小麦斑点叶基因lm5共分离的kasp分子标记及其应用
技术领域
1.本发明涉及分子遗传学领域,特别是涉及一种小麦斑点叶基因lm5共分离的kas p分子标记及其应用。
背景技术:
2.普通小麦(triticum aestivum l.)具有悠久的种植历史,并且其播种面积占我国耕地面积的20%
‑
30%,因此小麦在我国农作物中占据重要地位。粮食作物的生长发育离不开叶片为其提供的营养物质。叶片作为绿色植物进行光合作用的重要器官,里边进行着许多重要的生理生化反应。随着植物生理学的不断发展,光合作用、光合色素合成或降解、光形态建成、植物抗病性机制和细胞程序性死亡等与植物叶片都有着密切联系。叶绿体是植物进行光合作用的重要细胞器,叶绿体中的蛋白酶对叶绿体的发育和光形态建成等具有极其重要的作用。
3.叶色突变是一种在各种植物中广泛存的突变类型,它的种类有很多,大致可将其分为五类,分别是白化、黄化、浅绿、条纹和斑点。类病斑突变体又称为斑点叶突变体,它是指在无病原菌侵染、无机械损伤、无农药伤害及无逆境的环境条件下出现病斑表型的一种特殊的叶色突变类型。这些叶色突变体,不仅使叶绿体的超微结构发生变化,降低叶绿素的含量,色素蛋白复合体遭到破坏还阻碍了叶绿体
‑
细胞核之间的信号转导,严重影响到叶绿素的光合作用以及小麦的产量。因此,斑点叶突变体不仅在研究植物抗病防卫机制和细胞程序性死亡方面发挥着重要作用,而且对探索斑点叶突变体的分子机制、丰富类病斑突变体的基因及其功能,以及为小麦作物育种提供分子标记方面具有重大意义。
4.目前对于小麦斑点叶的研究还不是很深入,但是水稻中对斑点叶的研究比较深入,而且也有研究表明,水稻中斑点叶的产生可以增强植株对病原菌的广谱抗性。大麦中,mlo基因对白粉病的抗性已经广泛应用于大麦育种中。关于小麦斑点叶性状的研究报道还较少,但同为禾本科植物,小麦斑点叶在提高作物对病原菌的抗性方面也极有可能具有相似的作用。
5.bsr
‑
seq(bsa结合转录组测序),此方法在分离群体中选择极端性状的个体构建两个池,提取两个池的总rna,进行转录组测序,测序层数根据混池个数和物种的基因组大小确定。然后用经典贝叶斯算法分析转录组数据,开发snp标记,预测候选基因。
6.分子标记辅助育种,不依赖于表型选择,即不受环境、基因互作、基因与环境互作等因素的影响,而是直接对基因型进行选择,因而能大大提高育种效率。单核苷酸多态性(snp)指的是基因组内dna某一特定核苷酸位置上存在转换、颠换、插入、缺失等变化而引起的dna序列多态性。其技术是利用己知序列信息来比对寻找snp位点,再利用发掘的变异位点设计特异性的引物来对基因组dna或cdna进行pcr扩增,得到基于snp位点的特定的多态性产物,最后利用电泳技术分析产物的多态性。snp标记的优点是数量多,分布广泛;在单个基因和整个基因组中分布不均匀;snp等位基因频率容易估计。
7.竞争性等位基因特异性pcr技术(kompetitive allele specific pcr,kasp)是由
lgc公司(laboratory of the government chemist)(http://www.lgcgenomics.com)研发的低成本、高通量特点的新型基因分型技术,通过引物末端碱基的特异匹配来对sn p以及indel位点进行精准的双等位基因分型,在水稻、小麦、大豆等作物的分子标记辅助选择中得到广泛应用。
8.此前部分学者对斑点叶进行了基因定位,发现斑点叶基因在抗病育种中具有重要的潜在应用价值。然而目前与小麦斑点叶性状相关且可用于实际分子育种的紧密连锁的分子标记却并不多。因此研究获得有关斑点叶的qtl或基因,利用分子生物学技术,进而提高作物光合作用,筛选抗性优良的品种,最终达到选育增产小麦新品种的目的,在小麦育种工作中意义重大。
技术实现要素:
9.本发明的目的是提供一种小麦斑点叶基因lm5共分离的kasp分子标记及其应用,以解决上述现有技术存在的问题,该分子标记kasp
‑
sicau11与斑点叶基因lm5极显著相关,呈现紧密连锁标记特征,用于分子标记辅助选择的准确性高,可显著提高不同遗传背景中筛选含有lm5抗性材料的选择鉴定效率,且成功率高。
