322、wo87/02671、us5,859,205、us5,585,089、us4,816,567、ep0173494)。
[0253]
本公开的工程化抗体进一步包括其中对vh和/或vl内的框架残基进行修饰,例如,以改善抗体特性的抗体。典型地,进行该框架修饰以降低抗体的免疫原性。例如,一种途径是将一个或多个框架残基“回复突变”到相应的种系序列。更具体地,已经历体细胞突变的抗体可以含有与衍生抗体的种系序列不同的框架残基。可以通过将抗体框架序列与衍生抗体的种系序列进行比较来鉴定这些残基。为了使框架区序列恢复其种系构型,例如可以通过定点诱变或pcr介导的诱变将体细胞突变“回复突变”至种系序列。这种“回复突变”抗体也涵盖在本公开中。另一种类型的框架修饰涉及突变框架区内或甚至一个或多个cdr区内的一个或多个残基,以去除t细胞表位,从而降低抗体的潜在免疫原性。该途径也称为“去免疫化”,并且进一步详细描述于carr等的美国专利公开号20030153043中。
[0254]
fc工程化
[0255]
本公开的抗体可以通过上述方面的一个或多个功能或结构特征,或通过选择的功能和结构特征的任何组合来表征。
[0256]
本公开的抗体可以是任何同种型。同种型的选择典型地由所需的效应子功能(诸如adcc沉默)来引导。示例性同种型是igg1、igg2、igg3和igg4。可以使用人轻链恒定区中的任一个,κ或λ。如果需要,可以通过已知方法转换本公开的抗体类别。典型地,类别转换技术可用于将一个igg亚类转换成另一个,例如从igg1转变为igg2。因此,为各种治疗用途,可以通过同种型转换为例如igg1、igg2、igg3、igg4、igd、iga、ige或igm抗体来改变本公开抗体的效应子功能。在一些实施方案中,本文公开的抗体是全长抗体。在一些实施方案中,全长抗体是igg1抗体。在一些实施方案中,全长抗体是igg4抗体。在一些实施方案中,btn2a特异性igg4抗体是稳定化的igg4抗体。合适的稳定化的igg4抗体的实例是其中人igg4的重链恒定区中第409位的精氨酸(如在以上kabat等的eu索引中所示)被赖氨酸、苏氨酸、甲硫氨酸或亮氨酸,典型地是赖氨酸取代(描述于wo2006033386中)和/或其中铰链区包含cys
‑
pro
‑
pro
‑
cys序列的抗体。其他合适的稳定化的igg4抗体公开于wo2008145142中。
[0257]
在一些实施方案中,本公开的抗体不包含诱导抗体依赖性细胞毒性(adcc)的fc部分。术语“fc结构域”、“fc部分”和“fc区”是指抗体重链的c末端片段,例如,人γ重链从约第230位氨基酸(aa)至约第450位aa或其在其他类型的抗体重链(例如,人抗体的α、δ、ε和μ)中的对应序列,或其天然存在的同种异型。除非另有说明,否则在本公开内容中使用普遍接受的免疫球蛋白的kabat氨基酸编号(参见kabat等(1991)sequences of protein of immunological interest,5th ed,united states public health service,national institute of health,bethesda,md)。在一些实施方案中,本公开的抗体不包含能够基本上结合fcgriiia(cd16)多肽的fc结构域。在一些实施方案中,本公开的抗体缺少fc结构域(例如缺少ch2和/或ch3结构域)或包含igg2或igg4同种型的fc结构域。在一些实施方案中,本公开的抗体由以下组成或包含以下:fab、fab'、fab'
‑
sh、f(ab')2、fv、双体、单链抗体片段或包含多种不同抗体片段的多特异性抗体。在一些实施方案中,本公开的抗体不与毒性部分连接。在一些实施方案中,选自氨基酸残基的一个或多个氨基酸可以用不同的氨基酸残基替换,使得抗体具有改变的c2q结合和/或减少或消除的补体依赖性细胞毒性(cdc)。美国专利no.6,194,551进一步详细描述了该途径。
[0258]
本文预期的本文抗体的另一种修饰是聚乙二醇化。例如,可以将抗体聚乙二醇化以增加抗体的生物学(例如血清)半衰期。为了使抗体聚乙二醇化,抗体或其片段典型地在一个或多个peg基团变得与抗体或抗体片段连接的条件下与聚乙二醇(peg)反应,诸如peg的反应性酯或醛衍生物。聚乙二醇化可以通过酰化反应或与反应性peg分子(或类似的反应性水溶性聚合物)的烷基化反应来进行。如本文所用,术语“聚乙二醇”旨在涵盖用于衍生其他蛋白质的任何形式的peg,例如单(c1
‑
c10)烷氧基
‑
或芳氧基
‑
聚乙二醇或聚乙二醇
‑
马来酰亚胺。在一些实施方案中,待聚乙二醇化的抗体是无糖基化的抗体。使蛋白质聚乙二醇化的方法是本领域已知的,并且可以应用于本公开的抗体。例如,参见nishimura等的ep 0154 316和ishikawa等的ep 0 401 384。
[0259]
本文预期的抗体的另一种修饰是至少本公开抗体的抗原结合区与血清蛋白(诸如人血清白蛋白或其片段)的缀合物或蛋白质融合物,以增加所得分子的半衰期。
[0260]
在一些实施方案中,本公开还提供多特异性抗体。本公开的多特异性抗体分子的示例性形式包括但不限于(i)通过化学异源缀合交联的两种抗体,一种对btn2a具有特异性,另一种对第二抗原具有特异性;(ii)包含两个不同抗原结合区的单个抗体;(iii)包含
两个不同抗原结合区的单链抗体,例如,通过额外的肽接头串联连接的两个scfv;(iv)双可变结构域抗体(dvd
‑
ig),其中每个轻链和重链含有通过短肽连接串联的两个可变结构域(wu等generation and characterization of a dual variable domain immunoglobulin(dvd
‑
ig
tm
)molecule,in:antibody engineering,springer berlin heidelberg 2010);(v)化学连接的双特异性(fab')2片段;(vi)tandab,其是两个单链双体的融合物,产生对于每种靶标抗原具有两个结合位点的四价双特异性抗体;(vii)flexibody,其是scfv与双体的组合,产生多价分子;(viii)基于蛋白激酶a中的“二聚化和对接结构域”的所谓的“对接和锁定”分子,当应用于fab时,可以产生由与不同的fab片段连接的两个相同的fab片段组成的三价双特异性结合蛋白;(ix)所谓的scorpion分子,其包含例如与人fab臂的两个末端融合的两个scfv;和(x)双体。双特异性抗体的另一种示例性形式是具有互补ch3结构域的igg样分子,以推动异源二聚化。这些分子可以使用已知技术制备,诸如称为triomab/quadroma(trion pharma/fresenius biotech)、knob
‑
into
‑
hole(genentech)、crossmab(roche)和静电匹配(amgen)、luz
‑
y(genentech)、链交换工程化结构域体(seedbody)(emd serono)、biclonic(merus)和duobody(genmab a/s)技术的那些。在一些实施方案中,双特异性抗体典型地使用duobody技术通过受控的fab臂交换获得或可获得。通过受控的fab臂交换产生双特异性抗体的体外方法已描述于wo2008119353和wo 2011131746(均由genmab a/s)中。在wo 2008119353中描述的一种示例性方法中,在还原条件下孵育时,通过两种单特异性抗体之间的“fab
‑
臂”或“半分子”交换(交换重链和连接的轻链)形成双特异性抗体,所述两种单特异性抗体均包含igg4样ch3区域。所得产物是具有两个fab臂的双特异性抗体,其可包含不同的序列。在wo 2011131746中描述的另一种示例性方法中,通过包括以下步骤的方法制备本公开的双特异性抗体,其中第一和第二抗体中的至少一种是本公开的抗体:a)提供包含免疫球蛋白的fc区的第一抗体,所述fc区包含第一ch3区;b)提供包含免疫球蛋白的fc区的第二抗体,所述fc区包含第二ch3区;其中所述第一和第二ch3区的序列不同,并且所述第一和第二ch3区之间的异源二聚体相互作用比所述第一和第二ch3区之间的同源二聚体相互作用强;c)在还原条件下孵育所述第一抗体和所述第二抗体;和d)获得所述双特异性抗体,其中所述第一抗体是本公开的抗体,所述第二抗体具有不同的结合特异性,或反之亦然。例如,还原条件可以通过添加例如选自2
‑
巯基乙胺、二硫苏糖醇和三(2
‑
羧乙基)膦的还原剂来提供。步骤d)可以进一步包括将条件恢复为变成非还原或较少还原,例如通过去除还原剂,例如通过脱盐。典型地,第一和第二ch3区的序列不同,仅包含少数相当保守的不对称突变,使得所述第一和第二ch3区之间的异源二聚体相互作用比所述第一和第二ch3区的每个同源二聚体相互作用强。通过引用整体并入本文的wo 2011131746提供了关于这些相互作用及其如何实现的更多细节。以下是这种不对称突变的组合的示例性实施方案,任选地其中一个或两个fc区是igg1同种型。
[0261]
因此,本公开提出包含本文所述的抗btn2a抗体的双特异性或多特异性抗体(也称为双特异性或多特异性分子)。因此,本公开包括双特异性分子,其包含至少一种针对btn2a的第一结合特异性(例如本文公开的抗体的一个抗原结合部分)和针对第二靶标表位的第二结合特异性。例如,根据本公开的双特异性分子可以包括一个抗原结合部分,其包含至少:
[0262]
‑
包含seq id no:3的重链可变区cdr1、包含seq id no:4的重链可变区cdr2、包含
seq id no:5的重链可变区cdr3、包含seq id no:6的轻链可变区cdr1、包含seq id no:7的轻链可变区cdr2和包含seq id no:8的轻链可变区cdr3,或
[0263]
‑
包含seq id no:21的重链可变区cdr1、包含seq id no:22的重链可变区cdr2、包含seq id no:23的重链可变区cdr3、包含seq id no:24的轻链可变区cdr1、包含seq id no:25的轻链可变区cdr2和包含seq id no:26的轻链可变区cdr3。
[0264]
在一个实施方案中,双特异性分子包含针对btn3的第二结合特异性。更具体地,双特异性分子还可包括特异性结合btn3a并激活vγ9/vδ2t细胞的溶细胞功能的抗btn3a激活抗体的抗原结合部分。
[0265]
