与油脂代谢调控相关的大豆rna结合蛋白gmtsn990及其编码基因与应用
技术领域
1.本发明涉及生物技术领域中,与油脂代谢调控相关的大豆rna结合蛋白gmtsn990及其编码基因与应用。
背景技术:
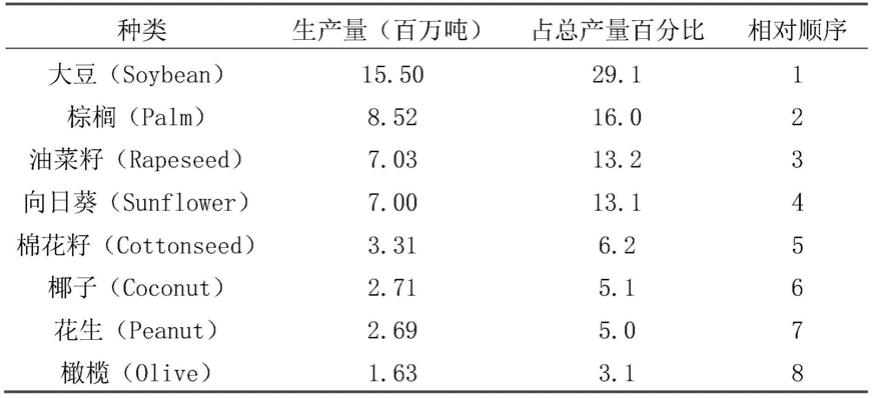
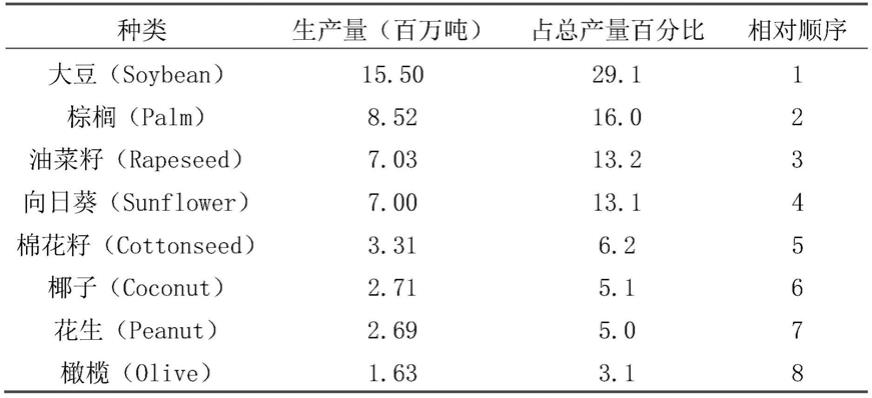
2.人类饮食中71%的油脂来自于植物。在世界上的几种主要产油作物中,大豆总产油量约占30%,居世界植物油产量的第一位(表1)。
3.表1世界上主要的产油作物
[0004][0005]
脂肪酸的合成是植物体中最重要的代谢途径之一,它存在于植物体的任何一个细胞中,是生长发育所必须的。对它的阻断会导致细胞的死亡,因而至今为止还没有发现一个阻断脂肪酸合成的植物突变体。
[0006]
大多数植物中,油脂都以三酰甘油(triacylglycerols,tag)的形式储藏,它的含量是一个非常重要的农艺性状,tag的生物合成称之为kennedy途径,如同真核生物中合成膜甘油酯的途径,脂肪酸去除coa后被转移到3-磷酸甘油的1和2位,形成中间产物pa。pa去磷酸化产生dag。在tag合成的最后一步,第三个脂肪酸分子被转移到空的dag 3
’-
oh位置,这一步反应是由二酰甘油乙酰转移酶(diacylglycerol acyltransferase,dgat)催化的,此反应被认为是tag生物合成中唯一的限速步骤。人们已对脂类合成途径有了认知,并且已经克隆了很多参与脂类合成的酶基因。然而,植物中,对脂类合成的调控机理及其相关基因仍然知之甚少。
[0007]
tudor-sn(tsn)rna结合蛋白,在动物细胞中为转录共激活因子,植物中其参与rna代谢过程、胞内对热反应、rna的基因沉默、mrna代谢和加工等过程调控生长发育的各个阶段。目前尚未见报道此类蛋白参与脂肪酸合成。
技术实现要素:
[0008]
本发明所要解决的技术问题是如何提高植物种子油脂含量。
[0009]
为解决上述技术问题,本发明首先提供了蛋白质或调控所述蛋白质活性或含量的物质的下述任一应用:
[0010]
d1)调控植物油脂含量;
[0011]
d2)制备调控植物油脂含量产品;
[0012]
d3)培育油脂含量增加植物;
[0013]
d4)制备培育油脂含量增加植物产品;
[0014]
d5)植物育种;
[0015]
所述蛋白质来源于大豆,其名称为gmtsn990,为如下a1)、a2)或a3):
[0016]
a1)氨基酸序列是序列2的蛋白质;
[0017]
a2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的蛋白质;
[0018]
a3)在a1)或a2)的n端或/和c端连接标签得到的融合蛋白质。
[0019]
为了使a1)中的蛋白质便于纯化,可在由序列表中序列2所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接上如下表所示的标签。
[0020]
表:标签的序列
[0021]
标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tag ii8wshpqfekc-myc10eqkliseedl
[0022]
上述a2)中的gmtsn990蛋白质,为与序列2所示蛋白质的氨基酸序列具有75%或75%以上同一性且具有相同功能的蛋白质。所述具有75%或75%以上同一性为具有75%、具有80%、具有85%、具有90%、具有95%、具有96%、具有97%、具有98%或具有99%的同一性。
[0023]
上述a2)中的gmtsn990蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
[0024]
上述a2)中的gmtsn990蛋白质的编码基因可通过将序列1所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5
′
端和/或3
′
端连上上表所示的标签的编码序列得到。其中,序列1所示的dna分子编码序列2所示的gmtsn990蛋白质。
