1.本发明属于分子生物学技术领域,涉及一种用于任意基因表达调控的组合物及方法。
背景技术:
2.随着人类基因组计划的完成,生物学研究已逐步进入了后基因组时代。后基因组时代的重要研究主题是探索基因功能,以及寻找调控网络;而基因功能和调控网络的系统表征则依赖于通过缺失、抑制或过表达来对基因进行准确操控。因此,dna和rna操控工具,如基因编辑工具和基因调控工具,可以深入推进相关研究的开展。
3.由zfns、talens、crispr系统为代表的基因编辑工具可以在dna链上进行切割,从而在目标基因的编码区域中插入缺失,造成阅读框移码,继而破坏特定基因或调控元件的功能。最近,我们也报告了一种新型基因编辑工具hpsgn系统,它是由fen1蛋白和引导探针组成的。不同于传统基因编辑工具的是,hpsgn系统对靶标底物的序列没有限制,且靶标底物的种类就可以是dna,也可以是rna。但这些“切割”方法有其弊端,以hpsgn系统为例,hpsgn会破坏靶标,造成的效果是不可逆转的,导致其无法被应用于致命突变和必须基因的研究。此外,由于dna双链断裂的形成会产生细胞毒性,可能造成不必要的损伤。
4.相较于基因编辑工具,不破坏改变靶序列的基因调控工具,可以以相对直接和可逆的方式进行基因功能调控。首先,在现有的基因调控工具中,rnai技术是使用最为广泛普遍的,通过与靶标rna结合下调目标基因的表达量,但该技术仅限于应用在具有适当宿主机制的特定生物体内。在某些生物体中,比如原核生物,就无法使用。其次,另一种基因调控工具,基于催化失活的cas蛋白(dcas)的crispr调控系统crispri/a,也已被广泛应用于细胞增殖、药物或毒素敏感性以及细胞转录组等研究。在dcas蛋白相关家族中,dcas9被用于开发靶向基因组dna的crispri系统,可通过干扰转录过程下调基因表达量;而dcasrx被用于开发靶向rna底物crispri系统,可通过干扰选择性剪接过程操纵基因表达量变化。
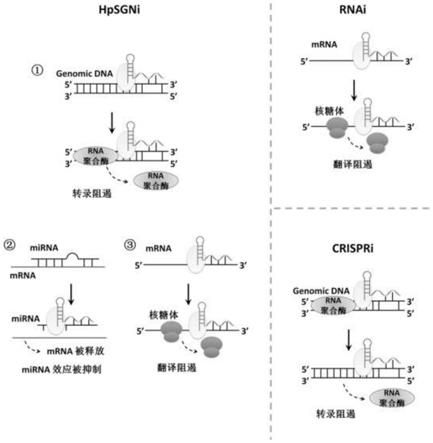
5.然而,到目前为止,尚没有一种基因调控工具,可以同时满足既可作用于dna靶标,也可作用于rna靶标;既可在原核细胞中工作,也可以在真核细胞中工作;既可以上调基因表达量,也可下调基因表达量。我们认为可以利用fen1蛋白可以同时靶向dna和rna的特点,设计一种基因调控系统hpsgni。一方面,当靶向基因组dna时,可以通过占据启动子区域或编码区域,阻遏转录的起始或延伸,达到下调基因表达的目的;另一方面,当靶向mirna时,可以通过占据mirna,阻遏其与特定mrna的结合,达到释放mrna从而上调基因表达的目的。另外,fen1类蛋白在原核和真核生物中均有亚型,物种实用范围广。
技术实现要素:
6.本发明的目的是针对现有基因调控工具的上述不足,提供一种新型基因调控工具及方法,可以同时满足既可作用于dna靶标,也可作用于rna靶标;既可在原核细胞中工作,也可以在真核细胞中工作;既可以上调基因表达量,也可下调基因表达量。
7.本发明的目的可通过以下技术方案实现:
8.一种用于任意基因表达调控的组合物,包括两个主要组分:
9.组分a:寡核苷酸探针:寡核苷酸探针由两部分组成,一部分具有核酸二级结构;另一部分为基本与靶标底物互补的单链区,但在其中某些特定位置上的碱基与靶标底物不互补。
10.组分b:蛋白分子:该蛋白分子可以识别组分a所述的寡核苷酸探针的核酸二级结构,从而与探针结合,再被探针引导,结合靶标底物形成“靶标底物
‑
蛋白分子
‑
探针”三元复合物;由于寡核苷酸探针某些特定位置上的碱基与靶标底物不互补,蛋白分子对靶标底物只发生结合反应,不会发生切割反应。
11.所述的与靶标底物不互补既包含与靶标底物相同所致的不互补,也包含与靶标底物不相同且不互补的情况,无论哪种情况,只要与靶标底物不互补,即可实现本发明。
12.作为本发明的一种优选,所述的特定位置为单链区的1个或多个碱基,优选单链区的1
‑
3个碱基。
13.作为本发明的一种优选,所述的蛋白分子含有核定位信号或核导出信号。
14.作为本发明的进一步优选,所述的组合物,包括两个主要组分:
15.组分a:寡核苷酸探针:寡核苷酸探针由两部分组成,一部分具有茎
