遗传修饰真菌和与其相关方法和用途
发明领域
1.本发明涉及工业生物技术、可再生原料材料和微生物生产生物体的领域。具体地说,本发明涉及生产乳酸或乳酸酯或一种或多种选自由聚合物、聚酯和聚乳酸组成的组的产物的方法。本发明还涉及包含增加的特定酶活性的遗传修饰真菌、制备所述遗传修饰真菌的方法、和所述真菌用于生产乳酸、乳酸酯或聚合物的用途。
2.发明背景
3.乳酸发酵是例如葡萄糖和其他己糖(六碳糖)或六碳糖的双糖(例如蔗糖或乳糖)藉以转化成能量和乳酸的厌氧代谢过程。目前在美国,乳酸由玉米淀粉生产,并且在其他地方,由诸如甜菜和甘蔗的其他糖来源生产。所述淀粉和糖来源主要包含简单碳水化合物。生产乳酸用于食物用途,但是也作为生产聚乳酸(pla)的前体。pla是越来越多地用于制造生物塑料的可再生聚合物。对于pla生产来说,需要通常不由野生型微生物生产的光学纯异构体。
4.需要用于生产乳酸的更便宜和生态相容的原料。举例来说,细菌唾液乳杆菌(lactobacillus salivarius)已用于将大豆糖蜜转化成乳酸(montelongo j等人,1993,journal of food science,第58卷,863-866)。然而,仍然存在对于能够将复杂碳水化合物诸如低聚半乳糖转化成乳酸的有效真菌的显著未满足的需要。
5.发明简述
6.本发明的目标,即获得生产乳酸和/或乳酸酯的有效方法以及获得能够有效地将碳水化合物转化成乳酸和/或乳酸酯的真菌通过使用真菌的遗传修饰来实现。
7.本发明使得能够克服现有技术的缺陷,包括但不限于缺乏能够将复杂碳水化合物(包括但不限于碳水化合物大豆糖蜜)转化成乳酸的真菌。事实上,与例如玉米淀粉和蔗糖相比,本发明的真菌和方法允许使用替代碳源物以工业规模生产乳酸。因此,本发明通过允许利用包含复杂碳水化合物的工业侧流来为生态发展提供价值。
8.目前,例如pla的成本与合成塑料不具有竞争性。然而,本发明允许降低聚合物诸如pla或聚酯的生产成本。
9.意外地,本发明的真菌和方法使得能够以有利于工业经济运行的高产率、滴度和生产力来生产纯l-乳酸异构体。
10.本发明涉及生产乳酸和/或乳酸酯的方法,所述方法包括
11.提供经遗传修饰来增加乳酸脱氢酶和α-半乳糖苷酶活性的真菌,
12.在包含碳源物(例如包含低聚半乳糖的碳源物)的培养基中培养所述真菌来获得乳酸和/或乳酸酯。
13.另外,本发明涉及包含增加的乳酸脱氢酶和α-半乳糖苷酶活性的遗传修饰真菌。
14.本发明还涉及制备包含增加的乳酸脱氢酶和α-半乳糖苷酶活性的本发明的遗传修饰真菌的方法,其中所述方法包括提供真菌和遗传修饰真菌来增加乳酸脱氢酶和α-半乳糖苷酶活性。
15.此外,本发明涉及包含增加的乳酸脱氢酶和α-半乳糖苷酶活性的本发明的真菌用
于生产乳酸和/或乳酸酯或用于生产聚合物,任选地聚酯或聚乳酸的用途。
16.并且此外,本发明涉及生产一种或多种选自由聚合物、聚酯和聚乳酸组成的组的产物的方法,所述方法包括在含有碳源物例如低聚半乳糖的培养基中培养本发明的遗传修饰真菌(包含增加的乳酸脱氢酶和α-半乳糖苷酶活性)以便生产乳酸,回收所得乳酸和使用回收的乳酸来生产聚合物、聚酯和/或聚乳酸。
17.本发明的其他目标、细节和优势由以下附图、详细说明和实施例而变得显而易知。
18.附图简述
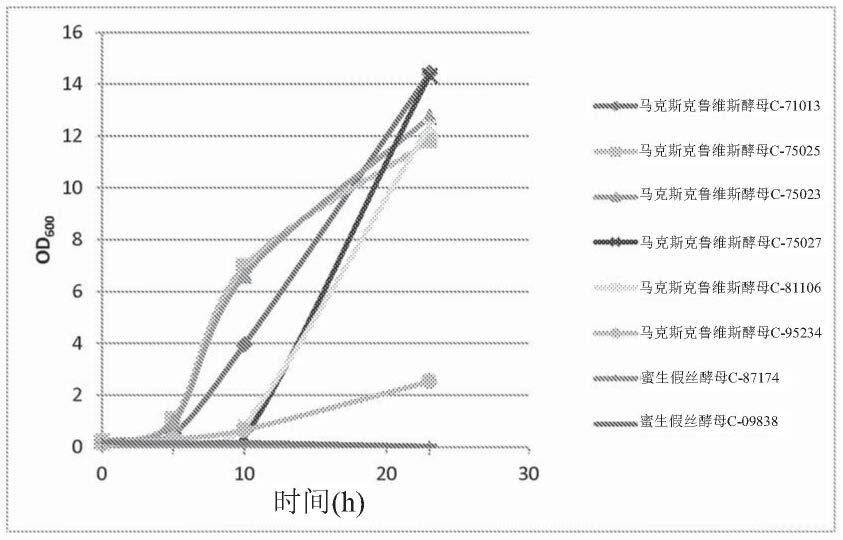
19.图1示出使用半乳糖作为唯一碳源,马克斯克鲁维酵母(kluyveromyces marxianus)和蜜生假丝酵母(candida apicola)的各种野生型真菌菌株的生长。菌株的生长通过量化od
600
来评估。
20.图2示出使用半乳糖作为唯一碳源,表达乳酸脱氢酶(ldh)的四种真菌菌株的生长。菌株的生长通过量化od
600
来评估。
21.图3示出在具有1%蜜二糖或蜜三糖作为碳源的sc-ura培养基上,表达编码α-半乳糖苷酶的不同基因的酿酒酵母(s.cerevisiae)菌株的生长。在30℃下,伴以220rpm振荡,菌株在24孔板中以4ml培养体积培养过夜。
22.图4示出来自亲代菌株(vtt-c-02453ura3δ/ura3δ)和表达不同α-半乳糖苷酶的衍生菌株的1:3稀释大豆糖蜜的24h培养物的通过hplc量化的乙醇滴度(g/l)。
23.图5示出来自亲代菌株(vtt-c-02453ura3δ/ura3δ)和表达不同α-半乳糖苷酶的衍生菌株的1:3稀释大豆糖蜜的24h培养物的通过hplc量化的残留糖(g/l)。
24.图6示出来自酿酒酵母e79-4和表达不同α-半乳糖苷酶的衍生菌株的生物反应器培养物的通过hplc量化的乳酸(g/l)。菌株使用大豆糖蜜作为唯一碳源来生长。
25.图7示出来自酿酒酵母e79-4和表达不同α-半乳糖苷酶的衍生菌株的生物反应器培养物的量化残留低聚半乳糖(g/l)。菌株使用大豆糖蜜作为唯一碳源来生长。结果报告为蜜三糖、水苏糖、毛蕊花糖、蜜二糖、甘露三糖和甘露四糖的浓度的总和。
26.图8示出用于实施例1-4中的质粒的图谱。
27.图9揭示来自使用大豆糖蜜作为修饰酵母菌株vtt c-191026和表达额外不同α-半乳糖苷酶基因拷贝的菌株的碳源的摇瓶培养物的量化残留四和三糖。
28.图10揭示来自使用大豆糖蜜作为修饰酵母菌株vtt c-191026和修饰库德毕赤酵母(p.kudriavzevii)菌株vtt c-201040的碳源的摇瓶培养物的量化所生产乳酸和残留三和二糖。
29.图11示出用于实施例6中的质粒的图谱。
30.序列表
31.seq id no:1:α-半乳糖苷酶(黑曲霉(a.niger)aglc)之氨基酸序列
32.seq id no:2:α-半乳糖苷酶(里氏木霉(t.reesei)agl1)之氨基酸序列
33.seq id no:3:α-半乳糖苷酶(米黑根毛霉菌(rhizomucor miehei)gal36)之氨基酸序列
34.seq id no:4:α-半乳糖苷酶(赤霉菌(gibberella)属f75 gal36)之氨基酸序列
35.seq id no:5:α-半乳糖苷酶(费氏曲霉(aspergillus fischeri)gal27b)之氨基酸序列
36.seq id no:6:α-半乳糖苷酶(酿酒酵母mel5)之氨基酸序列
37.seq id no:7:编码α-半乳糖苷酶(黑曲霉aglc)之多核苷酸序列
38.seq id no:8:编码α-半乳糖苷酶(里氏木霉agl1)之多核苷酸序列
39.seq id no:9:编码α-半乳糖苷酶(米黑根毛霉菌gal36)之多核苷酸序列
40.seq id no:10:编码α-半乳糖苷酶(赤霉菌属f75 gal36)之多核苷酸序列
41.seq id no:11:编码α-半乳糖苷酶(费氏曲霉gal27b)之多核苷酸序列
42.seq id no:12:编码α-半乳糖苷酶(酿酒酵母mel5)之多核苷酸序列
43.seq id no:13:引物32mel5-atg-f
44.seq id no:14:引物33mel5-stopr
45.seq id no:15:质粒pmie-16(黑曲霉aglc;q9uuz4)之密码子优化多核苷酸序列,
46.seq id no:16:质粒pmie-17(里氏木霉agl1;q92456)之密码子优化多核苷酸序列
