新型l-鼠李糖异构酶
技术领域
1.本发明涉及新型的l-鼠李糖异构酶及其制造方法、生产其的微生物、编码该酶的dna和包含该dna的重组载体和转化宿主细胞、l-鼠李糖异构酶变异体、以及利用l-鼠李糖异构酶或变异体的酮糖或醛糖的制造方法。
背景技术:
2.稀少糖根据国际稀少糖学会的定义,为“自然界中仅微量存在的糖”,即自然界中存在量少的单糖。
3.碳原子数为4的单糖(丁糖)有4种醛糖、2种酮糖和3种糖醇。碳原子数为5的单糖(戊糖)有8种醛糖、4种酮糖和4种糖醇。碳原子数为6的单糖(己糖)全部为34种,醛糖有16种,酮糖有8种、糖醇有10种。碳原子数为7个的单糖(庚糖)的种类,碳原子数为7的七醛糖、七酮糖、庚糖醇分别存在32种、16种、16种。
4.例如,六碳糖(己糖)中,通常在自然界中大量存在的醛糖为d-葡萄糖、d-半乳糖、d-甘露糖、d-核糖、d-木糖、l-阿拉伯糖的6种,除此以外的d-阿洛糖等的醛糖被定义为稀少糖。即,作为醛糖中被视为稀少糖的糖,可以列举l-阿洛糖、l-古洛糖、l-葡萄糖、l-半乳糖、l-阿卓糖、l-艾杜糖、l-甘露糖、l-塔罗糖、d-塔罗糖、d-艾杜糖、d-阿卓糖、d-古洛糖、d-阿洛糖。作为酮糖,d-果糖在自然界中大量存在,相对于此,其他酮糖在自然界不是大量地存在,因此可以被称为稀少糖。作为酮糖中被视为稀少糖的糖,可以列举d-阿洛酮糖(别名d-psicose)、d-塔格糖、d-山梨糖、l-果糖、l-阿洛酮糖(别名l-psicose)、l-塔格糖、l-山梨糖。
5.近来,确立了成为所有稀少糖生产的基本原料的d-阿洛酮糖(别名d-psicose)的大量生产技术,使得获得困难的稀少糖的生产成为可能。考虑进而通过酶反应,d-阿洛酮糖成为生产d-阿洛糖等的新的稀少糖的中心。
6.d-阿洛糖为仅3位碳的oh基方向与d-葡萄糖不同的异构体的醛糖,也已知是酮糖的d-阿洛酮糖的异构体的稀少糖单糖类。d-阿洛糖已知有以其为有效成分的用于治疗选自急性肾衰竭和尿毒症的肾疾病的医药组合物(专利文献1)、用于延迟因肌萎缩侧索硬化症引起的运动障碍的发病或恶化的医药品(专利文献2)、血压上升抑制剂(专利文献3)、以用于抑制血管新生为特征的制剂(专利文献4)、t淋巴细胞增殖抑制剂(专利文献5)、或在腹膜透析液中配合使用的腹膜劣化抑制剂(专利文献6),另外还已知作为能够吃的农药(专利文献7)。最近,申请了关于被摄入肾细胞癌细胞的抗肿瘤效果(日本特愿2019-52195号)、对人尿路上皮癌细胞的强抗肿瘤效果(日本特愿2019-58477号)的发明。由于具有这样的特征,d-阿洛糖在医药、农药领域中作为新一代的核心原料备受瞩目,继d-阿洛酮糖之后要求确立d-阿洛糖的大量生产技术。
7.作为使用微生物生产的酶生产d-阿洛糖的方法,本发明的发明人开发了使用从施氏假单孢菌(pseudomonas stutzeri)分离得到的l-鼠李糖异构酶(l-rhamnose isomerase)由阿洛酮糖生产阿洛糖的技术(非专利文献1)。l-鼠李糖异构酶是催化l-鼠
李糖-l-鼠李树胶糖之间的可逆的异构化反应的酶,但已知来自p.stutzeri的l-鼠李糖异构酶不仅能够作用于l-鼠李糖-l-鼠李树胶糖之间,还能够作用于l-来苏糖-l-木酮糖之间、l-甘露糖-l-果糖之间、d-古洛糖-d-山梨糖之间、d-核糖-d-核酮糖之间、d-阿洛糖-d-阿洛酮糖之间,l-塔罗糖-l-塔格糖之间,具有广泛的底物特异性。利用该广泛的底物特异性,以从d-阿洛酮糖向d-阿洛糖的转换为中心,能够生产何森图(izumoring)上的各种各样的稀少醛糖和酮糖。
8.现有技术文献
9.专利文献
10.专利文献1:日本专利第5330976号公报
11.专利文献2:日本专利第5317055号公报
12.专利文献3:日本专利第5158779号公报
13.专利文献4:日本专利第4943839号公报
14.专利文献5:日本专利第4724824号公报
15.专利文献6:日本特开2009-269887号公报
16.专利文献7:日本专利第5816871号公报
17.专利文献8:美国专利第10,480,018号说明书
18.非专利文献
19.非专利文献1:j.ferment.biоeng.(1997)vol.84,p.319
技术实现要素:
20.发明所要解决的技术问题
21.现在,伴随着大部分稀少糖的生产成为可能,稀少糖生产也逐渐产生了新的课题。例如,被发现了生理活性的稀少糖d-阿洛糖从实验室水平的大量生产向涉及产业化的市场规模大量生产的转变成为了本发明发明人的新的课题。即,为了实现稀少糖生产中的高效的大量生产,考虑首先需要筛选具有更高的耐热性活性和大范围的最适ph的酶。
22.如上所述在现有的技术中,以纯的稀少糖d-阿洛酮糖为原料,使用公知的l-鼠李糖异构酶(ec 5.3.1.14),生产稀少糖d-阿洛糖。l-鼠李糖异构酶是催化从l-鼠李糖向l-鼠李树胶糖的异构化反应的酶,也能够催化从l-鼠李树胶糖向l--鼠李糖的异构化。已知也作用于d-阿洛糖与d-阿洛酮糖之间的异构化。由于异构化酶根据显示最高活性的底物来命名,所以即使在被命名为l-鼠李糖异构酶的酶中,其底物特异性也是各种各样的。
23.另外,该公知的酶中有在生成d-阿洛糖时,作为副产物也生成作为醛糖的d-阿卓糖(d-altrose)的酶,作为副产物的d-阿卓糖的存在成为了d-阿洛糖纯化工序中的d-阿洛糖的收率降低的原因。因此,大量生产引起的稀少糖d-阿洛糖的纯度也称为课题之一。
24.本发明将提供来自在制造食品时承认使用且被视为无毒性的微生物且活性高、高收率且能够从d-阿洛酮糖异构化为d-阿洛糖的新型的l-鼠李糖异构酶、提供具有该酶的微生物、以及提供使用该酶的制造方法为其课题。
25.用于解决技术问题的技术方案
26.本发明的发明人着眼于不仅在日本而且在欧洲和美国,也登载于作为食品承认了使用的清单的被视为基本无毒性的菌种,采集各地的土壤,从其中分离微生物,持续具有l-鼠李糖异构酶活性的微生物的探索。
27.其结果,在众多分离的菌株中发现了产生新型的l-鼠李糖异构酶的属于欧文氏菌(erwinia)属的微生物。作为该欧文氏菌属微生物的欧文氏菌erwinia billingiae产生活性高且高耐热性的新型的l-鼠李糖异构酶。
28.来自该微生物的新型的l-鼠李糖异构酶催化醛糖与对应的酮糖之间的异构化反应,具有醛糖-酮糖之间的异构酶活性,即识别醛糖的c1的cho基和c2的oh基并与其反应,将c1的cho基转变为oh基,将c2的oh基转变为co基,或者识别酮糖的c1的oh基和c2的co基并与其反应,将c1的oh基转变为cho基,将c2的co基转变为oh基。另外,在以d-阿洛酮糖为底物制造d-阿洛糖时,由于不生成作为副产物的d-阿卓糖,所以适于d-阿洛糖的制造。
29.即,本发明涉及下述(1)至(4)中记载的l-鼠李糖异构酶、和下述(5)中记载的微生物。
30.(1)一种l-鼠李糖异构酶,其为来自属于欧文氏菌(erwinia)属的微生物的l-鼠李糖异构酶,由sds-page测定得到的亚基的分子质量为48kda,具有下述(a)和(b)的底物特异性:
31.(a)具有识别醛糖的c1的cho基和c2的oh基并与其反应,将c1的cho基转变为oh基,将c2的oh基转变为co基,或者识别酮糖的c1的oh基和c2的co基并与其反应,将c1的oh基转变为cho基,将c2的co基转变为oh基的异构酶活性;
32.(b)具有催化l-鼠李糖与l-鼠李树胶糖之间、l-来苏糖与l-木酮糖之间、l-甘露糖与l-果糖之间、d-核糖与d-核酮糖之间、l-塔罗糖与l-塔格糖之间、以及d-阿洛糖与d-阿洛酮糖之间的异构化反应的活性。
33.(2)如上述(1)所述的l-鼠李糖异构酶,其具有下述(c)和(d)的理化学性质:
34.(c)反应最适ph为9;
35.(d)反应最适温度为70℃。
36.(3)如上述(1)或(2)所述的l-鼠李糖异构酶,其中,属于欧文氏菌属的微生物为欧文氏菌erwinia billingiae。
37.(4)如上述(1)至(3)中任一项所述的l-鼠李糖异构酶,其中,属于欧文氏菌属的微生物为国家技术评估学会专利微生物保藏中心作为保藏编号nite bp-03142进行了国际保藏的欧文氏菌erwinia billingiae gual218-3。
38.另外,本发明涉及下述(5)至(7)所述的蛋白质、下述(8)至(11)所述的dna、重组载体、或转化宿主细胞、或者下述(12)所述的微生物。
39.(5)包含序列编号1所示的氨基酸序列的蛋白质。
40.(6)一种蛋白质,其为由序列编号1所示的氨基酸序列构成的蛋白质的氨基酸取代变异体,与序列编号1所示的氨基酸序列具有80%以上的同一性,且具有选自序列编号1所示的氨基酸序列的k3、l4、i5、y9、e10、l11、y16、d18、v19、i21、v23、q25、v26、m27、t28、g32、i33、r46、n52、e54、r68、h73、a77、i79、e80、k81、m83、a89、d102、t103、e106、d108、a109、e111、q113、s116、h117、q124、h125、k126、s134、s148、d151、k152、g153、c162、i168、h171、p179、v186、l193、i195、l198、a199、e202、a205、s206、v211、f216、d217、a218、s219、c245、
l246、a250、t257、t275、v280、l300、t310、a313、n315、k316、n319、k320、a345、s357、d358、q359、r361、k362、l365、e366、a371、l375、v387、a390、w391、l393、h395、v397、d400、a401、s402、s405、e406、h409、q412、q413、t414、r416、l417、419中的至少1个位点的氨基酸取代,具有下述(a)和(b)的l-鼠李糖异构酶活性,且在最适温度的l-鼠李糖异构酶活性比由序列编号1所示的氨基酸序列构成的蛋白质高,
41.(a)具有识别醛糖的c1的cho基和c2的oh基并与其反应,将c1的cho基转变为oh基,将c2的oh基转变为co基,或者识别酮糖的c1的oh基和c2的co基并与其反应,将c1的oh基转变为cho基,将c2的co基转变为oh基的异构酶活性;
42.(b)具有催化l-鼠李糖与l-鼠李树胶糖之间、l-来苏糖与l-木酮糖之间、l-甘露糖与l-果糖之间、d-核糖与d-核酮糖之间、l-塔罗糖与l-塔格糖之间、以及d-阿洛糖与d-阿洛酮糖之间的异构化反应的活性。
43.(7)一种蛋白质,其为由序列编号1所示的氨基酸序列构成的蛋白质的氨基酸取代变异体,与序列编号1所示的氨基酸序列具有78%以上的同一性,且具有选自序列编号1所示的氨基酸序列的k3、l4、i5、y9、e10、l11、y16、d18、v19、i21、v23、e24、q25、v26、m27、t28、g32、i33、r46、n52、e54、r68、h73、a77、i79、e80、k81、a82、m83、s84、a89、k90、i97、d102、t103、e106、d108、a109、e111、q113、s116、h117、e120、q124、h125、k126、s134、p139、l140、s148、a150、d151、k152、g153、i154、c162、r167、i168、h171、p179、v186、l193、i195、l198、a199、e202、a205、s206、e210、v211、k215、f216、d217、a218、s219、c245、l246、a250、t254、t257、t275、v280、r282、l300、t310、a313、n315、k316、n319、k320、a345、s357、d358、q359、r361、k362、l363、l365、e366、y369、a371、a374、l375、s381、v387、a390、w391、l393、h395、v397、d400、a401、s402、s405、e406、h409、q412、q413、t414、r416、l417、419中的至少1个位点的氨基酸取代,具有下述(a)和(b)的l-鼠李糖异构酶活性,且相比于由序列编号1所示的氨基酸序列构成的蛋白质,反应温度70℃与50℃时的l-鼠李糖异构酶活性之比(t70/t50)更高,或者在60℃保温1小时后的残存活性更高,或者反应温度80℃与最适温度时的l-鼠李糖异构酶活性之比(t80/最适温度)更高,
