1.本发明涉及微藻类培养方法,更详细地涉及在可实现光合作用的时间利用自养方式,且在无法实现光合作用的时间利用异养方式,如此分离其培养方式连续培养的微藻类培养方法。
背景技术:
2.为了确保二氧化碳的减排及利用(ccu)及经济性,重要的是微藻类生物工艺的商业化。微藻类培养方式中的自养方式大大依赖于大气中以0.04%的低浓度存在的co2的浓度。在高浓度的co2浓度中,顺畅地传递微藻类生长所需的物质,因而可供给高浓度的co2,并可满足co2供给的经济性,从发电站中得到的废气利用于微藻类的大规模培养。
3.为了微藻类生物工艺的商业化,持续进行用于提高生物质的生产率的战略和研究。在现有的自养方式下,即使持续供给光和co2,在规定光周期之后光合作用效率也降低,因而增加生物质生产率时受限。为了弥补这种缺点而提出的混养方式和异养方式还可利用有机碳源提高生物质生产率。其中,混养方式为自养方式和异养方式在一个培养过程内一同发生的营养方式。在混养方式中,同时发生通过光合作用使无机碳源进行固碳而利用为化学能的自养方式和从有机碳源中通过呼吸得到细胞活动所需的能量的异养方式,因而可得到高于各个培养方式的生物质生产率。但是,在这种混养方式中,虽然生物质生产率大,但存在抑制高浓度有机碳的吸收,光合作用效率减少导致的叶绿素损失以及有机碳源的供给费用问题之类的需要解决的主要问题。
技术实现要素:
4.本发明用于解决上述的现有技术的问题,本发明的一实施方式提供在光周期利用自养方式且在暗周期利用异养方式,如此按不同周期分离培养方式来培养微藻类的方法。
5.本发明的微藻类培养方法包括:步骤(a),在供给光的规定的第一时间内,以自养方式在培养基中培养微藻类;以及步骤(b),在中断供给上述光的规定的第二时间内,向上述培养基供给有机碳源,以异养方式培养上述微藻类。
6.并且,在本发明的微藻类培养方法中,可依次反复上述步骤(a)及上述步骤(b)。
7.并且,在本发明的微藻类培养方法中,上述步骤(a)及上述步骤(b)可连续进行24小时,上述第一时间可以为15~17小时。
8.并且,在本发明的微藻类培养方法中,在上述步骤(a)中,可供给无机碳源。
9.并且,在本发明的微藻类培养方法中,上述有机碳源可选自由乙酸(acetic acid)、葡萄糖(glucose)及它们的混合物组成的组中。
10.并且,在本发明的微藻类培养方法中,上述乙酸的浓度可以为0.5~1.8g/l。
11.并且,在本发明的微藻类培养方法中,上述葡萄糖的浓度可以为0.15~0.7g/l。
12.并且,在本发明的微藻类培养方法中,上述微藻类可以为原壳小球藻(chlorella protothecoides)。
13.通过根据附图的以下详细说明,本发明的特征及优点变得更加明确。
14.在此之前,本说明书及权利要求书中使用的术语或词汇不应被解释为常规且词典上的含义,应立足于发明人为了以最佳的方法说明自己的发明而可适当地定义术语的概念的原则而仅被解释为符合本发明的技术思想的含义和概念。
15.根据本发明,在大规模培养中,在光周期(light cycle)可利用自养方式将细胞生长性最大化,在暗周期(dark cycle)可利用异养方式将细胞生长性最大化。相比于现有的自养方式,不对光合受体系统(photosynthetic receptor system)施加负荷,可得到更高的生物质收率,可使作为混养方式的优点的生物质生产率相比于现有的常规的混养方式得到提高,在混养过程中,可改善供给有机碳源导致的光合作用抑制和有机碳的转化效率抑制。
16.并且,在本发明中,由于在异养方式期间较低地维持有机碳源的浓度,因而无需添加用于阻止细菌等引起的污染的抗生素等追加的化学物质,故而在回收生物质时,无需追加的去除化学物质的工序,具有经济性,在暗周期利用乙酸、葡萄糖作为有机碳源,还可利用多种废水供给碳源,因而还可确保经济性。