47.seq id no:17:质粒pmie-18(米黑根毛霉菌gal36;h8y263)之密码子优化多核苷酸序列
48.seq id no:18:质粒pmie-19(赤霉菌属f75 gal36;c6fjg8)之密码子优化多核苷酸序列
49.seq id no:19:质粒pmie-20(费氏曲霉gal27b;aja29661.1)之密码子优化多核苷酸序列
50.seq id no:20:质粒pmie-5(酿酒酵母mel5)之多核苷酸序列
51.seq id no:21:引物2scadh1-150f
52.seq id no:22:引物5scadh1 stopr
53.seq id no:23:质粒pmie-21b之多核苷酸序列
54.seq id no:24:质粒pmie-24b之多核苷酸序列
55.seq id no:25:质粒pmie-25b之多核苷酸序列
56.seq id no:26:质粒pmie-26a之多核苷酸序列
57.seq id no:27:质粒pmie-031之多核苷酸序列
58.seq id no:28:质粒pmie-032之多核苷酸序列
59.seq id no:29:质粒pmie-034之多核苷酸序列
60.seq id no:30:引物3scpdc5-210f
61.seq id no:31:引物6scpdc5 stopr
62.seq id no:32:引物4scpdc5-136f
63.seq id no:33:质粒pmie-8之多核苷酸序列
64.seq id no:34转化酶(酿酒酵母suc2)之氨基酸序列
65.seq id no:35编码转化酶(酿酒酵母suc2)之多核苷酸序列
66.seq id no:36质粒pmipk124之多核苷酸序列
67.seq id no:37质粒pekopa8之多核苷酸序列
68.seq id no:38质粒pekopa9之多核苷酸序列
69.发明详述
70.本发明的目标通过增加乳酸脱氢酶活性和α-半乳糖苷酶活性来实现。本公开的发明人能够提供经遗传修饰来增加乳酸脱氢酶和α-半乳糖苷酶活性的真菌。
71.在生产乳酸和/或乳酸酯的本发明的方法中,经遗传修饰来增加乳酸脱氢酶和α-半乳糖苷酶活性的真菌在包含碳源物的培养基中培养以便获得所述乳酸和/或乳酸酯。
72.如在此使用,“乳酸”是指具有分子式ch3ch(oh)co2h(化学式c3h6o3)的有机酸。在行业中,乳酸发酵通过将碳源物(例如简单碳水化合物诸如葡萄糖、蔗糖或半乳糖)转化成乳酸的微生物来执行。
73.乳酸以两个立体异构体形式,d和l乳酸,和这些异构体的所谓外消旋混合物出现。在一个实施方案中,通过本发明的方法或遗传修饰真菌来生产的乳酸是l-乳酸异构体或d-乳酸异构体或其组合。在一个实施方案中,乳酸是光学纯乳酸异构体,任选地l-乳酸异构体。如在此使用,“光学纯乳酸异构体”是指基本上只包含一种立体异构体形式的乳酸并且不包含其镜像(例如约95%或更多、约96%或更多、约97%或更多、约98%或更多、或约99%或更多(例如99.5%或更多)的一种立体异构体形式的乳酸)的溶液或固体。
74.本发明的有效真菌被工程化来水解碳水化合物并且将其转化成乳酸,例如光学纯l-乳酸。所述真菌用于在包含碳源物例如包含简单和/或复杂碳水化合物的碳源物的培养基中培养真菌来生产乳酸或乳酸酯的方法中。事实上,本发明使得能够在大规模生产过程期间操作和控制碳源,从而为制造商提供灵活性和对于所述过程的极好控制。如在此使用,“简单碳水化合物”是指简单糖,其可分类为单一糖(单糖),包括葡萄糖、果糖和半乳糖,或双重糖(双糖),包括蔗糖、乳糖和麦芽糖。如在此使用,“复杂碳水化合物”是指包含三个或更多个连接糖的多糖。事实上,与较短非多糖相比,分解多糖耗费更长时间。
75.意外地,在一个实施方案中,本发明的真菌和方法能够利用复杂碳水化合物,例如大豆糖蜜,作为碳源物。在本发明的具体实施方案中,碳源物包含复杂碳水化合物或是复杂碳水化合物。在更具体实施方案中,碳源物包含低聚半乳糖或是低聚半乳糖。发现于植物材料中的最常见低聚半乳糖是蜜三糖家族低聚糖(rfo)。这些分子是蔗糖的衍生物,具有额外α-(1
→
6)-连接半乳糖基部分。根据连接半乳糖基单元的数目的不同rfo糖包括蜜三糖(一个半乳糖单元)、水苏糖(两个半乳糖单元)、毛蕊花糖(三个半乳糖单元)和ajucose(四个半乳糖单元)。除了rfo以外,例如豆类可包含含有末端肌醇基团的其他低聚半乳糖,诸如属于碳水化合物的半乳糖肌醇、半乳糖松醇和荞麦醇系列的那些低聚半乳糖。在本发明的一个实施方案中,碳源物包含所述碳源物中的总碳水化合物的至少约10重量%、20重量%、30重量%、40重量%、50重量%、60重量%、70重量%、80重量%、或90重量%的复杂碳水化合物或低聚半乳糖,和/或所述碳源物中的总碳水化合物的至少约10重量%、20重量%、30重量%、40重量%、50重量%、60重量%、70重量%、80重量%或90重量%的简单碳水化合物(例如葡萄糖、果糖、半乳糖、蔗糖、乳糖或麦芽糖或其任何组合)。
76.在本发明的一个实施方案中,碳源物包含一种或多种低聚半乳糖,其选自由以下组成的组:蜜二糖、甘露三糖、甘露四糖、蜜三糖、水苏糖、毛蕊花糖、ajucose、半乳糖肌醇、双半乳糖基肌醇、半乳糖松醇a、半乳糖松醇b、鹰嘴豆糖醇、荞麦醇b1、荞麦醇b2和其任何组合。在具体实施方案中,低聚半乳糖是来自由蜜三糖、水苏糖、毛蕊花糖、蜜二糖、甘露三糖和甘露四糖的组成的组的一个或多个。
77.在一个实施方案中,碳源物包含葡萄糖、果糖、半乳糖、蔗糖、乳糖、麦芽糖、淀粉、纤维素和/或其任何组合。如在此使用,“淀粉”是指具有式(c6h
10
o5)
n-(h2o)的聚合物碳水化合物,即包含或由通过糖苷键接合的大量葡萄糖单元组成。如在此使用,“纤维素”是指具有
式(c6h
10
o5)n的有机化合物,由多个(例如一百个至数千个)β(1-4)连接d-葡萄糖单元的直链组成的多糖。
78.用于本发明中的碳源物可获自或可来自任何含碳材料,例如不同含碳材料的组合。在一个实施方案中,碳源物来自豆类诸如大豆(例如,扁豆)、蚕豆、豌豆、鹰嘴豆、玉米(例如,玉米芯仁)、甘蔗(例如,植物)、甜菜(甜菜的甜菜头)、木质纤维素或其任何组合;且/或碳源物包括大豆糖蜜、甘蔗糖蜜、甜菜糖蜜和/或柑橘糖蜜。如在此使用,“木质纤维素”是指包含纤维素、半纤维素和木质素的材料。例如大豆、甘蔗、甜菜或柑橘的“糖浆”是指分别将扁豆、植物、甜菜或果实提纯成糖所得的产物。
79.在一个实施方案中其中培养真菌以便生产乳酸和/或乳酸酯的碳源物或培养基包含5-100重量%大豆糖蜜(例如,至少约5重量%、10重量%、20重量%、30重量%、40重量%、50重量%、60重量%、70重量%、80重量%、或90重量%)。
80.举例来说,大豆糖蜜是生产大豆蛋白质浓缩物的副产物。这是通常注定用于生产动物饲料或甚至燃烧的低价值物流。然而,它可含有很高浓度的可增值的大豆碳水化合物(例如,》300g/l)。难题是这些糖是非传统低聚糖诸如蜜三糖和水苏糖,需要水解,然后所有产生的单糖葡萄糖、果糖和半乳糖需要代谢成产物。与例如玉米淀粉和蔗糖相比,大豆糖蜜是用于生产乳酸的更廉价原料的实例。大豆糖蜜可用作本身用于生产真菌乳酸的碳源物;没有额外营养物要求,从而进一步有助于最大限度地减少乳酸的生产成本。