44.(a)具有识别醛糖的c1的cho基和c2的oh基并与其反应,将c1的cho基转变为oh基,将c2的oh基转变为co基,或者识别酮糖的c1的oh基和c2的co基并与其反应,将c1的oh基转变为cho基,将c2的co基转变为oh基的异构酶活性;
45.(b)具有催化l-鼠李糖与l-鼠李树胶糖之间、l-来苏糖与l-木酮糖之间、l-甘露糖与l-果糖之间、d-核糖与d-核酮糖之间、l-塔罗糖与l-塔格糖之间、以及d-阿洛糖与d-阿洛酮糖之间的异构化反应的活性。
46.(8)编码上述(5)至(7)中任一项所述的蛋白质的dna。
47.(9)包含序列编号2所示的碱基序列的dna。
48.(10)含有上述(8)或(9)所述的dna的重组载体。
49.(11)由上述(10)所述的重组载体转化得到的转化宿主细胞。
50.(12)在国家技术评估学会专利微生物保藏中心作为保藏编号nite bp-03142进行了国际保藏的欧文氏菌erwinia billingiae gual218-3,其生产上述(1)至(4)中任一项所述的l-鼠李糖异构酶。
51.另外,本发明涉及下述(13)至(16)所述的固定化蛋白质。
52.(13)一种固定化蛋白质,其中,上述(1)至(4)中任一项所述的l-鼠李糖异构酶、或上述(5)至(7)中任一项所述的蛋白质固定化于载持体。
53.(14)一种固定化蛋白质,其中,上述(1)至(4)中任一项所述的l-鼠李糖异构酶以存在于菌体破碎物中的粗酶的状态、或上述(5)至(7)中任一项所述的蛋白质以存在于转化宿主细胞的菌体破碎物中的粗蛋白质的状态固定化于载持体。
54.(15)如上述(13)或(14)所述的固定化蛋白质,其中,上述载持体为离子交换树脂或合成吸附剂。
55.(16)如上述(15)所述的固定化蛋白质,其中,上述载持体为wa30、fpa54或fpa95。
56.另外,本发明涉及下述(17)至(19)所述的l-鼠李糖异构酶的制造方法、或酮糖或醛糖的制造方法。
57.(17)一种l-鼠李糖异构酶的制造方法,其中,在培养基中培养生产上述(1)至(4)中任一项所述的l-鼠李糖异构酶的欧文氏菌属微生物、或上述(11)所述的转化宿主细胞,使微生物菌体中积累该l-鼠李糖异构酶,并收集该l-鼠李糖异构酶。
58.(18)如上述(17)所述的l-鼠李糖异构酶的制造方法,其中,上述培养基为添加有l-鼠李糖的无机盐培养基。
59.(19)一种酮糖或醛糖的制造方法,其特征在于,使上述(1)至(4)中任一项所述的l-鼠李糖异构酶、上述(5)至(7)中任一项所述的蛋白质、或上述(13)至(16)中任一项所述的固定化蛋白质与含有选自醛糖或酮糖中的1种以上的溶液作用,生成对应的酮糖或醛糖,将其收集。
60.发明的效果
61.本发明的l-鼠李糖异构酶的特征在于,相比于来自微生物的现有的l-鼠李糖异构酶,具有特别高的耐热性,且为高活性。例如现有的来自施氏假单胞菌(pseudomonas stutzeri)的l-鼠李糖异构酶的最适温度为60℃,而相对于此,本发明的酶高达70℃,而且具有在60℃进行10分钟加热处理后的残存活性也为80%以上的热稳定性,因此适于工业生产中的使用。该酶具有的优异的耐热性能够通过制作该酶的氨基酸取代变异体,得到进一步提高。
62.另外,在底物为100mm的d-阿洛酮糖的情况下,在60℃每单位蛋白具有2.26u的高的d-阿洛糖的转换活性,通过本发明能够进行d-阿洛糖的大量生产。特别是在底物为d-阿洛酮糖的情况下,本发明的酶仅转变为d-阿洛糖,不生成副产物d-阿卓糖,收率相应地提高。
附图说明
63.图1是表示本发明的粗酶(来自gual218-3株,以下简称“本酶”)与对照粗酶(来自agm30株)的d-阿洛糖的生成量的比较的图。
64.图2表示用于确认本酶的分子质量的sds-page的结果。图中,左边的数字的单位为kda。
65.图3是表示本酶和对照酶的最适ph的图。
66.图4是表示本酶和对照酶的24小时后的ph稳定性的图。
67.图5是表示本酶和对照酶的最适温度的图。
68.图6是表示本酶和对照酶的保温10分钟的温度稳定性的图。
69.图7是表示本酶和对照酶的底物特异性的图。
70.图8是表示金属离子对本酶和对照酶的影响的图。
71.图9表示本酶利用固定化载持体得到的固定化酶的活性表现率的相对值。
72.图10表示利用大肠杆菌表达的重组酶的sds-page的结果。
73.图11表示利用重组酶进行d-阿洛酮糖向d-阿洛糖的转换反应的反应液的hplc的分析结果。
具体实施方式
74.本发明涉及能够从属于欧文氏菌(erwinia)属的微生物分离的l-鼠李糖异构酶,在高活性、耐热性上具有特征性的性质。
75.本发明的l-鼠李糖异构酶具有识别醛糖的c1的cho基和c2的oh基并与其反应,将c1的cho基转变为oh基,将c2的oh基转变为co基,或者识别酮糖的c1的oh基和c2的co基并与其反应,将c1的oh基转变为cho基,将c2的co基转变为oh基的异构酶活性。
76.本发明中所说的酮糖是指具有酮糖结构的六碳糖的己酮糖或五碳糖的戊酮糖。己酮糖包括阿洛酮糖(别名阿卢糖、psicose)、山梨糖、塔格糖、果糖,戊酮糖包括核酮糖、木酮糖。
77.本发明中所说的醛糖是指具有醛糖结构的六碳糖的己醛糖或五碳糖的戊醛糖。己醛糖包括葡萄糖、阿洛糖、阿卓糖、古洛糖、艾杜糖、塔罗糖、半乳糖、甘露糖,戊醛糖包括核糖、阿拉伯糖、木糖、来苏糖。d-或l-是指它们的d-体和l-体。
78.本发明的l-鼠李糖异构酶作用于l-鼠李糖、l-来苏糖、l-甘露糖、d-核糖、l-塔罗糖和d-阿洛糖,能够催化l-鼠李糖-l-鼠李树胶糖之间、l-来苏糖-l-木糖之间、l-甘露糖-l-果糖之间、d-核糖-d-核酮糖之间、l-塔罗糖-l-塔格糖之间以及d-阿洛糖-d-阿洛酮糖之间的转换,具有广泛的底物特异性。
79.本发明的l-鼠李糖异构酶能够通过培养属于欧文氏菌(erwinia)属、具有l-鼠李糖异构酶产生能力的微生物,从在培养液中生长的菌体中分离l-鼠李糖异构酶来制备。作为属于欧文氏菌(erwinia)属的微生物,例如欧文氏菌erwinia billingiae gual218-3和它们的变异株等能够被有效地利用。在提出该申请时,gual218-3株于2020年2月28日在位于日本千叶县木更津市上总(
かずさ
)镰足2-5-8的独立行政法人制品评价技术基盘机构国家技术评估学会专利微生物保藏中心进行国际保藏,作为收据编号nite abp-03142被接收,然后gual218-3株在2020年2月28日以保藏编号nite bp-03142正式基于布达佩斯公约进行了国际保藏。
80.关于本发明的l-鼠李糖异构酶,将欧文氏菌属的l-鼠李糖异构酶生产菌用添加有l-鼠李糖的无机盐培养基进行通气培养后,通过离心分离将菌体从培养液回收。将回收得到的菌体使用10mm tris-hcl缓冲液(ph7.5)清洗后,悬浮在10ml的10mm tris-hcl缓冲液(ph7.5)中,添加作为溶菌酶的lysozyme,利用酶处理将细胞破碎,或者一边将菌体悬浊液在冰水中冷却一边用超声波匀浆器进行细胞破碎。将这些破碎物离心分离,将其离心上清作为粗酶液。
81.纯化前的粗酶液的l-鼠李糖异构酶的活性能够通过以d-阿洛酮糖为底物,测定
d-阿洛糖的生成量来确认。
82.关于作为逆反应的从d-阿洛糖转变成d-阿洛酮糖的酶活性也以同样的条件进行测定。这些转变反应通常以如下条件进行。底物浓度为1~60%(w/v),优选为约5~50%(w/v),反应温度为30~80℃,优选为约50~70℃,反应ph为6~11,优选为约8~11,反应时间可以适当选择,在批式反应的情况下,通常选择4~20小时的范围。
83.粗酶液能够依次通过离子交换色谱和疏水色谱进行纯化,分离纯化酶。为了确认酶被纯化,进行sds-page(凝胶浓度12.5%),确认获得单一条带和表观的分子质量。
84.如上所述纯化得到的本发明的l-鼠李糖异构酶的利用sds-page测定的亚基分子质量约为48kda,该酶是活化度受金属离子调节的金属酶。与底物的反应能够在选自锰、钴、镍、镁、铁、铜、锌和钙中的金属离子以浓度0.5~5mm的存在下进行。
85.本发明的l-鼠李糖异构酶具有规定的氨基酸序列,作为其例子,可以列举具有序列编号1所示的氨基酸序列的蛋白质、或具有与其同源的氨基酸序列且维持同等的l-鼠李糖异构酶活性的蛋白质。同源的氨基酸序列是指例如与序列编号1的氨基酸序列具有75%以上、78%以上、80%以上、优选85%以上、更优选90%以上、更加优选95%以上的氨基酸序列的同一性。
86.两个氨基酸序列或两个核酸序列(碱基序列)的同一性(%)例如能够通过以下程序确定。首先,以能够最好地比较两条序列的方式排列。此时,例如可以在第一序列导入空位,以便于与第二序列的比对。在第一序列的特定位置的分子(氨基酸残基或核苷酸)与第二序列的对应位置的分子相同时,可以说该位置的分子相同。两个序列的同一性是该两个序列共通的同一位置的数量的函数(即,同一性(%)=同一位置的数/位置的总数
×
100),优选也考虑比对的优化所需要的空位的数量和大小。
87.另外,两个序列的比较和同一性的确定能够使用数学算法实现。作为序列的比较能够利用的数学算法的具体例,记载于karlin和altschul(1990)proc.natl.acad.sci.usa 87:2264―68,在karlin和altschul(1993)proc.natl.acad.sci.usa 90:5873―77中有改良的算法,但并不限定于此。这样的算法被编入altschul等(1990)j.mol.biol.215:403―10中记载的nblast程序和xblast程序(版本2.0)中。为了得到与本发明的核酸分子等价的核苷酸序列,例如用nblast程序设为score(得分)=100、wordlength(字长)=12进行blast核苷酸检索即可。
88.本发明的dna是编码上述蛋白质的基因,具有规定的碱基序列。作为其例,可以列举编码序列编号1所示的氨基酸序列的dna序列、具有序列编号2所示的碱基序列、或与序列编号2所示的碱基序列同源的碱基序列、且编码维持与序列编号1的蛋白质同等的l-鼠李糖异构酶活性的蛋白质的dna序列。同源的碱基序列例如是指与序列编号2的碱基序列具有75%以上、78%以上、80%以上、优选85%以上、更优选90%以上、进一步更优选95%以上的碱基序列的同一性。
89.本发明的dna也能够插入能够自主复制的适当载体来制成重组载体。重组载体由dna和能够自主复制的载体构成,只要能够获得dna,就能够通过常规方法的重组dna技术比较容易地制备。根据克隆或蛋白质的表达的使用目的或根据宿主细胞,选择适当的载体。作为这样的载体的例子,可以列举pbr322、puc18、pub110、ptz4、pc194、phv14、trp7、yep7、pbs7等的质粒载体或λgt
·
λc、λgt
·
λb、ρ11、等的噬菌体载体。
90.能够将这样得到的重组载体导入以大肠杆菌、枯草杆菌、放线菌、酵母为代表的适当的宿主细胞。导入方法能够使用磷酸钙共沉降法、电穿孔法、脂转染法、微注射法等公知的方法。为了取得转化的宿主细胞,适用菌落杂交法等。