[0025]
本发明还提供了与gmtsn990相关的生物材料的下述任一应用:
[0026]
d1)调控植物油脂含量;
[0027]
d2)制备调控植物油脂含量产品;
[0028]
d3)培育油脂含量增加植物;
[0029]
d4)制备培育油脂含量增加植物产品;
[0030]
d5)植物育种;
[0031]
所述生物材料为下述b1)至b7)中的任一种:
[0032]
b1)编码gmtsn990的核酸分子;
[0033]
b2)含有b1)所述核酸分子的表达盒;
[0034]
b3)含有b1)所述核酸分子的重组载体、或含有b2)所述表达盒的重组载体;
[0035]
b4)含有b1)所述核酸分子的重组微生物、或含有b2)所述表达盒的重组微生物、或含有b3)所述重组载体的重组微生物;
[0036]
b5)含有b1)所述核酸分子的转基因植物细胞系、或含有b2)所述表达盒的转基因植物细胞系;
[0037]
b6)含有b1)所述核酸分子的转基因植物组织、或含有b2)所述表达盒的转基因植物组织;
[0038]
b7)含有b1)所述核酸分子的转基因植物器官、或含有b2)所述表达盒的转基因植物器官。
[0039]
上述应用中,b1)所述核酸分子可为如下b11)或b12)或b13)或b14):
[0040]
b11)编码序列是序列表中序列1的cdna分子或dna分子;
[0041]
b12)序列表中序列1的cdna分子或dna分子;
[0042]
b13)与b11)或b12)限定的核苷酸序列具有75%或75%以上同一性,且编码gmtsn990的cdna分子或dna分子;
[0043]
b14)在严格条件下与b11)或b12)或b13)限定的核苷酸序列杂交,且编码gmtsn990的cdna分子或dna分子。
[0044]
其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
[0045]
本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码gmtsn990蛋白质的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的gmtsn990蛋白质的核苷酸序列75%或者更高同一性的核苷酸,只要编码gmtsn990蛋白质且具有gmtsn990蛋白质功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
[0046]
这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列2所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。
[0047]
上述应用中,所述严格条件可为如下:50℃,在7%十二烷基硫酸钠(sds)、0.5m napo4和1mm edta的混合溶液中杂交,在50℃,2
×
ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5m napo4和1mm edta的混合溶液中杂交,在50℃,1
×
ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5m napo4和1mm edta的混合溶液中杂交,在50℃,0.5
×
ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5m napo4和1mm edta的混合溶液中杂交,在50℃,0.1
×
ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5m napo4和1mm edta的混合溶液中杂交,在65℃,0.1
×
ssc,0.1%sds中漂洗;也可为:在6
×
ssc,0.5%sds的溶液中,在65℃下杂交,然后用2
×
ssc,0.1%sds和1
×
ssc,0.1%sds各洗膜一次;也可为:2
×
ssc,0.1%sds
的溶液中,在68℃下杂交并洗膜2次,每次5min,又于0.5
×