81.为了生产乳酸,遗传修饰真菌在培养基中培养,所述培养基包含一种或多种适合碳源和任选地选自由以下组成的组的其他成分:氮或氮源(诸如氨基酸、蛋白质、无机氮源诸如氨或铵盐)、酵母萃取物、胨、无机物和维生素。在一个实施方案中,真菌的培养在本领域技术人员已知的合适条件下执行。合适培养条件,诸如温度、ph、细胞密度、营养物的选择等在本领域技术人员的知识范围内并且所述本领域技术人员能够选择、修改或控制所述条件。在具体实施方案中,培养温度为约25至45℃(例如,约30-35℃)且/或培养基的ph为2-10(例如,3-6)。自然地,合适培养条件可取决于特定真菌。培养条件可在生产乳酸或乳酸酯的方法期间保持或替代地,其可定期调整。在一个实施方案中,当一个以上罐用于生产乳酸或乳酸酯的方法中时,培养条件可能在不同罐中有所不同。
82.在本发明的一个实施方案中,乳酸或乳酸酯通过厌氧、准厌氧或需氧发酵来生产。
83.在一个实施方案中,真菌的培养以连续发酵方法或以分批或分批补料发酵方法来执行。
84.在本发明的一个实施方案中,在培养基中培养遗传修饰真菌之后,方法还包括从培养基中回收所得乳酸或乳酸酯。事实上,可在不干扰细胞的情况下执行从培养基中的回收。在一个实施方案中,在培养基中培养真菌之后,方法还包括分离和/或纯化乳酸或乳酸酯。本领域技术人员已知的任何合适方法可用于分离乳酸或乳酸酯。举例来说,常见分离技术可用于从培养基中移除生物质,并且常见隔离程序可用于从无真菌培养基中获得乳酸或乳酸酯。乳酸或乳酸酯可在其生产的同时加以分离,或它可在乳酸或乳酸酯生产终止之后从培养基中分离。乳酸和乳酸酯可通过使用在本领域中已知的任何常规方法来回收、分离和/或纯化,诸如吸附、离子交换程序、色谱方法、双相萃取、分子蒸馏、熔融结晶、萃取、蒸馏或其任何组合。
85.在一个实施方案中,用于生产方法期间的真菌得以回收并且重复使用于后续生产
方法中。
86.pla,一种热塑性脂族聚酯,可从乳酸,例如,从通过本发明的方法产生并且任选地回收、分离和/或纯化的乳酸中,通过包括但不限于以下的不同方法来制备:丙交酯(从乳酸衍生)与各种金属催化剂的开环聚合、乳酸单体的直接缩合、乳酸的聚合、使乳酸与沸石接触、pla从乳酸的直接生物合成。在一个实施方案中,本发明的方法包括从所获得的乳酸来制备pla。
87.本发明涉及遗传修饰酵母和与其相关的方法和用途,其中酵母具有增加的乳酸脱氢酶和α-半乳糖苷酶活性。用于本发明中的遗传修饰至少用于修改,更具体地说增加,乳酸脱氢酶和α-半乳糖苷酶的活性。乳酸脱氢酶允许生产乳酸和乳酸酯并且α-半乳糖苷酶使得能够降解和消耗复杂碳水化合物包括但不限于大豆糖蜜碳水化合物。
88.如在此使用,“乳酸脱氢酶活性”是指催化将丙酮酸酯转换成乳酸酯的能力。因此,“乳酸脱氢酶“是指具有将丙酮酸酯转化成乳酸酯的活性的蛋白质。l-乳酸脱氢酶(l-ldh)将丙酮酸酯转化成l-乳酸酯并且d-乳酸脱氢酶(d-ldh)将丙酮酸酯转化成d-乳酸酯。l-乳酸脱氢酶和d-乳酸脱氢酶分别分类为ec 1.1.1.27和ec 1.1.1.28。乳酸脱氢酶(ldh)不仅是指真菌或细菌(诸如米根霉或瑞士乳杆菌)而且是指来自任何微生物、生物体或哺乳动物例如牛的任何其他ldh同系物。另外,所有同功酶、同种型和变体包括在ldh的范围内。在具体实施方案中,ldh是l-ldh。相应地,米根霉ldha(af226154)和ldhb(af226155)的ldh蛋白质和ldh基因在skory(2000appl.environ.microbiol.66:2343-2348)的文章中得以鉴定,并且瑞士乳杆菌ldhl(u07604)在savijoki k.,palva a.(1997.appl.environ.microbiol.63:2850-2856)的文章中得以鉴定。合适开放解读码组(orf)的实例包括但不限于米根霉ldha(q9p4b6)和ldhb(q9p4b5)和瑞士乳杆菌ldhl(cab03618)的orf。举例来说,ldh1、ldh2、ldh3、ldh4、ldh5、ldh6a、ldh6b、ldha、ldhb、ldhc和ldhl编码在ldh的范围内的相关但是不相同多肽。编码相关但是不相同多肽的基因的数目取决于所讨论的微生物或生物体。
89.如在此使用,“α-半乳糖苷酶活性”是指催化来自包括半乳糖和蜜三糖低聚糖、半乳甘露聚糖和半乳糖脂的各种α-半乳糖苷的非还原末端α-半乳糖基残基的水解的能力。因此,“α-半乳糖苷酶”是指具有水解来自各种α-半乳糖苷的非还原末端α-半乳糖基残基的活性的蛋白质。α-半乳糖苷酶分类为ec 3.2.1.22。α-半乳糖苷酶不仅是指真菌(诸如酿酒酵母)或细菌而且是指来自任何微生物或生物体的任何其他α-半乳糖苷酶同系物。另外,所有同功酶、同种型和变体包括在α-半乳糖苷酶的范围内。举例来说,(例如,里氏木霉)agl1、agl2和agl3,(例如,黑曲霉)agla、aglb、aglc和agld,和(例如,酿酒酵母)mel1、mel2、mel5、和mel6编码在α-半乳糖苷酶的范围内的相关但是不相同多肽。编码相关但是不相同多肽的基因的数目取决于所讨论的微生物或生物体。
90.本发明的工程化真菌包含增加蛋白质或酶活性的遗传修饰。如在此使用,“增加的蛋白质或酶活性”是指与野生型蛋白质相比,存在蛋白质的较高活性,或与未修饰细胞或真菌相比,细胞或真菌的较高总蛋白质活性。增加的蛋白质活性可由多肽表达的上调、基因表达的上调、基因的至少一部分的添加(包括基因拷贝的添加或通常在所述细胞或真菌中不存在的基因的添加)、蛋白质的增加和/或蛋白质的增加活性所引起。产生增加的蛋白质或酶活性的具体实例在实施例部分中提供。
91.细胞或真菌中的蛋白质活性的存在、不存在或量可通过在本领域中已知的任何合适方法来检测。合适检测方法的非限制实例包括市场上的商用试剂盒、酶测定、免疫检测方法(例如,对于所述蛋白质具有特异性的抗体)、基于pcr的测定(例如,qpcr、rt-pcr)、和其任何组合。在一个特定实施例中,乳酸脱氢酶的活性通过监测在乳酸锂和nad 存在下培育酶或真菌之后的吸光度来确定,例如如tokuhiro等人(2009,appl micro-biol biotechnol 82,883-890)所描述,且/或α-半乳糖苷酶的活性通过在将酶或真菌与对硝基苯基-α-吡喃半乳糖苷(pnpg)一起培育之后测量所释放的对硝基苯基(pnp)来确定,例如如chen等人(2015,protein expression and purification,110,107-114)所描述,和/或通过在将酶或真菌与甲基伞形基-α-d-吡喃半乳糖苷(mug)一起培育之后,测量所释放的甲基伞形基(mu),例如如等人(2010,j microbiol biotechnol,20(12),1653-1663)所描述。
92.导致增加的蛋白质活性的遗传修饰包括但不限于一或多个基因或其片段的遗传插入、缺失或中断或一或多个核苷酸的插入、缺失、中断或取代,或质粒的添加。如在此使用,“中断”是指将一个或多个核苷酸插入基因或多核苷酸序列中,导致不存在对应蛋白质或存在非功能性蛋白质或具有降低活性的蛋白质。