91.本发明的l-鼠李糖异构酶、具有l-鼠李糖异构酶活性的蛋白质的制造方法没有特别限定,能够采用公知的方法。具体而言,本发明的l-鼠李糖异构酶能够通过从在营养培养基中培养转化了本发明的具有l-鼠李糖异构酶生产能力的微生物、或编码本发明的具有l-鼠李糖异构酶活性的蛋白质的dna的宿主细胞得到的培养物中,采集具有l-鼠李糖异构酶活性的蛋白质来制造。培养方法可以使用公知的方法,例如液体培养和固体培养的任意一种都可以使用。
92.这样培养菌体后,将本发明的酶、蛋白质纯化
·
回收。酶、蛋白质的纯化
·
回收方法能够自由选择公知的方法来进行,例如在从培养液回收的情况下,例如能够通过将培养上清进行过滤、离心处理等,去除不溶物后,将利用超滤膜的浓缩、硫酸铵沉淀等的盐析、透析、离子交换树脂等各种色谱等适当组合,来进行分离、纯化。另外,在从菌体内回收的情况下,例如将菌体通过溶解酶处理、超声波处理等破碎后,同样地进行分离、纯化。
93.另外,本发明的l-鼠李糖异构酶能够与其他异构酶同样,将酶固定化后利用,能够通过各种固定化方法,得到活性高的固定化酶。通过使用固定化酶,能够连续地进行大量的异构化反应,能够利用公知的固定化手段例如、载持体结合法、交联法、凝胶包埋法、微胶囊化法等制成固定化酶,另外,载持体可以为公知的任意的载持体。
94.将本发明的l-鼠李糖异构酶固定化的方法的一个方式为使用粗酶液的固定化。通过将菌体悬液进行超声波处理得到的包含l-鼠李糖异构酶的粗酶液添加到离子交换树脂等中,在低温下使其结合,能够将l-鼠李糖异构酶固定化。
95.为了从菌体内取出l-鼠李糖异构酶,需要将菌体细胞壁破碎,有利用超声波处理或溶菌酶等的酶处理的破碎方法,其如上所述。可知在通过超声波处理得到的粗酶液中,含有大量具有活性的l-鼠李糖异构酶。作为从粗酶液将酶固定化的载持体,也能够使用作为固定化载持体公知的任意的载体,离子交换树脂、海藻酸钠、合成吸附剂等便利且经常被使用。
96.离子交换树脂中,例如作为碱性阴离子交换树脂,也可以使用强碱性阴离子交换树脂或弱碱性阴交换树脂的任意一种,例如,作为强碱性阴离子交换树脂,可以列举sa20a、pa418(三菱化学株式会社制)等,作为弱碱性阴离子交换树脂,可以列举wa30(三菱化学株式会社制)、fpa54、fpa95(organo公司制)等,作为合成吸附剂,可以列举xad7hp(organo公司制)等。
97.在载持体中使用弱碱性阴离子交换树脂制造固定化酶的情况下,固定化的l-鼠李糖异构酶能够在其反应后容易地洗脱,因此能够非常简便地进行固定化酶的再生,生产效率高。
98.本发明的l-鼠李糖异构酶能够通过从用营养培养基培养本发明的具有l-鼠李糖异构酶生产能力的微生物、或转化了编码具有l-鼠李糖异构酶活性的蛋白质的dna的宿主细胞得到的培养物中,收集具有l-鼠李糖异构酶活性的蛋白质来制造。培养方法可以使用公知的方法,例如可以使用液体培养和固体培养的任意一种。
99.将菌体用培养基培养后,纯化
·
回收本发明的l-鼠李糖异构酶。蛋白质的纯化
·
回收方法能够自由地选择公知的方法进行,例如在从培养液回收的情况下,例如能够通过将培养上清过滤、离心处理等,将不溶物除去后,将利用超滤膜的浓缩、硫酸铵沉淀等的盐析、透析、离子交换树脂等的各种色谱等适当组合进行分离、纯化。另外,在从菌体内回收的情况下,例如将菌体利用溶解酶处理、超声波处理等破碎后,同样地进行分离、纯化。
100.能够使用本发明的纯化后的l-鼠李糖异构酶或固定化后的该酶,使其作用于含有能够成为底物的选自醛糖或酮糖中的1种以上的溶液,生成对应的酮糖或醛糖,制造酮糖或醛糖。本发明的l-鼠李糖异构酶对d-阿洛酮糖的底物特异性比现有的l-鼠李糖异构酶高,因此在能够成为底物的d-阿洛酮糖的存在下,能够大量制造作为稀少糖的d-阿洛糖。
101.另外,通过在本发明的l-鼠李糖异构酶的基因中导入突变,将对应的氨基酸残基通过点突变(部位特异性变异)操作取代成其他氨基酸残基,制备具有各种氨基酸取代的变异体,能够得到相比于没有氨基酸取代的野生型酶,最适温度下的d-阿洛酮糖异构酶活性更高的酶或耐热性更高的酶。用最适温度、反应温度70℃和50℃时的d-阿洛酮糖异构酶活性之比(t70/t50)、在60℃保温1小时后的残存活性、或在反应温度80℃和最适温度时的d-阿洛酮糖异构酶活性之比(t80/最适温度)高这4个指标评价耐热性。
102.本发明的l-鼠李糖异构酶即使是氨基酸序列的同一性为78%左右的变异体,也能够得到保持酶活性且与序列编号1的野生型酶相比较最适温度提高或者反应温度70℃和50℃时的d-阿洛酮糖异构酶活性之比(t70/t50)较高、或者在60℃保温1小时后的残存活性较高、或者反应温度80℃和最适温度时的d-阿洛酮糖异构酶活性之比(t80/最适温度)较高的变异体。
103.关于氨基酸取代变异体的设计,参考专利文献8中记载的制作系统树的方法。
104.将编码与本发明的序列编号1所示的l-鼠李糖异构酶具有类似活性的酶或推定酶的dna序列全部提取,使用专利文献8的方法,制作从作为共通祖先dna的系统发生起源的dna序列以系统发生的方式演化的系统树。并且,与系统树中与本发明的序列编号1相同的系统的所得到的dna序列所编码的氨基酸序列相比较,着眼于序列不同的部分的氨基酸序列,预测氨基酸残基不同的位点对酶活性的贡献小,通过利用点突变导入的碱基取代使序列编号1的与该不同的位点对应的氨基酸等变化,制作各种l-鼠李糖异构酶变异体。从其中,选择相比于野生型酶,最适温度下的d-阿洛酮糖异构酶活性更高的氨基酸取代变异体或耐热性高的氨基酸取代变异体。
105.另外,本发明的氨基酸取代变异体例外地包括在c末端的第418位的氨基酸添加1个氨基酸残基的变异体。
106.点突变导入法例如能够利用反向pcr法或退火法等(村松等人编著、“修订第4版新基因工学手册”、羊土社、p.82-88)任意的方法进行。根据需要也可以使用stratagene公司的quickchange ii site-directed mutagenesis kit、quickchange multi site-directed mutagenesis kit等各种的市售的点突变导入用试剂盒。
107.点突变导入最通常地能够使用包含要导入的核苷酸变异的变异引物进行。这样的变异引物以如下方式设计即可:包括在包含编码基因内的改变对象的氨基酸残基的核苷酸序列的区域退火,且代替编码该改变对象的氨基酸残基的核苷酸序列(密码子)而具有编码改变后的氨基酸残基的核苷酸序列(密码子)的碱基序列。
108.氨基酸取代变异体是指相对于原始的序列的氨基酸,取代为不同的氨基酸的变异体,取代有保守取代也有非保守取代,没有特别限制,在发明的优选方式中,取代为保守取代。保守取代对应于以碱性取代为碱性、酸性取代为酸性、极性取代为极性的方式,具有相同性质(碱性、酸性或中性)、极性(亲水性或疏水性)的氨基酸彼此的取代、芳香族氨基酸彼此的取代或脂肪族氨基酸彼此的取代等。
109.保守取代例如通过在碱性氨基酸(arg、lys、his)、酸性氨基酸(glu、asp)、中性非极性氨基酸(gly、ala、val、leu、ile、met)、脂肪族氨基酸(ala、val、leu、ile、met)、极性氨基酸(gln、asn、ser、thr)、芳香族氨基酸(phe、trp、tyr)等组内进行。
110.另一方面,非保守取代是指与上述组外的成员的氨基酸交换,例如为了防止在三维结构中蛋白质中折叠,使cys缺失,或者取代成其他氨基酸。或者,为了保持亲水性/疏水性的平衡,或为了使合成容易而提高亲水度,考虑作为与氨基酸相关的疏水性/亲水性的指标的氨基酸的亲水指数(j.mol.biol.(1982)vol.157,p.105-132),对氨基酸进行取代。
111.另外,有时也取代成比原始氨基酸空间位阻小的氨基酸的取代,从具有电荷的氨基酸取代成不具有电荷的氨基酸。
112.关于取代,有保守取代也有非保守取代,例如可以列举:从ala到ser或thr的取代、从arg到gln、his或lys的取代、从asn到glu、gln、lys、his或asp的取代、从asp到asn、glu或gln的取代、从cys到ser或ala的取代、从gln到asn、glu、lys、his、asp或arg的取代、从glu到asn、gln、lys或asp的取代、从gly到pro的取代、从his到asn、lys、gln、arg或tyr的取代、从ile到leu、met、val或phe的取代、从leu到ile、met、val或phe的取代、从lys到asn、glu、gln、his或arg的取代、从met到ile、leu、val或phe的取代、从phe到trp、tyr、met、ile或leu的取代、从ser到thr或ala的取代、从thr到ser或ala的取代、从trp到phe或tyr的取代、从tyr到his、phe或trp的取代、以及从val到met、ile或leu的取代,但不限定于这些。关于这些氨基酸残基的取代,能够取代的残基的分类为例示,能够取代的氨基酸残基并不限定于该分类。
113.在具有序列编号1的氨基酸序列的野生型酶中,从取代了1处氨基酸的变异体制作取代了90处氨基酸的多个多重变异体,通过进行酶活性和耐热性试验,选择58个变异体。其中,在58种中确认了13种最适温度下的d-阿洛酮糖异构酶活性比没有氨基酸取代的野生型酶高的变异体。
114.另外,反应温度70℃和50℃时的d-阿洛酮糖异构酶活性之比(t70/t50)比野生型酶高的变异体在58种中确认到52种,在60℃保温1小时后的残存活性比野生型酶高的变异体在58种中确认到17种,反应温度80℃和最适温度时的d-阿洛酮糖异构酶活性之比(t80/最适温度)比野生型酶高的变异体在58种中确认到56种。
115.本发明的l-鼠李糖异构酶在可以得到大量耐热性比野生型l-鼠李糖异构酶高的变异体的方面也优异。
116.实施例
117.以下,通过实验详细地说明本发明,但本发明不受以下实施例任何限定。
118.<实验1:菌株的来源和鉴定>
119.本发明的发明人将通过筛选分离的大量菌接种于添加有l-鼠李糖的液体培养基中进行振荡培养,以l-鼠李糖作为底物,测定l-鼠李树胶糖的生成量,由此测定l-鼠李糖异构酶的活性。
120.这样操作,作为活性最高的菌株,发现了微生物gual218-3株,gual218-3株通过基于16srrna基因碱基序列同源性的系统解析,判明属于欧文氏菌。
121.菌株的鉴定
122.(1)16srrna基因碱基序列同源性
123.分析16srrna基因的1-500bp区域,确定碱基数500bp。
124.(2)同源性检索
125.关于该菌株的16srrna基因的碱基序列,利用blast检索(日本dna数据库)进行与被视为基准菌株的已知菌种的同源性检索。从与相对于上述确定的碱基数500bp的碱基序列、同源性为98%以上的菌株名的同源性(%)的值,确定gual218-3株的微生物为欧文氏菌erwinia billingiae。
126.该gual218-3株在2020年2月28日在日本国家技术评估学会专利微生物保藏中心进行国际保藏,作为保藏编号nite bp-03142进行了国际保藏。
127.<实验2:欧文氏菌erwinia billingiae gual218-3(保藏编号nite bp-03142)株的培养>
128.在以l-鼠李糖1.0%为碳素源、硫酸铵为氮源的无机盐液体培养基中,无菌添加欧文氏菌erwinia billingiae gual218-3(保藏编号nite bp-03142)株的菌种培养液1%(v/v),一边通气搅拌一边在30℃培养24小时。
129.作为用于与本发明的菌株比较的对照菌株,将球形节杆菌(arthrobacter globiformis)m30(nite bp-1111)株(以下,称为“agm30株”。)以与gual218-3株同样的培养基和培养条件培养。