附图说明
17.图1a为利用本发明的依次反复自养-异养方式(repeated sequential auto-and heterotrophic culture mode,rsah)的微藻类培养方法的示意图。
18.图1b为比较本发明的依次反复自养-异养方式(rsah)的细胞生长趋势和现有自养方式(autotrophy)及混养方式(mixotrophy)的图表。
19.图2a至图2f为供给乙酸作为实施例1的有机碳源,供给5%co2作为无机碳源时的自养方式、异养方式及混养方式的各个实验结果,图2a为生长曲线,图2b为以24小时间隔测定的比生长速率,图2c为有机碳源消耗量和生物质产量的对比图,图2d为以24小时间隔测定的atp/adp比率,图2e为在48小时时间点测定的碳酸酐酶比活度,图2f为在48小时时间点测定的核酮糖-1,5-二磷酸羧化酶(rubisco)比活度。
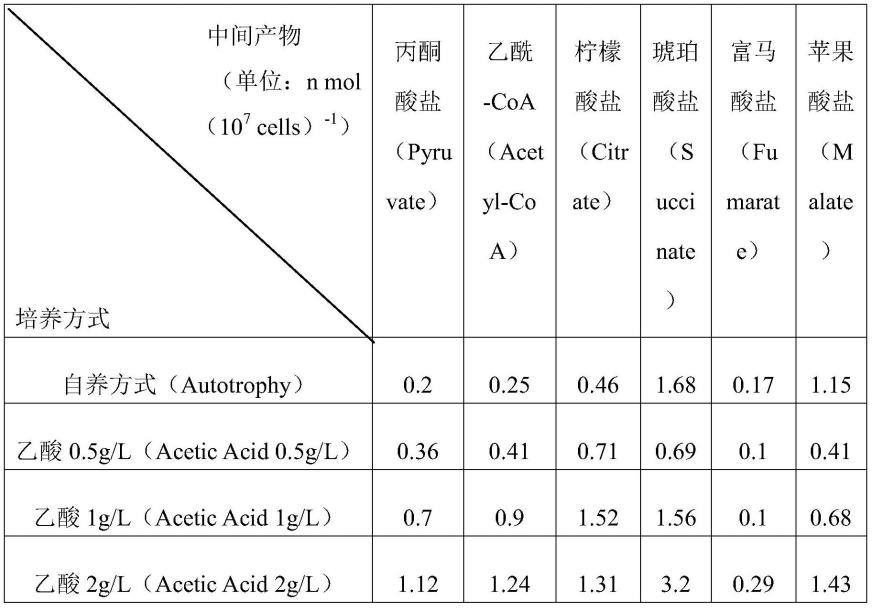
20.图3a表示实施例2的自养方式和将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式干菌量变化。
21.图3b表示实施例2的自养方式和将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式tca电路内中间产物的浓度(单位:n mol(107cells)-1
)。
22.图4a表示实施例3的自养方式和将葡萄糖用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式干菌量变化。
23.图4b表示实施例3的自养方式和将葡萄糖用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式tca电路内中间产物的浓度(单位:n mol(107cells)-1
)。
24.图5a表示实施例4的原壳小球藻的不同光周期/暗周期(24小时/0小时、16小时/8小时、14小时/10小时、12小时/12小时)的干重。
25.图5b表示在实施例4的依次反复自养-异养方式中将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的干菌量变化。
26.图5c表示在实施例4的依次反复自养-异养方式中将葡萄糖用作有机碳源时(0.167g/l、0.333g/l、0.667g/l)的干菌量变化(图下端所示的区间为0~16、24~40、48~
64、72~88、96~112小时的光周期及16~24、40~48、64~72、88~96、112~120小时的暗周期)。
27.图6a表示将乙酸用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的干菌量变化。
28.图6b表示将葡萄糖用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的干菌量变化(图下端所示的区间为0~16、24~40、48~64、72~88、96~112小时的光周期及16~24、40~48、64~72、88~96、112~120小时的暗周期)。
29.图6c表示将乙酸用作实施例5的有机碳源时的混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的有机碳源消耗量和比生长速率的对比值。
30.图6d表示将葡萄糖用作实施例5的有机碳源时的混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的有机碳源消耗量和比生长速率的对比值。
31.图6e表示将乙酸用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的atp/adp比。
