结合siglec15的抗体及其用途
1.相关申请和引用并入
2.本技术要求2020年3月27日提交的美国临时专利申请us 63/000,566的优先权。
3.上述申请、在其中引用或在其审查过程中引用的全部文件(“申请引用文件”),在本文中引用或提及的所有文件(包括但不限于本文引用的所有文献、专利、公开的专利申请)(“本文引用文件”),在本文引用文件中引用或提及的所有文件,以及本文或任意本文引用并入文件中提及的任何制造商手册、说明书、产品规格和产品页,均通过引用的方式并入本文,且可能在实施本发明时采用。更具体而言,所有参考文件均通过引用的方式并入,如同各文件具体且个别地通过引用的方式并入。在本公开中提及的任何genbank序列通过引用的方式并入,这些genbank序列为本公开最早有效递交日的序列。
发明领域
4.本技术大体涉及分离的单克隆抗体,例如全人、小鼠、嵌合或人源化单克隆抗体或其抗原结合部分,其以高亲和力和功能特异性与siglec15特异结合。还提供编码该抗体或其抗原结合部分的核酸分子,用于表达该抗体或其抗原结合部分的表达载体、宿主细胞以及方法。本技术还提供包含该抗体或其抗原结合部分的免疫偶联物、双特异性分子、嵌合抗原受体、溶瘤病毒、和药物组合物,以及使用本技术siglec15抗体的诊断和治疗方法。
背景技术:
5.免疫疗法是革命性的治疗手段,其激活免疫系统来对抗疾病例如癌症。其适用于多种适应症,且与其他标准治疗相比,毒性较低。pd-1/pd-l1通路是肿瘤免疫治疗中的热门靶点,多个pd-1或pd-l1抑制剂已经被临床批准,例如pd-1抗体和以及pd-l1抗体然而,一些患者对这种治疗没有反应。正如最新的报道所揭示的,对pd-1/pd-l1靶向治疗无反应的癌症患者来说,靶向siglec15可能是一种补充方案(jun wang et al.,(2019)nature medicine 25:656-666)。
6.siglec15是siglec家族的成员,其具有唾液酸结合免疫球蛋白型凝集素结构。它包含两个胞外免疫球蛋白样结构域、具有对与衔接蛋白dap12相互作用必要的赖氨酸残基的跨膜结构域、和胞质尾部(takashi angata et al.,(2007)glycobiology 17(8):838-846)。
7.siglec15表达在破骨细胞上,在破骨细胞的分化和骨重塑中发挥作用(hiruma y et al.,(2011)biochemical and biophysical research communications 409(3):424-429;takashi angata(2020)journal of biomedical science 27:10)。siglec15抗体的施用抑制破骨细胞骨吸收,并在啮齿动物模型中增加骨量(stuible m et al.,(2014)journal of biological chemistry 289(10):6498-6512;sato d et al.,(2018)bone 116:172-180)。
8.siglec15也表达在肿瘤相关巨噬细胞上,优先识别唾液酸-tn抗原,一种肿瘤相关糖类结构。唾液酸-tn-肿瘤细胞系与m-csf诱导的人巨噬细胞或siglec15
骨髓细胞系的共
培养,诱导转化生长因子-β的产生,促进肿瘤细胞的上皮间质转化和转移(takamiya r et al.,(2013)glycobiology 23(2):178-187)。lieping cheng等人最近发现,在非小细胞型肺癌的临床样品中,siglec15还在肿瘤细胞和/或肿瘤相关基质细胞上进一步表达。他们还发现,siglec15蛋白抑制t细胞增殖和激活,且siglec15抗体在体内逆转t细胞抑制及减弱肿瘤生长。如上所述,由于siglec15和pd-l1在肿瘤组织中是互斥的,siglec15可以作为补充性的治疗靶点并为耐受pd-1/pd-l1阻断的患者提供可选的治疗方案(jun wang et al.,(2019)同上)。nc318,人源化的siglec15抗体,已经在患有晚期实体瘤的患者中进行临床试验,包括非小细胞肺癌、卵巢癌、黑色素瘤、结直肠癌和乳癌,在54%的患者中观察到疾病稳定期得到延长,且客观应答率为5.4%(sun j et al.,(2021)clin cancer res.27(3):680-688)。
9.总观siglec15在骨重塑和肿瘤发展中的参与性,siglec15绝对是逐渐显露的有希望的治疗靶点。需要具有改进的药学特性的siglec15抗体。
10.本技术中任何文件的引用或识别,并不是承认这些文件是本技术的现有技术。
技术实现要素:
11.本技术提供分离的小鼠源、嵌合、人源或人源化的单克隆抗体或其抗原结合部分,其结合siglec15(例如,人siglec15和猴siglec15),与现有技术siglec15抗体例如siglec15-ch5g9(nextcure)相比,具有相当的,如果不是更高的话,与siglec15的结合亲和力/能力以及对siglec15与配体例如lrrc4c结合的阻断活性。抗体或其抗原结合部分能够逆转siglec15介导的t细胞抑制。
12.本技术的抗体或其抗原结合部分可以用于多种应用,包括siglec15蛋白的检测以及siglec15相关疾病(例如肿瘤和骨质疏松症)的治疗和预防。
13.因此,在一个方面,本技术涉及结合siglec15的分离的单克隆抗体(例如人源、小鼠源、嵌合或人源化的抗体)或其抗原结合部分,其具有i)重链可变区,其可以包含vh cdr1区、vh cdr2区和vh cdr3区,其中该vh cdr1区、vh cdr2区和vh cdr3区可以分别包含与(1)seq id nos:1、2(x1=d、x2=q)和3;(2)seq id nos:1、2(x1=e、x2=q)和3;(3)seq id nos:1、2(x1=d、x2=k)和3;(4)seq id nos:9、10和11;或(5)seq id nos:33、34和35具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列;和/或ii)轻链可变区,其可以包含vl cdr1区、vl cdr2区和vl cdr3区,其中该vl cdr1区、vl cdr2区和vl cdr3区可以分别包含与(1)seq id nos:4、5和6;(2)seq id nos:12、13和14;或(3)seq id nos:36、37和38具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列。
14.本技术的抗体或其抗原结合部分可以包含重链可变区和轻链可变区,重链可变区可以包含vh cdr1区、vh cdr2区和vh cdr3区,轻链可变区可以包含vl cdr1区、vl cdr2区和vl cdr3区,其中vh cdr1区、vh cdr2区、vh cdr3区、vl cdr1区、vl cdr2区和vl cdr3区可以分别包含与(1)seq id nos:1、2(x1=d、x2=q)、3、4、5和6;(2)seq id nos:1、2(x1=e、x2=q)、3、4、5和6;(3)seq id nos:1、2(x1=d、x2=k)、3、4、5和6;(4)seq id nos:9、10、11、12、13和14;或(5)seq id nos:33、34、35、36、37和38具有至少85%、86%、87%、88%、