130.<实验3:粗酶的制备>
131.从gual218-3株和agm30株的各自的培养液通过离心分离回收菌体。回收得到的菌体分别利用10mm甘氨酸-naoh缓冲液(ph9.0)清洗,然后将菌体悬浮于10ml的甘氨酸-naoh缓冲液(ph9.0),之后将该菌体悬液一边在冰水中冷却一边用超声波匀浆器(日本艾默生株式会社)进行细胞破碎。将破碎物以12,000rpm离心20分钟,将其离心上清作为各自的粗酶。
132.<实验4:利用粗酶的d-阿洛糖的生产>
133.使用所得到的2个粗酶液,分别作为底物以成为最终浓度100mm的方式添加d-阿洛酮糖,在30℃进行4小时酶反应后,测定d-阿洛糖的生成量,利用hplc的区域面积比较d-阿洛糖的生成量。具体而言,酶反应液组成使用50mm甘氨酸-naoh缓冲液(ph9.0)、100mm d-阿洛酮糖、粗酶液、1mm氯化锰,在30℃反应4小时,通过2分钟煮沸,停止反应后,进行hplc,测定反应后的液体组成。将通过hplc分析得到的所生成的d-阿洛糖的区域面积的比表示于图1。
134.关于使用单位该培养液量的粗酶时的d-阿洛酮糖到d-阿洛糖的转变量,来自gual218-3株的本粗酶为来自agm30株的对照粗酶的约23倍。
135.<实验5:酶的纯化>
136.1.利用离子交换色谱、疏水色谱的纯化
137.将2个粗酶液利用离子色谱进行纯化。使用的色谱柱是用缓冲液(20mm tris-hcl ph7.5)平衡化后的hitrapq hp,使用akta系统以流速5ml/min、1m nacl的浓度梯度0%至
100%分成各5ml的组分进行分级。将检测出酶活性的组分回收,得到利用离子交换色谱分离的纯化酶。
138.2.将通过离子交换色谱分离纯化的酶进一步通过疏水色谱进行纯化。使用的色谱柱为hitrap phenyl,酶液中以成为2m的方式加入硫酸铵使其溶解,以流速5ml/min将2m的硫酸铵的浓度从100%降低到0%进行洗脱,每5ml进行分级。将检测出酶活性的组分透析,去除硫酸铵,得到疏水色谱分离纯化酶。
139.3.聚丙烯酰胺凝胶电泳
140.按照常规方法进行sds-page(凝胶浓度12.5%),确认纯化酶的纯度。在图2中,左侧为标准的蛋白质,中央泳道和右泳道为使用疏水色谱纯化后的酶。其结果在约48kda确认到单一条带,由此确认酶被纯净地纯化,判明纯化后的本酶的利用sds-page的亚基分子质量约为48kda。
141.<实验6:酶的理化学性质的测定>
142.关于作为纯化酶的l-鼠李糖异构酶活性的测定,进行下述的实验,进行酶反应。作为底物,使用5mm l-鼠李糖,在各条件下进行10分钟酶反应,反应后,在反应液加入10%三氯乙酸溶液50μl,由此停止反应,测定通过半胱氨酸咔唑硫酸法生成的l-鼠李树胶糖。
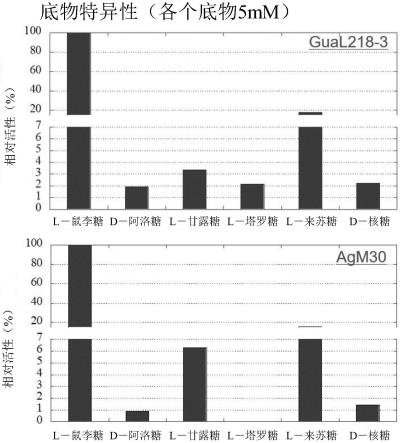
143.1.反应最适ph
144.在测定时,利用5mm l-鼠李糖作为底物,使用各种缓冲液ph3~11在30℃反应10分钟,将生成的l-鼠李树胶糖利用半胱氨酸咔唑硫酸法进行测定,求出反应的最适ph。
145.将反应条件表示于表1。使用的缓冲液表示于表2。
146.[表1]
[0147][0148]
[表2]
[0149][0150]
将结果表示于图3。本酶的反应最适ph为9,相对于此对照酶的反应最适ph为10在碱性侧,本酶的反应最适ph在更宽的范围,且在较为酸性的条件下显示高的活性。
[0151]
2.ph稳定性
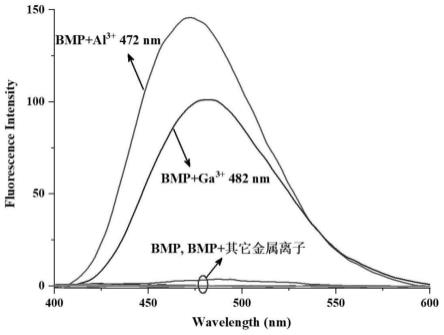
[0152]
接着,使用表2的ph4~11的4个缓冲液,将在各缓冲液中在30℃保持24小时后的残存活性表示于图4。
[0153]
本酶和对照酶在ph6~11都稳定,但对照酶的ph稳定性在ph6最高,而本酶在ph7.5和10最稳定。
[0154]
3.反应最适温度
[0155]
利用甘氨酸-naoh缓冲液将ph调整为9,在30-80℃的各个温度下进行反应。求出最适温度。反应条件表示于表3。40℃至80℃的温度范围为本酶的优选温度,从表示测定反应温度和相对活性的结果的图5可知本酶的最适温度为70℃。
[0156]
[表3]
[0157][0158]
对照酶的最适温度为50℃,但以本酶将特别显示最高活性的70℃设为100时的相对活性(%)计,在80℃为40%,在60℃为80%,在50℃为70%,相对于此,以对照酶显示最高活性的50℃设为100时的相对活性(%)计,在60℃为65%,在70℃为8%,本酶特别维持了在
高温时的活性,具有高的耐热性。
[0159]
4.热稳定性
[0160]
将利用表3所示的上述3.中求出最适温度的反应条件(10分钟),将本酶和对照酶在各温度保温10分钟后的残存活性表示于图6。在保温10分钟后的温度稳定性中,本酶在60℃的相对活性的减少低于20%,远远小于对照酶的80%的减少,因此可知本酶相比于对照酶,是在高温更为稳定的酶。
[0161]
5.d-阿洛糖或d-阿洛酮糖异构酶活性、
[0162]
关于本酶的d-阿洛糖异构酶活性的测定,进行与实验6同样的实验,进行酶反应。作为底物使用100mm d-阿洛糖,在60℃进行60分钟的酶反应,反应后,将反应液放入沸腾液中3分钟,由此停止反应,利用hplc测定反应后的液体组成。酶活性1单位(u)是在上述条件下将d-阿洛糖异构化在1分钟生成1μmol的d-阿洛酮糖的酶量。将反应条件表示于表4。
[0163]
在同样条件下,测定d-阿洛酮糖异构酶活性。作为底物使用100mm d-阿洛酮糖,在60℃进行60分钟酶反应,生成d-阿洛糖。酶活性1单位(u)是在上述条件下,将d-阿洛酮糖异构化在1分钟生成1μmol的d-阿洛糖的酶量。
[0164]
[表4]
[0165][0166]
其结果判明,本酶对于d-阿洛糖的比活性为5.21u/mg,对于d-阿洛酮糖的比活性为2.26u/mg。
[0167]
而且,在利用本酶将d-阿洛酮糖异构化为d-阿洛糖时,没有生成副产物的d-阿卓糖。
[0168]
6.l-鼠李糖异构酶的底物特异性
[0169]
对于本酶对l-鼠李糖和5种醛糖(d-阿洛糖、l-塔罗糖、l-来苏糖、d-核糖、l-甘露糖)的异构化活性进行了研究。酶反应组成如下述表5所示,在各底物最终浓度5mm、酶液(最终浓度50mm磷酸缓冲液ph8.0)中在70℃反应10分钟,通过利用hplc的分析,测定由各个醛糖异构化得到的酮糖。
[0170]
将l-鼠李糖的异构化活性设为100,将对于各个醛糖的活性作为相对活性表示。
[0171]
将d-阿洛糖(d-allose)、l-甘露糖(l-mannose)、l-塔罗糖(l-talose)、l-
来苏糖(l-lyxose)、d-核糖(d-ribose)作为底物,将活性以相对活性表示于表6和图7。
[0172]
[表5]
[0173][0174]
[表6]
[0175]
底物相对活性(%)l-鼠李糖100l-来苏糖17.9l-甘露糖3.3d-核糖2.2l-塔罗糖2.2d-阿洛糖1.9
[0176]
对于l-鼠李糖的活性最强,接着依次是l-来苏糖、l-甘露糖、d-核糖、l-塔罗糖、d-阿洛糖。另一方面,在对照酶时,依次为l-鼠李糖、l-来苏糖、l-甘露糖、d-核糖、d-阿洛糖,与l-塔罗糖不反应。本酶催化l-鼠李糖-l-鼠李树胶糖之间、l-来苏糖-l-木酮糖之间、l-甘露糖-l-果糖之间、d-核糖-d-核酮糖之间,l-塔罗糖-l-塔格糖之间、d-阿洛糖-d-阿洛酮糖之间的异构化反应。
[0177]
7.金属离子的影响
[0178]
接着,以调查金属离子对d-阿洛糖异构酶活性的影响为目的,将一部分酶透析,测定酶活性。透析通过将酶液加入纤维素膜,浸入包含20mm edta的甘氨酸-naoh缓冲液(ph9.0),将该缓冲液缓慢用16小时搅拌来进行,消除其他金属离子的影响。在各种二价金属离子1mm的存在下(表7的反应条件下)反应后,利用半胱氨酸咔唑法测定这样得到的酶的酶活性。
[0179]
其结果,cocl2使本酶活性显著上升,本酶显示金属依赖性(图7)。
[0180]
[表7]
[0181]
(金属离子的影响确认时的反应条件)
[0182][0183]
<实验7:固定化酶的制备>
[0184]
使用与实验2同样培养回收的欧文氏菌erwinia billingiae gual218-3株的菌体。
[0185]
1.菌体的利用超声波处理的粗酶的取得
[0186]
将对应于培养液1l的菌体悬浮于50mm甘氨酸-naoh缓冲液(ph9.0)40ml,将该菌体悬液一边在冰水中冷却一边用超声波匀浆器精细破碎。将破碎物以15,000
×
g离心30分钟,将其离心上清作为粗酶液。
[0187]
2.粗酶的固定化
[0188]
用纯水充分清洗膨润后,向由50mm甘氨酸-naoh缓冲液(ph9.0)平衡化后的离子交换树脂或合成吸附剂添加上述得到的粗酶液,在4℃缓慢混合20小时,由此使粗酶蛋白质结合。接着,用50mm甘氨酸-naoh缓冲液(ph9.0)清洗,得到固定化酶。
[0189]
作为固定化载持体,使用强碱性阴离子交换树脂(三菱化学株式会社制的sa10a、sa11a、nsa100、sa20a、pa306s、pa308、pa312、pa316、pa408、pa412、pa418、弱碱性阴离子交换树脂(三菱化学株式会社制的wa10、wa20、wa21j、wa30和organo公司制的fpa54、fpa60cl、fpa95)、或合成吸附剂(organo公司制的xad7hp、xad118on)。
[0190]
3.固定化酶的酶活性的测定方法
[0191]
固定化酶的酶活性的测定通过测定以d-阿洛酮糖为底物进行酶反应时的d-阿洛糖的生成量来进行。
[0192]
首先,以100mg的固定化酶树脂、甘氨酸-naoh缓冲液(ph9.0)(最终浓度50mm)、氯化锰(最终浓度1mm)、和d-阿洛酮糖(最终浓度100mm)的溶液(500μl)为反应液组成,在30℃恒温水槽反应24分钟后,通过在100℃煮沸2分钟,使反应立即停止。将该反应后的溶液冷却到室温后,用离子交换树脂(200ct和ira67的混合树脂(均为organo公司制))脱盐,进而进行过滤处理,制成分析样品。分析通过使用高效液相色谱(色谱柱:gl-c611(hitachi)、温度:60℃、洗脱液:0.1mm naoh、流速:1.0ml/分钟、检测器:rid-20a(岛津)),测定生成的d-阿洛糖的峰面积来进行。
[0193]
4.用于酶固定化的载持体对酶活性的比较
[0194]
为了确认酶固定化中使用的载持体造成的影响,比较了在上述2记载的各载持体固定化的本酶的酶活性。其中,相对值通过将比较样品中活性表现率最高的样品设为100计算而求得。
[0195]
图9表示了各种固定化酶的活性表现率的相对值。
[0196]
<实验8:编码酶的dna的克隆和包含该dna的重组载体和转化宿主细胞的制备>
[0197]
从欧文氏菌erwinia billingiae gual218-3株克隆编码具有d-阿洛酮糖异构酶活性的蛋白质的dna,进行能够自我复制的重组dna的制作、编码酶的dna的碱基序列的确定以及转化微生物的制备。
[0198]
<实验8-1:染色体dna的全碱基序列的确定>
[0199]
本菌的d-阿洛糖异构酶与现存的类似酶不同,无法使用pcr扩增的方法或现有的蛋白质数据库进行分离。因此,确定欧文氏菌erwinia billingiae gual218-3株的全基因组序列,之后进行该基因组中的全orf所编码的蛋白质群的数据库的构建。将欧文氏菌erwinia billingiae gual218-3株培养菌体作为供试菌体,委托株式会社macrogen japan,委托进行使用pacbio-rsii/sequel的新一代测序分析。
[0200]