32.图6f表示将葡萄糖用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的atp/adp比。
具体实施方式
33.通过与附图相关的以下详细说明和优选实施例,本发明的目的、特定的优点及新特征将变得更加明确。除非以其他方式定义,本说明书中使用的所有技术及科学术语具有与本发明所属技术领域的普通技术人员通常所理解的含义相同的含义。通常,本说明书中使用的命名法在本技术领域中熟知而常规使用。以下,在说明本发明的过程中,省略有可能不必要地混淆本发明的要旨的相关公知技术的详细说明。
34.图1a为利用本发明的依次反复自养-异养方式(repeated sequential auto-and heterotrophic culture mode,rsah)的微藻类培养方法的示意图,图1b为比较本发明的依次反复自养-异养方式(rsah)的细胞生长趋势和现有自养方式(autotrophy)及混养方式(mixotrophy)的图表。
35.如图1a所示,本发明的微藻类培养方法包括:在供给光的规定的第一时间内,以自养方式在培养基中培养微藻类的步骤;以及在中断供给光的规定的第二时间内,向培养基供给有机碳源,以异养方式培养微藻类的步骤。
36.微藻类培养中所使用的以往培养方式有自养方式(autotrophy)、异养方式(heterotrophy)、混养方式(mixotrophy)。本发明提出依次利用自养方式及异养方式的广义的混养方式。
37.其中,可连续进行自养方式的培养及异养方式的培养。即,在供给光的期间(上述
第一时间内)能够以自养方式培养微藻类,在中断供给光的期间(上述第一时间结束之后的上述第二时间内)能够以异养方式培养微藻类。
38.本发明的自养方式和异养方式可连续进行24小时。此时,进行自养方式的上述第一时间可以为15~17小时,进行异养方式的上述第二时间可以为7~9小时。例如,在可进行光合作用的白天能够以自养方式培养微藻类,在无法进行光合作用的夜间能够以异养方式培养微藻类。
39.并且,可依次反复本发明的自养方式和异养方式。即,可供给光的上述第一时间为光周期(light cycle),阻断光的供给的上述第二时间为暗周期(dark cycle),在光周期和暗周期依次反复的期间,在光周期能够以自养方式培养微藻类,在暗周期能够以异养方式培养微藻类。其中,光周期及暗周期以一天(day)为基准区分为白天及夜间,在微藻类的户外培养(outdoor culture)中可适用本发明。但是,本发明不一定仅局限于户外培养而适用,也可以人为供给光来组成光周期,中断光的供给来组成暗周期,以适用本发明。另一方面,光周期/暗周期优选为16小时/8小时。
40.混养相比于自养和异养,其生物质生产率优秀,但其生物质生产率始终与自养和异养之和不同。根据生长条件,在混养过程内自养和异养相互起作用,各个过程的生长被抑制。代表性的是,添加有机碳源影响微藻类的呼吸过程,相比于无机碳源,容易转化为能源,因而有机碳源的浓度变高时,光合作用的效率下降。并且,有机碳源的存在改变tca电路的活性,致使光合作用效率下降。作为利用于异养的有机碳源,主要使用乙酸盐(乙酸、乙酸钠),乙酸盐在同化过程中促进琥珀酸的生产,因而投入高浓度的乙酸盐时,细胞生长被抑制。并且,有机碳源的供给相比于利用废气供给的co2供给方式,存在消耗供给碳源所需的额外费用的问题。并且,在自养过程中,持续的光照射(16小时以上)对光合受体系统(photosynthetic receptor system)引起损伤,因而光合作用效率减少,细胞生长性也被限制。
41.为了克服这种以往混养方式的问题,在本发明中,按周期分离自养方式和异养方式,可将各个培养方式的效率最大化,而且设计按各个不同异养周期以低浓度添加有机碳源来将细胞生长最大化的周期性自养-异养方式。