此外,对于双特异性分子是多特异性的实施方案,除了第一和第二靶标表位之外,分子还可以包括第三结合特异性。
[0266]
在一个实施方案中,本文公开的双特异性分子包含作为结合特异性的至少一种抗体或其抗体片段,包括例如fab、fab'、f(ab')2、fv、unibody或单链fv。抗体也可以是轻链或重链二聚体,或其任何最小片段,例如如ladner等人美国专利号4,946,778所述的fv或单链构建体。
[0267]
可用于本文公开的双特异性分子的其他抗体是鼠、嵌合和人源化单克隆抗体。
[0268]
药物组合物
[0269]
在另一方面,本公开提供组合物,例如药物组合物,其包含与药学上可接受的载体一起配制的至少一种如本文所公开的抗体。此类组合物可包含如上所述的(例如,两种或更多种不同的)抗体的组合。本文公开的药物组合物也可以联合疗法,即与其他试剂联合施用。
[0270]
例如,本发明的抗体典型地可以与至少一种抗病毒剂、抗炎剂或另一种抗增殖剂联合。可用于联合治疗的治疗剂的实例在下文关于本公开的抗体的用途的部分中更详细地描述。
[0271]
如本文所用,“药学上可接受的载体”包括生理学相容的任何和所有溶剂、分散介质、包衣、抗菌剂和抗真菌剂、等渗剂和吸收延迟剂等。载体应适合静脉内、肌内、皮下、肠胃外、脊髓或表皮施用(例如,通过注射或输注)。在一个实施方案中,载体应适用于皮下途径。
[0272]
基于施用途径,活性化合物(即抗体)可以包衣于材料中以保护化合物免受酸和其他可能使化合物失活的自然条件的作用。药物组合物的形式、施用途径、剂量和方案自然取决于待治疗的病症、疾病的严重程度、患者的年龄、体重和性别等。
[0273]
本公开的药物组合物可以配制用于局部、口服、肠胃外、鼻内、静脉内、肌内、皮下或眼内施用等。
[0274]
本发明的用途和方法
[0275]
本公开的抗体具有体外和体内诊断和治疗效用。例如,这些分子可以施用于培养中的细胞,例如体外、离体或体内,或施用于受试者,例如体内,以治疗、预防或诊断多种病症。
[0276]
本公开的抗体是在表型、细胞因子分泌和/或t细胞抑制特性方面抑制单核细胞向促肿瘤m2巨噬细胞分化的抗btn2a1抗体。
[0277]
或者,或优选另外,本公开的抗体在nk细胞的质膜处直接结合btn2a(特别是btn2a1)并触发它们对癌细胞的活化和细胞毒性。
[0278]
在一些实施方案中,本公开的抗体可以进一步激活vγ9/vδ2t细胞的溶细胞功能、细胞因子产生和增殖。
[0279]
因此,本公开的抗体可用于克服在癌症患者和慢性感染期间观察到的免疫抑制机制。
[0280]
在一些实施方案中,本公开的抗体可用于降低肿瘤环境的免疫抑制作用。
[0281]
本文公开的抗btn2a抗体还可以通过作用于肿瘤微环境(通过m1极化和/或m2抑制)和直接作用于nk细胞区室来增强nk和/或th1细胞的细胞毒性作用。
[0282]
在一些实施方案中,本公开的抗体(例如本文所述的107g3抗体及其变体)进一步激活vγ9/vδ2t细胞的溶细胞功能、细胞因子产生和增殖。因此,此类抗体有可能联合作用于免疫的3个细胞区室:nk细胞、巨噬细胞和γδt细胞,因此代表癌症治疗,尤其是实体瘤治疗的强大工具。
[0283]
临床前研究已经证实,nk细胞可以杀死髓系白血病细胞。但是,例如在cml中,nk细胞的数量随着疾病进展而减少,对刺激的反应减弱,并且溶细胞活性降低。在aml中,nk细胞更高的溶细胞活性也预示患者在诊断和缓解期的长期预后更好(carlsten m,m.natural killer cells in myeloid malignancies:immune surveillance,nk cell dysfunction,and pharmacological opportunities to bolster the endogenous nk cells.front immunol.2019)。因此,在一些实施方案中,呈现nk细胞活化特性的本发明抗体可与nk细胞疗法(如过继转移nk细胞疗法)组合使用以恢复nk细胞功能和/或触发或改进它们的细胞毒性。特别地,本技术的抗体可用于治疗通常对nk细胞杀伤具有抗性的实体瘤。如本文所用,术语“癌症”、“过度增殖的”和“肿瘤的”是指具有自发生长能力的细胞,即以快速增殖的细胞生长为特征的异常状态或病症。过度增殖和肿瘤疾病状态可归类为病理性的,即表征或构成疾病状态,或可归类为非病理性的,即偏离正常但与疾病状态无关。该术语意在包括所有类型的癌性生长或致癌过程、转移组织或恶性转化的细胞、组织或器官,而不管组织病理学类型或侵袭阶段。
[0284]
术语“癌症”或“肿瘤”包括各种器官系统的恶性肿瘤,例如影响肺、乳腺、甲状腺、淋巴、胃肠道和泌尿生殖道的恶性肿瘤,以及包括恶性肿瘤(如大多数结肠癌、肺、皮肤或阴道的鳞状细胞癌、肾细胞癌、前列腺癌和/或睾丸肿瘤、非小细胞肺癌、小细胞肺癌、子宫内膜癌、卵巢癌、宫颈管腺癌、胰腺癌、小肠癌和食道癌,以及更一般地可以通过在患有所述癌症的受试者中体内刺激γδt细胞的活化和/或增殖来治疗的任何癌症)。
[0285]
癌症的实例包括但不限于血液恶性肿瘤,如b细胞淋巴肿瘤、t细胞淋巴肿瘤、非霍奇金淋巴瘤(nhl)、b
‑
nhl、t
‑
nhl、慢性淋巴细胞白血病(cll)、小淋巴细胞淋巴瘤(sll)、套细胞淋巴瘤(mcl)、nk细胞淋巴肿瘤和髓细胞系肿瘤,包括急性髓细胞白血病。
[0286]
非血液癌症的实例包括但不限于结肠癌、乳腺癌、肺癌、卵巢癌、脑癌、前列腺癌、头颈癌、胰腺癌、膀胱癌、结肠直肠癌、骨癌、宫颈癌、肝癌、口腔癌、食道癌、甲状腺癌、肾癌、胃癌、睾丸癌和皮肤癌。
[0287]
慢性感染的实例包括但不限于病毒、细菌、寄生虫或真菌感染,如慢性肝炎、肺部感染、下呼吸道感染、支气管炎、流感、肺炎和性传播疾病。
[0288]
因此,本公开内容涉及在有需要的受试者中治疗上文公开的病症之一的方法,所述方法包括治疗有效量的如上文公开的抗btn2a1抗体。
[0289]
用于如上公开的抗体可以作为唯一的活性成分或与例如佐剂或与例如其他药物联合施用以用于治疗或预防上述疾病,所述其他药物例如细胞因子、抗病毒剂、抗炎剂或细胞毒剂、抗增殖剂、化疗剂或抗肿瘤剂、细胞治疗产品(例如γδt细胞组合物或nk细胞组合物)。
[0290]
例如,如上所公开使用的抗体可以与细胞疗法,特别是γδt细胞疗法、nk细胞疗法、化疗、抗肿瘤剂或免疫治疗剂联合使用。
[0291]
如本文所用,术语“细胞疗法”是指包括向有需要的受试者体内施用至少一种治疗有效量的细胞组合物的疗法。向患者施用的细胞可以是同种异体的或自体的。术语“γδt细胞疗法”是指一种细胞疗法,其中细胞组合物包括作为活性成分的γδt细胞,特别是vγ9/vδ2t细胞(例如过继性γδt细胞转移或表达嵌合抗原受体的γδt细胞)。术语“nk细胞疗法”是指一种细胞疗法,其中细胞组合物包括作为活性成分的nk细胞,例如过继性nk细胞转移或表达嵌合抗原受体的nk细胞(car
‑
nk)。
[0292]
细胞治疗产品是指为了治疗目的而向所述患者施用的细胞组合物。所述细胞治疗产品包括治疗有效剂量的细胞和任选的附加赋形剂、佐剂或其他药学上可接受的载体。
[0293]
合适的抗肿瘤剂可以包括但不限于,烷化剂(例如环磷酰胺、氮芥、苯丁酸氮芥、美法仑、亚硝基脲、替莫唑胺)、蒽环类(例如柔红霉素、多柔比星、表柔比星、伊达比星、米托蒽醌、戊柔比星)、紫杉烷(例如紫杉醇、多西他赛)、埃坡霉素、拓扑异构酶i抑制剂(如伊立替康或托泊替康)、拓扑异构酶ii抑制剂(如依托泊苷、替尼泊苷或他氟泊苷)、核苷酸类似物和前体类似物(如阿扎胞苷、硫唑嘌呤、卡培他滨、阿糖胞苷、氟尿嘧啶、吉西他滨、羟基脲、巯基嘌呤、甲氨蝶呤或硫鸟嘌呤)、肽抗生素(如卡铂、顺铂和奥沙利铂)、类视黄醇(如维甲酸、阿利维a酸、贝沙罗汀)、长春花生物碱及其衍生物(如长春碱、长春新碱、长春地碱、长春瑞滨)、靶向治疗如激酶抑制剂(如依鲁替尼、艾代拉里斯、厄洛替尼、吉非替尼、伊马替尼、维木拉非尼、维莫德吉)、蛋白酶体抑制剂(如硼替佐米、卡非佐米)、组蛋白脱乙酰酶抑制剂(如伏立诺他或罗米地辛)。
[0294]
免疫治疗剂的实例包括但不限于磷酸抗原(例如唑来膦酸或其他双膦酸盐)、抗pd
‑
1抗体、抗pd
‑
l1抗体、抗ctla
‑
4抗体和细胞因子(诸如白介素2(il
‑
2)(choudhry h等,2018,biomed res int.)、白介素15(il
‑
15)(patidar m等,cytokine growth factor rev.2016)、白介素21(il
‑
21)(caccamo n.等,plos one.2012)或白介素33(il
‑
33)(duault c等,j immunol.2016),或其重组形式及其衍生物,或任何能够诱导淋巴细胞活性(例如增殖或细胞因子产生或代谢变化)的细胞因子。术语衍生物用于可依赖于peg化(例如与聚乙二醇(peg)链缀合)的任何细胞因子修饰,诸如氨基酸缺失、取代或插入的突变,或与增强剂的结合(例如与igg1 fc融合的il15/il15ra复合物,其中il
‑
15另外发生突变(asn72asp),进一步增强了生物活性,使该复合物成为il
‑
2和il
‑
15rβγ超级激动剂(rhode pr等,cancer immunol res.2016)(barroso
‑
sousa r等,curr oncol rep.2018)。
[0295]
术语“il
‑
2”具有其一般含义,并且是指人白介素
‑
2。il
‑
2主要通过与il
‑
2受体结合来调节淋巴细胞活性。
[0296]
术语“il
‑
15”具有其一般含义,并且是指人白介素
‑
15。与il
‑
2一样,il
‑
15结合并通过由il
‑
2/il
‑
15受体β链(cd122)和通用γ链(γ
‑
c,cd132)组成的复合物发出信号。il
‑
15调节t细胞和自然杀伤(nk)细胞的活化和增殖。
[0297]
术语“il
‑
21”具有其一般含义,并且是指人白介素
‑
21。il
‑
21已被归因于多效性,包括但不限于增强nk细胞和cd8 t细胞的细胞毒性、调节浆细胞分化和抑制treg细胞。
[0298]
术语“il
‑
33”具有其一般含义,并且是指人白介素33。被认为是在组织受到压力或破坏时释放的警报蛋白的il
‑
33是il
‑
1家族的成员,并与st2受体结合。已知il
‑
33是t
h
1免疫细胞、自然杀伤(nk)细胞、inkt细胞和cd8 t淋巴细胞的有效刺激剂。