其结果,得到碱基数4,301,131bp和1,024,754bp、280,568bp、126,938bp的4个重叠群。在该4个重叠群中约为5.7mb,因此考虑可以覆盖欧文氏菌erwinia billingiae gual218-3株的全基因组序列。
[0201]
<实验8-2:蛋白质数据库的构建>
[0202]
基于所得到的约5.7mb的dna序列,使用prokka程序,推测5,357个orf,对于各个orf预测氨基酸序列。将该5,357个氨基酸序列作为欧文氏菌erwinia billingiae gual218-3株的蛋白质数据库。
[0203]
<实验8-3:具有d-阿洛酮糖异构酶活性的蛋白质的鉴定>
[0204]
使用上述蛋白质数据库,登录蛋白质鉴定系统mascotserver(matrix science公司),进行观察到d-阿洛酮糖异构酶活性的蛋白质的鉴定。供试样品使用国际公开第[0045]段的sds-page的48kda的条带,将还原处理、烷基化处理后进行了胰蛋白酶消化的片段进行maldi-tof-ms解析。
[0205]
其结果,由于观察到与在下述的由418个氨基酸构成的序列1(序列表的序列编号1)的氨基酸序列中画下划线表示的氨基酸序列78%的相似性,所以强烈表明本蛋白质为欧文氏菌erwinia billingiae gual218-3株的l-鼠李糖异构酶。
[0206]
[序列1]
[0207]
mtklieqayelakqryadvgidveqvmtqldgipvsmhcwqgddvrgfenpngeltggiqatgnypgrarnahelradiekamslipgakrlnlhaiylesdtpverdaiepqhfshwvewakqhklgldfnpscfshplsadgftlshadkgirqfwidhckasrrisahfgeqlgtpsvmniwvpdgmkdltidrlaprerlasaldevisekfdashhidavesklfgigaesytvgsnefclgyaasrqtaltldaghfhptevisdkistamlyvprlllhvsrpvrwdsdhvvllddetqaiateiarnklfnkvhigldffdasinriaawvigtrnakkallrallepsdqlrklelegdytarlalleeqkslpwqavweawclrhdvpadaswlsevrhyeqqtlrlr
[0208]
(序列1使用欧文氏菌erwinia billingiaegual218-3株的蛋白质数据库鉴定的氨基酸序列。以下划线部表示的氨基酸序列是与maldi-tof-ms解析得到的峰一致的序列。)
[0209]
<实验8-4:具有d-阿洛酮糖异构酶活性的酶基因的分离>
[0210]
合成通过氨基酸序列鉴定的基因的序列2的dna序列(序列表的序列编号2),整合入pqe60载体(qiagen公司),转化表达用大肠杆菌。使用构建的大肠杆菌表达体系进行诱导酶的确认。sds-page的结果如图10所示,在可溶性组分中确认到诱导蛋白质。另外,使用该
诱导得到的重组酶,以60%(w/v)的d-阿洛酮糖为底物在30℃反应24小时,将用hplc确认d-阿洛酮糖异构酶活性的结果表示于图11。该图的保留时间22.48分钟的峰为d-阿洛糖,保留时间29.67分钟的峰为d-阿洛酮糖,本重组酶催化从d-阿洛酮糖到d-阿洛糖的异构化反应,并不生产可能作为副产物的d-阿卓糖。
[0211]
[序列2]
[0212]
atgacaaagctgattgaacaagcttatgaactggctaaacaacgttacgctgacgtaggaattgatgtggagcaggtgatgacgcaattggatgggattcctgtgtcgatgcactgctggcagggggatgacgtgcgcggcttcgaaaaccccaatggcgaactcaccggcgggattcaggccacaggtaactatccgggacgcgcgcgtaatgcgcatgaactgcgcgccgatatcgaaaaagccatgtcgctgattccaggcgctaagcgcctcaatctgcacgccatctatctggaaagcgacacgccggtagaacgtgatgctatcgaaccgcaacacttcagccactgggtggagtgggcaaaacagcataaattagggctcgattttaatccaagctgcttctcgcatcctttaagtgccgatggttttaccctgtcacacgctgacaaaggcatccgccagttctggattgatcactgcaaggccagtcgccgcatctcggcacattttggtgaacaactgggcacgccctcagtgatgaacatctgggttccggacggtatgaaagatctcaccattgatcgcctggcgccgcgtgaacgtttagccagcgcgctggatgaagtgatcagcgaaaaattcgatgccagtcatcatatcgacgccgtcgaaagtaagttattcgggattggtgctgagagttataccgtggggtccaatgagttttgccttggctacgctgccagccgccagaccgcgctgacacttgatgccgggcatttccatcccactgaagtgatctccgacaaaatctccaccgcgatgctgtatgtcccgcgcctgttattgcacgtcagccgtccggtgcgttgggacagcgaccatgtggtgctgctggatgatgaaacccaggccattgccactgaaattgcgcgtaacaagctgttcaacaaagtgcatatcggcctcgacttctttgatgcttccatcaaccgcatcgcggcgtgggtgattggtacccgcaatgccaaaaaggcgttactgcgtgcgctgttggagccgagtgaccaactgcgcaaactggagcttgagggggattacaccgcacgtctggcgctgctggaggagcaaaagtcattgccatggcaggctgtctgggaagcctggtgcttgcgtcacgatgtccctgctgatgccagttggctcagcgaagtccgtcattatgaacaacaaacactgcgtctacgttaa
[0213]
<实验9:本酶的氨基酸取代变异体的制作及其酶活性、耐热性>
[0214]
比较本发明的序列编号1所示的l-鼠李糖异构酶的氨基酸序列、与以专利文献8所记载的方法制作系统树、分类为与本发明的序列编号2的dna序列相同的系统的dna序列所编码的氨基酸序列和序列编号1的氨基酸序列,通过点突变导入的碱基取代而改变对应于与序列编号1不同的位点的氨基酸等,制作各种l-鼠李糖异构酶变异体。本发明的氨基酸取代变异体中例外地包括在c末端的第418位的氨基酸添加1个氨基酸残基(g)的变异体。
[0215]
通过点突变导入,首先制作单氨基酸取代变异体。制作具有在目标变异导入位点发生氨基酸取代的1到2个碱基对加入碱基取代的碱基序列的两个dna链的pcr引物,使用变异导入目的地的质粒dna和这两个引物进行pcr反应。通过测序分析确认将所得到的pcr片段转化到宿主大肠杆菌、从所得到的克隆提取质粒dna、导入目标变异。对于多重取代变异体,通过重复上述方法来制作。
[0216]
表达包含点突变酶的重组酶的重组大肠杆菌用包含以最终浓度为100μg/ml的方式加入了氨苄西林的3.5%多聚蛋白胨、2.0%酵母浸膏、1.0%氯化钠、2mm氯化锰的培养基在30℃、以200rpm进行12小时前培养。将前培养液以1/20量接种于以最终浓度100μg/ml的方式加入有氨苄西林的表达用培养基,在30℃以200rpm进行2小时正式培养。正式培养后,以最终浓度0.1mm的方式添加iptg,在30℃以200rpm进行一晩重组酶的表达诱导。
[0217]
将重组大肠杆菌的表达用培养基的组成表示于表8。
[0218]
[表8]
[0219][0220]
通过重组得到的野生型酶和氨基酸取代变异体的粗酶的酶活性如下测定:底物使用最终浓度100mm的d-阿洛酮糖,作为缓冲液使用最终浓度50mm的tris-hcl缓冲液(ph8.0),在各温度下反应10分钟,对反应液中的糖组成,使用hplc测定所生产的d-阿洛糖的量。
[0221]
另外,作为这些粗酶在60℃保温1小时后的残存活性,将酶液在60℃热处理1小时后,以表9的反应组成在60℃进行10分钟酶反应,测定残存活性。
[0222]
将反应液的组成表示于下述表9。
[0223]
[表9]
[0224][0225]
用上述方法,测定各变异体的反应最适温度和相对活性,选择在各变异体的反应最适温度下,d-阿洛酮糖异构酶活性比野生型酶高的氨基酸取代变异体。
[0226]
野生型酶是由重组制造的粗酶,d-阿洛糖生产的最适温度为60℃,纯化酶对l-鼠李糖的最适温度小于70℃,但也存在通过插入变异恢复最适温度的变异体。
[0227]
作为结果,得到了13个最适温度比野生型酶高或最适温度下的d-阿洛酮糖异构酶活性比野生型酶高的变异体(表10)。
[0228]
另外,作为耐热性的指标,与上述各变异体的反应最适温度和相对活性的测定同样地测定了各变异体在反应温度70℃和50℃时的d-阿洛酮糖异构酶活性之比(t70/t50)、在60℃保温1小时后的残存活性、以及在反应温度80℃和最适温度时的d-阿洛酮糖异构酶活性之比(t80/最适温度)。并且选择了任何一个指标比野生型酶高的变异体(表11~13)。
[0229]
其结果,取得了56个上述4个指标的任意一个比野生型酶高的变异体。将这56个变
异体的各氨基酸序列表示于序列表的序列编号3~8、10~28、30~60。具有序列编号9和29的氨基酸序列的变异体虽然具有活性,但4个指标都达不到野生型酶的水平。
[0230]
将这些具有序列表的序列编号3~60的氨基酸序列的各变异体命名为“序列3~60”。
[0231]
以下,分别表示序列3~60的从野生型酶(序列编号1)的氨基酸取代位点(位置)和取代氨基酸。该情况下,以下所示的氨基酸取代位点等与序列表的序列编号3~60所示的不同的情况下,序列表所示的氨基酸序列为正确的序列。另外,各序列的最后的括号内的数字表示氨基酸取代的数量。
[0232]
序列3 419g(1)
[0233]
4v280i(1)
[0234]
5v387i(1)
[0235]
6l198f(1)
[0236]
7s405e(1)
[0237]
8n52e(1)
[0238]
9q124r(1)
[0239]
10d102t(1)
[0240]
11q359l(1)
[0241]
12k152p(1)
[0242]
13d18a、f216l、t310s、l365q(4)
[0243]
14d18a、f216l、l365q(3)
[0244]
15y9f、d18a、v19i、i21v、q25k、t28a、h73d、a77s、i79v、t103k、e106d、q113k、h117n、q124t、p179a、i195v、a199g、e202q、f216l、d217n、a218p、s219a、a250s、l300i、n315h、n319d、s357t、l365s、a371g、l375m、l417q(31)
[0245]
16y9f、d18a、v19i、i21v、t28a、h73d、a77s、i79v、t103k、e106d、q113k、h117n、q124t、p179a、i195v、e202q、f216l、d217n、a218p、s219a、a250s、l300i、n315h、n319d、s357t、l365n、a371g、l375m、l417q(29)
[0246]