42.在本发明的异养过程中供给的有机碳源可选自由乙酸(acetic acid)、葡萄糖(glucose)及它们的混合物组成的组中。其中,乙酸的浓度可以为0.5~1.8g/l,适当为0.8~1.2g/l,可更优选为1g/l。另一方面,葡萄糖的浓度可以为0.15~0.7g/l,适当为0.5~0.7g/l,可更优选为0.667g/l。为了在光周期维持完整的自养方式,将有机碳源的浓度设定为可在暗周期期间消耗所有有机碳源,以便于在不妨碍细胞生长性的情况下可最大限度地添加有机碳源。
43.在本发明的自养方式中,可利用tap-c培养基,在异养方式中可在tap-c培养基中将葡萄糖或乙酸利用为有机碳源。
44.在本发明的自养方式培养过程中,可供给无机碳源,该无机碳源可以为co2。
45.本发明可使用为培养原壳小球藻(chlorella protothecoides)的方法,但对象微藻类不受特别限制。
46.根据本发明,当在培养过程中分离自养方式和异养方式时,通常在不产生光合作用的暗周期区间仅进行异养方式,因而存在如下优点,在无光合受体系统的损伤的情况下
可增大生物质生产率,在混养方式中可阻止有机碳源引起的光合率的减少和生长抑制效果。
47.如图1b所示,在本发明的依次反复自养-异养方式(repeated sequential auto-and heterotrophic culture mode,rsah)中,在暗周期也顺畅地进行细胞生长,因而可期待高于现有的自养方式及混养方式的生物质生产率。
48.综合来说,在本发明中,利用依次反复自养-异养方式在供给光的周期适用自养方式的环境,在不供给光的周期适用供给有机碳源,维持低的浓度,以防止细胞生长被抑制的异养方式的环境。由此相比于常规的混养方式,不仅增加生物质生产率,还防止光合作用被抑制,由此可维持固碳比率(carbon fixation rate)使其适合于自养方式,并可保存二氧化碳减排效率。并且,通过本专利的培养方式在异养方式下通过维持低的有机碳源的浓度,可较高地维持从有机碳源到生物质的转化效率。进而无需供给高浓度的有机碳源,因而利用废水供给更低廉的费用的有机碳源,由此可确保经济性。
49.以下,通过实施例更详细说明本发明。
50.实施例1.自养方式、异养方式、混养方式对原壳小球藻(chlorella protothecoides)的细胞生长带来的影响
51.为了掌握各个培养方式(自养方式、异养方式、混养方式)对原壳小球藻(chlorella protothecoides)的细胞生长及物质代谢带来的影响而做实验。为此,在本实验中,将乙酸用作有机碳源,将co2用作无机碳源,在自养方式中作为碳源使用无机碳源,在异养方式中作为碳源使用有机碳源,在混养方式中作为碳源同时使用有机碳源和无机碳源,来培养原壳小球藻。并且,同等地维持相当于不同物理化学变量的温度、ph、培养时间、接种量等。
52.图2a至图2f为供给乙酸作为实施例1的有机碳源,供给5%co2作为无机碳源时的自养方式、异养方式及混养方式的各个实验结果,图2a为生长曲线,图2b为以24小时间隔测定的比生长速率,图2c为有机碳源消耗量和生物质产量的对比图,图2d为以24小时间隔测定的atp/adp比率,图2e为在48小时时间点测定的碳酸酐酶比活度,图2f为在48小时时间点测定的核酮糖-1,5-二磷酸羧化酶比活度。
53.根据在上述培养条件下做120小时实验的结果,可得到在生长活跃的时间点(72小时)以混养方式得到的生物质量多于以自养方式及异养方式得到的生物质量,但少于以自养方式及异养方式得到的生物质之和的结果(参照图2a)。