89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,其中该抗体或其抗原结合部分与siglec15结合。
15.本技术的抗体或其抗原结合部分的重链可变区可以包含与seq id nos:7(x1=d、x2=q;x1=e、x2=q;或x1=d、x2=k)、15、39、40或41具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,其中该抗体或其抗原结合部分与siglec15结合。seq id no:7(x1=d、x2=q)的氨基酸序列可由seq id no:27的核苷酸序列编码,seq id no:15的氨基酸序列可由seq id no:29的核苷酸序列编码。
16.本技术的抗体或其抗原结合部分的轻链可变区可以包含与seq id nos:8、16、42或43具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,其中该抗体或其抗原结合部分与siglec15结合。seq id nos:8和16的氨基酸序列可由seq id nos:28和30的核苷酸序列编码。
17.本技术的抗体或其抗原结合部分可以包含重链可变区和轻链可变区,其可以分别包含与(1)seq id nos:7(x1=d、x2=q)和8;(2)seq id nos:7(x1=e、x2=q)和8;(3)seq id nos:7(x1=d、x2=k)和8;(4)seq id nos:15和16;(5)seq id nos:39和42;(6)seq id nos:40和42;或(7)seq id nos:41和43具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,其中该抗体或其抗原结合部分与siglec15结合。
18.本技术分离的单克隆抗体或其抗原结合部分可以包含经二硫键连接的重链和轻链,重链可以包含重链可变区和重链恒定区,轻链可以包含轻链可变区和轻链恒定区,其中重链可变区的c端与重链恒定区的n端相连,轻链可变区的c端与轻链恒定区的n端相连,其中重链可变区和轻链可变区可以包含上述氨基酸序列,该抗体或其抗原结合部分与siglec15结合。重链恒定区可以是igg1、igg2或igg4重链恒定区,例如人igg1、igg或igg4重链恒定区,具有seq id no:17所示氨基酸序列的。重链恒定区,例如fc片段,可以加工成具有降低或增强的fcr结合亲和力。轻链恒定区可以是κ恒定区,例如具有seq id no:18所示氨基酸序列的人κ恒定区。seq id nos:17和18的氨基酸序列可分别由seq id nos:31和32的核苷酸序列编码。
19.在一些实施方式中,本技术的抗体可以包含两条重链和两条轻链,或者由两条重链和两条轻链组成,其中各重链可以包含上述重链恒定区、重链可变区或cdr序列,各轻链可以包含上述轻链恒定区、轻链可变区或cdr序列,其中抗体与siglec15结合。本技术的抗体或其抗原结合部分可以是全长抗体,例如igg1、igg2或igg4同种型的。在其他实施方式中,本技术的抗体或其抗原结合部分可以是单链可变片段(scfv)抗体或抗体片段,例如fab或f(ab
′
)2片段。
20.本技术还提供一种双特异性分子,其可以包含本技术的抗体或其抗原结合部分,与具有不同于本抗体或其抗原结合部分的结合特异性的第二功能基团(例如,第二抗体)连接。本技术还提供免疫偶联物,例如抗体-药物偶联物,其可以包含本技术的抗体或其抗原结合部分,与治疗剂例如细胞毒素连接。在另一方面,本技术的抗体或其抗原结合部分可以制备成嵌合抗原受体(car)的一部分。还提供包含该抗原嵌合受体的免疫细胞,例如t细胞和nk细胞。本技术的抗体或其抗原结合部分也可以由溶瘤病毒编码或与溶瘤病毒一起使
用。
21.本技术还涉及编码本技术抗体或其抗原结合部分的核酸分子,以及包含该核酸的表达载体、和包含该表达载体的宿主细胞。本技术还提供利用该宿主细胞制备siglec15抗体或其抗原结合部分的方法,其可以包括以下步骤:(i)在该宿主细胞中表达抗体,以及(ii)从宿主细胞或其细胞培养物中分离抗体。
22.还提供组合物,其可以包含本技术的抗体或其抗原结合部分、免疫偶联物、双特异性分子、溶瘤病毒、car、car-t细胞、核酸分子、表达载体或宿主细胞,以及药学上可接受的载体。在一些实施方式中,药物组合物还可以包含例如抗癌剂的治疗剂。
23.在另一方面,本技术提供一种在受试者中调节免疫应答的方法,包括向受试者施用治疗有效量的本技术抗体或其抗原结合部分、或者能够在受试者中表达这些的核酸分子,从而调节受试者中的免疫应答。优选地,本技术的抗体或其抗原结合部分加强、刺激或增加受试者的免疫应答,例如通过逆转siglec15介导的t细胞抑制。在一些实施方式中,该方法包括施用本技术的双特异性分子、免疫偶联物、car-t细胞、或者编码或携带抗体的溶瘤病毒。
24.在另一方面,本技术提供一种在受试者中抑制骨流失或者增加骨密度的方法,包括向受试者施用治疗有效量的本技术抗体或其抗原结合部分、或者能够在受试者中表达这些的核酸分子。
25.在再一个方面,本技术提供一种在有需求受试者中抑制肿瘤生长的方法,包括向受试者施用治疗有效量的本技术抗体或其抗原结合部分、或者能够在受试者中表达这些的核酸分子。在一些实施方式中,该方法包括施用本技术的双特异性分子、免疫偶联物、car-t细胞、或者编码或携带抗体的溶瘤病毒。肿瘤可以是实体瘤或非实体瘤。在一些实施方式中,肿瘤是实体瘤,包括但不限于,非小细胞肺癌、卵巢癌、黑色素瘤、结直肠癌、乳癌(包括三阴性乳癌)、头颈部鳞状细胞瘤、子宫内膜癌、和鳞状细胞癌。在一些实施方式中,至少一种其他的抗癌抗体可以与本技术抗体或其抗原结合部分一起施用,例如vista抗体、pd-1抗体、pd-l1抗体、lag-3抗体、ctla-4抗体、tim-3抗体、stat3抗体、和/或ror1抗体。在另一实施方式中,本技术的抗体或其抗原结合部分与细胞因子(例如il-2、il-21、gm-csf和/或il-4)或共刺激抗体(例如cd137和/或gitr抗体)一起施用。在其他实施方式中,本技术的抗体或其抗原结合部分与化疗剂一起施用,其可以是细胞毒性剂例如表阿霉素、奥沙利铂和/或5-氟尿嘧啶(5-fu)。本技术的抗体或其抗原结合部分可以是例如小鼠源、人源、嵌合的或人源化的。
26.基于以下具体描述和实施例,当前公开内容的其他特征和优势之处将会更加明晰,具体描述和实施例不应解读为限制性的。在本技术中引用的所有文献、genbank记录、专利和已公开专利申请的内容通过引用的方式明确地包含在本文中。
27.另外,本技术的目标是不在本技术中包含任何先前已知的产品、制造该产品的工艺或使用该产品的方法,从而申请人保留权利,并在此公开对任何先前已知的产品、过程或方法的弃权声明。需要进一步指出的是,本技术并不打算在本技术的范围内包含任何不符合uspto(35 u.s.c.