ssc,0.1%sds的溶液中,在68℃下杂交并洗膜2次,每次15min;也可为:0.1
×
sspe(或0.1
×
ssc)、0.1%sds的溶液中,65℃条件下杂交并洗膜。
[0048]
上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。
[0049]
上述应用中,b2)所述的含有编码gmtsn990蛋白质的核酸分子的表达盒(gmtsn990基因表达盒),是指能够在宿主细胞中表达gmtsn990蛋白质的dna,该dna不但可包括启动gmtsn990基因转录的启动子,还可包括终止gmtsn990基因转录的终止子。进一步,所述表达盒还可包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子,组织、器官和发育特异的启动子,和诱导型启动子。启动子的例子包括但不限于:花椰菜花叶病毒的组成型启动子35s;来自西红柿的创伤诱导型启动子,亮氨酸氨基肽酶("lap",chao等人(1999)plant physiol 120:979-992);来自烟草的化学诱导型启动子,发病机理相关1(pr1)(由水杨酸和bth(苯并噻二唑-7-硫代羟酸s-甲酯)诱导);西红柿蛋白酶抑制剂ii启动子(pin2)或lap启动子(均可用茉莉酮酸甲酯诱导);热休克启动子(美国专利5,187,267);四环素诱导型启动子(美国专利5,057,422);种子特异性启动子,如谷子种子特异性启动子pf128(cn101063139b(中国专利200710099169.7)),种子贮存蛋白质特异的启动子(例如,菜豆球蛋白、napin,oleosin和大豆beta conglycin的启动子(beachy等人(1985)embo j.4:3047-3053))。它们可单独使用或与其它的植物启动子结合使用。此处引用的所有参考文献均全文引用。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv 35s终止子、tml终止子、豌豆rbcs e9终止子和胭脂氨酸和章鱼氨酸合酶终止子(参见,例如:odell等人(i
985
)nature 313:810;rosenberg等人(1987)gene,56:125;guerineau等人(1991)mol.gen.genet,262:141;proudfoot(1991)cell,64:671;sanfacon等人genes dev.,5:141;mogen等人(1990)plant cell,2:1261;munroe等人(1990)gene,91:151;ballad等人(1989)nucleic acids res.17:7891;joshi等人(1987)nucleic acid res.,15:9627)。
[0050]
可用现有的表达载体构建含有所述gmtsn990基因表达盒的重组载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。如pahc25、pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa、psn1301或pcambia1391-xb(cambia公司)等。所述植物表达载体还可包含外源基因的3
′
端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3
′
端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂碱合成酶基因nos)、植物基因(如大豆贮存蛋白基因)3
′
端转录的非翻译区均具有类似功能。使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、抗生素的标记基因(如赋予对卡那霉素和相关抗生素抗性的nptii基因,赋予对除草剂膦丝菌素抗性的bar基因,赋予对抗生素潮霉素抗性的hph基因,和赋予对氨甲喋呤抗性的
dhfr基因,赋予对草甘磷抗性的epsps基因)或是抗化学试剂标记基因等(如抗除莠剂基因)、提供代谢甘露糖能力的甘露糖-6-磷酸异构酶基因。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