54.对于比生长速率(μ)而言,可得到在生长活跃的48~72小时区间混养方式的比生长速率大于以自养方式及异养方式得到的比生长速率,但少于以自养方式及异养方式得到的生物质之和的结果(参照图2b)。
55.不仅如此,可确认有机碳源消耗和生物质生长比(y
a/b
)的对比值在异养方式中高于混养方式(参照图2c)。换句话说,当消耗等量的有机碳源时,相比于混养方式,在异养方式中将有机碳源转化为生物质的比率高。为了掌握其相关原因,以24小时间隔测定各个培养环境下的atp/adp比率。atp/adp比率为可确认tca电路的活化程度的指标,atp/adp比率越高,tca电路的活性度可以越高。根据测定结果,呈现异养方式的atp/adp比率高于混养方式的atp/adp比率(参照图2d)。当将其与图2c一同考虑时,可确认异养方式下的有机碳源在tca电路中有效使用,以使有机碳源转化为生物质的比率高于混养方式。另一方面,由于在
微藻类无需光合作用过程的情况下利用有机碳源,因而在无机碳源和有机碳源共存的状态下,更喜欢利用有机碳源。即,在无机碳源和有机碳源同时存在的条件下,微藻类的光合作用量减少,这导致来自无机碳源的固碳(carbon fixation)量的减少。碳酸酐酶(carbonic anhydrase,ca)和核酮糖-1,5-二磷酸羧化酶(rubisco)为二氧化碳浓缩机制(ccm)中两种核心酶,根据两种酶的活性,可测定固碳的活化度。在tca电路最活化的48小时区间测定各个培养方式(自养方式、异养方式、混养方式)的碳酸酐酶活性度(参照图2e)和核酮糖-1,5-二磷酸羧化酶的活性度(参照图2f)。确认到碳酸酐酶的酶活化度在自养方式中相比于异养方式和混养方式高出2.67倍左右,在异养方式和混养方式中其值相似。核酮糖-1,5-二磷酸羧化酶活化度也同样在自养方式中相比于异养方式和混养方式高出3.5倍左右,异养方式和混养方式呈现相似的值。在混养方式中通过二氧化碳浓缩机制的无机碳源的固碳效率低于自养方式,当考虑二氧化碳浓缩机制和从有机碳源到生物质的有效转化时,若存在等量的碳源,则相比于混养方式,分离自养方式和异养方式的方式可被视为更有效的培养方式。
56.实施例2.利用作为有机碳源的乙酸的混养方式和利用无机碳源的自养方式的生物质及tca电路中间产物的比较
57.将乙酸用作有机碳源时,将基于有机碳源供给的混养方式的生物质和tca电路中间产物与自养方式比较。为了光合作用,在生长期间内,混养方式和自养方式中持续供给光。并且,在本实验中,将co2用作无机碳源,将乙酸用作有机碳源,将其浓度分别变更为0.5g/l、1g/l、2g/l来做实验。作为混养方式的对照组,使用自养方式的烧瓶,将co2用作无机碳源。对于混养方式和自养方式而言,如同在一般的实验室单位的培养中执行一样继续供给光。
58.图3a表示实施例2的自养方式和将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式干菌量变化,图3b表示实施例2的自养方式和将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式的tca电路内中间产物的浓度(单位:n mol(107cells)-1
)。
59.通过以提出的所有培养方式得到的生物质(图3a)和在生长最活跃的时间点(60小时)测定的tca电路中间产物表和图(参照图3b、[表1])呈现。通常,呈现供给的有机碳源的量越多,可得到的生物质的量增加的倾向。但是,将乙酸用作有机碳源时,与通常的倾向存在大大差异。确认到当供给最多的碳源(乙酸2g/l)时,在72~120小时区间,干菌量(dcw)最少,当将乙酸1g/l供给为有机碳源时,在72~120小时区间,干菌量(dcw)最大,呈现相比于将乙酸2g/l供给为有机碳源的情况,多出12%的干菌量(参照图3a)。因此,可知即使ph、光量等其他条件相同,乙酸的初始浓度和细胞生长不成正比。