17d18a、v19i、i21v、t28a、h73d、a77s、i79v、t103k、e106d、q113k、h117n、q124t、p179a、i195v、e202q、f216l、d217n、a218p、s219a、a250s、l300i、n315h、n319d、s357t、l365s、a371g、l375m(27)
[0247]
18d18a、v19i、i21v、t28a、i79v、q113k、h117n、q124p、i195v、e202q、f216l、a218p、s219a、a250t、t310s、n315h、n319d、s357t、l365n、l417q(20)
[0248]
19d18a、v19i、i21v、t28a、h73d、t103k、q113k、h117n、q124n、h125n、e202q、f216l、d217n、a218p、s219q、a250t、n315h、n319d、l365n(19)
[0249]
20d18a、v19i、i21v、t28a、h73d、t103k、q113k、h117n、q124n、h125n、i195v、e202q、f216l、d217n、a218p、s219q、a250t、n315h、n319d、s357t、l365n(21)
[0250]
21y9f、d18a、v19i、i21v、m27l、t28g、g32r、n52q、e54a、h73d、i79l、k81q、t103k、e106a、a109e、q113e、h117n、q124n、k126q、s134t、h171y、i195v、e202q、v211i、f216l、d217n、a218p、s219q、a250t、t275a、t310n、n315q、k316n、n319d、k320r、s357t、d358a、k362q、l365n、e406d(40)
[0251]
22d18a、v19i、i21v、m27l、t28g、g32r、n52q、e54a、h73d、i79l、k81q、t103k、e106a、a109e、q113e、h117n、q124n、k126q、s134t、h171y、i195v、e202q、v211i、f216l、d217n、a218p、s219q、a250t、t275a、t310n、n315q、k316n、n319d、k320r、s357t、d358a、k362q、l365n、e406d(39)
[0252]
23y9f、d18a、v19i、i21v、t28g、n52q、e54a、h73d、i79l、t103k、q113e、h117n、q124n、s134t、h171y、i195v、e202q、f216l、d217n、a218p、s219q、a250t、t275a、t310n、n315q、n319d、s357t、k362q、l365n、e406d(30)
[0253]
24y9f、d18a、v19i、i21v、t28a、n52q、e54a、h73d、i79l、t103k、q113e、h117n、q124n、h125n、s134t、h171y、i195v、e202q、v211i、f216l、d217n、a218p、s219q、a250t、t310n、n315q、n319d、s357t、k362q、l365n、e406d(31)
[0254]
25d18a、v19i、i21v、t28a、n52q、e54a、h73d、i79l、t103k、q113e、h117n、q124k、s134t、h171y、i195v、e202q、v211i、f216l、d217n、a218p、s219q、a250s、t310n、n315q、n319d、s357t、l365n(27)
[0255]
26d18a、v19i、i21v、v26a、t28a、g32r、n52q、e54a、h73d、i79l、e80d、t103k、q113e、h117n、q124k、s134t、h171y、i195v、e202q、a205v、f216l、d217n、a218p、s219q、a250t、t310n、n315q、n319d、s357t、k362q、l365n(31)
[0256]
27l4q、y9f、v19i、i21v、v26a、m27i、t28r、g32r、n52q、e54a、h73d、a77s、e80d、t103k、q113a、s116a、h117n、e120a、q124e、l140m、a150s、g153t、i154v、h171y、l193i、i195v、e202q、a205m、f216l、d217n、a218p、s219q、t257c、t275a、t310s、a313v、n315h、k316q、n319d、a345t、s357t、q359r、k362q、l363a、l365n、a374t、s405g、e406d(48)
[0257]
28v19i、i21v、v26a、m27i、t28r、g32r、n52q、e54a、h73d、a77s、e80d、t103k、q113a、s116a、h117n、e120a、q124e、l140m、a150s、g153t、i154v、h171y、l193i、i195v、e202q、a205m、f216l、d217n、a218p、s219q、t257c、t275a、t310s、a313v、n315h、k316q、n319d、a345t、s357t、q359r、k362q、l363a、l365n、a374t、s405g、e406d(46)
[0258]
29d18e、v19n、i21v、e24d、q25l、t28a、g32e、e54q、h73d、m83i、k90m、d102s、t103q、q113k、h117n、e120a、q124s、h125n、k126r、s134t、p139a、l140k、k152t、r167h、h171y、l198f、e202q、e210a、v211i、k215q、f216l、d217n、a218p、s219q、a250s、t254i、t310h、n315q、n319d、q359r、k362r、l365s、y369f(43)
[0259]
30k3t、l4q、i5l、y9w、e10d、y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、a82t、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、l365a、e366n、a390m、w391y、l393q、h395n、v397a、d400g、a401s、s402q、s405d、e406n、h409a、q412e、q413d、t414v、r416s、l417q、419g(84)
[0260]
31k3t、l4q、i5l、y9w、e10d、y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、a82t、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、
t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、l365a、e366n、a390m、w391y、l393q、h395n、v397a、d400g、a401s、s402q、s405d、e406n、h409a、q412e、q413d、t414v、r416s、l417q、419g(84)
[0261]
32k3t、l4q、i5l、y9w、e10d、y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113g、s116t、h117n、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、l365a、e366n、a390m、w391y、l393q、h395n、v397a、d400g、a401s、s402q、s405d、e406n、h409a、q412k、q413d、t414v、r416s、l417q、419g(84)
[0262]
33k3t、l4q、i5l、y9w、e10d、d18a、i21v、e24d、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、e106a、d108n、a109e、e111k、q113a、s116k、h117n、e120a、q124a、h125n、k126q、s148a、d151n、k152d、g153d、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310n、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358s、q359a、r361k、k362q、l365e、e366n、a374v、s381t、a390m、w391y、l393q、h395n、v397t、d400g、a401s、s402q、s405d、e406n、h409m、q412k、q413d、t414v、r416s、l417q、419g(90)
[0263]
34l11i、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54s、r68k、h73g、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、d151n、k152d、g153e、i168v、h171y、v186i、l193i、i195v、l198f、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、v280i、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366d(60)
[0264]
35d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54s、r68k、h73g、i79l、k81q、m83l、a89p、i97l、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124t、h125n、d151n、k152n、g153e、i168v、h171y、v186i、l193i、i195v、l198f、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、v280i、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366d、s381c(61)
[0265]
36y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54a、r68k、h73t、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、d151n、k152d、g153e、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206d、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、k362q、l365a、e366d(57)
[0266]
37y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54a、r68k、h73t、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、d151n、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206d、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、k362q、l365a、e366d(59)