[0299]
术语“pd
‑
1”在本领域中具有其一般含义,并且是指程序性死亡
‑
1受体。术语“pd
‑
1”还指i型跨膜蛋白,其属于cd28
‑
b7信号转导受体家族的成员,该家族包括cd28、细胞毒性t淋巴细胞相关抗原4(ctla
‑
4)、诱导型共刺激物(icos)以及b和t淋巴细胞减毒剂(btla)(greenwald rj等,2005;riley jl等,2005)。
[0300]
术语“抗pd
‑
1抗体”或“抗pd
‑
l1”具有本领域的一般含义,并且是指分别对pd
‑
1或pd
‑
l1具有结合亲和力且对pd
‑
1具有拮抗活性的抗体,即其抑制与pd
‑
1有关的信号转导级联并抑制pd
‑
1配体结合(pd
‑
l1;pd
‑
l2)。相比于其与受体的cd28
‑
b7信号转导家族的其他亚型或同工型(cd28;ctla
‑
4;icos;btla)的相互作用,此类抗pd
‑
1抗体或抗pd
‑
l1抗体分别优先地以更大的亲和力和效力来灭活pd
‑
1。用于确定化合物是否为pd
‑
1拮抗剂的测试和测定法是本领域技术人员公知的,例如描述于greenwald等(2005)、riley等(2005)中。
[0301]
此类抗pd1抗体的实例包括但不限于尼古拉单抗、派姆单抗、奥维单抗、杜鲁伐单抗、西米普利单抗或阿妥珠单抗。
[0302]
根据前述内容,本公开在又另一方面提供:
[0303]
如上所定义的方法,其包括共同施用(如伴随使用或顺次使用)治疗有效量的本公开的抗btn2a1抗体,和至少一种第二药物,所述第二药物是抗病毒或抗增殖剂或免疫治疗剂或细胞因子或细胞治疗产品(诸如γδt细胞),例如如上文所述。
[0304]
在一个实施方案中,本公开的抗体可用于检测可溶性btn2a1的水平或表达btn2a1的细胞的水平。例如,可通过在允许抗体和btn2a1形成复合物(如在细胞表面或可溶性btn2a1,如血液样本中表达)的条件下,将样品(诸如体外样品)和对照样品与抗btn3a抗体接触来实现。检测并比较样品和对照中抗体与btn2a1之间形成的任何复合物。例如,可用本公开的组合物来进行本领域公知的标准检测方法(诸如elisa和流式细胞仪)进行测定。
[0305]
因此,一方面,本公开进一步提供了用于检测样品中btn2a1(例如人btn2a1抗原)的存在或测量btn2a1的量的方法,其包括在允许抗体或其部分与btn2a1形成复合物的条件下,使样品和对照样品与本公开的抗体或蛋白质或特异性结合btn2a1的抗原结合区接触。然后,检测复合物的形成,其中,与对照样品比较,样品中复合物形成的差异指示样品中btn2a1的存在。
[0306]
试剂盒也包括在本公开的范围内,所述试剂盒由本文公开的组合物(例如人源化抗体、缀合的抗体和多特异性分子)和使用说明书组成。试剂盒可进一步含有至少一种其他试剂,或一种或多种其他抗体或蛋白质。试剂盒典型地包括标明试剂盒内容物预期用途的标签。术语标签包括试剂盒上、随试剂盒提供或随试剂盒附带的任何文字或记录材料。如上定义,试剂盒可进一步包括用于诊断患者是否属于对抗btn2a1抗体治疗有反应的人群的工具。
[0307]
另一种治疗方法是基于将本公开的人源化抗体作为选择性活化从人受试者样品分离的nk细胞的试剂的用途。
[0308]
因此,本公开涉及用于治疗有需要的受试者的方法,包括:
[0309]
(a)从受试者的血液样本中分离出包含nk细胞的血细胞,例如pbmc,
[0310]
(b)在本文公开的抗btn2a1,以及任选地,其他肿瘤或辅助细胞的存在下,体外扩增nk细胞,
[0311]
(c)收集扩增的nk细胞,
[0312]
(d)任选地,配制扩增的nk细胞并向受试者施用治疗有效量的所述nk细胞。
[0313]
本公开进一步涉及本文公开的人源化抗体作为试剂的用途,其选择性地激活嵌合抗原受体(car)nk细胞。car nk细胞及其在过继性nk细胞癌症免疫疗法中的用途例如描述于rezvani,k et al.“adoptive cell therapy using engineered natural killer cells”(bone marrow transplant 2019)中。
[0314]
本公开还涉及本文公开的抗btn2a1抗体,其在有需要的受试者(通常患有癌症)体内用作nk细胞疗法中的增强剂的用途。
[0315]
如本文所用,术语nk细胞疗法是指一种疗法,其包括向有需要的受试者施用至少一种有效量的nk细胞。这种nk细胞可以是同种异体的或自体的。在具体实施方案中,nk细胞可以通过特定基因的缺失或敲除或插入或敲入基因工程化。在具体实施方案中,所述nk细胞包括表达嵌合抗原受体的nk细胞。nk细胞可能已经离体扩增和/或纯化。或者,nk细胞也可包含在包含其他血细胞,例如免疫系统的其他细胞的细胞组合物中。有关γδt细胞疗法的参考资料,请参见rezvani,k.bone marrow transplant 2019。
[0316]
因此,本公开涉及治疗患有癌症(例如血液恶性肿瘤,特别是白血病,诸如急性髓样白血病,并具有肿瘤细胞,例如血液肿瘤细胞)的受试者的方法,所述方法包括:
[0317]
(i)在所述受试者中施用有效量的本文公开的抗btn2a1抗体,和,
[0318]
(ii)在所述受试者中施用有效量的nk细胞组合物,
[0319]
其中所述有效量的抗btn2a1抗体具有增强由所述nk细胞组合物介导的针对所述肿瘤细胞的抗肿瘤溶细胞的能力。本公开还涉及治疗有需要的受试者的方法,所述方法包括联合(同时或顺次)施用nk细胞,例如car nk细胞,和如本文公开的人源化抗体。
[0320]
在替代或另外的实施方案中,治疗策略还可以基于使用如本文公开的人源化抗体作为试剂,其选择性扩增和/或激活从人受试者的样品分离的vγ9/vδ2t细胞。
[0321]
因此,本公开涉及用于治疗有需要的受试者的方法,包括:
[0322]
(a)从受试者的血液样本中分离出包含vγ9/vδ2t细胞的血细胞,例如pbmc,
[0323]
(b)在本文公开的抗btn2a1,以及任选地,其他肿瘤或辅助细胞的存在下,体外扩增vγ9/vδ2t细胞,
[0324]
(c)收集扩增的vγ9/vδ2t细胞,
[0325]
(d)任选地,配制扩增的vγ9/vδ2t细胞并向受试者施用治疗有效量的所述vγ9/vδ2t细胞。
[0326]
本公开进一步涉及如本文公开的人源化抗体作为试剂的用途,其选择性地扩增嵌合抗原受体(car)vγ9vδ2t细胞。carγδt细胞及其在过继性t细胞癌症免疫疗法中的用途描述于例如mirzaei et al,cancer lett 2016中。
[0327]
本公开的抗体还可用于制备人造t细胞受体(也称为嵌合t细胞受体,或嵌合抗原受体(car))。例如,抗体的可变区可用于形成fab或scfv,其通过间隔基连接至tcr的跨膜结
构域和信号内结构域并可在t细胞表面产生。此类car可用于过继性转移疗法,例如用于治疗增殖性疾病。
[0328]
本公开还涉及抗btn2a1抗体在有需要的受试者(典型地患有癌症)体内用作γδt细胞疗法中的肿瘤细胞增强剂的用途。
[0329]
如本文所用,术语γδt细胞疗法是指包括向有需要的受试者施用至少有效量的γδt细胞的疗法。这些γδt细胞可以是同种异体的或自体的。在特定实施方案中,可以通过缺失或敲除或插入或敲入特定基因来对γδt细胞进行基因工程化改造。在特定实施方案中,所述γδt细胞包括表达嵌合抗原受体的γδt细胞。γδt细胞可能已经被离体扩增和/或纯化。或者,γδt细胞也可以包含在含其他血细胞,例如免疫系统的其他细胞的细胞组合物中。有关γδt细胞疗法的参考资料,请参见pauza cd.et al,front immunol.2018j saudemont a.et al,frontiers immunol 2018。
[0330]
实际上,不受任何特定理论的束缚,本公开的抗btn2a1抗体的提议的作用方式是它与在肿瘤细胞表面表达的btn2a1的结合触发构象变化,使其能够在vγ9vδ2t细胞上向其反受体发出信号。
[0331]
因此,本公开涉及治疗患有癌症(例如血液恶性肿瘤,特别是白血病,诸如急性髓样白血病,并具有肿瘤细胞,例如血液肿瘤细胞)的受试者的方法,所述方法包括:
[0332]
(i)在所述受试者中施用有效量的本文公开的抗btn2a1抗体,和
[0333]
(ii)在所述受试者中施用有效量的γδt细胞组合物,
[0334]
其中所述有效量的抗btn3a抗体具有增强由所述γδt细胞组合物介导的针对所述肿瘤细胞的抗肿瘤细胞溶解的能力。本公开还涉及用于治疗有需要的受试者的方法,所述方法包括car t细胞(例如carγδt细胞)和本文公开的人源化抗体的联合(同时或顺次)施用。
[0335]
通过以下附图和实施例将进一步说明本发明。然而,这些实施例和附图不应以任何方式解释为限制本公开的范围。
附图说明
[0336]
图1.抗btn2a1 107g3 mab的鉴定。a.从小鼠免疫到mab测序筛选抗btn2a1 mab的级联。b.条形图显示在主要命中选择期间在luminex上测量的每个亲和力(k
d
)范围内的克隆数。c.堆积条形图显示根据它们在初级(第一轮)和次级(第二轮)命中筛选期间调节通过vγ9/vδ2t细胞产生ifn
‑
γ的能力分类为中性(灰色)、拮抗剂(白色)或激动剂(黑色)的克隆数。
[0337]
图2.抗btn2a1 107g3 mab增强vγ9/vδ2t细胞的溶细胞功能。在存在抗cd107a/b抗体和高尔基体终止,有或没有指定抗体的情况下,将vγ9/vδ2t细胞从3个健康供体的pbmc中扩增(参见材料和方法),并在37℃下与靶细胞共培养,其中效应物:靶标(e:t)比为1:1。4小时后,收集、固定细胞并在流式细胞仪上分析。在a中,在有或没有抗btn2a1 107g3上清液或对照杂交瘤培养基的情况下,使用不同的靶细胞系,包括daudi(伯基特淋巴瘤)、jurkat(急性t细胞白血病)、l
‑
ipc(胰腺癌)和mda
‑
mb
‑
134(乳腺癌)。条形图显示cd107 细胞的百分比,描述了vγ9/vδ2t细胞脱粒。在b中,在存在指定浓度的纯化抗btn2a1 107g3 mab或作为同种型对照的不相关的小鼠igg1的情况下,将daudi细胞用作靶细胞。图显示了
允许ec
50
计算的剂量反应曲线。
[0338]
图3.anti
‑
btn2a1 107g3 mab识别btn2a1但不识别btn3。hek