10.为实现上述目的,本发明提供了如下方案:
11.基于以上目的,申请人利用斑点叶突变体材料mc21为母本,以小麦品系3642为父本杂交,得到杂种f1,f1代单株自交获得f2,f2单株全部收获,每个株系单独播种一行,获得含有313个株系的f
2:3
家系,构成遗传作图群体。在作图群体中选取极端斑点叶和绿色叶材料进行rna混池测序(bsr
‑
seq)。对f
2:3
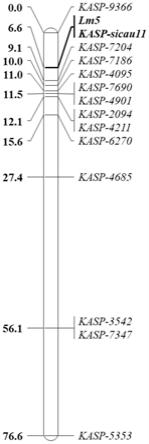
家系群体叶片颜色进行鉴定,提取亲本mc21、3642和f2代群体株系的dna,筛选亲本和极端混池之间的多态性snp标记。根据筛选的snp标记,利用joinmap4.0构建遗传图谱。结合群体的斑点叶表型数据定位斑点叶基因lm5,计算lm5基因的位置和分子标记之间的遗传距离,发现mc21突变体材料2a染色体长臂上定位到一个斑点叶基因,经过基因克隆开发了一个与斑点叶基因lm5共分离的kasp标记。
12.本发明提供一种小麦斑点叶基因lm5在调控小麦叶色颜色性状中的应用,所述小麦斑点叶基因lm5位于小麦2a染色体长臂上。
13.本发明还提供一种小麦斑点叶基因的kasp分子标记kasp
‑
sicau11,所述kasp分子标记kasp
‑
sicau11为snp分子标记,多态性为c/t,其与小麦斑点叶基因lm5共分离于2a染色体长臂。
14.进一步地,所述kasp分子标记kasp
‑
sicau11可通过如seq id no.31
‑
33所示的引物组合扩增得到。
15.进一步地,seq id no.31
‑
32所示的引物序列在5’端分别添加不同的荧光修饰基团,或在3’端分别添加不同的荧光修饰基团。
16.进一步地,荧光修饰基团包括但不限于fifc、fam、tet、hex、joe、tamra或bhq。
17.本发明还提供一种引物组合物,含有如seq id no.31
‑
32所示的两条特异性引物和如seq id no.33所示的通用引物。
18.本发明还提供一种含有上述的引物组合物的试剂盒。
19.本发明还提供一种上述的kasp分子标记kasp
‑
sicau11、上述的引物组合物或上述的试剂盒在作物分子育种、培育转基因小麦或小麦种质资源改良中的应用。
20.本发明还提供一种上述的kasp分子标记kasp
‑
sicau11、上述的引物组合物或上述的试剂盒在培育斑点叶性状的小麦品种或品系、或在小麦特异叶片颜色材料创制中的应用。
21.本发明还提供一种鉴定小麦斑点叶基因lm5的分子标记的方法,以待鉴定材料的dna为模板,用上述的引物组合物进行荧光定量pcr扩增,根据扩增产物进行基因型分型,判断小麦材料中是否有斑点叶基因lm5。
22.具体地,所述荧光定量pcr的反应体系为:5μl master mix、1.4μl混合引物、5ng模板dna、双蒸水加至总量为10μl。其中,所述混合引物是由seq id no.31
‑
seq id no.33所示的三条引物按照10ng/μl的浓度分别加入120μl,120μl和300μl,并添加ddh2o 460μl进行混合而成。
23.进一步的,所述荧光定量pcr至少有3个独立的以双蒸水代替dna模板的空白。
24.进一步的,所述荧光定量pcr的反应程序为:94℃预变性15min;94℃变性20s、65℃复性/延伸60s,共10个循环;94℃变性20s、55℃复性/延伸60s,共30个循环。
25.本发明公开了以下技术效果:
26.本发明首次公开了来自小麦突变体mc21的斑点叶基因lm5,位于小麦2a染色体长臂上,含有该基因的材料可以显著提高小麦的抗条锈病和白粉病能力。该基因在小麦产量(调控叶片)育种中具有较高的利用价值。本发明公开了基于荧光定量pcr平台精确检测小麦mc21的斑点叶基因lm5的分子标记kasp
‑
sicau11,且为共显性标记,检测准确高效、扩增方便稳定。本发明公开的分子标记kasp
‑