[0060]
当利用高浓度乙酸时,为了分析细胞生长的抑制,通过超高效液相色谱(uplc)设备分析各个不同浓度tca电路的中间产物。在tca电路中,有机碳源按丙酮酸盐(pyruvate)、乙酰-coa(acetyl-coa)、柠檬酸盐(citrate)、琥珀酸盐(succinate)、富马酸盐(fumarate)、苹果酸盐(malate)的顺序转化。但是,据熟知,在微藻类的生长中将乙酸用作有机碳源时,琥珀酸盐的过度生成抑制细胞生长。将乙酸2g/l供给为有机碳源时,相比于自养方式和供给更低的浓度的乙酸时,可知琥珀酸盐的中间产物的量多(参照图3b及[表1])。并且,高浓度的乙酸增加抑制atp的效果,抑制tca电路的活化,由此妨碍细胞生长,因而特定浓度以上的有机碳源的供给有可能妨碍用于大量培养的细胞的急速生长。
[0061]
[表1]自养方式和将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式的tca电路内中间产物的浓度(单位:n mol(107cells)-1
)
[0062][0063]
实施例3.利用作为有机碳源的葡萄糖的混养方式和利用无机碳源的自养方式的生物质及tca电路中间产物的比较
[0064]
在实施例3中,将葡萄糖用作有机碳源时,比较基于有机碳源的供给的自养方式和混养方式的生物质和tca电路内中间产物。在混养方式和自养方式中均为了光合作用而在生长期间内持续供给光。并且,将co2供给为无机碳源,将葡萄糖(0.167g/l、0.333g/l、0.667g/l)供给为有机碳源。其中,葡萄糖0.167g/l在与乙酸0.5g/l相同的浓度中具有相同的碳量,葡萄糖0.333g/l、0.667g/l分别在与乙酸1g/l、2g/l相同的浓度中具有相同的碳量。
[0065]
图4a表示自养方式和将葡萄糖用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式干菌量变化,图4b表示自养方式和将葡萄糖用作有机碳源时(0.5g/l、1g/l、2g/l)的混养方式的各个不同培养方式的tca电路内中间产物的浓度(单位:n mol(107cells)-1
)。
[0066]
将葡萄糖用作有机碳源时,呈现与低浓度下将乙酸用作有机碳源的情况相似的倾向性,但越使有机碳源的浓度变高,越呈现不同的倾向。以所有培养方式得到的产量示于图4a中。培养4天之后,与乙酸不同,将葡萄糖用作有机碳源时,与通常的倾向相同,投入的有机碳源的量越多,混养方式中的生长量呈现越大。但是,投入的有机碳源的量并不与生物质的量成正比地增加。
[0067]
通过依次表示在生长最活跃的时间点(60小时)测定的tca电路的中间产物的表和图(图4b、[表2])分析。当供给葡萄糖0.667g/l时,相对高于供给葡萄糖0.333g/l时的柠檬酸盐(citrate)浓度使细胞的生长变慢。并且,引起过量的乙酰-coa(acetyl-coa)和柠檬酸
盐(citrate)糖异生作用(gluconeogenesis),抑制葡萄糖的糖酵解过程(glycolysis)。由于该糖酵解过程的抑制,减少对细胞生长起到重要作用的丙酮酸盐和乙酰-coa,因而可确认投入的有机碳源的量变多时,生物质的生产收率降低。
[0068]
[表2]自养方式和将葡萄糖用作有机碳源时(0.167g/l、0.333g/l、0.667g/l)的混养方式的各个不同培养方式的tca电路内中间产物的浓度(单位:n mol(107cells)-1
)
[0069][0070]
实施例4.利用乙酸、葡萄糖的依次反复自养-异养方式
[0071]
根据实施例2和实施例3的结果,可通过适量的有机碳源的供给实现相比于有机碳源消耗高的细胞生长。长时间的曝光可对光合受体系统产生损伤,由此光合作用有可能被抑制。