‑
293t btn2 ko细胞用编码btn2a1
‑
cfp融合蛋白的质粒瞬时转染。a.柱状图显示用纯化的抗btn2a1 107g3 mab(顶部,黑线)、抗btn3 103.2mab(底部,黑线)或migg1或igg2a(虚线)对照染色的指定细胞和细胞转染子的叠加。对于转染的细胞,染色在cfp 细胞上门控后显示。b.图显示了纯化的抗btn2a1 107g3mab与用编码btn2a1
‑
cfp的质粒转染的hek
‑
293t btn2 ko细胞结合的剂量反应曲线。在cfp 细胞上门控后分析所有染色。
[0339]
图4:nk细胞和单核细胞btn2a的表达以及对参考抗btn2a 101g5和107g3 mab的单核细胞到m2巨噬细胞极化的影响。(a)来自未刺激的hd
‑
pbmc的nk细胞和单核细胞上btn2a1和btn2a2表达(白色)对比对照同种型(灰色)的代表性柱状图,通过流式细胞术评估。(b)在m
‑
csf存在下诱导的体外m1/m2巨噬细胞或巨噬细胞101g5和107g3 mab的代表性cd14/cd163点图谱。分化5天后,通过流式细胞术分析生成cd14和cd163点图。
[0340]
图5:参考抗btn2a 101g5和107g3 mab以剂量依赖性方式抑制m2巨噬细胞极化。将m1、m2、用gm
‑
csf和ifnγ恢复的m2以及在不同浓度的101g5或107g3 mab(或其同种型对照)存在下的m
‑
csf诱导的巨噬细胞极化5天,并用或不用lps再刺激2天。通过流式细胞术在未刺激的细胞(a
‑
c)或lps刺激的细胞(d)上分析cd14(a)、cd163(b)、pdl1(c)和cd86(d)的表达。结果以中值荧光强度值(mfi)减去相应的同种型对照表示。通过elisa在lps刺激的巨噬细胞上清液中定量il
‑
10(e)和tnfα(f)。结果以pg/ml表示。
[0341]
图6:参考抗btn2a 101g5和107g3 mab抑制从单核细胞的“m2 il
‑
4”诱导的极化。将m1、m2、“m2 il4”、用gm
‑
csf和ifnγ恢复的m2以及用10μg/ml的m
‑
csf il
‑
4和101g5或107g3 mab(或其同种型对照)诱导的巨噬细胞产生5天,并用或不用lps再刺激2天。通过流式细胞术在未刺激的细胞(a
‑
d)或lps刺激的细胞(e)上分析cd14(a)、cd163(b)、pdl1(c)、dc
‑
sign(d)和cd86(e)的表达。结果以中值荧光强度值(mfi)减去相应的同种型对照表示。通过elisa在lps刺激的巨噬细胞上清液中定量il
‑
10(f)和tnfα(g)。结果以pg/ml表示。
[0342]
图7:参考抗btn2a 101g5和107g3 mab抑制癌细胞诱导的m2极化。将m1、m2、panc
‑
1条件培养基诱导的m2、用gm
‑
csf和ifnγ恢复的m2以及用10μg/ml的panc
‑
1条件培养基和101g5或107g3 mab(或其同种型对照)诱导的巨噬细胞产生5天,并用或不用lps再刺激2天。通过流式细胞术在未刺激的细胞上分析cd14(a)、cd163(b)的表达。结果以中值荧光强度值(mfi)减去相应的同种型对照表示。通过elisa在lps刺激的巨噬细胞上清液中定量il
‑
10(c)和tnfα(d)。结果以pg/ml表示。
[0343]
图8:参考抗btn2a 101g5和107g3 mab将m2巨噬细胞恢复为促炎m1巨噬细胞:表型和细胞因子分泌。将m1、m2由单核细胞产生5天。5天后,将10μg/ml的101g5或107g3 mab(或同种型对照)或ifnγ添加到m2巨噬细胞上2天,并用或不用lps再刺激2天。通过流式细胞术在未刺激的细胞(a
‑
c)或lps刺激的细胞(d)上分析cd14(a)、cd163(b)、pdl1(c)和cd86(d)的表达。结果以中值荧光强度值(mfi)减去相应的同种型对照表示。通过elisa在lps刺激的巨噬细胞上清液中定量il
‑
10(e)和tnfα(f)。结果以pg/ml表示。
[0344]
图9:参考抗btn2a 101g5和107g3 mab释放m2介导的t细胞增殖和ifnγ分泌的抑制。将在101g5和107g3 mab(或其同种型对照)存在下诱导的分化的m1、m2或巨噬细胞与同种异体okt3激活的ctv标记的cd3 t细胞共培养5天。共培养后,用pma/离子霉素和
golgistop蛋白抑制剂刺激细胞5小时,然后通过流式细胞术定量cd3 t细胞数(a和b)、胞内ifnγ产生(c
‑
f)和增殖(celltrace violet,ctv dim)(g
‑
j)。通过稀释ctv染料(第0天的ctv信号作为基线)定量增殖。结果是在countbright绝对计数珠上校准的cd3 t细胞的绝对数(b、e、f、i和j)或cd3 t细胞的百分比(c、d、g和h)。
[0345]
图10:参考抗btn2a 101g5和107g3 mab对纯化nk细胞的活化和细胞毒性的影响。(a
‑
b)将纯化的nk细胞与参考抗btn2a 101g5和107g3 mab(或对照同种型)在il
‑
2或il
‑
2/il
‑
15刺激下培养5天。在指定的mab或对照同种型存在下,通过评估未刺激和il
‑
2/il
‑
15刺激的nk细胞内的cd69(a)和cd25(b)表达(mfi)来评估nk细胞活化。(c
‑
d)在存在或不存在il
‑
2/il
‑
15刺激下,将纯化的nk细胞与参考抗btn2a 101g5和107g3 mab(或对照同种型)预孵育过夜,然后与人肿瘤细胞系共培养4个小时。在指定的mab或对照同种型存在下,通过流式细胞术将nk细胞脱颗粒评估为未刺激(c)和il
‑
2/il
‑
15刺激的nk细胞(d)中cd107αβ相对于每个肿瘤细胞系的百分比。(e)与添加到共培养中未预孵育的mab相比,当参考抗btn2a 101g5和107g3 mab(或对照同种型)在共培养4小时之前预先在nk细胞或靶细胞上预孵育时,nk细胞针对a549细胞系脱粒。
[0346]
图11:参考抗btn2a 101g5和107g3 mab增强nk细胞脱粒和杀死腺癌细胞系。(a)在存在或不存在il
‑
2/il
‑
15刺激下,将纯化的nk细胞与参考抗btn2a 101g5和107g3 mab(或对照同种型)预孵育过夜,然后与du
‑
145细胞系共培养4小时。通过流式细胞术将nk细胞脱粒评估为cd107αβ 细胞的百分比。使用prism软件上的四参数剂量反应曲线计算指定mab的nk细胞脱粒增强的ec
50
。(b)纯化的nk细胞与参考抗btn2a 101g5和107g3 mab(或对照同种型或il
‑
2/il
‑
15刺激)预孵育过夜,然后与hl
‑
60和a549细胞系共培养4小时。在指定的mab或对照同种型存在下,通过获得caspase 3/7 细胞的百分比来评估nk细胞介导的癌细胞死亡。
[0347]
图12:参考抗btn2a1 101g5和107g3 mab针对btn2a1的分箱实验。分箱实验在基于生物层干涉测量(bli)技术的octet red96平台上进行。将107g3和101g5以成对组合方式针对rhbtn2a1
‑
his蛋白进行测试。a.107g3饱和,101g5竞争;b:101g5饱和并且107g3竞争;c:以任意单位结合和自阻断mab对的测量值。
[0348]
图13:btn2a1的胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶肽。96.37%的序列被鉴定的肽覆盖。
[0349]
图14:参考mab 107g3和101g5与人btn2a1之间的相互作用。a.107g3/btn2a1。b.101g5/btn2a1。
[0350]
图15:相互作用btn2a1/107g3。btn2a1 pdb结构4f9p在表位位点上呈灰色。蓝色的btn2a1氨基酸对应于提供的btn2a1序列的65
‑
78(rwfrsqfspavfvy)和84
‑
100(rteeqmeeyrgrttfvs)。a、b、c、d、e:前视图(a)、后视图(b)、侧视图1(c)、侧视图2(d)和顶视图(e)的带/表面表示。f、g、h、i、j:前视图(f)、后视图(g)、侧视图1(h)、侧视图2(i)和顶视图(j)的带表示。
[0351]
图16:相互作用btn2a1/101g5。btn2a1 pdb结构4f9p在表位位点上呈灰色。蓝色的btn2a1氨基酸对应于提供的btn2a1序列的212
‑
229(ksvrnmscsinntllgqk)。a、b、c、d、e:前视图(a)、后视图(b)、侧视图1(c)、侧视图2(d)和顶视图(e)的带/表面表示。f、g、h、i、j:前视图(f)、后视图(g)、侧视图1(h)、侧视图2(i)和顶视图(j)的带表示。
[0352]
图17:评估参考抗btn2a1 101g5和107g3 mab针对食蟹猴btn2a1直系同源物的交叉反应性。基于elisa测量107g3和101g5与重组人btn2a1
‑
fc融合蛋白或包被在elisa板上的重组食蟹猴btn2a1
‑
fc融合蛋白的结合。图描绘了允许使用可变斜率模型通过非线性回归计算ec
50
的剂量反应曲线。
具体实施方式
[0353]
材料和方法
[0354]
细胞培养、单核细胞和nk细胞分选:
[0355]
外周血单核细胞(pbmc)来自edta(乙二胺
‑
四乙酸)
‑
当地血库(etablissementdu sang(efs)
‑
marseille
‑
france)提供的健康供体(hd)的血沉棕黄层,并在密度梯度上(eurobio)通过离心分离。将新鲜的pbmc在37℃、5%co2下在补充有10%胎牛血清(fbs)和1%青霉素/链霉素(p/s)的roswell park memorial institute培养基1640(rpmi;lonza)中培养。
[0356]
按照制造商的说明,使用easysep
tm
人nk细胞富集试剂盒(stemcell technologies)通过负选择从新鲜pbmc中分选自然杀伤(nk)细胞。按照制造商的说明,使用cd14 微珠试剂盒(miltenyi)分选人cd14 单核细胞。在37℃下将单核细胞以106个细胞/ml的密度在补充有1%l
‑
谷氨酰胺、100u/ml青霉素/链霉素、1mm丙酮酸钠、10mm hepes、0.1mm非必需氨基酸和10%fbs(均来自thermofisher)的rpmi中培养5天。进胰腺癌细胞系panc