§
112,第一段)或epo(epc,第83条)书面描述要件和可实施性要求的产品、工艺、或产品制造方法或产品使用方法,从而申请人保留权利,并在此公开对任何先前描述的产品、产品制备工艺、或产品使用方法的弃权声明。在本发明的实施中,符合epc第53
条(c)和细则第28条(b)和(c)是有利的。明确保留对本技术同族或任何其他同族或任何第三方在先申请中涉及本技术人任何已授权专利的主题的任何实施方式做出明确的弃权声明的所有权利。本文中的任何内容都不应被解释为承诺。
28.应当注意的是,在本技术中,特别是在权利要求和/或段落中,术语例如“包含”、“包括”等可以具有美国专利法所赋予的意义;例如它们可以表示“包含在内”等;而术语例如“基本由...组成”或“基本由...构成”具有美国专利法所赋予的意义,例如允许没有明确表述的元素的存在,但将现有技术中存在的元素、或影响本发明基本或新特性的元素排除在外。
附图说明
29.以下以示例方式给出详细的描述,但不意在将本技术限制于所述具体实施方式的具体描述,可以结合附图更好地进行理解。
30.图1a-1c示出在捕获elisa中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)与人siglec15的结合力。
31.图2a-2c示出在基于细胞的结合facs检测中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)与表达人siglec15的人siglec15-2d3-1e1细胞的结合力。
32.图3a-3c示出在间接elisa中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)与猴siglec15的结合力。
33.图4a-4c示出在间接elisa中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、ale7g5d1、ale10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)与小鼠siglec15的结合力。
34.图5a-5c示出在竞争elisa中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)阻断人siglec15-lrrc4c结合的能力。
35.图6a-6c示出在竞争elisa中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)阻断对照基准物-人siglec15结合的能力。
36.图7a-7c示出在基于细胞的阻断facs检测中抗体a1c8c6h1、a1d1b7h9、a1d5e2h1和a1d11a7h10(a)、a1e7g5d1、a1e10g7h9、a2a1d2f1和a2a5c7e8(b)、a2a6b1c2、a2g4c8g7和a2h5f1a1(c)阻断人siglec15与细胞表面人lrrc4c结合的能力。
37.图8a和8b示出在基于细胞的功能检测中抗体a2a5c7e8和a1e10g7h9逆转了siglec15诱导的cd8
细胞(a)和cd4
细胞(b)抑制。
38.图9示出在捕获elisa中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3对人siglec15的结合力。
39.图10示出在基于细胞的结合facs检测中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3对表达人siglec15的人siglec15-2d3-1e1细胞的结合力。
40.图11示出在间接elisa中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3对猴siglec15的结合力。
41.图12示出在间接elisa中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3对小鼠siglec15的结合力。
42.图13示出在竞争elisa中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3阻断人siglec15-lrrc4c结合的能力。
43.图14示出在竞争elisa中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3阻断对照基准物与人siglec15结合的能力。
44.图15a-15d示出在基于细胞的功能检测中抗体a2a5c7e8-1、a2a5c7e8-2和a2a5c7e8-3逆转了由93.5nm siglec15诱导的cd8
细胞(a)和cd4
细胞(b)抑制,以及由186.9nm siglec15诱导的cd8
细胞(c)和cd4
细胞(d)抑制。
45.图16示出在捕获elisa中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7对人siglec15的结合力。
46.图17示出在基于细胞的结合facs检测中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7对表达人siglec15的人siglec15-2d3-1e1细胞的结合力。
47.图18示出在间接elisa中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7对猴siglec15的结合力。
48.图19示出在间接elisa中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7与小鼠siglec15的结合力。
49.图20示出在竞争elisa中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7阻断人siglec15-lrrc4c结合的能力。
50.图21示出在竞争elisa中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7阻断对照基准物与人siglec15结合的能力。
51.图22a和22b示出在基于细胞的功能检测中小鼠源抗体b2d7h7a3c1、b2g12h3e8和b2h2h1h7逆转了siglec15介导的cd8