sicau11与斑点叶基因lm5极显著相关,呈现紧密连锁标记特征,用于分子标记辅助选择的准确性高,可显著提高在不同遗传背景中筛选含有lm5抗性材料的选择鉴定效率,且成功率高。
27.本发明公开了位于小麦2a染色体上的与小麦斑点叶连锁的分子标记kasp
‑
sicau11,该分子标记是小麦2a染色体长臂上斑点叶lm5基因的共分离标记,连锁度高。该标记可用来检测小麦2a染色体上的斑点叶基因,快速筛选具有该位点的植株,进而方便进行高产小麦的分子辅助育种。本发明提供的分子标记kasp
‑
sicau11与小麦2a上的斑点叶基因lm5紧密连锁,可用来对小麦斑点叶这一性状进行定位,从而在育种过程中筛选含有斑点叶的抗性植株,提高育种工作效率,并为小麦斑点叶基因的研究提供基础。
附图说明
28.为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
29.图1为小麦斑点叶基因lm5在2a染色体上的定位;
30.图2为mc21
×
3642的f2代株系中分子标记kasp
‑
sicau11检测的荧光读值结果;其中,fam(正方形,“3642”)荧光为有正常叶片的株系,hax(圆形,“mc21”)荧光为斑点叶片的株系;三角形荧光为杂合株系;菱形荧光为空白对照;
31.图3为小麦斑点叶突变体mc21
×
小麦品种cn16的株系中分子标记kasp
‑
sicau11检测的荧光读值结果;其中,fam(正方形,“cn16”)荧光为正常叶片的株系,hax(圆形,“mc21”)
荧光为斑点叶片的株系,三角形荧光为杂合株系;菱形荧光为空白对照。
具体实施方式
32.现详细说明本发明的多种示例性实施方式,该详细说明不应认为是对本发明的限制,而应理解为是对本发明的某些方面、特性和实施方案的更详细的描述。
33.应理解本发明中所述的术语仅仅是为描述特别的实施方式,并非用于限制本发明。另外,对于本发明中的数值范围,应理解为还具体公开了该范围的上限和下限之间的每个中间值。在任何陈述值或陈述范围内的中间值以及任何其他陈述值或在所述范围内的中间值之间的每个较小的范围也包括在本发明内。这些较小范围的上限和下限可独立地包括或排除在范围内。
34.除非另有说明,否则本文使用的所有技术和科学术语具有本发明所述领域的常规技术人员通常理解的相同含义。虽然本发明仅描述了优选的方法和材料,但是在本发明的实施或测试中也可以使用与本文所述相似或等同的任何方法和材料。本说明书中提到的所有文献通过引用并入,用以公开和描述与所述文献相关的方法和/或材料。在与任何并入的文献冲突时,以本说明书的内容为准。
35.在不背离本发明的范围或精神的情况下,可对本发明说明书的具体实施方式做多种改进和变化,这对本领域技术人员而言是显而易见的。由本发明的说明书得到的其他实施方式对技术人员而言是显而易见得的。本发明说明书和实施例仅是示例性的。
36.关于本文中所使用的“包含”、“包括”、“具有”、“含有”等等,均为开放性的用语,即意指包含但不限于。
37.实施例1小麦斑点叶基因lm5及其分子标记kasp
‑
sicau11的获得
38.(1)利用斑点叶突变体材料mc21,以小麦品系3642为父本杂交,得到杂种f1,f1代单株自交获得f2,获得含有313个植株的f
2:3
家系,构成遗传作图群体。
39.(2)f
2:3
家系群体斑点叶表型鉴定:小麦拔节期对每个株系的叶片颜色进行分析鉴定,整行植株的叶片全部为绿色的记为“a”,整行植株的叶片全部为斑点叶的记为“b”,整行植株的叶片中既有斑点又有绿色的记为“h”。每一个株系逐一进行判定,判定结果记为斑点叶基因lm5的表型标记数据。这样在连锁图上定位到的位置就是lm5基因所在位置。
40.(3)bsr
‑
seq分析
41.a)dna提取:用ctab法提取亲本mc21、3642和f2群体植株dna。
42.b)采用极端混池分离分析法,在mc21
×
3642群体中分别选取极端的新鲜斑点叶和绿色叶材料各30株混合构建斑点池和正常池,提取rna。bsr
‑