‑
1在补充有10%fbs的rpmi中培养。一旦生长至90%汇合,弃去培养基,并将细胞在pbs 1x中冲洗两次。然后,将panc
‑
1细胞在补充有5%fbs的rpmi中再培养24小时(每175cm2烧瓶30ml,以获得浓缩的上清液)。然后,收集panc
‑
1条件培养基,过滤(0.2μm)并储存在
‑
20℃直至使用。其他人细胞系及其相应的培养基总结于下表1中:
[0357]
表1
[0358][0359][0360]
以下人细胞系来自美国典型培养物保藏中心:daudi(伯基特淋巴瘤)、jurkat(急性t细胞白血病)、mda
‑
mb
‑
134(乳腺导管癌)和hek
‑
293t(胚胎肾)。人胰腺癌细胞系l
‑
ipc
(pdac087t)由juan iovanna博士友情提供。将daudi和jurkat细胞以及pbmc在补充有10%胎牛血清(fcs)、1%丙酮酸钠、1%l
‑
谷氨酰胺(均来自life technologies)的rpmi 1640培养基中培养。hek
‑
293t btn2 ko细胞通过crispr
‑
cas9介导的btn2所有亚型失活产生(数据未显示)。将mda
‑
mb
‑
134、l
‑
ipc、hek
‑
293细胞和hek
‑
293t btn2 ko细胞在含有10%fcs的dmem培养基(life technologies)中培养。将杂交瘤在dmem/ham's f12(1:1)(thermofisher scientific)、4%fetalclone i(hyclone)、化学定义脂质浓缩物(1:250)、1%谷氨酰胺、1%丙酮酸钠和100μg/ml penstrep(全部来自thermofisher scientific)中培养。为了收集杂交瘤上清液,在没有fetalclone的情况下培养杂交瘤4
‑
5天。
[0361]
为了评估抗btn2a1 mab的特异性,按照制造商的说明,使用lipofectamine 3000试剂(thermofisher scientific),用编码btn2a1和btn2a2 cfp(nter)
‑
融合蛋白的pcdna3
‑
zeo
‑
btn2a1
‑
cfp质粒独立转染hek
‑
293t btn2 ko细胞。
[0362]
参考抗btn2a1 mab 107g3的鉴定
[0363]
通过用重组人btn2a1
‑
fc融合蛋白免疫48只带有6种不同mhc组合的小鼠而产生小鼠抗人btn2a1抗体。21天后将小鼠放血,并通过luminex测定确定btn2a1特异性多克隆抗体的血清滴度。将显示最高btn2a1特异性抗体滴度的小鼠安乐死。通过阳性选择分离脾b细胞,并进行peg诱导的骨髓瘤细胞融合以产生杂交瘤。
[0364]
通过有限稀释克隆杂交瘤,并且杂交瘤上清液针对靶特异性及其诱导vγ9vδ2
‑
t细胞脱粒的能力进行两轮筛选(图1c和2),并鉴定出参考mab 107g3。对这些亚克隆的vh和vl区进行测序(见表1)。
[0365]
vγ9vδ2
‑
t细胞的扩增
[0366]
通过从第0天开始在唑来膦酸盐(sigma,1μm)和重组人(rh)il
‑
2(proleukin,200iu/ml)存在下培养来自hv的pbmc建立效应vγ9/vδ2
‑
t细胞。从第5天开始,rhil
‑
2每隔一天更新一次,并且将细胞密度保持在1.15x 106个/ml,总共15天。最后一天,通过流式细胞术评估vγ9/vδ2
‑
t细胞的纯度。仅选择达到vγ9/vδ2
‑
t细胞纯度高于80%的细胞培养物用于功能测试。冷冻纯化的vγ9/vδ2
‑
t细胞直至使用。
[0367]
luminex测定
[0368]
根据制造商的说明,将磁性cooh珠(biorad)与rhbtn2a1蛋白(r&d)缀合,并将珠储存在
‑
20℃的储存缓冲液(biorad)中直至使用。为了滴定小鼠血清,在luminex测定缓冲液(nanotools)中从1:50开始,通过1:4的稀释步骤进行系列血清稀释;将100μl珠悬浮液与100μl血清稀释液混合并在室温下孵育1小时,然后将珠在洗涤缓冲液中洗涤3次,在luminex测定缓冲液中与1μg/ml生物素化山羊抗小鼠igg
‑
fc一起孵育,和在luminex测定缓冲液中再洗涤3次。最后,将珠在luminex测定缓冲液中与1μg/ml链霉亲和素pe一起孵育1小时,然后在luminex读取缓冲液(nanotools)中最后洗涤3次。将珠重悬于luminex读取缓冲液中,并在luminex 100/200系统上采集数据。对于命中鉴定,将30μl上清液转移到96孔板中,并添加90μl luminex测定缓冲液。将100微升珠悬浮液与100μl上清液稀释液混合,并在室温下孵育16小时,然后继续进行上述方案。对于命中识别,选择对靶标具有最高亲和力和对不相关对照蛋白(rank
‑
fc)具有最低亲和力的那些。对于亲和力/kd计算,将杂交瘤上清液在luminex测定缓冲液中从40.000pm开始,通过1:4的稀释步骤进行连续稀释,并如上所述进行分析。kd对应于相应结合曲线的中点。
eurofins)以验证抗btn2a mab,然后大规模(maxi)制备用于进一步转染的质粒。将编码每个抗btn2a克隆的匹配轻链和重链的载体瞬时转染到hek
‑
293细胞(2.9x106个细胞/ml)中,其中重链/轻链比例为1:1.2,并且18小时后更新培养基。转染后7天,收集培养物上清液用于mab纯化。抗体的亲和纯化使用protein a sepharose fast flow(ge healthcare)进行,44℃下过夜。结合缓冲液是0.5m甘氨酸,3m nacl,ph8.9。使用以下缓冲液进行洗脱:0.1m柠檬酸盐,ph3。样品在洗脱后立即用1m tris
‑
hcl,ph9(10%v/v)中和。最后,将嵌合抗btn2a mab透析到pbs 1x中并通过0.22μm过滤器(millex gv亲水性pvdf,millipore)过滤。考虑到抗体的消光系数,在nanodrop 2000分光光度计(thermoscientific)中测定嵌合抗btn2a mab浓度。使用acquity uplc
‑
hclass bio(waters)和acquity uplc protein
‑
beh
‑
200a,1.7μm 4.6x 50mm色谱柱(waters)通过uplc
‑
sec测定由mab单体的分数定义的纯度。使用反相色谱柱(plrp
‑
s 4000a,5μm,50x 2.1mm(agilent technologies)在xevo g2
‑
s q
‑
tof质谱仪(waters)中测定抗体质量。所有样品均在去糖基化后使用pngase f糖苷酶(new england biolabs)在37℃下分析。当发现意外质量时,使用生物信息学工具分析初级氨基酸序列以鉴定fab区内推定的糖基化位点。纯化抗体的sds
‑
page允许检测最终材料无染色mini protean tgx凝胶4
‑
15(biorad)的碎片和/或聚集。使用clariostar分光光度计(bmg labtech)使用chromogenic lal limulus amebocyte lysate动力学分析(charles river endosafe)测定内毒素水平。
[0379]
体外巨噬细胞极化测定:
[0380]
将m1或m2巨噬细胞从健康供体的分选单核细胞中极化。为此,在gm
‑
csf或m
‑
csf(40ng/ml;miltenyi)存在的情况下培养分选的单核细胞,以分别产生m1或m2巨噬细胞。5天后,收集产生的巨噬细胞用于表型分析,或用lps(200ng/ml)再刺激2天。在一些实验中,在第4天将il
‑
4(20ng/ml)添加到m2巨噬细胞中,导致产生“m2 il
‑
4”或巨噬细胞。在一些实验中,通过在没有m
‑
csf补充的情况下在panc
‑
1癌细胞条件培养基(在培养基中稀释30%v/v,在第0天和第3天)下培养单核细胞产生m2巨噬细胞。由此产生的m2巨噬细胞在本技术中称为“tum
‑
ind
‑
m2”。为了筛选抗btn2a mab调节m2分化的能力,如上所述在有或没有指定浓度的嵌合抗btn2a mab或其同种型对照(人igg1;sigma)的情况下,从单核细胞产生m2巨噬细胞。将所有mab湿法包被(在室温下在pbs 1x中过夜)。作为m2分化抑制的对照,在m
‑
csf诱导的m2极化过程中,将gm
‑
csf(40ng/ml)和ifnγ(100ng/ml,biotechne)添加到单核细胞中。在gm
‑
csf存在下极化的m1巨噬细胞用作表型对照。极化后,收集得到的巨噬细胞及其培养上清液,并通过流式细胞术评估质膜上m1和m2相关标志物的表达。此外,按照制造商的说明,使用il
‑
10和tnfαelisa试剂盒(thermofisher scientific)定量培养上清液中的细胞因子含量。
[0381]
体外m2巨噬细胞逆转试验:
[0382]
在不存在参考mab的情况下如上所述在m
‑
csf存在下从单核细胞产生m2巨噬细胞。收集m2巨噬细胞并在有或没有lps的情况下在培养孔上培养2天,所述培养孔之前用10μg/ml参考抗体或其对照同种型mab(来自sigma的人igg1)湿法包被过夜。作为m2逆转的对照,将gm
‑
csf(40ng/ml)和ifnγ(100ng/ml)添加到m2巨噬细胞培养物中2天。在gm
‑
csf存在下极化的m1巨噬细胞用作表型对照。逆转实验后,收集未添加lps逆转的巨噬细胞,通过流式细胞术进行表型分析。使用elisa对来自lps刺激的逆转巨噬细胞的培养物上清液进行细胞
因子定量收集。
[0383]
m2巨噬细胞介导的t细胞增殖和ifnγ产生抑制的体外测定
[0384]
如上所述,在添加或不添加参考抗体或其同种型对照mab的情况下产生m1和m2巨噬细胞。根据制造商的说明,使用cd3 微珠试剂盒(miltenyi)从健康供体pbmc中分选cd3 t细胞并冷冻直至共培养。活化的cd3 t细胞如下产生:将cd3 t细胞用5μm celltrace violet染料(thermofisher scientific)染色,然后在之前用1μg/ml抗cd3 mab(克隆okt3,bd biosciences)包被的96孔平板中在补充有20u/ml il
‑