细胞(a)和cd4
细胞(b)抑制。
具体实施方式
52.为确保更容易地理解本技术,首先定义一些术语。其他的定义贯穿具体描述而给出。
53.术语“siglec15”包括变体、亚型、同系物、直系同源物和旁系同源物。例如,对人siglec15蛋白特异的抗体可能在某些情况下与来自人以外物种(例如猴)的siglec15蛋白交叉反应。在其他实施方式中,对人siglec15蛋白特异的抗体可能完全对人siglec15蛋白特异,而对其他物种或其他类型表现出无交叉反应性,或者可能与来自某些其他物种但不是所有其他物种的siglec15交叉反应。
54.术语“人siglec15”是指具有来自人的氨基酸序列的siglec15蛋白,例如genbank登录号为q6zmc9的人siglec15的氨基酸序列。术语“猴或恒河猴siglec15”和“小鼠siglec15”分别是指猴和小鼠的siglec15序列,例如具有genbank登录号分别为xp_028694069.1和np_001094508.1的氨基酸序列的那些。
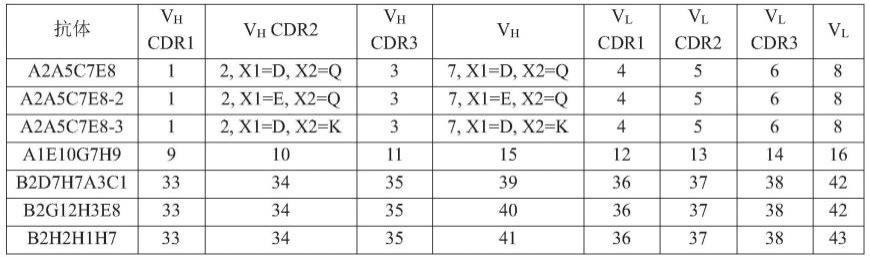
55.术语“免疫应答”是指例如淋巴细胞、抗原递呈细胞、吞噬细胞、粒细胞和由上述细
胞或肝脏产生的可溶性大分子(包括抗体、细胞因子和补体),引发对于入侵病原体、被病原体感染的细胞或组织、癌细胞,或在自身免疫或病理性炎症情况下对于正常人体细胞或组织的选择性损坏、破坏或从人体中消除的过程。
56.本文所用的术语“抗体”是指通过至少一个抗原结合位点识别和特异性结合靶标(例如siglec 15)的免疫球蛋白分子,其中抗原结合位点通常位于免疫球蛋白分子的可变区内。如本文所用的,术语包括完整的多克隆抗体、完整的单克隆抗体、单链fv(scfv)抗体、重链抗体(hcab)、轻链抗体(lcab)、多特异性抗体、双特异性抗体、单特异性抗体、单价抗体、包含抗体的抗原结合位点的融合蛋白、以及任何其他含抗原结合位点的修饰免疫球蛋白分子(例如:双可变结构域免疫球蛋白分子),只要抗体展现出所需的生物活性。抗体还包括,但不仅限于,小鼠源抗体、嵌合抗体、人源化抗体和人源抗体。基于分别称为α、δ、ε、γ和μ的其重链恒定结构域的特征,抗体可以是五类主要免疫球蛋白类iga、igd、ige、igg和igm或其亚类(同种型)(例如:igg1、igg2、igg3、igg4、iga1和iga2)中的任意种。不同种类的免疫球蛋白具有不同的众所周知的亚基结构和三维构象。抗体可以是裸露的或与其他分子偶联,包括但不限于毒素和放射性同位素。除非另有明确说明,本文所用的术语“抗体”包括完整抗体的“抗原结合部分”。igg是可以包含经二硫键而内部连接的两条重链(h)和两条轻链(l)的糖蛋白。各重链可以由重链可变区(缩写成vh)和重链恒定区组成。重链恒定区可以由c
h1
、c
h2
和c
h3
这三个结构域组成。各轻链可以由轻链可变区(本文中缩写成v
l
)和轻链恒定区组成。轻链恒定区可以由cl这一个结构域组成。vh和v
l
区可以进一步细分为高变区,称为互补决定区(cdr),其间穿插着更加保守的称为骨架区(fr)的区域。各vh和v
l
由三个cdr和四个fr组成,从氨基端到羧基端以fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4的顺序排布。重链和轻链的可变区包含与抗原相互作用的结合域。抗体的恒定区可以介导免疫球蛋白与宿主组织或因子,包括多种免疫系统细胞(例如,效应细胞)和传统补体系统的第一组分(c1q),的结合。
57.本文中所用的术语,抗体的“抗原结合部分”(或简称为“部分抗体”),是指抗体上保留有特异性结合抗原(例如siglec15蛋白)能力的一个或多个片段。已显示出,抗体的抗原结合功能可以通过全长抗体的片段来实施。包含在术语抗体的“抗原结合部分”中的结合片段的例子包括(i)fab片段,由v
l
、vh、c
l
和c
h1
结构域构成的单价片段;(ii)f(ab
′
)2片段,包含由铰链区二硫键连接的两个fab片段的二价片段;(iii)由vh和c
h1
结构域构成的fd片段;(iv)由抗体单臂的v
l
和vh结构域构成的fv片段;(v)由vh结构域构成的dab片段(ward et al.,(1989)nature 341:544-546);(vi)分离的互补决定区(cdr);以及(viii)纳米抗体,包含单个可变结构域的重链可变区和两个恒定结构域。此外,尽管fv片段的两个结构域,v
l
和vh,由不同的基因编码,它们可以通过重组的方法经合成接头而连接,其中合成的接头使得它们可以制备为单个蛋白链,其中v
l
和vh区配对形成单价分子(称为单链fv(scfv);参见例如bird et al.,(1988)science 242:423-426;和huston et al.,(1988)proc.natl.acad.sci.usa 85:5879-5883)。这样的单链抗体也意在包括在术语抗体的“抗原结合部分”中。这些抗体片段通过本领域技术人员已知的常用技术而得到,且片段经与完整抗体相同的方式进行应用筛选。
58.本文所用的“分离的抗体”是指基本不含具有不同抗原特异性的其他抗体的抗体(例如,分离出的与siglec15蛋白特异性结合的抗体,基本不含特异结合siglec15之外蛋白的抗体)。然而,分离出的与人siglec15蛋白特异性结合的抗体可以与其他抗原,例如其他
物种的siglec15蛋白,有交叉反应性。此外,分离的抗体可以基本不含其他细胞材料和/或化学物质。
59.本文所用的术语“单克隆抗体”或“单克隆抗体组成”是指单分子组成的抗体分子的制品。单克隆抗体组成表现出对于特定表位的单结合特异性和亲和力。
60.本文所用的术语“人源抗体”意在包括骨架和cdr区得自人种系免疫球蛋白序列的可变区的抗体。此外,如果抗体包含恒定区,恒定区也得自人种系免疫球蛋白序列。本技术的人源抗体可以包含不由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机突变或点突变或通过体内体细胞突变而引入的突变)。然而,本文所用的术语“人源抗体”不意在包括在人骨架序列中植入得自另一哺乳动物物种种系的cdr序列的抗体。