‑
环核酸结构,另一部分与靶标底物基本互补,但在某个特定位置上的碱基与靶标底物不互补;寡核苷酸探针数量包括但不限于1条;
16.组分b:蛋白分子:该蛋白分子可以识别所述的寡核苷酸探针的茎
‑
环核酸结构,从而与探针结合。
17.作为本发明的进一步优选,所述寡核苷酸探针的长度不受限制,优选为16nt以上。
18.作为本发明的进一步优选,所述的蛋白分子选自afufen、pfufen、mjafen、mthfen、homo sapiens fen中任意一种的部分功能域或全酶片段。
19.本发明所述的组合物在制备基因调控试剂中的应用。
20.本发明所述的组合物在基因调控中的应用,所述的基因调控不包括以疾病诊断和或治疗为目的的基因调控。
21.一种用于任意基因表达调控的方法,根据靶多核苷酸,设计并合成本发明所述的寡核苷酸探针,向含有靶多核苷酸的体系中加入所述的寡核苷酸探针和本发明所述的蛋白分子,所述的蛋白分子识别寡核苷酸探针的二级结构,从而与探针结合,再被探针引导,结合靶标底物形成“靶标底物
‑
蛋白分子
‑
探针”三元复合物;基于寡核苷酸探针某个特定位置上的碱基与靶标底物不互补,蛋白分子对靶标底物只发生结合反应,不会发生切割反应。
22.作为本发明的一种优选,所述的靶多核苷酸为dna或rna,优选是基因组dna、质粒dna,mrna或microrna(又称为mirna)。
23.有益结果:
24.第一,本发明提供的新型基因调控工具及方法,可以既作用于dna靶标,也作用于rna靶标,具有极强的通用性。
25.第二,本发明提供的新型基因调控工具及方法,既可以在原核细胞中工作,也可以在真核细胞中工作,具有广泛的适用性。
26.第三,本发明提供的新型基因调控工具及方法,既可以用于上调基因表达量,也可
以用于下调基因表达量,还可以同时对不同的基因进行上调和下调发生。
27.第四,本发明中蛋白分子比crispr调控系统中的dcas蛋白小,有利于体内递送。
28.第五,本发明涉及的方法不会受到靶标底物的序列限制。
附图说明
29.图1
‑
本发明与最接近现有技术的原理示意图
30.图2
‑
是关于实施例1的结果图
31.图3
‑
是关于实施例2的结果图
32.图4
‑
是关于实施例2的另一个结果图
33.图5
‑
是关于实施例3的结果图
34.图6
‑
是关于实施例4的结果图
35.图7
‑
是关于实施例5的结果图
36.图8
‑
是关于实施例6的结果图
37.图9
‑
是关于实施例7的结果图
38.图10
‑
是关于实施例8的结果图
39.图11
‑
是关于实施例9的结果图
40.图12
‑
是关于实施例10的结果图
41.图13
‑
是关于实施例11的结果图
42.图14
‑
是关于实施例12的结果图
43.图15
‑
是关于实施例13的结果图
具体实施方式
44.参照下列实施例更详细描述本发明。然而,本发明不应解释为限制于此。实施例中的mis
‑
hpdna探针是本发明所述核酸探针中的一种。实施例中的afufen1是本发明所述蛋白分子中的一种。
45.实施例1〈afufen1蛋白能够在mis
‑
hpdna探针的引导下,结合而非切割靶标dna〉
46.为了验证afufen1蛋白(核苷酸序列如seq id no.1所示,编码的氨基酸序列如seq id no.2所示)能否成功在mis
‑
hpdna探针的引导下靶向结合目标底物ssdna,如图2所示,我们设计了几种探针,这些探针的3’端具有茎
‑
环结构,5’端基本与靶标ssdna互补,但在某些特定的位置上与靶标ssdna不匹配。将表达纯化出的afufen1蛋白分别与这些探针在37℃条件下孵育20min,再加入ssdna共孵育30min,反应产物进行emsa分析。
47.如图2所示,当探针上不存在错配碱基时(pc组,hp4
‑
17),泳道中的底物几乎被完全切割(标注为切割产物);当探针上的错配碱基处于位置1(hp4
‑
17
‑
m1)时,泳道中没有切割条带;当探针上的错配碱基处于位置2(hp4
‑
17
‑
m2)时,泳道中有微弱的切割条带;当探针上的错配碱基处于位置3(hp4
‑
17
‑
m3)时,泳道中有较明显的切割条带;当探针上的错配碱基处于位置1到3(hp4