[0051]
上述应用中,所述载体可为质粒、黏粒、噬菌体或病毒载体。所述质粒具体可为pbin438载体。
[0052]
b3)所述重组载体具体可为pbin438-gmtsn990。所述pbin438-gmtsn990为在pbin438中插入为序列表中序列1所示的gmtsn990基因得到的重组载体,该重组载体表达gmtsn990,且gmtsn990基因的表达由35s驱动。
[0053]
上述应用中,所述微生物可为酵母、细菌、藻或真菌。其中,细菌可为农杆菌,如农杆菌gv3101。
[0054]
上述应用中,所述转基因植物细胞系、转基因植物组织和转基因植物器官均不包括繁殖材料。
[0055]
上述应用中,所述植物可为m1)或m2)或m3):
[0056]
m1)双子叶植物或单子叶植物;
[0057]
m2)豆科植物;
[0058]
m3)大豆。
[0059]
本发明还提供了下述任一方法:
[0060]
x1)培育油脂含量增加植物的方法,包括使受体植物中表达gmtsn990,或提高受体植物中gmtsn990的含量,或提高受体植物中gmtsn990的活性,得到油脂含量增加的目的植物;
[0061]
x2)提高植物油脂含量的方法,包括使受体植物中表达gmtsn990,或提高受体植物中gmtsn990的含量,或提高受体植物中gmtsn990的活性,得到油脂含量增加的目的植物,实现植物油脂含量的提高。
[0062]
上述方法中,x1)和x2)中提高受体植物中gmtsn990的含量可通过向所述受体植物导入gmtsn990的编码基因并使所述编码基因得到表达实现。
[0063]
上述方法中,所述编码基因可为b1)所述核酸分子。
[0064]
上述方法中,其中所述gmtsn990的编码基因可先进行如下修饰,再导入受体植物中,以达到更好的表达效果:
[0065]
1)根据实际需要进行修饰和优化,以使基因高效表达;例如,可根据受体植物所偏爱的密码子,在保持本发明所述gmtsn990的编码基因的氨基酸序列的同时改变其密码子以符合植物偏爱性;优化过程中,最好能使优化后的编码序列中保持一定的gc含量,以最好地实现植物中导入基因的高水平表达,其中gc含量可为35%、多于45%、多于50%或多于约60%;
[0066]
2)修饰邻近起始甲硫氨酸的基因序列,以使翻译有效起始;例如,利用在植物中已知的有效的序列进行修饰;
[0067]
3)与各种植物表达的启动子连接,以利于其在植物中的表达;所述启动子可包括组成型、诱导型、时序调节、发育调节、化学调节、组织优选和组织特异性启动子;启动子的选择将随着表达时间和空间需要而变化,而且也取决于靶物种;例如组织或器官的特异性表达启动子,根据需要受体在发育的什么时期而定;尽管证明了来源于双子叶植物的许多
启动子在单子叶植物中是可起作用的,反之亦然,但是理想地,选择双子叶植物启动子用于双子叶植物中的表达,单子叶植物的启动子用于单子叶植物中的表达;
[0068]
4)与适合的转录终止子连接,也可以提高本发明基因的表达效率;例如来源于camv的tml,来源于rbcs的e9;任何已知在植物中起作用的可得到的终止子都可以与本发明基因进行连接;
[0069]
5)引入增强子序列,如内含子序列(例如来源于adhl和bronzel)和病毒前导序列(例如来源于tmv,mcmv和amv)。
[0070]
所述gmtsn990的编码基因可利用含有所述gmtsn990的编码基因的重组载体导入受体植物。所述重组载体具体可为所述pcambia1301-gmtsn990。
[0071]
所述重组载体可通过使用ti质粒,ri质粒,植物病毒载体,直接dna转化,显微注射,电穿孔,农杆菌介导等常规生物方法导入植物细胞或组织,并将转化的植物组织培育成植株。被转化的植物宿主既可以是单子叶植物,也可以是双子叶植物。
[0072]
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以干旱处理筛选转化植株。
[0073]