61.本文所用的术语“小鼠源抗体”意在包括骨架和cdr区得自小鼠种系免疫球蛋白序列的可变区的抗体。此外,如果抗体包含恒定区,恒定区也得自小鼠种系免疫球蛋白序列。本技术的小鼠源抗体可以包含不由小鼠种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机突变或点突变或通过体内体细胞突变而引入的突变)。然而,本文所用的术语“小鼠源抗体”不意在包括在小鼠骨架序列中植入得自另一哺乳动物物种种系的cdr序列的抗体。
62.术语“嵌合抗体”是指通过组合非人源遗传物质与人源遗传物质而制备的抗体。或者更笼统地说,嵌合抗体是含有某一物种的遗传物质以及另一物种遗传物质的抗体。
63.本文所用的术语“人源化抗体”是指来源于非人物种但其蛋白序列已经经修改而增加与人中天然生成的抗体变体的相似度的抗体。
64.术语“同种型”是指由重链恒定区基因编码的抗体类别(如igm或igg1)。
65.词组“识别抗原的抗体”和“对抗原特异的抗体”在本文中与术语“特异结合抗原的抗体”交替使用。
66.如本文所用的,“特异结合人siglec15”的抗体是指与人siglec15蛋白(以及可能的来自一种或多种非人物种的siglec15蛋白)结合但基本不与非siglec15蛋白结合的抗体。优选地,抗体以“高亲和力”,即以kd值为5.0x10-9
m以下、更优选1.0x10-9
m以下、更优选1.0x10-10
m以下,结合人siglec15蛋白。
67.如本文所用的,术语“基本不结合”蛋白或细胞是指,不与蛋白或细胞结合,或者不以高亲和力与其结合,即,以kd为1x10-6
m以上、更优选1x10-5
m以上、更优选1x10-4
m以上、更优选1x10-3
m以上、更优选1x10-2
m以上,结合蛋白或细胞。
68.术语“高亲和性”对于igg抗体而言,是指抗体对于靶抗原的kd为5.0x10-9
m以下、更优选1.0x10-9
m以下、更优选5.0x10-10
m以下、甚至更优选1.0x10-10
m以下,甚至更优选5.0x10-11
m以下。然而,对于其他抗体同种型而言,“高亲和性”结合可以会变化。例如,对于igm同种型的“高亲和性”结合是指抗体的kd为10-6
m以下、更优选10-7
m以下、甚至更优选10-8
m以下。
69.本文所用术语“k
assoc”或“k
a”是指特定抗体-抗原相互作用的结合速率,而本文所用的术语“k
dis”或“k
d”是指特定抗体-抗原相互作用的解离速率。如本文所用,术语“k
d”是指从kd与ka的比(即kd/ka)获得的解离常数,表达为摩尔浓度(m)。可以使用本领域公知的方法确定抗体的kd值。用于确定抗体kd的优选方法是使用表面等离子体共振,优选使用生物传感器系统如biacore
tm
系统。
70.术语“ec
50”,也称为半最大效应浓度,是指在特定暴露时间后引起在基线和最高值之间的中间值反应的抗体浓度。
71.术语“ic
50”,也称为半最大抑制浓度,是指相对于不存在抗体而言,抑制50%特定生物学或生化功能的抗体浓度。
72.术语“受试者”包括任何人或非人动物。术语“非人动物”包括所有脊椎动物,例如哺乳类和非哺乳类,例如非人灵长类、羊、狗、猫、牛、马、鸡、两栖类、和爬行类,尽管优选哺乳动物,例如非人灵长类、羊、狗、猫、牛和马。
73.术语“治疗有效量”是指足以防止或减缓与疾病或病症(例如癌症)相关的症状和/或减轻疾病或病症的严重程度的本技术抗体的用量。治疗有效量在所治疗疾病的背景下进行理解,其中本领域技术人员可以方便地判别出实际有效量。
74.本技术的多个方面在以下小节中作进一步详细描述。
75.本技术的抗体或其抗原结合部分与人或猴siglec15特异性结合,结合亲和力与之前所述的siglec15抗体例如siglec15-ch5g9相当,如果不是更佳的话。
76.其他的功能性特征包括阻断siglec15与其配体结合、以及逆转siglec15介导的t细胞抑制的能力。
77.本技术的示例性抗体或其抗原结合部分在结构上和化学上如下文和实施例中表征。抗体的重/轻链可变区的氨基酸序列id编号总结在表1中。抗体的重链恒定区可以是具有例如seq id no:17所示的氨基酸序列的人igg1重链恒定区,抗体的轻链恒定区可以是具有例如seq id no:18所示氨基酸序列的人κ恒定区。本技术的抗体可以是人源、小鼠源、嵌合或人源化的抗体。
78.表1中的重链可变区cdr和轻链可变区cdr经kabat编号体系确定。然而,如本领域所公知的,cdr区也可以基于重链/轻链可变区序列经其他体系例如chothia、imgt、abm或contact编号体系/方法确定。
79.表1.重链/轻链可变区的氨基酸序列id编号
[0080][0081]
结合人siglec15的其他siglec15抗体的vh和v
l
序列(或cdr序列)可以与本公开的siglec15抗体的vh和v
l
序列(或cdr序列)“混合和匹配”。优选地,当vh和v
l
链(或这些链中的cdr)混合并匹配时,来自特定vh/v
l
配对的vh序列被结构上相似的vh序列代替。同样地,优选来自特定vh/v
l
配对的v
l
序列经结构类似的v
l
序列替换。
[0082]
因此,在一个实施方式中,本技术的抗体或其抗原结合部分包括:
[0083]
(a)包含表1中所列氨基酸序列的重链可变区;和
[0084]
(b)包含表1中所列氨基酸序列的轻链可变区、或另一siglec15抗体的v
l
,其中该抗体特异性结合人siglec15。
[0085]
在另一实施方式中,本技术的抗体或其抗原结合部分包括:
[0086]
(a)列于表1中的重链可变区的cdr1、cdr2和cdr3;以及
[0087]
(b)列于表1中的轻链可变区的cdr1、cdr2和cdr3,或者另一siglec15抗体的cdr,其中该抗体特异结合人siglec15。
[0088]
在另一实施方式中,抗体或其抗原结合部分包括siglec15抗体的重链可变cdr2区,结合有其他结合人siglec15抗体的cdr区,例如,不同siglec15抗体的重链可变区cdr1和/或cdr3、和/或轻链可变区cdr1、cdr2和/或cdr3。
[0089]