[0072]
图5a表示实施例4的原壳小球藻的不同光周期/暗周期(24小时/0小时、16小时/8小时、14小时/10小时、12小时/12小时)的干重,图5b表示在实施例4的依次反复自养-异养方式中将乙酸用作有机碳源时(0.5g/l、1g/l、2g/l)的干菌量变化,图5c表示在实施例4的依次反复自养-异养方式中将葡萄糖用作有机碳源时(0.167g/l、0.333g/l、0.667g/l)的干菌量变化(图下端所示的区间为0~16、24~40、48~64、72~88、96~112小时的光周期及16~24、40~48、64~72、88~96、112~120小时的暗周期)。
[0073]
原壳小球藻在光周期/暗周期为16小时/8小时的情况下,具有最优秀的生长性(参照图5a)。并且,现有的混养方式的缺点在于,基质的高浓度抑制光合作用,由此降低微藻类的生产率和固碳效率。因此,为了阻止固碳效率减少和光合作用抑制,提出在光周期利用自养方式且在暗周期利用通过供给低浓度的有机碳源的异养方式的依次反复自养-异养方式。利用这种方式克服混养方式的缺点,在利用高浓度二氧化碳的大规模培养中还利用有机碳源提高生物质生产率的同时可缩短细胞培养的周期。
[0074]
在依次反复自养-异养方式中,将乙酸(0.5g/l、1g/l、2g/l)和葡萄糖(0.167g/l、0.333g/l、0.667g/l)用作有机碳源进行实验,在图5b及图5c中比较其结果。将光周期/暗周期设定为16小时/8小时,在相当于光周期的0~16、24~40、48~64、72~88、96~112小时内
供给光和5%co2,在相当于暗周期的16~24、40~48、64~72、88~96、112~120小时内,在各个暗周期供给规定浓度的有机碳源作为基质。并且,作为对照组,利用继续供给光和5%co2的自养方式。
[0075]
将乙酸用作基质时,在暗周期持续供给1g/l的乙酸时生长性最优秀,与对照组比较确认到以96小时为基准的生物质提高27.2%(在96小时之后因氮源枯竭而未引起追加的生物质生长)。在暗周期持续供给2g/l的乙酸时,因琥珀酸盐的形成而使细胞生长被抑制,相比于持续供给1g/l的乙酸时,生物质生长变慢(参照图5b)。
[0076]
将葡萄糖用作基质时,供给最多基质而在暗周期持续供给0.667g/l的葡萄糖时生物质生长性优秀,与对照组比较确认到以88小时为基准的生物质进一步增加58.1%(在88小时之后因氮源枯竭而未引起追加的生物质生长)(参照图5c)。
[0077]
因此,可确认依次反复自养-异养方式是比起现有的自养方式优秀的培养方式。
[0078]
实施例5.将乙酸和葡萄糖利用为有机碳源时,用于将生物质转化最大化的依次反复自养-异养方式
[0079]
在微藻类的依次反复自养-异养方式中,投入等量的基质时,随着培养时间的经过,每个微藻类细胞基质的量减少,细胞的生长速度减少。因此,为了使细胞的生长速度持续,与微藻类细胞的量成正比地增加暗周期放入的基质的量。作为对照组,利用供给5%co2,并使光周期持续24小时的自养方式。将乙酸用作有机碳源时,在依次反复自养-异养方式中作为暗周期的16~24、40~48、64~72、88~96小时内分别投入0.5g/l、1g/l、1.5g/l、1.75g/l,在混养方式中投入4.75g/l而使基质的量相同,测定干重。同样,将葡萄糖用作有机碳源时,在依次反复自养-异养方式中作为暗周期的16~24、40~48、64~72、88~96小时内分别投入0.167g/l、0.333g/l、0.5g/l、0.583g/l,在混养方式中投入葡萄糖1.583g/l,使给药的基质的量相同之后,测定干重。
[0080]