93.如在此使用,“基因或多肽表达的上调”是指通过与未修饰真菌相比,产生更多产物(例如,相应地mrna或蛋白质)而导致基因或多肽的过度表达。举例来说,一种或多种基因的一或多个拷贝可转化至细胞中以便上调基因表达。术语还涵盖实施方案,其中调控区域诸如启动子或启动子区域得以修饰或改变或插入不天然存在于真菌中的调控区域(例如,启动子)以便允许基因的过度表达。另外,在“遗传修饰”中包括表观遗传修饰诸如减少dna甲基化或组蛋白修饰,导致基因或多肽的表达上调。如在此使用,“增加或上调的表达”是指与没有遗传修饰的野生型真菌相比,所关注的基因或多肽的增加的表达。表达或增加的表达可例如通过蛋白质印迹、rna印迹或dna印迹或定量pcr或本领域技术人员已知的任何其他合适方法来证实。
94.在某些实施方案中,工程化真菌包含至少一个(例如,一个、两个、三个、四个、五个、六个或更多个)异源多核苷酸。任何插入多核苷酸或基因(例如,一个、两个、三个、四个、五个、六个或更多个)可对于宿主真菌来说是异源或同源的。真菌可通过将它用编码异源蛋白质的异源多核苷酸转化来遗传修饰。或者,例如异源启动子或其他调控序列可用于本发明的真菌中。如在此使用,“异源多核苷酸”是指不天然存在于细胞或真菌中的多核苷酸,即细胞或真菌正常地不包含所述多核苷酸。典型地所述异源多核苷酸通过重组技术来插入或修饰。
95.另一方面,任何插入多核苷酸或基因(例如,一个、两个、三个、四个、五个、六个或更多个)可与待遗传修饰的真菌相同或非常同源的。这样,例如与未遗传修饰真菌相比,真菌中的多核苷酸或基因的拷贝数可增加。或者,例如可利用与待遗传修饰的真菌相同或非常同源的启动子或其他调控序列。事实上,取决于所讨论的真菌,本发明的真菌可用通常包含在所述真菌中的多核苷酸来修饰。
96.在具体实施方案中,遗传修饰的真菌最初(即在所述遗传修饰之前)不包含ldh基因(例如,l-ldh基因)和/或α-半乳糖苷酶基因。
97.在本发明的方法、用途或遗传修饰真菌的一个实施方案中,α-半乳糖苷酶是异源α-半乳糖苷酶且/或乳酸脱氢酶是异源乳酸脱氢酶。
98.如果在本发明中利用异源α-半乳糖苷酶,那么它可以是来自任何合适生物体的α-半乳糖苷酶。在这种情况下,所述异源α-半乳糖苷酶必须在本发明中具有功能性。在一个实施方案中,异源α-半乳糖苷酶是例如选自以下属类的酵母或丝状真菌的α-半乳糖苷酶:曲霉(aspergillus)、赤霉菌(gibberella)、小克银汉霉(cunninghamella)、镰刀菌(fusarium)、球囊霉(glomus)、腐质霉(humicola)、被孢霉(mortierella)、毛霉菌(mucor)、青霉菌(penicillium)、腐霉(pythium)、根毛霉(rhizomucor)、根霉(rhizopus)、木霉(trichoderma)和酵母属(saccharomyces),尤其由以下组成的组:玉米赤霉(gibberella zeae)、中间赤霉(gibberella intermdia)、串珠状赤霉(gibberella moniliformis)、藤仓赤霉(gibberella fujikuroi)、尼加迈赤霉(gibberella nygamai)、赤霉菌属f75、镰刀菌属2f75、尖孢镰刀菌(fusarium oxysporum)、芒果镰刀菌(fusarium mangiferae)、层生镰刀菌(fusarium proliferatum)、轮状镰刀菌(fusarium verticilloides)、构巢曲霉(aspergillus nidulans)、米曲霉(aspergillus oryzae)、土曲霉(aspergillus terreus)、黑曲霉(aspergillus niger)、费氏曲霉(aspergillus fischeri)、米黑根霉(rhizopus miehei)、米黑根毛霉菌(rhizomucor miehei)、米根霉(rhizopus oryzae)、里氏木霉(trichoderma reesei)、哈茨木霉(trichoderma harzianum)、长枝木霉(trichoderma longibrachiatum)和酿酒酵母(saccharomyces cerevisiae)。在具体实施方案中,异源α-半乳糖苷酶是,或α-半乳糖苷酶基因是功能α-半乳糖苷酶基因,所述基因编码与例如黑曲霉、赤霉菌属f75、费氏曲霉、里氏木霉、酿酒酵母、米黑根毛霉菌物种中的任何一个的α-半乳糖苷酶基因所编码的蛋白质至少60%、70%、80%、85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、或99%相同的蛋白质。
99.如果在本发明中利用异源乳酸脱氢酶,那么它可以是来自任何合适生物体,包括哺乳动物诸如牛的乳酸脱氢酶。在这种情况下,所述异源乳酸脱氢酶必须在本发明中具有功能性。在具体实施方案中,异源乳酸脱氢酶来自生物体、哺乳动物、微生物、真菌、或细菌,例如任选地来自哺乳动物诸如牛属(bos)(例如黄牛(bos taurus)),真菌诸如克鲁维斯酵母或根霉(例如耐热克鲁维斯酵母(kluyveromyces thermotolerans)或米根霉),或细菌诸如乳杆菌(例如瑞士乳杆菌(lactobacillus helveticus)或干酪乳杆菌(l casei)),片球菌(例如乳酸片球菌(pediococcus acidilactici))或芽孢杆菌属(例如巨大芽孢杆菌(bacillus megaterium)),或来自单细胞原生动物寄生虫例如疟原虫(例如恶性疟原虫(plasmodium falciparum))。在具体实施方案中,异源乳酸脱氢酶是,或ldh基因是功能ldh基因,所述基因编码与例如瑞士乳杆菌、干酪乳杆菌、乳酸克鲁维斯酵母、巨大芽孢杆菌、乳酸片球菌、黄牛、米根霉或恶性疟原虫物种中的任何一个的l-ldh基因所编码的蛋白质至少40%、50%、60%、70%80%、85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、或99%相同的蛋白质。特定d-ldh基因的实例是从瑞士乳杆菌、约氏乳杆菌(l.johnsonii)、保加利亚乳杆菌(l.bulgaricus)、德氏乳杆菌(l.delbrueckiii)、植物乳杆菌(l.plantarum)、戊糖乳杆菌(l.pentosus)和嗜酸乳杆菌(p.acidilactici)获得的那些基因。与这些l-ldh或d-ldh基因相同或在氨基酸水平下与这些基因至少35%、60%、70%或80%相同的功能基因是合适的。在具体实施方案中,l-ldh基因从瑞士乳杆菌获得或与所述基因至少35%、60%、70%、80%、85%、90%或95%相同。另一个合适l-ldh基因从巨大芽孢杆菌获得或与所述基因至少35%、60%、70%、80%、85%、90%或95%相同。合适d-ldh基因
从瑞士乳杆菌获得或与所述基因至少45%、60%、70%、80%、85%、90%或95%相同。