另外,如本领域所公知的,cdr3结构域,独立于cdr1和/或cdr2结构域,可以单独决定抗体对同源抗原的结合特异性,且基于相同的cdr3序列,可预见能产生具有相同结合特异性的多个抗体。参见例如klimka et al.,british j.of cancer 83(2):252-260(2000);beiboer et al.,j.mol.biol.296:833-849(2000);rader et al.,proc.natl.acad.sci.u.s.a.95:8910-8915(1998);barbas et al.,j.am.chem.soc.116:2161-2162(1994);barbas et al.,proc.natl.acad.sci.u.s.a.92:2529-2533(1995);ditzel et al.,j.immunol.157:739-749(1996);berezov et al.,biajournal 8:scientific review 8(2001);igarashi et al.,j.biochem(tokyo)117:452-7(1995);bourgeois et al.,j.virol 72:807-10(1998);levi et al.,proc.natl.acad.sci.u.s.a.90:4374-8(1993);polymenis and stoller,j.immunol.152:5218-5329(1994)以及xu and davis,immunity 13:37-45(2000)。也可参见美国专利6,951,646;6,914,128;6,090,38;6,818,216;6,156,313;6,827,925;5,833,943;5,762,905和5,760,185。这些参考文献中的每一个都通过引用的方式整体并入本文。
[0090]
因而,在另一个实施方式中,本技术的抗体包含siglec15抗体重链可变区的cdr2,至少地siglec15抗体重链和/或轻链可变区的cdr3,或另一siglec15抗体重链和/或轻链可变区的cdr3,其中该抗体能够与人siglec15特异性结合。优选这些抗体(a)竞争结合siglec15;(b)保留功能特性;(c)结合相同表位;和/或(d)具有与本技术siglec15抗体相似的结合亲和力。在另一实施方式中,抗体还可以包含siglec15抗体轻链可变区的cdr2、或另一siglec15抗体轻链可变区的cdr2,其中该抗体能够与人siglec15特异性结合。在另一实施方式中,本技术的抗体可以包含siglec15抗体的重链和/或轻链可变区的cdr1,或另一siglec15抗体的重链和/或轻链可变区的cdr1,其中该抗体能够与人siglec15特异性结合。
[0091]
在另一个实施方式中,本技术的抗体包含cdr1、cdr2和cdr3序列的重链和/或轻链可变区序列,其与本技术的siglec15抗体的可变区相比,区别在于具有一个或多个保守修饰。如本领域所理解的,可以进行一些不消除抗原结合的保守序列修饰。参见例如brummell et al.,(1993)biochem 32:1180-8;de wildt et al.,(1997)prot.eng.10:835-41;komissarov et al.,(1997)j.biol.chem.272:26864-26870;hall et al.,(1992)j.immunol.149:1605-12;kelley and o
′
connell(1993)biochem.32:6862-35;adib-conquy et al.,(1998)int.immunol.10:341-6和beers et al.,(2000)clin.can.res.6:2835-43。
[0092]
因而,在一个实施方式中,抗体包含具有cdr1、cdr2和cdr3序列的重链可变区和/或具有cdr1、cdr2和cdr3序列的轻链可变区,其中:
[0093]
(a)重链可变区cdr1序列包含如上表1所列的序列,和/或其保守修改;和/或
[0094]
(b)重链可变区cdr2序列包含如上表1所列的序列,和/或其保守修改;和/或
[0095]
(c)重链可变区cdr3序列包含如上表1所列的序列,及其保守修改;和/或
[0096]
(d)轻链可变区cdr1、和/或cdr2、和/或cdr3序列包含如上表1所列的序列,和/或其保守修改;和
[0097]
(e)该抗体特异结合人siglec15。
[0098]
本技术的抗体或其抗原结合部分具有以下一个或多个上述的功能特点,例如与人siglec15的高结合亲和力、以及逆转siglec15介导的t细胞抑制的能力。
[0099]
在多个实施方式中,抗体或其抗原结合部分可以是例如小鼠源、人源、人源化或嵌合抗体。
[0100]
本文所用的术语“保守的序列修饰”是指不会显著影响或改变含有这类氨基酸序列的抗体的结合特性的氨基酸修饰。这样的保守修饰包括氨基酸替换、添加和缺失。可以通过领域内已知的标准技术,例如点突变和pcr介导的突变,将修饰引入本技术抗体中。保守氨基酸替换是指将氨基酸残基用具有相似侧链的氨基酸残基替换。已定义出领域内具有相似侧链的氨基酸残基组。这些组包括具有碱性侧链(例如,赖氨酸、精氨酸、组氨酸)、酸性侧链(例如,天冬氨酸、谷氨酸)、不带电极性侧链(例如,甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸、色氨酸)、非极性侧链(例如,丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸)、β-支链侧链(例如,苏氨酸、缬氨酸、异亮氨酸)和芳香族侧链(例如,酪氨酸、苯丙氨酸、色氨酸、组氨酸)的氨基酸。因此,本技术抗体的cdr区中的一个或多个氨基酸残基可以用同一侧链组的其他氨基酸残基替换,且改造后的抗体可以使用本文所述的功能检测测试其保留下来的功能(即上述的功能)。
[0101]
本技术的抗体可以用具备本技术siglec15抗体的一个或多个vh/v
l
序列的抗体为起始原料,制备成修饰的抗体。抗体可以通过修饰一个或两个可变区(即,vh和/或v
l
)内(例如,在一个或多个cdr区和/或一个或多个骨架区)的一个或多个残基来进行基因修饰。此外以及可选地,抗体可以通过修饰恒定区的残基来进行工程改造,以例如改变抗体的效应功能。
[0102]