‑
17
‑
m)时,泳道中没有切割条带。上述结果说明,当探针上的错配碱基处于位置1到3时,afufen1不会被引导切割靶标底物ssdna。另外,如图2虚线框标注所示,尽管不产生切割反应,但能观察到由“ssdna
‑
探针
‑
fen1蛋白”组成的三元复合物,该复合物的分子量大约在65kda(m是蛋白marker,其中最粗的条带是70kda)。上述结果说明,当探针上
的错配碱基处于位置1到3时,afufen1可以在探针的引导下,不切割靶标底物ssdna,而只发生结合反应。在后文中,我们将这种错配碱基处于位置1到3的探针称为mis
‑
hpdna。
[0048][0049]
实施例2〈afufen1蛋白可以在mis
‑
hpdna探针的引导下,高效结合靶标底物dna〉
[0050]
为了考察afufen1蛋白能否在mis
‑
hpdna的引导下,充分结合靶标底物ssdna,如图3所示,我们逐渐增加afufen1蛋白的量,随着fen1蛋白相对靶标ssdna的比例不断增大,“ssdna
‑
探针
‑
fen1”三元复合物的量逐渐变多,说明afufen1在mis
‑
hpdna探针(hp4
‑
17
‑
m)的引导下,可以较好地结合靶标底物(ssdna
‑
s2)。
[0051]
为了进一步提高结合效率,我们在mis
‑
hpdna探针的3’端增加茎
‑
环结构的复杂性,分别为版本1.0(hp4
‑
19
‑
m
‑
ver1.0),1.1(hp4
‑
19
‑
m
‑
ver1.1),1.2(hp4
‑
19
‑
m
‑
ver1.2)和1.3(hp4
‑
19
‑
m
‑
ver1.3)号探针。如图4所示,随着茎
‑
环结构的复杂性增加,“ssdna
‑
探针
‑
fen1”三元复合物的量也逐步增加。上述结果说明,afufen1可以在mis
‑
hpdna探针的引导下,高效结合靶标底物dna。
[0052][0053][0054]
实施例3〈afufen1蛋白可以在mis
‑
hpdna探针的引导下,结合靶标底物rna〉
[0055]
为了考察afufen1能否在mis
‑
hpdna探针的引导下,结合靶标底物rna,如图5所示,我们将表达纯化出的afufen1蛋白分别与1.0(hp4
‑
19
‑
m
‑
ver1.0),1.1(hp4
‑
19
‑
m
‑
ver1.1),1.2(hp4
‑
19
‑
m
‑
ver1.2)和1.3(hp4
‑
19
‑
m
‑
ver1.3)版本的mis
‑
hpdna探针在37℃条件下孵育20min,再加入靶标底物ssrna(ssrna
‑
s2)共孵育30min,反应产物进行emsa分析。能观察到虚线框标注的,由“ssrna
‑
探针
‑
fen1蛋白”组成的三元复合物;该结果说明,afufen1可以在mis
‑
hpdna探针的引导下,高效地结合rna靶标底物。
[0056][0057]
实施例4〈“ssdna
‑
探针
‑
fen1蛋白”三元复合物的形成对靶底物序列没有偏爱性〉
[0058]
为了确认“ssdna
‑
探针
‑
fen1蛋白”三元复合物的形成对靶底物序列有无偏爱性,如图6所示,我们在靶标底物(ssdna
‑
s2)的不同区域设计探针(hp4
‑
19
‑
1,hp4
‑
19
‑
2,hp4
‑
19
‑
3,hp4
‑
19
‑
4),观察三元复合物的生成效率会否因为探针结合区域底物序列的不同而不同。实验结果所示,无论针对靶标底物哪个区域设计探针,“ssdna
‑
探针
‑
fen1蛋白”三元复合物都能很好地形成;说明“ssdna
‑
探针
‑
fen1蛋白”三元复合物的形成对靶底物的序列没有偏爱性。
[0059][0060][0061]
实施例5〈afufen1蛋白和mis
‑
hpdna探针在原核细胞中可以调控外源报告基因的表达〉
[0062]
假使afufen1蛋白和mis
‑
hpdna探针能够在原核细胞中调控基因表达,那么预测其工作机制如图7
‑
a所示,一方面,当mis
‑
hpdna探针结合在目标基因的启动子区域时,会抑制rna聚合酶的结合,使转录不能启动发生,从而下调靶基因的表达;另一方面,当mis