100.在本发明的一个实施方案中,将异源ldh和/或α-半乳糖苷酶基因整合到真菌细胞的基因组中。在具体实施方案中,将ldh和/或α-半乳糖苷酶基因整合到原生pdc基因的基因座中。异源ldh和/或α-半乳糖苷酶基因可例如在对于真菌细胞是原生或异源的启动子的转录控制下。在一个实施方案中,所述方法、用途或真菌可利用转化载体,所述载体包含与例如对于待遗传修饰的真菌是原生的启动子序列可操作地连接的功能ldh和/或α-半乳糖苷酶基因。可使用在不同类型启动子和/或终止子的控制下的不同异源ldh和/或α-半乳糖苷酶基因。
101.在一个实施方案中,转化真菌细胞可含有单一ldh基因和/或α-半乳糖苷酶基因,或多个ldh和/或α-半乳糖苷酶基因,诸如1-10个ldh和/或α-半乳糖苷酶基因,尤其1-5个ldh和/或α-半乳糖苷酶基因。当转化细胞含有多个ldh和/或α-半乳糖苷酶基因时,个别基因可以是同一基因的拷贝,或包括两个或更多个不同ldh和/或α-半乳糖苷酶基因的拷贝。异源和/或内源ldh和/或α-半乳糖苷酶基因的多个拷贝可整合在单一基因座处(以使得其彼此相邻),或真菌细胞的基因组内的多个基因座处。举例来说,类似或不同ldh基因和/或α-半乳糖苷酶基因的两个拷贝可整合在二倍体真菌的同源等位基因处。
102.识别含有所关注的异源多核苷酸的细胞的方法是本领域技术人员熟知的。这些方法包括但不限于pcr和核酸杂交技术诸如rna和dna分析。在一些情况下,免疫组织化学和生物化学技术可用于通过检测所述特定核酸分子所编码的编码酶性多肽的表达来确定是否细胞含有特定核酸。举例来说,具有对于编码酶的特异性的抗体可用于确定是否特定细胞或真菌含有所述编码酶。此外,生物化学技术可用于通过检测由于酶性多肽的表达而生产的有机产物来确定是否细胞含有编码酶性多肽的特定核酸分子。
103.在本发明的方法、用途或真菌的一个实施方案中,真菌被遗传修饰来过度表达编码乳酸脱氢酶的基因和/或编码α-半乳糖苷酶的基因。“过度表达基因”是指当与没有所述修饰的真菌相比时,由于遗传修饰而导致的所述基因的上调表达。在具体实施方案中,所述修饰真菌包含编码乳酸脱氢酶的基因和/或编码α-半乳糖苷酶的基因的一或多个拷贝。
104.在本发明的方法、用途或真菌的一个实施方案中,编码乳酸脱氢酶的基因选自由以下组成的组:ldh1、ldh2、ldh3、ldh4、ldh5、ldh6a、ldh6b、ldha、ldhb、ldhc和ldhl,且/或编码α-半乳糖苷酶的基因选自由以下组成的组:agl1、agl2、agl3、agla、aglb、aglc、agld、mel1、mel2、mel5、和mel6。
105.在一个实施方案中,除了导致乳酸脱氢酶和α半乳糖苷酶酶活性增加的遗传修饰以外,本发明的真菌可还包含一个或多个遗传修饰。在一个实施方案中,真菌进一步遗传修饰来降低乙醇生产。在具体实施方案中,真菌被遗传修饰以便通过修饰或缺失与乙醇生产相关的基因的至少一部分或通过灭活与乙醇生产相关的基因来降低乙醇生产。任选地与乙醇生产相关的一种或多种基因选自由以下组成的组:pdc1、pdc5、pdc6、adh1、adh2、adh3、adh4、和adh5、和其任何组合。在一个具体实施例中,pdc1和adh1已缺失或修饰。在另一个具体实施方案中,pdc1和pdc5已缺失或修饰。在一个非常具体实施方案中,pdc1;pdc1和adh1;pdc1和pdc5;adh1和pdc5;或pdc5中的一或多个等位基因已缺失或修饰。
106.如在此使用,pdc基因是指编码丙酮酸酯脱羧酶的基因,所述酶催化将丙酮酸酯降解成乙醛和二氧化碳。至少pdc1、pdc5、和pdc6编码丙酮酸酯脱羧酶的不同同功酶。丙酮酸
酯脱羧酶分类为ec 4.1.1.1。所有同功酶、同种型和变体包括在pdc的范围内。
107.如在此使用,adh是指编码醇脱氢酶的基因,所述酶催化将乙醛转化成乙醇。酵母和大多数细菌使碳源物诸如葡萄糖发酵成乙醇和co2。事实上,由糖酵解所产生的丙酮酸酯被转化成乙醛和二氧化碳,并且然后通过醇脱氢酶使乙醛还原成乙醇。至少adh1、adh2、adh3、adh4、和adh5编码醇脱氢酶的不同同功酶。醇脱氢酶分类为ec 1.1.1.1。所有同功酶、同种型和变体包括在adh的范围内。
108.在一个实施方案中,与乙醇生产相关的一种或多种基因被修饰或至少部分地缺失或失活。在另一个实施方案中,不同于与乙醇生产相关的基因的任何其他基因被修饰或至少部分地缺失或失活。在本发明的一个实施方案中,真菌包含减少蛋白质或酶活性的遗传修饰。“降低活性”是指分别与野生型蛋白质或真菌相比,在特定蛋白质或修饰真菌中存在较小活性(如果有的话),或与未修饰细胞或真菌相比,细胞或真菌中的较低活性(如果有的话)。降低活性可由多肽表达的下调、基因表达的下调、基因的至少一部分的缺乏、蛋白质的缺乏和/或蛋白质的降低活性所导致。存在用于减少蛋白质的活性的各种遗传技术并且所述技术是本领域技术人员熟知的。这些技术利用基因的核苷酸序列或基因附近的核苷酸序列。
109.在本发明的具体实施方案中,一或多个蛋白质失活。如在此使用,“失活”是指其中蛋白质的活性完全失活的情况,即细胞没有特定蛋白质的活性。基因可例如通过预防其表达或通过基因或其部分的突变或缺失而失活。在本发明的一个实施方案中,一或多个基因或其任何片段已缺失。在具体实施方案中,真菌通过缺失基因的至少一部分来遗传修饰。如在此使用,“基因的一部分”是指基因或其任何片段的一个或多个核苷酸。举例来说,基因敲除方法适合于缺失编码具有特定活性的多肽的核苷酸序列、其任何部分。
110.pdc和/或adh基因的缺失或修饰可以各种方法实现,包括但不限于同源重组、破坏基因位点、反义分子或存在于细胞中的例如用于减少pdc和/或adh基因的表达的杀伤质粒。
111.在本发明的方法、用途或真菌的一个实施方案中,真菌还包含选自由以下组成的组的一种或多种基因的遗传修饰:cyb2、gpd1、gpd2、gpp1、gpp2和其任何组合。cyb2编码使乳酸酯氧化的l-乳酸酯:细胞色素c氧化还原酶。gpd1、gpp1和gpp2是与甘油生物合成相关的基因。gpd1编码甘油-3-磷酸酯脱氢酶。gpp1和gpp2分别编码甘油-1-磷酸酯磷酸水解酶1和2。
112.本发明的遗传修饰真菌通过执行特定遗传修饰来获得。在一个实施方案中,遗传修饰真菌是重组真菌。如在此使用,“重组真菌”是指被遗传修饰以便与修饰之前的真菌相比,含有不同遗传物质(例如,与修饰之前的真菌相比,包含包括整个基因或其一部分的一种或多种核酸的缺失、取代、破坏或插入)的任何真菌。“重组真菌”另外是指包含所述遗传修饰的宿主细胞。
113.编码已知多肽的多核苷酸可使用常见分子或遗传技术来突变。核酸和氨基酸数据库(例如genbank)可用于识别编码具有酶活性的多肽的多核苷酸序列。序列比对软件诸如blast(蛋白质或核苷酸)可用于比较各种序列。简单地说,与具有酶活性的多肽具有某种同源性的任何氨基酸序列,或与编码具有酶活性的多肽的序列具有某种同源性的任何核酸序列可用作搜索例如genbank的查询。序列的一致性百分比可方便地使用具有默认参数的blast软件来计算。使用具有默认参数的blast算法,具有给定百分比的一致性评分和阳性
评分的序列被认为所述百分比一致或同源的。