所述目的植物理解为不仅包含gmtsn990蛋白或其编码基因被改变的第一代植物,也包括其子代。对于所述目的植物,可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述目的植物包括种子、愈伤组织、完整植株和细胞。
[0074]
上述方法中,所述受体植物可为m1)或m2)或m3):
[0075]
m1)双子叶植物或单子叶植物;
[0076]
m2)豆科植物;
[0077]
m3)大豆。
[0078]
本发明还提供了提高植物油脂含量产品,所述产品含有gmtsn990或所述生物材料。
[0079]
所述产品可以gmtsn990或所述生物材料为其活性成分,还可将gmtsn990或所述生物材料与具有相同功能的物质组合在一起作为其活性成分。
[0080]
上述产品中,所述植物可为m1)或m2)或m3):
[0081]
m1)双子叶植物或单子叶植物;
[0082]
m2)豆科植物;
[0083]
m3)大豆。
[0084]
gmtsn990或所述生物材料,也属于本发明的保护范围。
[0085]
本发明中,所述油脂含量可为种子中油脂含量。所述油脂可为亚油酸、油酸、亚麻酸、棕榈酸、硬脂酸、贡多酸、花生四烯酸和/或芥子酸。
[0086]
本发明的实验证明,本发明的与植物组织油脂含量相关的rna结合蛋白gmtsn990及其编码基因,在植物中转入表达该转基因可以提高种子中的油脂含量。说明rna结合蛋白gmtsn990及其编码基因可以正调控植物种子中油脂含量。该基因对提高种子油脂含量,培
育高油脂品种具有重要的理论和现实意义。
附图说明
[0087]
图1为gmtsn990基因在大豆不同器官的表达分析。
[0088]
图2为植物表达载体pbin438-gmtsn990示意图。
[0089]
图3为gmtsn990相对表达量检测结果。对照表示野生型拟南芥。
[0090]
图4为gmtsn990转基因株系总油脂含量检测结果。对照表示野生型拟南芥。
具体实施方式
[0091]
下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本技术领域普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。
[0092]
下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂、仪器等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中,如无特殊说明,序列表中各核苷酸序列的第1位均为相应dna/rna的5
′
末端核苷酸,末位均为相应dna/rna的3
′
末端核苷酸。
[0093]
大豆材料:大豆williams 82(w82)记载在如下文献中:scott a jackson等,genome sequence of the palaeopolyploid soybean,nature,2010,vol.463,178-183,该大豆材料为美国栽培品种,由purdue university,department of agronomy,scott jackson教授赠送,经urdue university,department of agronomy,scott jackson教授同意后公众可从中国科学院遗传与发育生物学研究所获得。
[0094]
pbin438载体(双元表达载体)由方荣祥院士惠赐,记载于“李太元,田颖川,秦晓峰,等.高效抗虫转基因烟草的研究[j].中国科学(b辑),1994,24(3):276-282.)”一文中,公众经方荣祥院士同意后可以从中国科学院遗传与发育生物学研究所获得。
[0095]
农杆菌gv3101菌株记载在“clough-sj,bent-af.floral dip:a simplified method for agrobacterium-mediated transformation of arabidopsis thaliana.plant-journal.1998,16:6,735-743”一文中,公众可从中国科学院遗传与发育生物学研究所获得。
[0096]