[0267]
38y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54a、r68k、
h73t、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、d151n、k152n、g153e、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366d(58)
[0268]
39y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54a、r68k、h73t、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、k152n、g153e、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366n(57)
[0269]
40y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54s、r68k、h73t、i79l、k81q、m83l、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152n、g153k、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206d、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313v、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366s(56)
[0270]
41y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54s、r68k、h73t、i79l、k81q、m83l、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152n、g153k、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313v、n315h、k316n、n319d、k320r、a345m、s357t、d358e、k362q、l365a、e366s(56)
[0271]
42y16f、d18a、i21v、v26a、m27l、t28p、g32r、i33l、r46a、n52q、e54a、r68k、h73t、i79l、k81q、a89p、d108n、e111k、q113e、s116k、h117n、q124e、k152n、g153d、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、k362q、l365a、e366n(50)
[0272]
43y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46s、n52e、e54s、r68k、h73s、i79l、k81q、s84r、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152n、g153s、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、r282q、t310s、a313v、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、l365a、e366a(57)
[0273]
44y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46s、n52e、e54s、r68k、h73s、i79l、k81q、s84r、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152n、g153s、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、a250t、t257c、t275a、r282q、t310s、a313v、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、l365a、e366a(57)
[0274]
45y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46s、n52e、e54s、r68k、h73s、a77t、i79l、k81q、s84r、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152n、g153s、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、r282q、t310s、a313v、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、l365a、e366a(59)
[0275]
46y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54a、r68k、
h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、k362q、l365a、e366d(61)
[0276]
47y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54a、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、k362q、l365a、e366d(62)
[0277]
48y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54a、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152n、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、k362q、l365a、e366n(62)
[0278]
49e10d、y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、k362q、l365a、e366n(65)
[0279]
50e10d、y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、k362q、l365a、e366n(64)
[0280]
51y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、s148a、k152n、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、r361k、k362q、l365a、e366n(63)
[0281]
52e10d、y16f、d18a、i21v、q25e、v26a m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、k126r、s148a、d151n、k152d、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357v、d358a、q359a、r361k、k362q、l365e、e366n(66)
[0282]
53y16f、d18a、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、
h73t、i79l、k81l、m83l、a89p、t103e、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124a、h125n、k126r、s148a、d151n、k152n、g153e、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357v、d358a、q359a、r361k、k362q、l365e、e366n(65)
[0283]
54e10d、d18a、i21v、e24d、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52g、e54s、r68k、h73t、i79l、k81l、m83l、a89p、e106a、d108n、a109e、e111k、q113a、s116k、h117n、e120a、q124a、h125n、k126q、s148a、d151n、k152d、g153d、c162v、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、v211a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316d、n319d、k320r、a345m、s357t、d358s、q359a、r361k、k362q、l365e、e366n、a374v、s381t(69)
[0284]
55k3t、l4q、i5l、y9w、l11i、d18a、v19i、i21v、v23a、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54s、r68k、h73q、i79l、k81q、m83l、a89p、d102t、e106a、d108n、a109e、e111k、q113e、s116k、q124r、k126q、d151n、k152p、g153e、h171y、l193i、i195v、l198f、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、t257c、t275a、v280i、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358a、q359l、k362q、l365n、e366d、v387i、a390m、w391y、l393q、v397t、d400g、a401s、s402q、s405e、e406n、h409t、q412k、q413d、t414v、r416s、l417q、419g(81)
[0285]
56k3t、l4q、i5l、y9w、d18a、v19i、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52d、e54s、r68k、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124p、k126q、d151n、k152e、g153e、h171y、l193i、i195v、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、a250t、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358a、k362q、l365n、e366d、a390m、w391y、l393q、v397t、d400g、a401s、s402q、s405d、e406n、h409t、q412k、q413d、t414v、r416s、l417q、419g(75)
[0286]