114.在本发明的具体实施方案中,用于本发明中的多肽包含与seq id no:1、2、3、4、5、或6具有至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%的序列一致性的序列,或其酶活性片段或变异体。序列id no 1-6是α-半乳糖苷酶的多肽序列。在本发明的具体实施方案中,用于本发明中的多核苷酸包含与seq id no:7、8、9、10、11或12具有至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%的序列一致性的序列,或其活性片段或变异体。序列id no 7-12是α-半乳糖苷酶基因的核苷酸序列。
115.众所周知一个或少许氨基酸的缺失、添加或取代不一定改变酶类蛋白质的催化性质。因此本发明还涵盖具有规定酶活性的给定氨基酸序列的变体和片段。如在此使用的术语“变体”是指如与给定序列相比,具有氨基酸序列的轻微变化的序列。这种变体可例如作为相同菌株、物种或属类内的等位基因变体而自然地发生,或它可通过诱变或其他基因修饰来产生。它可包含氨基酸取代、缺失或插入,但是它仍然以与给定酶大致上相同的方式起作用,具体地说它保留其作为酶的催化功能。
116.给定蛋白质或多肽序列的“片段”意指所述序列的部分,例如在n和/或c末端处截短的序列。它可以例如是包含信号序列的蛋白质的成熟部分,或它可以只是成熟蛋白质的酶活性片段。
117.本发明基于真菌和与其相关方法和用途。各种真菌适合用于本发明中。在一个实施方案中,真菌是酵母或丝状真菌。在具体实施方案中,真菌是选自以下属类的酵母或丝状真菌:曲霉、酵母属、克鲁维斯酵母、毕赤酵母、汉逊酵母、假丝酵母、丝孢酵母属(trichosporon)、根霉、圆孢菌(torulaspora)、伊萨酵母(issatchenkia)和舍弗氏菌(scheffersomyces),例如尤其来自由以下组成的组:酿酒酵母、葡萄汁酵母(s.uvarum)、耐热克鲁维斯酵母、乳酸克鲁维斯酵母、马克斯克鲁维斯酵母、多形汉逊酵母、树干舍弗氏菌(scheffersomyces stipitis)、米根霉、比勒陀利亚圆孢菌(torulaspora pretoriensis)、东方伊萨酵母(issatchenkia orientalis)、发酵毕赤酵母(pichia fermentans)、盔状毕赤酵母(p.galeiformis)、沙漠毕赤酵母(p.deserticola)、膜醭毕赤酵母(p.membranifaciens)、杰丁毕赤酵母(p.jadinii)、库德毕赤酵母(p.kudriavzevii)、异常毕赤酵母(p.anomala)、乙醇假丝酵母(candida ethanolica)、索诺拉假丝酵母(c.sonorensis)和蜜生假丝酵母(c.apicola)。
118.在本发明的方法、用途或真菌的一个实施方案中,真菌在登录号vtt c-191026或vtt c-201040下寄存于vtt保藏。根据用于专利程序目的寄存微生物的国际公认布达佩斯条约,以下菌株寄存于vtt培养物保藏,p.o.box 1000(vuorimiehentie 3),fl-02044vtt,finland:登录号vtt c-191026和登录号vtt c-201040。(关于vtt c-191026,参见实例3的e143-4;关于vtt c-201040,参见实例6。)
119.本发明的遗传修饰真菌可通过本领域技术人员已知的任何遗传方法来制备。所述方法包含至少提供真菌和遗传修饰真菌来增加乳酸脱氢酶和α-半乳糖苷酶活性。真菌或真菌细胞的遗传修饰在一或多个步骤中通过设计和构造合适载体和用所述载体来转化真菌细胞来实现。可使用电穿孔和/或化学(诸如基于氯化钙或乙酸锂)转化方法。转化真菌细胞的方法在本领域技术人员的知识范围内。可能遗传修饰的实例如上在本公开中描述。在一
个实施方案中,将编码一或多个异源酶的一或多个多核苷酸添加至真菌或真菌细胞,并且任选地将编码一或多个内源酶的一或多个多核苷酸修饰(例如通过一或多个核苷酸的插入、缺失或取代)以便增加或降低所述真菌中的所述酶的活性。编码多肽的多核苷酸序列或多肽序列的知识可用于遗传修饰合适真菌。
120.当真菌存在于包含低聚半乳糖的发酵培养基中时,本发明的遗传修饰真菌能够水解来自各种α-半乳糖苷的非还原末端半乳糖基残基,消耗丙酮酸酯和生产乳酸和/或乳酸酯。在一个非常具体实施方案中,所述真菌可以高生产力和产率来生产l-乳酸。在一个实施方案中,本发明的真菌耐受高乳酸浓度。在一个非常具体实施方案中,真菌是被修饰以便最小程度地生产天然发酵产物乙醇并且替代地生产乳酸的酸耐受性真菌。
121.在本发明的一个实施方案中,真菌具有增加的乳酸生产。生产乳酸的方法可导致约30、40、50、60、70、80、90、100、110、120、或130克/l或更大的乳酸滴度和/或约0.5、1.0、1.5、2.0、2.5、3.0g l-1
h-1
或更大的乳酸生产力。
122.在一个实施方案中,本发明的真菌具有极优异性能,以超过80%产率(即,g有机产物/g消耗碳源)、超过2g l-1
h-1
生产率来转化糖(例如大豆糖蜜糖)并且达到高滴度(多达129g/l乳酸)。
123.生产乳酸酯的方法可导致约30、40、50、60、70、80、90、100、110、120、或130克/l或更大的乳酸酯滴度和/或约0.5、1.0、1.5、2.0、2.5、3.0g l-1
h-1
或更大的乳酸酯生产力。
124.检测乳酸、乳酸酯和/或低聚半乳糖的方法是本领域技术人员熟知的。举例来说,可使用色谱方法诸如hplc和离子色谱。乳酸酯的存在可确定例如如witte等人(1989,j.basic microbiol.29:707-716)所描述。
125.本发明的真菌可用于生产乳酸和/或乳酸酯或生产聚合物,任选地聚酯或聚乳酸。
126.生产一种或多种选自由聚合物、聚酯和聚乳酸组成的组的产物的本发明方法包括在含有碳源物(例如低聚半乳糖)的培养基中培养本发明的遗传修饰真菌以便生产乳酸,回收所得乳酸和使用回收的乳酸来生产聚合物、聚酯和/或聚乳酸。聚合物的生产是本领域技术人员熟知的方法,包括但不限于例如乳酸的聚合。
127.在本公开中,术语“多肽”和“蛋白质”可互换使用来意指任何长度的氨基酸的聚合物。如在此使用,“酶”是指能够加速或催化化学反应的蛋白质或多肽。
128.如在此使用,“多核苷酸”是指包含编码所讨论的多肽的核酸序列或其保守序列变异体的任何多核苷酸,诸如单链或双链dna(基因组dna或cdna)或rna。保守核苷酸序列变体(即,不显著改变所编码的多肽的生物性质的核苷酸序列修饰)包括由遗传密码的简并和沉默突变所产生的变体。
129.本领域技术人员显而易知,随着技术进步,本发明概念可以不同的方式实施。本发明和其实施方案不限于以下描述的实施例而是可在权利要求的范围内变化。
实施例
130.实施例1-半乳糖上的不同真菌物种的生长
131.在摇瓶培养中研究多个野生型和ldh表达真菌菌株在半乳糖上的生长。菌株在具有10ml sc培养基、酵母氮源和20g/l半乳糖作为碳源的50ml锥形瓶中培养。菌株的生长通过在培养过程期间量化光密度(od
600
)来评估。在野生型菌株(图1)之中,所有马克斯克鲁维