图6a表示将乙酸用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的干菌量变化,图6b表示将葡萄糖用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的干菌量变化(图下端所示的区间为0~16、24~40、48~64、72~88、96~112小时的光周期及16~24、40~48、64~72、88~96、112~120小时的暗周期)。图6c表示将乙酸用作实施例5的有机碳源时的混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的有机碳源消耗量和比生长速率的对比值,图6d表示将葡萄糖用作实施例5的有机碳源时的混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的有机碳源消耗量和比生长速率的对比值。图6e表示将乙酸用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的atp/adp比,图6f表示将葡萄糖用作实施例5的有机碳源时的自养方式(autotrophy)、混养方式(mixotrophy)、依次反复自养-异养方式(repeated sequential auto-and heterotrophy)中的atp/adp比。
[0081]
当使用如上所述的培养方式时,若将乙酸用作基质,则在96小时内依次反复自养-异养方式的细胞生长相比于混养方式高32.3%,相比于自养方式高50.2%(参照图6a)。在
反复自养-异养方式中,由于与微藻类细胞的量成正比地投入有机碳源,因而收率(y
x/a
)相比基质消耗量,相似地维持在40~96小时,相比于混养方式高2.59倍以上(参照图6c)。并且,确认到作为tca电路的活化指标的atp/adp比率也相比于反复自养-异养方式高2.17倍以上(参照图6e)。
[0082]
将葡萄糖用作基质时,同样在96小时区间,依次反复自养-异养方式的细胞生长相比于混养方式高12.6%,相比于自养方式高50.1%(参照图6b)。在与微藻类细胞的量成正比地投入有机碳源的方式中,以高的状态维持tca电路的活化,呈现更高的相比于基质消耗量的收率(y
x/a
)(参照图6d及图6f)。
[0083]
并且,确认到作为碳浓缩机制(ccm)的指标的碳酸酐酶比活度(carbnonic anhydrase specific activity)和核酮糖-1,5-二磷酸羧化酶比活度(rubisco specific activity)也在反复自养-异养方式中相比于混养方式分别高84.8%、149%。不仅如此,如下列[表3]所示,通过生产率求得的最大固碳比率也因高的生产率而相比于自养方式高42.3%。
[0084]
[表3]将乙酸用作基质时的不同培养方式的干重、比生长速率、碳酸酐酶比活度、核酮糖-1,5-二磷酸羧化酶比活度、最大固碳比率(最大固碳比率,p
co2
=p
max
×
1.88。依次反复自养-异养方式,将除了异养方式引起的生产率之外的剩余生产率值乘以1.88倍)
[0085]
[0086]
因此,可知依次反复自养-异养方式防止光合作用效率下降,促进tca电路的活化,由此贡献于细胞生长。进而,光合作用效率的保持对固碳(carbon fixation)产生大的影响,因而在ccu观点上也可被视为有意义的培养方式。
[0087]
以上,通过具体实施例详细说明本发明,但其用于具体说明本发明,本发明不局限于此,在本发明的技术思想内可由本发明所属技术领域的普通技术人员变形或改良,这是显而易见的。
[0088]
本发明的简单变形至变更均属于本发明的领域,本发明的具体保护范围将根据所附的权利要求书变得明确。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。