2(miltenyi)、lps(200ng/ml)和countbright绝对计数珠(每孔5x103个,thermofisher scientific)的x
‑
vivo 10培养基中培养105个这样的细胞。为了与巨噬细胞共培养,将2x104个同种异体m1、m2或在m
‑
csf和参考mab或其对照同种型存在下极化的巨噬细胞添加到活化的同种异体cd3 细胞中。共培养5天后,在golgistop蛋白转运抑制剂存在下将20ng/ml pma和0.5μg/ml离子霉素添加到共培养物中以增强细胞因子的产生,持续5小时。然后,通过流式细胞术回收细胞用于表型分析。celltrace稀释度用作cd3 t细胞增殖的指标。表型和增殖的结果以百分比或每ml绝对细胞数表示(在使用绝对计数珠校准后)。
[0385]
使用参考抗btn2a1 mab进行自然杀伤(nk)挑战:
[0386]
将来自健康供体的分选自然杀伤(nk)细胞标记,然后在37℃、5%co2下在补充有10%fbs和1%p/s、il
‑
2(50ui/ml)的rpmi中在有或没有il
‑
15(10ng/ml)刺激下培养。在第0天将参考抗btn2a mab或对照同种型(10μg/ml)添加到培养物中。5天后,对nk细胞进行细胞外表型分析以表达活化标志物。通过诱导cd69和cd25表达(百分比和中值荧光强度mfi)来评估nk激活。nk细胞的门控策略如图4所示。对于nk细胞毒性测量,将来自3个健康供体的分选nk细胞在37℃、5%co2下在补充有10%fbs和1%p/s的rpmi中在有或没有il
‑
2(50ui/ml)和il
‑
15(10ng/ml)的条件下培养。将参考抗btn2a mab或对照同种型(10μg/ml)添加到未刺激或il
‑
2/il
‑
15刺激的nk细胞上过夜。第二天,将nk细胞与指定的血液或癌细胞系以1:1的比例共培养,并将fitc标记的抗cd107a和抗cd107b mab(均来自bd biosciences)添加到共培养物中并孵育4小时。通过流式细胞术将nk细胞脱粒评估为未刺激或il
‑
2/il
‑
15刺激的nk细胞上cd107ab 细胞百分比。为了计算nk细胞脱粒增强的ec
50
,参考抗btn2a mab及其同种型对照mab以0.005nm
‑
300nm的浓度使用。对于癌细胞nk细胞介导的杀伤评估,将纯化的nk细胞10μg/ml的参考101g5和107g3 mab或相应的igg1对照在37℃、5%co2下在补充有10%fbs和1%p/s il
‑
2(50ui/ml)和il
‑
15(10ng/ml)的rpmi中预孵育过夜,并用作阳性对照。第二天,将nk细胞与指定的celltrace标记的癌细胞系以1:1的比例共培养4小时。通过使用cellevent
tm caspase
‑
3/7绿色检测试剂(thermofisher scientific)在肿瘤细胞系上获得caspase 3/7 细胞的百分比来评估癌细胞死亡。
[0387]
流式细胞术:
[0388]
在染色之前,将pbmc/nk细胞和单核细胞/巨噬细胞与人fc阻断(miltenyi)或人igg1(sigma)孵育10分钟以使fc受体饱和。使用的标记mab如下:cd14
‑
fitc和
‑
apc
‑
vio770(miltenyi)、cd163
‑
vioblue(miltenyi)、dc
‑
sign
‑
pe
‑
vio770(miltenyi)、cd80
‑
pe(bd biosciences)、pdl1
‑
apc(bd biosciences)、cd3
‑
pe
‑
cf594(bd biosciences)和cd3
‑
bv605(biolegend)、cd56
‑
pe
‑
vio770(miltenyi)和
‑
bv605(bd biosciences)、cd69
‑
bv421(bd biosciences)、cd25
‑
apc(bd biosciences)。将细胞与抗体混合物在4℃下孵育30分钟。使
用活/死近红外染料(thermofisher scientific)来定义“活”门,而排除死细胞。对于细胞内ifnγ染色,使用细胞内固定和透化缓冲液组(ebioscience)固定和透化细胞外染色的细胞,并与apc标记的抗ifnγ(bd biosciences)一起孵育。使用flowjo 10软件在fortessa流式细胞仪(bd biosciences)上进行采集。对于btn2a1和btn2a2表型分析,使用10μg/ml的纯化抗btn2a1特异性(mab5)和抗btn2a2特异性(mab17),并用pe标记的抗igg(h l)(jackson immunoresearch)显示。选择单个细胞后,nk细胞是“活”门内的cd45 cd14
‑
cd3
‑
cd56 细胞。选择单个细胞后,单核细胞是“活”门内的cd45 cd19
‑
cd3
‑
cd56
‑
cd14 细胞。在cytoflex ls(beckman coulter)、ique screener(intellicyt)或fortessa(bd biosciences)流式细胞仪上进行采集,并使用flowjo 10软件分析数据。结果表示为减去相应染色对照获得的值后的中值荧光强度(mfi)。
[0389]
基于octet的btn2a1表位亲和力测量和分箱测定:
[0390]
在生成嵌合igg1形式的参考抗btn2a抗体后,评估了该靶标的2种不同亚型(btn2a1和btn2a2)的亲和力,并进行了竞争测定以确定这些mab是否识别btn2a1的相同表位区域。在基于生物层干涉测量(bli)技术的octet red96平台(fortebio/pall)上进行亲和力和分箱实验。对于亲和力实验,根据制造商的说明,使用ez
‑
link
tm
nhs
‑
peg4生物素化试剂盒将重组人(rh)btn2a1
‑
fc(gtp)生物素化,并且生物素化的rhbtn2a2
‑
fc购自r&d systems。在btn2a1亲和力测定的情况下,将生物素化的rhbtn2a1
‑
fc加载到在kinetic buffer 1x(fortebio)中稀释的链霉亲和素(sa)生物传感器(fortebio)中,加载靶标水平约为1nm,并使用嵌合抗btn2a抗体作为分析物。对于btn2a2亲和力测定,如上所述将嵌合抗btn2a抗体加载到fab2g传感器(抗人ch1;fortebio)中,并且使用生物素化的rhbtn2a2
‑
fc作为分析物。在这两种情况下,分析物保留在溶液中,并且将其工作浓度在kinetic buffer 10x(fortebio)中稀释。对于第一次运行,标准工作浓度范围为200
‑
3.125nm。当第二次运行需要时,将工作浓度从80调整到1.25nm。所有运行(包括加载、平衡、传感器结合/浸入分析物、解离和再生)均在30℃下以1000rpm的转速进行。使用由octet软件计算的1:1或2:1langmuir模型(分别为btn2a1或btn2a2)进行分析,这允许更好的拟合计算。对于分箱实验,his标记的btn2a1(rhbtn2a
‑
his)购自r&d systems。以成对方式针对btn2a1测试参考抗btn2a抗体。通过遵循“串联”格式进行分箱实验,意味着将rhbtn2a
‑
his固定在生物传感器(抗penta
‑
his《his1k生物传感器;fortebio/pall)上,并在连续步骤中呈递给2个竞争抗体。对于此动力学筛选,将rhbtn2a
‑
his加载到his1k(信号强度:1nm)之后,然后是与10μg/ml抗体结合3分钟的步骤,然后是3分钟的解离步骤。rhbtn2a1
‑
his活性通过以与分箱测定相同的格式进行的动力学筛选测定而证实(btn2a1作为传感器上的配体/捕获物,并且抗体作为分析物)。所有抗体(饱和抗体或竞争抗体)均以10μg/ml的浓度使用,在kinetic buffer 1x中稀释。对于该动力学筛选,在his1k上加载rhbtn2a1
‑
his(信号强度:1nm),然后是与抗体结合3分钟的步骤,然后是3分钟的解离步骤。测定步骤如下:基线
‑
>抗原捕获
‑
>基线
‑
>饱和抗体
‑
>基线
‑
>竞争抗体
‑
>遵循“串联”方案的再生。使用表位分箱操作使用octet数据分析ht 11.1分析分箱数据。
[0391]
参考mab 107g3和101g5的表位定位
[0392]
btn2a1与参考mab 107g3和101g5之间的相互作用通过对单独的btn2a1或与107g3或101g5的肽质量指纹的差异评估来评估。在开始表位定位之前,已在rhbtn2a1
‑
fc蛋白
101g5和107g3或对照人igg1在2%pbs bsa中稀释10倍级联稀释,从1μm
‑
1pm,并且每孔添加100μl每种稀释液,并在室温下在振动筛上孵育90分钟。将所有孔在pbs中洗涤3次,然后添加山羊抗小鼠igg hrp(jackson immunoresearch,在pbs bsa 2%中以1:10000稀释)并在室温下孵育1小时。然后,将所有孔在pbs中洗涤3次,并添加1步abts溶液(thermofisher)用于结合显示,如在spark光谱仪(tecan)中通过405nm处的吸光度评估。所有样品一式两份进行评估。
[0395]
结果:
[0396]
参考抗体抗btn2a1 107g3的鉴定
[0397]
参考抗btn2a1 107g3抗体鉴定如下:用btn2a1
‑
fc抗原免疫小鼠,收集表现出最高滴度btn2a1特异性血清的小鼠的脾细胞,并与骨髓瘤细胞系融合以获得杂交瘤。根据它们调节vγ9/vδ2
‑
t细胞分泌ifn
‑
γ的能力,选择对btn2a1表现出最高亲和力的杂交瘤培养上清液进行第一轮筛选。将从第一轮筛选中选出的克隆亚克隆并测试它们诱导ifn
‑
γ分泌和vγ9/vδ2
‑
t细胞脱粒和ifn
‑
γ分泌的能力,特别是它们诱导vγ9vδ2
‑
t细胞脱粒的能力(图1c和2),并导致参考mab 107g3的鉴定。对这些亚克隆的vh和vl区进行测序(见下表2)。
[0398]
抗btn2a1 107g3抗体诱导针对不同癌细胞靶标的vγ9vδ2
‑
t细胞脱粒
[0399]
从健康供体的pbmc中扩增纯化的vγ9/vδ2t细胞,并在有或没有抗btn2a1 107g3杂交瘤培养上清液的情况下与不同的癌细胞系共培养,包括daudi(伯基特淋巴瘤)、jurkat(急性t细胞白血病)、l
‑
ipc(胰腺癌)和mda
‑
mb
‑