‑
hpdna探针结合在目标基因的编码区域时,会抑制rna聚合酶的前进延伸,使转录不能完整完成,从而下调靶基因的表达。
[0063]
为了验证上述调控能否发生,我们将表达afufen1蛋白的原核质粒转化进大肠杆菌bl21
‑
egfp细胞中,通过抗性筛选阳性克隆;再将分别靶向egfp基因启动子区和编码区的mis
‑
hpdna探针(egfp
‑
p1、egfp
‑
np1、egfp
‑
p2、egfp
‑
np2、egfp
‑
t1、egfp
‑
t2)电转进bl21
‑
egfp
‑
fen1细胞中。电转后的菌液在37℃条件下复苏1小时,再加入四环素诱导剂诱导fen1蛋白的表达,分别在诱导表达1、4和7小时后收集菌体,流式细胞仪分析egfp的表达量。结果如图7
‑
b所示,我们在6个不同的位置设计了探针,分别标记为p1、p2、np1、np2、t1、t2。当mis
‑
hpdna探针结合在目标基因的启动子区域时(如p1),可以观察到egfp蛋白的表达下调了80%;当mis
‑
hpdna探针结合在目标基因的编码区域时(如t1),可以观察到egfp蛋白的表达下调了50%。实验组与对照组有显著性差异。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在原核细胞中可以调控外源报告基因的表达。
[0064][0065]
实施例6〈afufen1蛋白和mis
‑
hpdna探针在原核细胞中可以调控内源基因的表达〉
[0066]
为了验证afufen1蛋白和mis
‑
hpdna探针在原核细胞中能否调控内源基因的表达,如图8所示,我们将表达afufen1蛋白的原核质粒转化进大肠杆菌mg1655细胞中,通过抗性筛选阳性克隆,制备电转感受态细胞;再针对大肠杆菌mg1655中内源基因folp和sula启动子区域设计了mis
‑
hpdna探针(folp
‑
p1和sula
‑
p1),将探针电转进感受态细胞中,电转后的菌液在37℃条件下复苏1小时,再加入四环素诱导剂诱导fen1蛋白的表达,诱导表达4小时后收集菌体,folp和sula的表达量用real
‑
time qpcr定量(引物分别为folp
‑
f/r,sula
‑
f/r,内参基因为reca,引物为reca
‑
f/r),结果表明,与对照组相比,实验组中folp和sula的表达量下调25%左右。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在原核细胞中可以调控内源基因的表达。
[0067][0068]
实施例7〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中抑制转录,从而下调外源报告基因的表达〉
[0069]
为了验证afufen1蛋白和mis
‑
hpdna探针能够在真核细胞中抑制转录从而下调基因表达,如图9所示,我们将表达afufen1蛋白的真核质粒以及egfp质粒转染进人肾细胞hek293a中,再将分别靶向egfp基因增强子区和编码区的mis
‑
hpdna探针转染进人肾细胞hek293a中,48小时候收细胞,流式细胞仪分析egfp的表达量。结果如图9所示,无论当mis
‑
hpdna探针结合在目标基因的增强子区时(如a1、a2、na1和na2),还是当mis
‑
hpdna探针结合在目标基因的编码区域时(如t1、nt1和t2),可以观察到egfp蛋白的表达下调了70
‑
80%,实验组与对照组有显著性差异。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在真核细胞中可以调控外源报告基因的表达。
[0070][0071]
实施例8〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中抑制转录,从而下调内源基因的表达〉
[0072]
为了验证afufen1蛋白和mis
‑
hpdna探针在真核细胞中能否调控内源基因的表达,如图10所示,我们将表达afufen1蛋白的真核质粒转染进人hela细胞中,再针对人基因组中内源基因cd71和cxcr4各设计了四组mis
‑