酵母菌株能够在半乳糖上生长,而两种测试蜜生假丝酵母菌株都不展示可证明的生长。在表达编码l-乳酸脱氢酶的瑞士乳杆菌ldhl的菌株之中,仅酿酒酵母h5037(从野生型菌株c-02453衍生)生长较好,而属于毕赤酵母属、杰丁毕赤酵母、库德毕赤酵母、或异常毕赤酵母的菌株中没有一个能够在此糖上生长(图2)。最后,在真菌或酵母物种之间,存在其利用半乳糖作为碳源的能力的显著变化。
132.实施例2-真菌中的α-半乳糖苷酶活性的展示
133.酿酒酵母菌株vtt-c-02453从vtt培养物保藏接收。所有其他菌株是vtt-c-02453的后代。
134.酿酒酵母vtt-c-02453的尿苷营养缺陷型衍生物通过将ura3基因的蛋白质编码区域替换成赋予潮霉素抗性的hph基因来构建。hph表达盒侧接有loxp位点以便促进通过cre重组酶来进行标志物切除。两种ura3等位基因在二倍体宿主中缺失。
135.对于α-半乳糖苷酶的多拷贝游离基因表达,酿酒酵母mel5基因(genbank登录号z37511)通过来自质粒pmlv18(pmel5-39衍生物,naumov等人1990.mol gen genet224:119-128;turakainen等人1994yeast 10:1559-1568)使用引物32mel5-atg-f(seq id no:13)和33mel5-stopr(seq id no:14)的pcr来扩增,用ecori和asci消化,并且在酿酒酵母eno1启动子与终止子之间克隆至pmi529中(ilm
é
n等人2011biotech for biofuels 4:30),从而产生pmie-005。通过genscript(usa),其他α-半乳糖苷酶编码基因的蛋白质编码区域得以合成并且优化以便在酿酒酵母中表达,并且pmie-5中的mel5基因通过合成基因来替换,产生质粒pmie-16(黑曲霉aglc;q9uuz4)(seq id no:15)、pmie-17(里氏木霉agl1;q92456)(seq id no:16)、pmie-18(米黑根毛霉菌gal36;h8y263)(seq id no:17)、pmie-19(赤霉菌属f75 gal36;c6fjg8)(seq id no:18)、和pmie-20(费氏曲霉gal27b;aja29661.1)(seq id no:19)。
136.使用乙酸锂方法(gietz等人1992nucleic acids res.20:1425.),vtt-c-02453ura3δ/ura3δ用ura3可选择α-半乳糖苷酶表达载体pmie-5(酿酒酵母mel5)(seq id no:20)、pmie-16(黑曲霉aglc)、pmie-17(里氏木霉agl1)、pmie-18(米黑根毛霉菌gal36;h8y263)、pmie-19(赤霉菌属f75 gal36;c6fjg8)、或pmie-20(费氏曲霉gal27b;aja29661.1)中的每一个来转化。转化株在scd-ura培养基上选择,基于补充有5-溴基-4-氯基-3-吲哚基-α-d-吡喃半乳糖苷(α-x-gal)的琼脂板上的菌落的蓝色的形成,观察到α-半乳糖苷酶活性。
137.α-x-gal上的α-半乳糖苷酶基因活性在表达α-半乳糖苷酶的每个酵母转化株中观察到(数据未展示)。α-x-gal阳性转化株在含有1%蜜二糖或蜜三糖作为唯一碳源的液体sc-ura-培养基中生长的能力在30℃下在220rpm振荡下在24孔板上在4ml o/n培养物中测试。包括含有功能ura3基因的亲本菌株作为阴性对照。表达酿酒酵母、黑曲霉、赤霉菌属、或费氏曲霉的α-半乳糖苷酶的转化株在蜜二糖上生长较好至8至12的od
600
,而缺少α-半乳糖苷酶的亲本菌株和带有里氏木霉或米黑根毛霉菌α-半乳糖苷酶基因的转化株的od
600
具有低于1的od
600
(图3)。相比之下,在蜜三糖上的生长不单独取决于α-半乳糖苷酶,因为转化酶将蜜三糖裂解成果糖和蜜二糖,并且果糖可被亲本菌株消耗。
138.pmie-5(酿酒酵母mel5)、pmie-16(黑曲霉aglc)、pmie-17(里氏木霉agl1)、pmie-19(赤霉菌属f75 gal36;c6fjg8)、和pmie-20(费氏曲霉gal27b;aja29661.1)转化株(参见
实例2)在24孔板上在1:3稀释大豆糖蜜中以4ml培养24小时以便证明菌株将不同糖转化成乙醇的能力。过滤样品在35℃,0.3ml/min流量的5mm h2so4下在aminex hpx-87h管柱(bio rad)上运作以便量化所生产乙醇和残留糖。所述方法不区分三糖(蜜三糖/甘露三糖)或双糖(蔗糖,蜜二糖),并且不将果糖与半乳糖分离。当表达酿酒酵母mel5、黑曲霉aglc、赤霉菌属f75 gal36或费氏曲霉gal27b时,相对于亲本菌株vtt-c-02453ura3δ/ura3δ,乙醇生产大大增加(图4)。通过这些菌株的大豆糖蜜低聚半乳糖(gos)的消耗从hplc结果中也是明显的(图5)。亲本菌株和表达里氏木霉agl1的菌株展示显著残留二和三糖,而这些对于表达酿酒酵母mel5、黑曲霉aglc、赤霉菌属f75 gal36或费氏曲霉gal27b的菌株来说是不明显的。
139.实施例3-构建表达ldh和不同α-半乳糖苷酶的真菌
140.vtt-c-02453中的adh1基因通过将编码区域替换成pcr产物来缺失,所述pcr产物包含侧接有loxp位点的kanmx遗传霉素抗性盒,其使用引物2scadh1-150f(seq id no:21)和5scadh1stopr(seq id no:22)从pug6(=b901)扩增,获得缺失构建体2 5-scadh1。
141.为了将不同α-半乳糖苷酶表达盒整合至酿酒酵母can1基因座中,pmie-5、pmie-16、pmie-19、pmie-20用smal和swal消化,去磷酸化,并且将含有α-半乳糖苷酶的片段连接至含有kanmx盒和can1同源性区域的b3033=pmi-503的5177bp mscl-ecorv片段,分别产生pmie-21b(seq id no:23)、pmie-24b(seq id no:24)、pmie-25b(seq id no:25)、pmie-26a(seq id no:26)。
142.为了将编码l-乳酸脱氢酶的瑞士乳杆菌ldhl整合至pdc1基因座中,构建表达载体pmie-8(seq id no:33)。它含有酿酒酵母pgk1启动子与adh1终止子之间的瑞士乳杆菌ldhl和棉蚜(a.gossypii)tef1启动子与终止子之间的赋予潮霉素抗性的大肠杆菌hph基因,围绕有用于标志物切除的loxp位点,和促进在pdc1基因座中同源重组的pdc1的5'和3'区域。
143.对于标志物切除,将cre重组酶在来自诺尔丝菌素可选择着丝粒载体cre-nat的gal1启动子下表达。
144.酿酒酵母使用peg-乙酸锂方法(gietz等人1992nucleic acids res.20:1425)来转化。转化株在视情况补充有200μg/ml潮霉素、300μg/ml遗传霉素、或200μg/ml诺尔丝菌素的琼脂固化ypd培养基中选择。