134(乳腺癌)作为靶细胞。如图2和表3所示,与单独与靶细胞共培养或在对照杂交瘤培养基存在的情况下相比,添加抗btn2a1 107g3杂交瘤上清液会诱导vγ9/vδ2t细胞的溶细胞功能,如通过cd107 脱粒细胞的百分比所测量。如所预期,pma/离子霉素处理vγ9/vδ2t细胞最大程度地诱导其独立于靶细胞的溶细胞功能。
[0400]
由抗btn2a1 107g3杂交瘤上清液诱导的cd107 细胞百分比范围从daudi细胞中的71.1
±
7.4%到mda
‑
mb
‑
134细胞中的17.1
±
2.9%vs分别在与对照杂交瘤培养基共培养的24.9
±
4.7%和4.9
±
0.4%。在vγ9/vδ2t细胞的共培养物中,以所有测试的癌细胞系为靶标,在对照杂交瘤培养基存在下,与相同的共培养物相比,抗btn2a1 107g3诱导的vγ9/vδ2t细胞脱粒增加了2
‑
8倍。
[0401]
在增加浓度(0
‑
18μg/ml)的抗btn2a1 107g3 mab存在下与作为靶标的daudi细胞共培养,纯化的抗btn2a1 mab 107g3的诱导vγ9/vδ2t细胞脱粒的ec
50
为0.77μg/ml(95%ic 0.32
‑
13.22μg/ml),如cd107 细胞百分比所示(图2b)。
[0402]
表3
[0403][0404][0405]
抗btn2a1 107g3抗体识别btn2a1但不识别btn3
[0406]
为了确定抗btn2a1 mab 107g3仅对btn2a1亚型具有特异性,将具有crispr
‑
cas9介导的两种btn2亚型失活的hek
‑
293t btn2 ko细胞用编码btn2a1的质粒瞬时转染,作为cfp
‑
融合蛋白。如图3a所示,仅在用btn2a1编码质粒转染的hek
‑
293t btn2ko细胞中检测到纯化的抗btn2a1 mab 107g3染色,而在单独的hek
‑
293t btn2 ko细胞中未检测到。
[0407]
识别所有btn3亚型的抗btn3 mab 103.2容易检测hek
‑
293t btn2 ko细胞中btn3表达。因此,抗btn2a1 107g3对btn2a1亚型具有特异性,并且不会与btn3发生交叉反应。
[0408]
抗btn2a1 107g3 mab对细胞中btn2a1的亲和力
[0409]
为了测量抗btn2a1 107g3 mab对其靶标的亲和力,用btn2a1编码质粒转染的hek
‑
293t btn2 ko细胞用增加浓度(5ng/ml
‑
75μg/ml)的纯化抗btn2a1 107g3 mab或对照migg1染色(图3b)。平均荧光强度数据的非线性回归分析发现抗btn2a1 107g3 mab的ec
50
为0.32μg/ml(95%ic 0.21
‑
0.46μg/ml)。
[0410]
嵌合抗btn2a mab的产生、亲和力测量和btn2a亚型特异性:
[0411]
在hek
‑
293t细胞中瞬时产生23种单克隆抗体,实现了不同范围的生产力。大多数抗btn2a抗体以高水平(>100mg/l和高达430mg/l)产生。一种抗体,抗btn2a mab3(表3),在hek
‑
293t细胞中的产量非常低,范围为6
‑
8mg/l。氨基酸序列分析揭示了其fab部分中的n
‑
糖基化位点,在其vh的cdr1中。其他两种抗体,抗btn2a mab9和mab11,也在其fab区内(在mab9的cdr1_vh和mab11的cdr1_vl中)表现出n
‑
糖基化位点。六种抗体的纯度水平较低(单体中<95%),但只有抗btn2a mab1和mab3的纯度水平<90%(分别为86%和75%)。所有最终
纯化的抗btn2a mab都表现出非常低的内毒素水平(在0.1eu/mg的范围内)。只有mab3的内毒素水平(0.73eu/mg)高于其他单克隆抗体,但仍处于可接受的标准范围内(<1eu/mg)。通过使用生物素化重组fc融合可溶性蛋白,使用octet技术测定23种抗btn2a嵌合mab的亲和常数(k
d
、k
on
和k
off
)。表4概括了每种抗btn2a mab的k
d
常数。对于mab6和mab9,由于在测量期间没有观察到解离(k
off
<10
‑7s
‑1),因此无法计算k
d
,这可以通过这些抗体的亲合力效应减缓它们与靶标的解离来解释。发现8种抗btn2a mab仅结合btn2a1亚型(mab2、mab3、mab4、mab5、mab6、mab8、mab9和mab10),发现8种抗btn2a mab仅结合btn2a2亚型(mab16、mab17、mab18、mab19、mab20、mab21、mab22和mab23)并发现7种mab结合两种亚型(mab1、mab7、mab11、mab12、mab13、mab14和mab15)。
[0412]
表4:嵌合抗btn2a mab产生和亲和力总结。
[0413]
[0414][0415]
*在vh或vl中存在n
‑
糖基化位点。
[0416]
单核细胞和nk细胞上的btn2a1和btn2a2质膜表达:
[0417]
我们试图确定抗btn2a mab是否可以针对外周血的非vγ9vδ2t细胞区室,即单核细胞和nk细胞进行治疗。因此,我们使用在我们的octet测定中发现的分别仅结合btn2a1或btn2a2的mab5和mab17进行来自外周血的单核细胞和nk细胞的表型分析。如图4所示,只有抗btn2a1 mab5染色单核细胞和nk细胞的质膜,其中在单核细胞中观察到更强的信号。因此,在单核细胞和nk细胞的质膜上检测到btn2a1而不是btn2a2,这为筛选识别btn2a1的mab提供了理由,因为它们能够调节这些免疫细胞区室的免疫功能。
[0418]
筛选抗btn2a mab调节单核细胞至巨噬细胞极化的能力
[0419]
为响应来自其微环境的信号,单核细胞可以极化为m1或m2巨噬细胞。m1巨噬细胞具有促炎和抗肿瘤特性,而m2巨噬细胞具有抗炎特性并与肿瘤发展有关。鉴于在单核细胞的质膜上仅发现btn2a1亚型,在m
‑
csf存在下评估仅识别btn2a1或识别btn2a1/btn2a2亚型的抗btn2a mab在体外干扰单核细胞极化为m2巨噬细胞的能力。在gm
‑
csf(cd14 /
‑
cd163
‑
)存在下产生的m1巨噬细胞和在m
‑
csf存在下产生的m2(cd14 cd163 )巨噬细胞(两者均没有mab)用作巨噬细胞极化的对照。体外极化5天后,在抗btn2a mab或其对照igg1存在下极化
的m1、m2和m
‑
csf诱导的巨噬细胞质膜上cd14和cd163的表达通过流式细胞术评估(表4)。如所预期,m1细胞呈现出低cd14表达和无法检测到的cd163表达(表5和图4),而m2巨噬细胞呈现出两种标志物的高表达。有趣的是,从现在起将被称为101g5的抗btn2a mab1在m
‑
csf存在下诱导了cd14和cd163表达的最强降低,从而使m
‑
csf诱导的巨噬细胞向m1样表型极化(表4和图4b)。m
‑
csf诱导的m2巨噬细胞极化的第二个最佳抑制剂是mab2,即107g3(表5和图4b)。这与在m
‑
csf和对照igg1存在下获得的巨噬细胞的表型相反,后者类似于未处理的m2巨噬细胞。
[0420]
表5:抗btn2a mab对单核细胞向m2
‑
巨噬细胞极化后cd14和cd163表达的影响。
[0421]
[0422][0423]
图5显示了在cd14和cd163表达抑制方面与同种型对照相比参考101g5和107g3抗btn2a mab的m2抑制作用的剂量依赖性(图5a和5b)以及作为m1表型的特征的pdl1和cd86的增加的表达(图5c和5d)。细胞因子分泌特征也是m2与m1巨噬细胞之间的区别特征。因此,在lps刺激来自m
‑
csf诱导的巨噬细胞的培养物上清液后(有或没有参考抗
‑
btn2a mab),通过elisa评估il
‑
10(抗炎,m2相关)和tnfα(促炎,m1相关)分泌。如图5e和5f所示,与同种型对照相比,参考抗btn2a mab以剂量依赖性方式抑制il
‑
10的分泌并增加tnfα的分泌。这些观察结果证实,在表型和细胞因子分泌方面,101g5和107g3通过偏向m1样表型来抑制m
‑
csf诱导的单核细胞极化为m2巨噬细胞。此外,101g5和107g3的这些作用是剂量依赖性的。每个mab的ic
50
和ec
50
显示于表6中。值得注意的是,与107g3相比,101g5获得了除pd
‑
l1之外的所有参数的最低ic
50
和ec
50
。
[0424]
表6:参考抗btn2a mab对m2与m1相关表型和细胞因子分泌的ic
50
和ec
50
[0425][0426]
来自肿瘤微环境的其他刺激已被描述为诱导m2巨噬细胞极化(mosser和edwards,nat rev immunol 2008;mantovani和allavena,j exp med 2015)。除了m
‑
csf,最常用的诱导m2极化的刺激之一是il
‑
4。我们确定了101g5和107g3对用m
‑
csf和il
‑
4刺激后由单核细胞产生的所谓的促肿瘤“m2 il
‑
4”巨噬细胞分化的影响。在这种条件下培养5天后,101g5和107g3抑制“m2 il
‑
4”相关标志物(cd14、cd163和dc
‑
sign,图6a、6b和6d)的表达和il
‑
10分泌(图6f),同时增加m1相关标志物(cd86、pdl1)的表达和tnfα的分泌(图6c、6e和6g)。因此,在促肿瘤环境(m
‑
csf和il
‑
4)中,101g4和107g3抑制“m2 il
‑
4”分化并增强促炎性m1巨噬细胞分化。
[0427]
此外,101g5和107g3对癌细胞诱导的单核细胞m2极化的影响通过在panc
‑
1(胰腺腺癌细胞系)条件培养上清液存在下培养分选的单核细胞来评估。当在此设置中添加101g5或107g3时,m2极化被抑制,如m2相关标志物(cd14、cd163)的表达和il
‑
10分泌(图7a
‑
c)减少以及m1相关促炎性tnfα的表达增加(图7d)所示。
[0428]
参考抗btn2a mab 101g5和107g3对m2
‑
巨噬细胞向m1重编程的影响
[0429]
评估了参考101g5和107g3 mab将m2极化巨噬细胞逆转为m1表型的潜力。为此目的,将先前在m
‑
csf存在下极化5天的m2巨噬细胞接种到先前包被有101g5和107g3 mab的孔中,并再培养2或4天。用ifnγ处理m2巨噬细胞作为m2
‑