hpdna探针(cd71
‑1‑
version1.0、cd71
‑1‑
version1.1、cd71
‑1‑
version1.2、cd71
‑1‑
version1.3、cxcr4
‑1‑
version1.0、cxcr4
‑1‑
version1.1、cxcr4
‑1‑
version1.2、cxcr4
‑1‑
version1.3)。cd71和cxcr4的表达量用real
‑
time qpcr定量(引物分别为cd71
‑
f/r,cxcr4
‑
f/r,内参基因为gapdh,引物为gapdh
‑
f/r),结果表明,与对照组相比,实验组中cd71和cxcr4的表达量下调40%左右。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在真核细胞中可以调控内源基因的表达。
[0073][0074]
[0075]
实施例9〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中抑制翻译,从而下调外源报告基因的表达〉
[0076]
为了验证afufen1蛋白和mis
‑
hpdna探针能够在真核细胞中抑制翻译从而下调基因表达,如图11
‑
a所示,我们在编码afufen1蛋白序列的c端融合表达核导出序列nes,构建只在细胞核外表达蛋白的真核质粒,将构建好的质粒以及egfp质粒转染进人肾细胞hek293a中,再将靶向egfp基因编码区的mis
‑
hpdna探针nt1(nt1
‑
version1.2)转染进人肾细胞hek293a中,48小时候收细胞,流式细胞仪分析egfp的表达量。结果如图11
‑
b所示,靶向egfp基因编码区的mis
‑
hpdna探针nt1会结合在egfp的mrna上,实验组与对照组有显著性差异,可以观察到egfp蛋白的表达下调了62%。图11
‑
c所示,afufen1蛋白和mis
‑
hpdna探针不仅可以在细胞核内通过结合egfp基因组编码区阻止转录延伸,还可以在细胞核外通过结合egfp的mrna阻止翻译发生,从而下调egfp的表达量。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在真核细胞中通过抑制翻译调控外源报告基因的表达。
[0077][0078][0079]
实施例10〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中抑制翻译,从而下调内源基因的表达〉
[0080]
为了验证afufen1蛋白和mis
‑
hpdna探针在真核细胞中能否抑制翻译,从而调控内源基因的表达,如图12所示,我们在编码afufen1蛋白序列的c端融合表达核导出序列nes,构建只在细胞核外表达蛋白的真核质粒,将该真核质粒转染进人hela细胞中,再针对人基因组中内源基因cxcr4的mrna设计了mis
‑
hpdna探针(cxcr4
‑5‑
version1.3)。cd71的表达量分别用流式细胞仪定量其蛋白水平,以及real
‑
time qpcr定量其转录水平(引物分别为cxcr4
‑
f/r,内参基因为gapdh,引物为gapdh
‑
f/r),结果表明,与对照组相比,实验组中cxcr4的蛋白表达量下调50%左右,但转录水平没有明显差异变化。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在真核细胞中可以通过抑制翻译,下调内源基因的表达。
[0081][0082]
实施例11〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中上调内源基因的表达〉上述实施例均是下调基因表达量,为了考察afufen1蛋白和mis
‑
hpdna探针在真核细胞中能否上调内源基因的表达,我们选取microrna(简称mirna)为靶标。如图13
‑
a所示,在真核细胞中,某些mirna可以特异性结合到对应的mrna上形成双链rna分子,继而引起降解或者抑制其翻译;而afufen1蛋白和mis
‑
hpdna探针可以结合mirna分子形成三元复合物,从而减少mirna对mrna的抑制作用,继而提高了对应基因的表达量。如图13