在某些实施方式中,cdr植入可以用来工程改造抗体的可变区。抗体主要通过位于六个重链和轻链互补决定区(cdr)中的氨基酸残基与靶标抗原进行相互作用。出于这个原因,在个体抗体之间,cdr内的氨基酸序列比cdr外的序列更加多样。因为cdr序列负责大部分抗体-抗原相互作用,可以通过构建包含有将来自特定天然抗体的cdr序列植入到具有不同特性的不同抗体的骨架序列中的表达载体,来表达模拟特定天然抗体的特性的重组抗体(参见例如riechmann et al.,(1998)nature 332:323-327;jones et al.,(1986)nature 321:522-525;queen et al.,(1989)proc.natl.acad。又可参见u.s.a.86:10029-10033;美国专利5,225,539;5,530,101;5,585,089;5,693,762和6,180,370)。
[0103]
因此,本技术的另一实施方式涉及分离的单克隆抗体或其抗原结合部分,其包含重链可变区和/或轻链可变区,重链可变区包含具有本技术上述序列的cdr1、cdr2和cdr3序列,轻链可变区包含具有本技术上述序列的cdr1、cdr2和cdr3序列。尽管这些抗体包含本技术单克隆抗体的vh和v
l cdr序列,它们可以含有不同的骨架序列。
[0104]
这些骨架序列可以从包括种系抗体基因序列的公开dna数据库或公开参考文献中获得。例如,人重链和轻链可变区基因的种系dna序列可以在“vbase”人种系序列数据库获
得(可得自www.mrc-cpe.cam.ac.uk/vbase),也可以在kabat et al.,(1991),同上;tomlinson et al.,(1992)j.mol.biol.227:776-798;和cox et al.,(1994)eur.j.immunol.24:827-836获得;上述文件各自的内容通过引用的方式明确并入本文。作为另一个例子,用于人重链和轻链可变区基因的种系dna序列可以在genbank数据库中得到。例如,下列hco7 humab小鼠中的重链种系序列可得自所附的genbank登录号1-69(ng-0010109、nt
‑‑
024637&bc070333)、3-33(ng
‑‑
0010109&nt
‑‑
024637)和3-7(ng
‑‑
0010109&nt
‑‑
024637)。作为另一例子,以下来自hco12 humab小鼠的重链种系序列可得自genbank登录号1-69(ng-0010109、nt
‑‑
024637&bc070333)、5-51(ng
‑‑
0010109&nt
‑‑
024637)、4-34(ng
‑‑
0010109&nt
‑‑
024637)、3-30.3(caj556644)&3-23(aj406678)。
[0105]
通过使用本领域公知的称为空格blast的序列相似性搜索方法(altschul et ak.l(1997),同上),将抗体蛋白序列与汇编蛋白序列数据库进行比较。
[0106]
用于本技术抗体的优选骨架序列,是结构上与本技术抗体所用的骨架序列相似的那些。v
h cdr1、cdr2和cdr3序列可以植入到与得到该骨架序列的种系免疫球蛋白基因具有相同序列的骨架区中,或者cdr序列可以植入到与种系序列相比包含一个或多个突变的骨架区中。例如,发现了,在一些情况下,为保持或增强抗体的抗原结合能力,在骨架区中突变残基是有益的(参见例如美国专利5,530,101;5,585,089;5,693,762和6,180,370)。
[0107]
另一类的可变区修饰是将vh和/或v
l cdr1、cdr2和/或cdr3区内的氨基酸残基进行突变,从而改进目标抗体的一种或多种结合特性(例如,亲和力)。可以进行点突变或pcr介导的突变来引入突变,且其对于抗体结合或其他目标功能特性的影响可以在本领域所知的体外或体内检测中进行评价。优选地,引入(本领域所知的)保守修饰。突变可以是氨基酸替换、添加或缺失,但优选为替换。此外,通常对cdr区的改变不多于一个、两个、三个、四个或五个残基。
[0108]
此外,在另一实施方式中,本技术提供分离的siglec15单克隆抗体或其抗原结合部分,其包含重链可变区,其包含:(a)v
h cdr1区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列;(b)v
h cdr2区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列;(c)v
h cdr3区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列;(d)v
l cdr1区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列;(e)v
l
cdr2区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列;和(f)v
l cdr3区,其包含本技术的序列,或含有一个、两个、三个、四个或五个氨基酸替换、缺失或添加的氨基酸序列。
[0109]
本技术的基因改造抗体包括,例如为提高抗体特性,在vh和/或v
l
的骨架残基中做出基因修饰的抗体。通常而言,做出这些骨架修饰,以降低抗体的免疫原性。例如,一种方法是将一个或多个骨架残基“回复突变”成相应的种系序列。更加具体而言,经历体细胞突变的抗体可能包含不同于已得到抗体的种系序列的骨架残基。这些残基可以通过比较抗体骨架序列和得到抗体的种系序列而识别出来。
[0110]
另一类的骨架修饰涉及对骨架区的、或者甚至一个或多个cdr区的一个或多个残基进行突变,以去除t细胞表位,从而减少抗体的潜在免疫原性。该方法也称为“去免疫化”,
在美国专利公开20030153043中有更加详细的描述。
[0111]
此外,或者作为对骨架或cdr区内修饰的另一种选择,本技术的抗体可以基因修饰成包含fc区内的修饰,通常是为改变抗体的一个或多个功能特性,例如血清半衰期、补体结合、fc受体结合、和/或抗体依赖的细胞毒性。此外,可以对本技术的抗体进行化学修饰(例如,可以向抗体附加一个或多个化学基团),或者修饰成改变其糖基化,同样是为改变抗体的一个或多个功能特性。
[0112]
在一个实施方式中,c