哥伦比亚生态型拟南芥(col-0):arabidopsis biological resource center(abrc),以下简称野生型拟南芥。
[0097]
实施例1、大豆油脂代谢调控相关蛋白gmtsn990编码基因gmtsn990的克隆和组织特异性表达检测
[0098]
本发明以种子脂肪含量有差异的大豆品种为材料,分析了种子发育中油脂快速积累的前期和中期的转录组,获得一个参与调控籽粒油脂含量的候选基因,根据williams 82(w82)基因组序列,设计引物从w82中克隆了目的基因,并检测了其在大豆不同器官中的表达水平。
[0099]
1、gmtsn990的克隆
[0100]
采用ctab提取w82种子总rna,将rna用逆转录酶反转录合成cdna。
[0101]
根据w82基因组序列中glyma.04g203000全长cdna序列的信息,设计引物,引物序列如下:
[0102]
glyma.04g203000-f:5
’-
atggcatcaacggcgagt-3’;
[0103]
glyma.04g203000-r:5
’-
ttatttacggccaccagcag-3’。
[0104]
以w82的cdna为模板,用glyma.04g203000-f和glyma.04g203000-r为引物,进行pcr扩增,得到约3.0kb的pcr产物。经过测序,该pcr产物为2973bp,具有序列表中序列1所示的核苷酸,经比对,该核苷酸序列包含tudor-sn结构域,为rna结合蛋白,将该基因编码的蛋白命名为gmtsn990,该蛋白包含990个氨基酸,其氨基酸序列为序列表中的序列2。
[0105]
2、gmtsn990在大豆不同器官中的表达分析
[0106]
取大豆w82根、苗、叶、荚和籽粒的总rna,用逆转录酶反转录合成cdna,进行real time-pcr鉴定,引物步骤1。以大豆tublin基因为内标,所用引物为primer-tf:5
’-
aacctcctcctcatcgtact3’,和primer-tr:5
’-
gacagcatcagccatgttca-3’。
[0107]
结果如图2所示,gmtsn990基因的转录在种子中表达量最高,依次是荚、叶、苗、和根。
[0108]
实施例2、gmtsn990基因可以调控拟南芥种子中油脂的合成和积累
[0109]
一、重组农杆菌的获得
[0110]
gmtsn990基因表达植物载体的构建:所用植物表达载体为pbin438,运用同源重组连接,将gmtsn990基因插入至该载体中。
[0111]
所用pbin438的同源臂序列为:
[0112]
f:5
’-
caattactgcagggatccgtcgac-3’;
[0113]
r:5
’-
cttatcgtcgtcatccttgtaatc-3’。
[0114]
具体的是利用pbin438的同源臂序列将序列1所示的gmtsn990基因替换pbin438的上述两条同源臂间的dna片段,将所得到的重组载体记为pbin438-gmtsn990,pbin438-gmtsn990的结构示意图如图2所示,pbin438-gmtsn990能表达序列2所示的gmtsn990蛋白质,其中,gmtsn990基因的表达由35s驱动。
[0115]
重组菌的构建:将上述含有gmtsn990基因的重组载体pbin438-gmtsn990导入农杆gv3101,得到重组农杆菌,将该重组农杆菌命名为gv3101/gmtsn990;将pbin438导入农杆gv3101,得到重组农杆菌,将该重组农杆菌命名为gv3101/pbin438,作为空载体对照。
[0116]
二、转gmtsn990拟南芥的获得及鉴定
[0117]
将重组农杆菌gv3101/gmtsn990培养至对数期,然后用抽真空法将其转化哥伦比亚生态型拟南芥(col-0)中。经培育后收获种子,将种子播于含卡那霉素(50mg/l)的ms筛选培养基上,待筛选得到的t1代植株长至4-6叶时移到蛭石上生长,收获t1代单株,各单株种子分别播种,用相同的ms筛选培养基继续筛选以观察t2代的分离情况,如此重复数代直至获得遗传稳定的转基因纯合株系,获得10个转gmtsn990拟南芥纯系。
[0118]