seq测序由诺禾致源公司(https://www.novogene.com/)完成。亲本3642的带型记为“a”,亲本mc21的带型记为“b”,杂合f1记为“h”;群体材料中,带型来源于3642的记为“a”,来源于mc21的记为“b”,杂合类型的记为“h”,缺失的记为
“‑”
。
43.c)亲本混池之间多态性标记筛选:根据bse
‑
seq测序数据选取亲本和混池之间有纯合差异的snp,转化成kasp标记,进行荧光定量pcr基因分型。
44.d)mc21
×
3642的f
2:3
家系群体分析:以上步骤获得的多态性kasp标记为引物,利用每对引物同时扩增亲本3642、mc21和f
2:3
家系群体的dna,进行基因型鉴定,获得分子标记数据。亲本3642的基因型记为“a”,亲本mc21的基因型记为“b”,杂交f1的基因型记为“h”,群体
材料中,带型来源于3642的记为“a”,来源于mc21的记为“b”,杂合类型的记为“h”。
45.e)连锁图谱的构建:根据开发的标记,利用joinmap4.0构建遗传图谱。以lod值3
‑
10依次构建连锁群,寻找最优的标记数和标记顺序,确定连锁群及顺序。利用软件mapqtl6.0的区间作图模型和多qtl作图模型,结合群体的斑点叶表型数据定位斑点叶基因lm5,并计算lm5基因的位置和分子标记之间的遗传距离,发现斑点叶突变体mc21染色体2a染色体长臂上存在一个斑点叶基因lm5。
46.f)遗传图谱的密化和紧密连锁分子标记的获得:前人报道与斑点叶相关的基因较少。例如:受隐形基因控制的lm基因被定位到1bl染色体上,含此位点的qtl可以解释60.8%的抗叶锈病表型变异。隐形的类病斑基因lm1和lm2分别定位在3bs和4al染色体上。受光照强度和光照长度影响的显性基因lm3被定位到3bl染色体。最近研究表明,位于2ds染色体上的显性类病斑基因lm4可以显著性提高小麦的抗条锈病能力。此外,在6a染色体和1b染色体上也存在与类病斑相关的基因。
47.为了密化遗传图谱并获得与斑点叶基因lm5紧密连锁的分子标记,利用bsr
‑
seq数据定位结果对侧翼标记进行物理定位并筛选位于区间内的基因。开发获得高效的ka sp分子标记,需要如下多个步骤:
48.(i)设计扩增特定小麦基因型目标同源染色体(2a)候选基因序列的引物。虽然目前已有六倍体小麦“中国春”参考基因组,但是因为小麦进化过程中可能有染色体结构变异(ma j,stiller j,wei y,zheng y
‑
l,devos km,j,liu c(2014)extensive pericentric rearrangements in the bread wheat(triticum aestivum l.)genotype“chinese spring”revealed from chromosome shotgun sequence data.genome biol evol 6:3039
‑
3048),不同小麦基因型的基因序列结构和多态性位点差异可能较大,要高效、快速、低成本地获得特定小麦基因型的基因序列,最简单的方法便是同源序列克隆。而因为小麦具有三个部分同源染色体组,要分离获得特异于其中某个同源染色体上的序列具有很大难度(bagge m,xia x,l
ü
bberstedt t(2007)functional markers in wheat.curr opin plant biol 10:211
‑
216),需要通过设计特异于某个染色体组上的引物。在熟练掌握比较基因组学技术、生物信息学技术的基础上,对六倍体小麦的供体二倍体亲本乌拉尔图小麦和节节麦、四倍体野生二粒小麦和六倍体“中国春”等参考基因组进行序列截取、比对、分析,获得特异于某个染色体上的多态性位点,从而设计扩增目标区域的特异引物。在设计好引物后,需依靠比较基因组学技术对引物特异性、退火温度、扩增长度等进一步分析,从而确定引物的可用性。
49.(ii)利用特异引物对目标小麦基因型进行扩增。在进行扩增时,需要凭借熟练的分子生物学技术对扩增条件进行优化,进一步克隆测序,以获得目标区域的基因序列。