‑
b所示,已有研究表
明,内源基因egfr蛋白的表达受mirna
‑
21的抑制,我们设计了靶向mirna
‑
21的mis
‑
hpdna探针(mis
‑
hp
‑
mi21
‑
5p
‑
version1.0、mis
‑
hp
‑
mi21
‑
5p
‑
version1.1、mis
‑
hp
‑
mi21
‑
5p
‑
version1.2、mis
‑
hp
‑
mi21
‑
5p
‑
version1.3),通过对mirna
‑
21的抑制上调egfr基因的表达量。结果如图13
‑
b所示,当细胞内转入afufen1蛋白和mis
‑
hpdna探针时,egfr的表达量上调了3倍左右,与对照组有显著性差异。上述结果表明,afufen1蛋白和mis
‑
hpdna探针在真核细胞中可以上调内源基因的表达。
[0083][0084]
实施例12〈afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中同时上调和下调内源基因的表达〉
[0085]
为了考察afufen1蛋白和mis
‑
hpdna探针能否在真核细胞中同时上调和下调内源基因的表达,如图14
‑
a所示,我们分别设计了靶向cxcr4基因mrna,以及靶向mirna
‑
21的mis
‑
hpdna探针(cxcr4
‑5‑
version1.3、mis
‑
hp
‑
mi21
‑
5p
‑
version1.3);前者可以通过抑制转录下调cxcr4基因的表达,后者通过抑制mirna
‑
21与egfr基因mrna的结合上调egfr的表达。实验结果如图14
‑
b,c和d所示,当细胞内转入afufen1蛋白和mis
‑
hpdna探针时,cxcr4的表达量下调45%,egfr的表达量上调了197%左右,与对照组有显著性差异。上述结果表明,afufen1蛋白和mis
‑
hpdna探针可以在真核细胞中同时上下调不同内源基因的表达。
[0086][0087]
实施例13〈hpsgni与现有基因调控方法rnai,orfs和crispri的比较〉
[0088]
在有些情况下,为了全面表征基因功能和调控网络,我们需要同时上调和下调不同基因的表达。对传统方法来说,需要一种基因上调工具和一种基因下调工具同时在同一个细胞中工作,这会给操作带来不便,甚至可能造成不同工具之间的相互干扰。然而,到目前为止,尚没有一种基因调控工具,既可以上调基因表达量,也可下调基因表达量。hpsgni系统较传统基因调控方法的优势在于,一方面,当靶向基因组dna时,可以通过占据启动子区域或编码区域,阻遏转录的起始或延伸,达到下调基因表达的目的;另一方面,当靶向mirna时,可以通过占据mirna,阻遏其与特定mrna的结合,达到释放mrna从而上调基因表达
的目的。
[0089]
为了更好地证明hpsgni系统的优势,我们比较了rnai、开放阅读框(orfs)、crispri和hpsgni同时上调egfr蛋白的表达和下调egfp的表达的可能性。在这个平行实验中,首先将编码egfp的质粒转染hek293a细胞。将靶向egfp基因编码区的sirna(sirna
‑
sense、sirna
‑
antisense)、靶向egfp基因编码区的sgrna表达dcas9蛋白的质粒、靶向egfr蛋白的质粒、表达fen1蛋白的质粒及靶向egfp基因编码区和靶向mirna
‑
21
‑
5p的探针(mis
‑
hp
‑
mi21
‑
5p
‑
version1.3、egfp
‑
nt1
‑
version1.2)转染到各组。流式细胞仪检测egfp和egfr的表达水平。如图15所示,sirna、orfs、crispri和hpsgni组的egfp表达水平分别下调约48%、0%、64%和30%。同时,sirna、orfs、crispri和hpsgni组egfr表达水平分别上调约3%、420%、0%和30%。hpsgni系统是唯一能同时抑制egfp表达和egfr活性表达的系统。
[0090]
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。