按照相同的转化方法,利用重组农杆菌gv3101/pbin438转化哥伦比亚生态型拟南芥(col-0),得到空载体对照株系。经检测,野生型和空载体对照株系全生育期的生长发育及其它性状均未见明显差异。因此下述对照为野生型拟南芥。
[0119]
提取上述各个株系苗rna,反转录得到cdna作为模板,利用引物(f:5
’-
atggcatcaacggcgagt-3’和r:5
’-
ttatttacggccaccagcag-3’)进行real time-pcr鉴定。以
野生型拟南芥(col-0)为对照。拟南芥atactin2基因为内标,所用引物为primer-tf:5
’-
atgcccagaagtcttgttcc-3’,和primer-tr:5
’-
tgctcatacggtcagcgata-3’。
[0120]
结果显示,oe1、oe9、oe3和oe13中gmtsn990基因的相对表达量分别约为0.126、0.053、0.045、0.036,作为对照的野生型拟南芥(col-0)中均未能检测出gmtsn990基因的表达量,图3。表明,gmtsn990成功基因转入拟南芥中,且得到了表达。选取纯系oe1、oe9、oe3和oe13做进一步检测。
[0121]
三、转gmtsn990基因拟南芥的表型分析
[0122]
测定gmtsn990转基因纯系oe1、oe9、oe3和oe13种子中总油脂含量,以野生型拟南芥为对照,每个株系取30株的籽粒,生物学实验重复三次,结果取平均值
±
标准差。
[0123]
步骤如下:
[0124]
彻底干燥待测籽粒,研磨成粉,取10mg加入螺口的2ml离心管中,每份样品平行称取四份;然后向样品中加入10μl的17:0十七烷酸(10mg/ml)做内标,而后再加入含2.5%浓硫酸的甲醇溶液1ml,85℃水浴中保温1小时,期间晃动数次;自然冷却后,取上清500μl到新管中,加入600μl的0.9%(质量百分比)nacl水溶液、300μl正己烷,震荡混匀几分钟,4000转离心10分钟,取上清至新管中;通风橱中过夜使正己烷挥发完全,然后加入50μl乙酸乙酯溶解甲酯化的脂肪酸;将甲酯化的脂肪酸样品用气相色谱质谱联用仪测各个组分的相对含量,然后将各个组分的脂肪酸与加入的内标比较得出绝对含量,而后计算总油脂含量,即各个组分绝对含量之和。(shen,b.,et al.,the homeobox gene glabra2 affects seed oil content in arabidopsis,plant mol.biol.,60,377-387,2006.)
[0125]
其中,气相色谱仪:日本岛津公司(gc-2014,shimadzu)。检测条件:色谱柱:famewax,载气:氮气,柱流量:4.21ml/min,柱温:170℃,进样温度:250℃,检测器温度:250℃,压力:163.8kpa,分析时间:25min,内标:十七烷酸(17:0,四川省维克奇生物科技有限公司,货号:506-12-7),进样体积:1μl,溶剂:乙酸乙酯。
[0126]
所检测的脂肪酸为亚油酸、油酸、亚麻酸、棕榈酸、硬脂酸、贡多酸、花生四烯酸和芥子酸。
[0127]
结果如图4所示,野生型拟南芥籽粒总油脂含量为29.5
±
0.3%,即为籽粒总重量的百分比。转gmtsn990拟南芥株系oe1、oe9、oe3和oe13种子总油脂质量含量分别为33.9
±
1.8、33.1
±
2.0、31.8和35.1
±
0.3%。结果表明,4个转基因株系种子中的油脂含量均显著高于野生型对照。
[0128]
上述实验表明,大豆rna结合蛋白gmtsn990可以正调控种子中油脂的合成和积累,其编码基因gmtsn990的表达,可提高转基因植株种子中总油脂的含量。
[0129]
以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本技术欲包括任何变更、用途或对本发明的改进,包括脱离了本技术中已公开范围,而用本领域已知的常规技术进行的改变。按以下附带的权利要求的范围,可以进行一些基本特征的应用。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。