145.vtt-c-02453用pmie-8转化并且分离潮霉素抗性转化株e16。潮霉素抗性标志物通过将cre-重组酶表达载体psk-70转化至e16中来切除并且分离诺尔丝菌素抗性转化株e23。e23用pmie-8转化并且分离潮霉素抗性转化株e51-6。pcr分析指示pdc1编码区域不存在于e51-6中。e51-6用adh1缺失盒转化并且分离g418抗性转化株e79-4、e79-5、e79-9和e79-10。pcr分析指示adh1编码区域存在于e79-5、e79-9和e79-10中但是不存在于e79-4中,表明两种adh1等位基因在e79-4中缺失。根据此情况,与e79-5、e79-9和e79-10相比,e79-4形成较小菌落。抗性标志物通过将cre-重组酶表达载体psk-70转化至e79-4中来切除并且分离诺尔丝菌素抗性转化株。
146.转化株e79-4的无标记衍生物用sacll-scal消化pmie-24b、pmie-25b、和pmie-26a转化,以便分别表达黑曲霉、赤霉菌属、和费氏曲霉的α-半乳糖苷酶基因。靶向α-半乳糖苷酶基因以便整合至can1基因座中。转化株基于遗传霉素抗性来选择,基于补充有α-x-gal的琼脂板上的菌落的蓝色的形成,观察到α-半乳糖苷酶活性。
147.菌株e142-1、e143-4(vtt c-191026)和e144-4分别表达黑曲霉、赤霉菌属f75和费
no:32)和6scpdc5stopr(seq id no:31)和pug6质粒作为模板的pcr来制备,并且分离g418抗性转化株e82。转化株中pdc5编码区域的缺乏通过pcr来验证。
156.并行地,e22用pdc5缺失盒转化并且分离g418抗性株。pcr分析指示pdc5编码区域不存在于转化株e78-1中,表明两种pdc5等位基因在e78-1中缺失。e78-1用notl消化pmie-008转化以便缺失其余pdc1等位基因并且分离潮霉素抗性转化株。转化株e94中pdc1编码区域的缺乏通过pcr来验证。
157.缺失pdc1和pdc5的两个拷贝的转化株e82和e94用cre-重组酶表达载体psk-70转化以便切除kanmx和潮霉素抗性标志物。转化株e82和e94的无标记衍生物用sacll-scal消化pmie-24b、pmie-25b、和pmie-26a转化,以便分别表达黑曲霉、赤霉菌属、和费氏曲霉的α-半乳糖苷酶基因。靶向α-半乳糖苷酶基因以便整合至can1基因座中。转化株基于遗传霉素抗性来选择,基于补充有α-x-gal的琼脂板上的菌落的蓝色的形成,观察到α-半乳糖苷酶活性。乳酸的生产在如实施例3所描述的使用大豆糖蜜作为唯一碳源的生物反应器中展示。
158.图8示出实施例1-4中描述或提及的质粒的图谱。
159.实施例5-通过表达一种以上α-半乳糖苷酶的菌株来生产乳酸
160.菌株vtt c-191026(e143-4,参见实施例3)和含有另外α-半乳糖苷酶基因的三个菌株在摇瓶中使用大豆糖蜜作为碳源来培养。三个菌株包含赤霉菌属f75 gal36,或黑曲霉aglc或费氏曲霉gal27b的另外拷贝,如实施例3所描述。不同菌株的预培养物在30℃下在ypd培养基中生长过夜。细胞通过离心来收获并且重新悬浮于ro-h2o中以便给出20的od
600
值。大豆糖蜜用ro-h2o来稀释至其原始浓度三分之一并且使用标准高压釜液体循环(121℃,20min)来灭菌。将50毫升的这份经灭菌、稀释的大豆糖蜜添加至250ml锥形烧瓶,所述烧瓶用2.5g caco3使用干燥循环(160℃,3h)来预先灭菌。将500微升的细胞悬浮液用于接种每个培养瓶,以获得对应于大约0.2的od
600
值的原始细胞密度。
161.伴以200rpm搅动,将烧瓶在30℃下保持在振荡培养器中,并且将样品定期取出。将样品离心并且所得上清液浸没于沸水浴中10分钟。沸腾之后,将样品再次离心,并且所得上清液在hplc洗脱剂(5mm h2so4)中稀释10倍。样品在55℃和0.5ml流量下在aminex hpx-84h管柱(bio rad)上运作。水苏糖用作四糖的标准,而麦芽三糖和麦芽糖分别用作三和二糖的标准。所获得的结果在图9中给出并且表明与vtt c-191026相比,α-半乳糖苷酶基因的另外拷贝可进一步增强大豆糖蜜低聚半乳糖的水解速率。
162.实施例6-通过替代酵母库德毕赤酵母来生产乳酸
163.为了证明在除了酿酒酵母以外的酵母中表达α-半乳糖苷酶和乳酸脱氢酶也可导致较高水平地从大豆糖蜜生产乳酸,合适菌株(vtt c-201040)从库德毕赤酵母vtt-c-79090产生。因为酵母天然地不能够水解蔗糖,所以需要转化酶上的另外表达。
164.为了将编码l-乳酸脱氢酶的瑞士乳杆菌ldhl整合至pdc1基因座中,构建表达载体pmipk124(seq id no:36,图11)。它含有库德毕赤酵母pgk1启动子与酿酒酵母adh1终止子之间的瑞士乳杆菌ldhl和库德毕赤酵母pgk1启动子与酿酒酵母mel5终止子之间的赋予潮霉素抗性的大肠杆菌hph基因,围绕有用于标志物切除的loxp位点,和促进在pdc1基因座中同源重组的库德毕赤酵母pdc1的5'和3'区域。通过notl消化,将表达盒从载体序列释放。库德毕赤酵母使用peg-乙酸锂方法(gietz等人1992nucleic acids res.20:1425)来转化。转化株在视情况补充有500μg/ml潮霉素、或200μg/ml诺尔丝菌素的琼脂固化ypd培养基中选
择。潮霉素抗性标志物通过将cre-重组酶表达载体pklnatcrelopgk转化至中来切除并且分离诺尔丝菌素抗性转化株。pklnatcrelopgk通过在非选择性培养基上生长细胞来移除,导致分离菌株h4927。h4927再次用pmipk124转化以便将二倍体基因组中的两个pdc1等位基因用ldhl表达载体替换,并且分离h4948。
165.以如上所述类似的方式,用cre-重组酶将潮霉素抗性标志物从菌株h4948中移除并且所获得的菌株被命名为h5661。h5661是用于将转化酶和α-半乳糖苷酶整合至adh1基因座中的亲代菌株。构建两种表达载体pekopa8(seq id no:37,图11)和pekopa9(seq id no:38,图11),其含有编码转化酶(seq id no:34)的酿酒酵母suc2(seq id no:35)以及各自编码α-半乳糖苷酶的赤霉菌gibgal36(pekopa8)或黑曲霉aglc(pekopa9),以及促进在adh1基因座中同源重组的库德毕赤酵母adh1的5'和3'区域。将两个表达盒从载体中释放以便用notl限制酶转化。表达转化酶和α-半乳糖苷酶的转化株在补充有20g/l d( )-蔗糖和40μg/mlα-x-gal的琼脂固化yp培养基中选择。
166.为了表明从大豆糖蜜生产乳酸,与vtt c-191026并行地,将表达转化酶和赤霉菌gibgal36α-半乳糖苷酶的库德毕赤酵母菌株vtt-c-201040在使用大豆糖蜜作为碳源的摇瓶中培养。培养条件与实施例5所描述相同。如以前实施例所描述,量化来自培养样品的所产生的乳酸和残留低聚糖,并且结果在图10中给出。对于两种菌株,获得可比较的乳酸生产水平。结果指示来自大豆糖蜜的高水平的乳酸生产可使用具有类似遗传修饰的另一种酵母菌株来实现。
167.图11示出实施例6中描述或提及的质粒的图谱。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。