>m1逆转的阳性对照。如图8所示,在101g5和107g3存在下培养的m2巨噬细胞获得了m1样表型,类似于ifnγ处理。实际上,用参考101g5和107g3 mab处理m2巨噬细胞导致cd14(图8a)和cd163(图8b)表达降低,并且cd86表达增加(图8c)。分别用101g5或107g3处理后,观察到pdl1适度至无上调(图8d)。此外,用101g5和107g3处理m2巨噬细胞可抑制il
‑
10分泌(图8e)并增强tnfα分泌(图8f),指示m1表型。
[0430]
图8g
‑
i显示在cd163表达抑制方面与同种型对照相比(图8g),参考101g5和107g3 mab对m2
‑
巨噬细胞重编程为m1巨噬细胞的影响的剂量依赖性,il
‑
10降低和tnfα分泌增加(图8h和i)。对于相关的特定m1/m2标志物,每种mab的ic
50
和ec
50
显示于表7中,其中抗btn2a 101g5 mab对m2
‑
巨噬细胞重编程为m1巨噬细胞显示出最佳活性。
[0431]
表7:参考抗btn2a mab对m2逆转的ic
50
和ec
50
[0432][0433][0434]
参考抗btn2a mab 101g5和107g3释放m2介导的t细胞增殖抑制
[0435]
研究了参考101g5和107g3 mab影响m2巨噬细胞功能的能力,即抑制m2介导的t细胞增殖。为此,将同种异体预活化的cd3 t细胞与m1、m2或在mab存在下产生的m2共培养。如所预期,与m1巨噬细胞相比,与常规m2巨噬细胞共培养导致cd3 t细胞数减少,t细胞增殖(通过ctv稀释评估)和ifnγ产生减少(图9a、c、g、e和i)。相比之下,在101g5和107g3存在下产生的m2巨噬细胞似乎没有抑制t细胞增殖,如在对照igg1存在下产生的m2巨噬细胞培养物相比,cd3 t细胞的百分比和绝对数更高(图9b、9h和9j)所示。此外,与对照igg1相比,在含有在101g5和107g3存在下产生的巨噬细胞的共培养物中,产生ifnγ的t细胞的百分比和数量也更高(图9d和9f)。
[0436]
因此,与单独由m
‑
csf诱导的m2巨噬细胞相比,除了m
‑
csf外,在101g5或107g3存在下产生的巨噬细胞允许同种异体cd3 t细胞的增殖和th1功能(ifnγ产生),类似于m1巨噬细胞。
[0437]
参考抗btn2a mab 101g5和107g3触发nk细胞活化和细胞毒性
[0438]
由于在nk细胞的质膜上发现了btn2a1,因此研究了101g5和107g3调节nk细胞活化的潜在能力。来自健康供体的纯化nk细胞在101g5或107g3存在下培养5天,有或没有进一步激活(分别为il
‑
2和il
‑
15或仅仅il
‑
2)。如图10所示,101g5和107g3在所有测试条件下都增强nk细胞质膜上cd69的表达(图10a)。此外,101g5和107g3还增强了由il
‑
2和il
‑
15处理nk细胞诱导的cd25的表达(图10b)。由于101g5和107g3能够激活纯化的nk细胞,我们研究了这些mab是否也能增强nk细胞的细胞毒性。因此,在存在101g5或107g3,有或没有il
‑
2和il
‑
15刺激的情况下评估了针对癌细胞系hl
‑
60(骨髓性白血病)、ht
‑
29(结肠癌)、mda
‑
mb
‑
231(乳腺癌)和a549(肺腺癌)的nk细胞脱粒(%cd107 细胞)。如所预期,在存在对照igg1的情况下,只有hl
‑
60细胞触发nk细胞脱粒(图10c),这通过用il
‑
2和il
‑
15刺激而增强(图10d)。在存在对照igg1和il2 il
‑
15的情况下,也观察到针对实体瘤细胞系ht
‑
29、mda
‑
mb
‑
231和a549的适度nk细胞脱粒。表8总结了针对这些和测试的其他(raji,hct116,du
‑
145)癌细胞系的nk细胞脱粒。有趣的是,参考mab 101g5和107g3增强了针对实体瘤细胞系mda
‑
mb
‑
231和a549的nk细胞脱粒,并且在没有il
‑
2 il
‑
15刺激的情况下,对ht
‑
29的作用较小。添加il
‑
2和il
‑
15增强101g5和107g3在mda
‑
mb
‑
231、a549和du145细胞中的作用(表8)。通过添加参考101g5或107g3 mab,在hl
‑
60和raji血癌细胞系中没有观察到这种增强(表8和图10c和
d)。此外,在共培养前与nk细胞预孵育时,参考mab 101g5和107g3能够触发针对a549细胞的nk细胞脱粒,而无需进一步将mab添加至共培养(图10e)。这表明参考107g3和101g5 mab通过直接与nk细胞结合引发针对癌细胞的细胞毒性的直接影响。此外,评估了101g5和107g3对增强nk细胞脱粒针对抗前列腺腺癌du
‑
145细胞系的剂量依赖性。实际上,101g5和107g3以剂量依赖性方式增强nk细胞针对du
‑
145细胞的脱粒作用(分别对于101g5和107g3,ec
50(无刺激)
=0.14和0.54nm;ec
50(il
‑
2 il
‑
15)
=0.08和0.2nm)。
[0439]
表8:在没有il
‑
2和il
‑
15刺激的情况下,针对不同癌细胞系的nk细胞脱粒(%cd107 细胞)
[0440][0441][0442]
最后,我们通过评估纯化nk细胞与白血病细胞系hl
‑
60或肺腺癌细胞系a459共培养后caspase 3/7细胞的百分比,测试了101g5和107g3增强nk细胞介导的癌细胞杀伤的能力。如图11所示,101g5和107g3增强nk细胞介导的对腺癌a549细胞的杀伤作用(~2倍),但对hl
‑
60白血病细胞没有作用。总之,这些观察结果表明101g5和107g3优先增强nk细胞针对来自实体瘤的癌细胞的细胞毒性。
[0443]
参考抗btn2a 101g5和107g3 mab识别btn2a1的不同表位
[0444]
101g5和107g3均结合btn2a1并共享抑制m2巨噬细胞极化和增强nk细胞活化和细胞毒性的能力。因此,我们研究了这些mab是否识别btn2a蛋白上的相同表位区域。因此,进行了基于octet的分箱实验,其中101g5和107g3使用“串联”设置竞争btn2a1结合。如图12所示,101g5和107g3没有阻断彼此与btn2a1的结合,表明这两种mab不与btn2a1上的相同表位区域结合。
[0445]
参考mab 101g5和107g3的表位定位
[0446]
为了表征btn2a1,我们将样品提交给胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶水解,然后进行nlc
‑
ltq
‑
orbitrap ms/ms分析。胰蛋白酶水解后,在btn2a1序列中鉴定出32个肽,覆盖序列的79.84%;胰凝乳蛋白酶水解后鉴定出27个肽,覆盖btn2a1序列的94.76%;asp
‑
n蛋白水解后鉴定出2个肽,覆盖btn2a1序列的12.50%;弹性蛋白酶水解后鉴定出33个肽,覆盖btn2a1序列的89.11%;嗜热菌蛋白酶水解后鉴定出29个肽,覆盖btn2a1序列的78.23%。基于获得的结果,设计了胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶肽的重叠定位(图13)。结合胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶蛋白水解的肽,覆盖了btn2a1序列的96.37%。为了以高分辨率确定btn2a1/107g3和btn2a1/101g5复合物的表位,将蛋白质复合物与氘化交联剂一起孵育,然后进行多酶裂解。在蛋白质复合物btn2a1/107g3的胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶蛋白水解后,nlc
‑
orbitrap ms/ms分析检测到btn2a1和抗体107g3之间的17个交联肽。
[0447]
表9:btn2a1/107g3之间交联的序列和位置
[0448]
[0449]
[0450][0451]
因此,我们的分析表明btn2a1和107g3 mab之间的相互作用包括btn2a1上的以下氨基酸:65、68、69、72、78、84、85、95、97、100。这些结果在图14a和图15中说明。
[0452]
在蛋白质复合物btn2a1/101g5的胰蛋白酶、胰凝乳蛋白酶、asp
‑
n、弹性蛋白酶和嗜热菌蛋白酶蛋白水解后,nlc
‑
orbitrap ms/ms分析检测到btn2a1和抗体101g5之间的14个交联肽。
[0453]
表10:btn2a1/101g5之间交联的序列和位置
[0454]
[0455][0456]
因此,我们的分析表明btn2a1和101g5之间的相互作用包括btn2a1上的以下氨基酸:212、213、218、220、224、229。这些结果如图14b和图16所示。
[0457]
参考抗btn2a 101g5和107g3 mab与食蟹猴btn2a1直系同源物存在交叉反应性
[0458]
btn2a1直系同源物存在于大多数非人灵长类动物中,包括食蟹猴(macaca fascicularis)。为了确定参考mabs 101g5和107g3与食蟹猴btn2a1直系同源物(cynobtn2a1;ncbi ref.xm_015448906.1,与人btn2a1具有93.31%的同一性)的交叉反应性,我们产生了含有cynobtn2a1胞外域的重组fc融合蛋白(cynobtn2a1
‑
fc),并且我们进行了elisa测定以评估参考mab与该蛋白质的结合。我们还使用重组人btn2a1
‑
fc蛋白进行了elisa,以比较人和食蟹猴btn2a1直系同源物之间参考mab的亲和力。如图13所示,101g5和107g3均能够结合cynobtn2a1胞外域,ec
50
分别为0.60和0.57nm,这与在hubtn2a1上获得的相应ec
50
(分别为0.82和0.56nm)相当。
[0459]
表11:用于实践的有用氨基酸和核苷酸序列的简要说明
[0460][0461]
本发明:
[0462]
[0463]
[0464][0465]
参考文献:
[0466]
在本技术中,各种参考文献描述了与本发明相关的现有技术。这些参考文献的公开在此通过引用并入本公开。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。