50.(iii)目标同源染色体候选基因序列多态性位点的获得。在获得两个亲本候选基因序列后,需要进一步对序列进行详细分析,检测是否有多态性位点,如果没有,则需要重新回到第i步,选择其他可能的候选区域进行分离克隆。
51.(iv)多态性位点上下游kasp引物的设计。在获得多态性位点之后,需要设计kasp特异引物。由如前所述,小麦为异源六倍体植物,则仍然要凭借熟练的生物信息学技术对abd染色体序列进行分析,从而获得特异于目标染色体的kasp引物。
52.(v)kasp引物扩增条件的优化。引物合成之后,需要凭借经验进一步对引物扩增条
件进行调试优化,以便能达到在亲本之间能够显著区分的效果。
53.综上所述,虽然kasp标记技术已经广泛应用于二倍体物种,但是要想在六倍体小麦中获得高效的kasp标记时,对本领域技术人员而言绝非易事。
54.最终经过多次克隆测序,引物设计和扩增,共设计kasp引物15对(表1),最终得到标记kasp
‑
sicau11与斑点叶基因lm5共分离。
55.表1 15对kasp引物序列
56.57.[0058][0059]
注:下划线部分为fam标签序列,波浪线部分为hex标签序列。
[0060]
e)进行分析。设计的15对kasp引物中最终得到了1个分子标记kasp
‑
sicau11,其与斑点叶基因lm5共分离。结果如图1和图2所示。
[0061]
实施例2分子标记kasp
‑
sicau11在选择控制斑点叶基因lm5的应用
[0062]
(1)利用有斑点叶的小麦突变体材料mc21为母本,叶片正常的普通小麦品种cn16为父本构建f3代群体,在后代株系中随机混合选取100个株系,每个株系包含10个单株。
[0063]
(2)对所获得的100个株系进行kasp
‑
sicau11标记检测,具体方法为:在苗期用ctab法提取100个株系的dna;以其作为底物模板,以分子标记kasp
‑
sicau11的特异性引物对为引物进行荧光定量pcr扩增,所述引物为:
[0064]
fam标签上引物:(下划线部分为fam标签序列)5
’‑
gaaggtgaccaagttcatgcttcgcagttaaagcgtcagc
‑3’
(seq id no.31);
[0065]
hex标签上引物:(波浪线部分为hex标签序列)5
’‑
gaaggtcggagtcaacggatttcgcagttaaagcgtcagt
‑3’
(seq id no.32);
[0066]
通用下游引物:5
’‑
cacatgcttgggagtaac
‑3’
(seq id no.33)。
[0067]
上述pcr扩增的扩增体系为:5μl master mix、混合引物1.4μl、5ng模板dna、双蒸水加至总量为10μl,同时需添加至少3个独立的以双蒸水代替dna模板的空白。上述混合引
物是由三条引物seq id no.1、2和3按照10ng/μl的浓度,分别加入120μl,120μl和300μl并添加ddh2o 460μl进行混合后制备而成的。
[0068]
上述pcr扩增的程序为:94℃预变性15min;94℃变性20s、65℃复性/延伸60s,共10个循环;94℃变性20s、55℃复性/延伸60s,共30个循环;完成后进行荧光读值。
[0069]
荧光读值结果(见图3),将检测到与cn16一致的fam(圆形)荧光的植株基因型记为“a”,为正常叶片的株系,同mc21一样表现为hax(正方形)荧光的基因型记为“b”,为斑点叶类型的株系,绿色三角形荧光为杂合株系,基因型记为“h”。各个株系基因型与田间表型值如表2所示。
[0070]
表2 mc21
×
cn16 f3代混取株系kasp
‑
sicau11分型结果统计
[0071]
[0072]
[0073][0074]
由表2可见,对比斑点叶性状数据发现,株系基因型与斑点叶表型数据性状表型一致。
[0075]
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利要求书确定的保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。