57k3t、l4q、i5l、y9w、d18a、v19i、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54t、r68k、h73n、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124g、k126q、d151n、k152e、g153e、h171y、l193i、i195v、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358a、q359l、k362q、l365n、e366d、a390m、w391y、l393q、v397t、d400g、a401s、s402q、s405d、e406n、h409t、q412k(71)
[0287]
58y9w、d18a、v19i、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54t、r68k、h73n、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124g、k126q、d151n、k152e、g153e、h171y、l193i、i195v、e202q、a205l、s206n、f216l、a218p、s219a、c245y、l246m、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358a、q359l、k362q、l365n、e366d、a390m、w391y、l393q、v397t、d400g、a401s、s402q、s405d、e406n、h409t、q412k(68)
[0288]
59y9w、d18a、v19i、i21v、q25e、v26a、m27l、t28r、g32r、i33l、r46a、n52e、e54t、r68k、h73n、i79l、k81q、m83l、a89p、e106a、d108n、a109e、e111k、q113e、s116k、h117n、q124g、k126q、d151n、k152e、g153e、h171y、l193i、i195v、e202q、a205l、s206n、f216l、
a218p、s219a、c245y、l246m、t257c、t275a、t310s、a313i、n315h、k316n、n319d、k320r、a345m、s357t、d358a、q359l、k362q、l365n、e366d、a390m、w391y、l393q、v397t、d400g、a401s、s402q、s405d、e406n、h409t(67)
[0289]
60k3t、l4q、i5l、y9w、y16f、d18a、q25e、v26a、m27l、t28r、g32r、i33l、r46s、n52e、e54s、r68k、h73s、i79l、k81q、s84r、a89p、e106s、a109q、e111k、q113e、s116k、h117n、q124a、h125n、k126q、k152d、g153s、i168v、h171y、v186i、l193i、i195v、e202q、a205l、s206a、f216l、d217n、a218p、s219a、c245y、l246m、a250t、t257c、t275a、r282q、t310s、a313v、n315h、k316d、n319d、k320r、a345m、s357t、d358a、q359e、l365a、e366p、a390m、w391y、l393q、v397t、d400g、a401s、s402e、s405e、e406s、h409a、q412k、q413e、t414i、r416s、l417r、419g(78)
[0290]
将这些总结一下,序列3~60的氨基酸取代变异体在以下的122个位点发生了特定的氨基酸取代。
[0291]
k3t、l4q、i5l、y9f/w、e10d、l11i、y16f、d18a/e、v19i/n、i21v、v23a、e24d、q25k/l/e、v26a、m27l/i、t28a/g/r/p、g32e/r、i33l、r46a/s、n52d/e/g/q、e54a/q/s/t、r68k、h73d/g/n/s/t/q、a77s/t、i79l/v、e80d、k81q/l、a82t、m83i/l、s84r、a89p、k90m、i97l、d102s/t、t103e/k/q、e106a/d/s、d108n、a109e/q、e111k、q113a/e/k/g、s116a/k/t、h117n、e120a、q124a/e/g/k/n/r/p/s/t、h125n、k126r/q、s134t、p139a、l140k/m、s148a、a150s、d151n、k152d/e/n/p/t、g153d/e/k/s/t、i154v、c162v、r167h、i168v、h171y、p179a、v186i、l193i、i195v、l198f、a199g、e202q、a205l/m/v、s206a/d/n、e210a、v211a/i、k215q、f216l、d217n、a218p、s219a/q、c245y、l246m、a250s/t、t254i、t257c、t275a、v280i、r282q、l300i、t310n/h/s、a313i/v、n315h/q、k316d/n/q、n319d、k320r、a345m/t、s357t/v、d358a/e/s、q359a/e/l/r、r361k、k362q/r、l363a、l365a/e/n/s/q、e366a/d/n/p/s、y369f、a371g、a374t/v、l375m、s381c/t、v387i、a390m、w391y、l393q、h395n、v397a/t、d400g、a401s、s402q/e、s405d/e/g、e406d/n/s、h409a/t/m、q412e/k、q413d/e、t414v/i、r416s、l417q/r、419g
[0292]
进而,序列3~60的氨基酸取代变异体中在以下的122个位点进行了氨基酸取代和1个氨基酸残基的添加。
[0293]
k3、l4、i5、y9、e10、l11、y16、d18、v19、i21、v23、e24、q25、v26、m27、t28、g32、i33、r46、n52、e54、r68、h73、a77、i79、e80、k81、a82、m83、s84、a89、k90、i97、d102、t103、e106、d108、a109、e111、q113、s116、h117、e120、q124、h125、k126、s134、p139、l140、s148、a150、d151、k152、g153、i154、c162、r167、i168、h171、p179、v186、l193、i195、l198、a199、e202、a205、s206、e210、v211、k215、f216、d217、a218、s219、c245、l246、a250、t254、t257、t275、v280、r282、l300、t310、a313、n315、k316、n319、k320、a345、s357、d358、q359、r361、k362、l363、l365、e366、y369、a371、a374、l375、s381、v387、a390、w391、l393、h395、v397、d400、a401、s402、s405、e406、h409、q412、q413、t414、r416、l417、419
[0294]
序列3~60中氨基酸取代的数量最多的序列33为90个,与序列1的野生型酶的氨基酸序列的同一性为78%。
[0295]
另外,在下述的表10中表示的最适温度下的d-阿洛酮糖异构酶活性比序列1的野生型酶高的13个序列中,氨基酸取代的数量最多的是序列32的84个,与序列1的氨基酸序列的同一性为80%。
[0296]
[表10]
[0297] 最适温度下的活性(u)最适温度序列32107.3270℃46104.9470℃5598.6260℃1397.0970℃2696.9270℃1596.1670℃5194.0970℃1783.7770℃1080.2670℃2180.0570℃2079.2370℃3777.3260℃4975.7970℃175.5460℃
[0298]
表11表示在反应温度70℃和50℃时的d-阿洛酮糖异构酶活性之比(t70/t50)比序列1的野生型酶高的52个序列的编号。
[0299]
[表11]
[0300]
序列t70/t50423.05272.07141.98561.91211.90481.87171.84521.83261.83581.82191.80391.79221.7681.75511.69461.6951.67571.67501.66
311.63401.63201.62181.61591.60411.59491.59231.58341.56541.55331.55251.55381.50441.48531.47431.46101.4471.43131.42151.4155140601.40471.36241.35371.33451.30351.29161.28301.27321.21121.21361.19281.1711.16
[0301]
表12表示在60℃保温1小时后的残存活性比序列1的野生型酶高的17个序列的编号。
[0302]
[表12]
[0303]
序列在60℃保温1小时的残存活性41115.2
22113.646113.036112.247110.46110.434109.528107.330107.240107.227107.042105.719105.417104.851104.521103.849103.61103.4
[0304]
表13表示在反应温度80℃和最适温度时的d-阿洛酮糖异构酶活性之比(t80/最适温度)比序列1的野生型酶高的56个序列的编号。
[0305]
[表13]
[0306]
序列t80/最适温度下的相对活性(%)4592.05183.95683.74382.63381.14679.94179.41776.64875.63173.13871.12268.62168.03667.33766.75266.33266.15063.7
4460.23959.74959.02058.85558.45356.84752.83052.32349.75949.01048.51947.41344.12843.4841.72640.24239.94038.41637.25835.91435.15434.83434.1533.41832.4631.31530.95730.52530.16028.81224.1724.02421.7319.62717.43514.0413.5118.317.7
[0307]
产业上的可利用性
[0308]
本发明的l-鼠李糖异构酶的特征在于,与来自微生物的现有的l-鼠李糖异构酶相比,特别是具有高的耐热性,且活性高。例如,相对于来自现有的施氏假单胞菌(pseudomonas stutzeri)的l-鼠李糖异构酶的最适温度为60℃,而本发明的酶高达70℃,且在60℃进行了10分钟加热处理后的残存活性也为80%以上,具有热稳定性,因此适于工业的生产上的使用。
[0309]
另外,在底物为d-阿洛酮糖的情况下,在60℃具有每mg蛋白质为2.26u的高的向d-阿洛糖的转换活性,通过本发明开拓了d-阿洛糖的大量生产的道路。
[0310]
另外,本发明的l-鼠李糖异构酶能够通过各种固定化方法得到活性高的固定化酶,通过使用固定化酶,能够连续地进行大量的异构化反应。通过能够在工业上固定化,能够实现目标醛糖的大量生产。
[0311]
进而,本发明的l-鼠李糖异构酶的氨基酸取代变异体中包括大量酶活性或耐热性比变异前的本酶高的变异体。
[0312]
因此,本发明的l-鼠李糖异构酶及其变异体、以及其制造方法的确立不仅在制糖产业,而且在与其相关的食品、化妆品、医药品、农药产业上的工业意义也极大。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。