h1
铰链区经修饰,改变,例如增加或减少,铰链区半胱氨酸残基的数量。该方法在美国专利5,677,425中作进一步描述。改变c
h1
铰链区中的半胱氨酸残基的数量,以例如促进轻链和重链的组装或者增加或降低抗体的稳定性。
[0113]
在另一个实施方式中,抗体的fc铰链区经突变,以降低抗体的生物半衰期。更加具体地,将一个或多个氨基酸突变引入fc铰链片段的c
h2-c
h3
结构域连接区,从而相较于与天然fc-铰链结构域的金黄色葡萄球菌蛋白a(spa)结合,该抗体具有削弱的spa结合。该方法在美国专利6,165,745中有更详细的描述。
[0114]
在另一实施方式中,对抗体的糖基化进行修饰。例如,可以制备去糖基化的抗体(即,抗体缺少糖基化)。可以改变糖基化,例如增加抗体对抗原的亲和性。这样的糖化修饰可以通过例如改变抗体序列中的一个或多个糖基化位点来达成。例如,可以进行一个或多个氨基酸替换,以消除一个或多个可变区骨架糖基化位点,从而消除该位置的糖基化。这样的去糖基化可以增加抗体对抗原的亲和性。参见,例如美国专利5,714,350和6,350,861。
[0115]
另外地或者可选地,可以制备糖基化类型改变的抗体,例如岩藻糖残基量减少的低岩藻糖基抗体或者具有增加的平分型glcnac结构的抗体。经证实,这些改变的糖基化形式增加抗体的adcc活性。这样的糖基化修饰可以通过例如在糖基化系统改变的宿主细胞中表达抗体而完成。糖基化系统改变的细胞在领域中已知,且可以用作表达本技术重组抗体的宿主细胞,以制备糖基化改变的抗体。例如,细胞系ms704、ms705和ms709缺乏岩藻糖基转移酶基因fut8(α(1,6)-岩藻糖基转移酶),从而在ms704、ms705和ms709细胞系中表达的抗体在其糖中缺失岩藻糖。使用两种替换载体靶向破坏cho/dg44细胞的fut8基因,制备ms704、ms705和ms709 fut8-/-细胞系(参见美国专利公开20040110704和yamane-ohnuki et al.,(2004)biotechnol bioeng 87:614-22)。作为另一个例子,ep 1,176,195记载了具有功能破坏的fut8基因(其编码岩藻糖基转移酶)的细胞系,从而通过降低或消除α-1,6键相关酶,在该细胞系中表达的抗体表现出低岩藻糖基化。ep 1,176,195也描述了一种细胞系,其具有较低的向结合抗体fc区的n-乙酰氨基葡萄糖中添加岩藻糖的活性,或不具有酶活性,例如大鼠骨髓瘤细胞系yb2/0(atcc crl 1662)。pct公开文本wo 03/035835描述了cho变体细胞系,lec13细胞,其向asn(297)-连接糖添加岩藻糖的能力降低,从而使得宿主细胞中表达的抗体的低岩藻糖基化(参见shields et al.,(2002)j.biol.chem.277:26733-26740)。具有改变的糖基化图谱的抗体也可以在鸡蛋中制备,如在pct公开文本wo 06/089231中所述的。或者,具有改变的糖基化图谱的抗体可以在植物细胞如浮萍中制备。在植物体系中制备抗体的方法在对应alston&bird llp律师记录号040989/314911的2006年8月11日提交的美国专利申请中有过记载。pct公开文本wo 99/54342记载了一种细胞系,其基因改造成表达修饰糖蛋白的糖基转移酶(例如,β(1,4)-n-乙酰葡糖胺转移酶iii(gntiii)),从而在基因改造细胞系中表达的抗体表现出增加的平分型glcnac结构,其引起
抗体的adcc活性增强(umana et al.,(1999)nat.biotech.17:176-180)。或者,可以使用岩藻糖苷酶来切除抗体的岩藻糖残基,例如α-l-岩藻糖苷酶从抗体移除岩藻糖残基(tarentino et al.,(1975)biochem.14:5516-23)。
[0116]
包含在本技术中的本文抗体的另一修饰是聚乙二醇化。抗体可以聚乙二醇化,例如增加抗体的生物(例如,血清)半衰期。为使抗体聚乙二醇化,抗体或其片段通常与聚乙二醇(peg),例如peg的活性酯或醛类衍生物,在使一个或多个peg基团附于抗体或抗体片段的条件下反应。优选地,聚乙二醇化通过与活性peg分子(或类似的有反应性的水溶性聚合物)的酰化反应或烷化反应进行。本文中所用的术语“聚乙二醇”意在包括任何形式的用于衍生其他蛋白的peg,例如单(c
1-c
10
)烷氧基-或芳氧基聚乙二醇或聚乙二醇马来酰亚胺。在某些实施方式中,需要聚乙二醇化的抗体是去糖基化的抗体。蛋白聚乙二醇化的方法在领域内已知,且可以应用于本技术的抗体。参见,例如epo 154 316和ep 0 401 384。
[0117]
本技术的抗体可以用它们的多种物理特性进行表征,以检测和/或区别其不同类别。
[0118]
例如,抗体可以在轻链或重链可变区包含一个或多个糖基化位点。这些糖基化位点可能引起抗体的免疫原性增高,或由于抗原结合的改变引起抗体的pk变化(marshall et al(1972)annu rev biochem 41:673-702;gala and morrison(2004)j immunol 172:5489-94;wallick et al(1988)j exp med 168:1099-109;spiro(2002)glycobiology 12:43r-56r;parekh et al(1985)nature 316:452-7;mimura et al.,(2000)mol immunol 37:697-706)。已知糖基化发生在含有n-x-s/t序列的基序中。在一些情况下,优选不包含可变区糖基化的siglec15抗体。这可以通过选择在可变区不包含糖基化基序的抗体或者通过使糖基化区域内的残基发生突变来实现。
[0119]
在优选实施方式中,抗体不包含天冬酰胺异构位点。天冬酰胺的脱酰胺可能发生在n-g或d-g序列处,引起异天冬氨酸残基的生成,其向多肽链引入链接并降低其稳定性(异天冬氨酸效果)。
[0120]
各抗体将具有独特的等电点(pi),其基本落在6-9.5的ph范围内。igg1抗体的pi通常落在7-9.5的ph范围内,而igg4抗体的pi基本落在6-8的ph范围内。推测pi在正常范围外的抗体可能在体内条件下具有一些展开结构和不稳定性。因此,优选pi值落在正常范围内的siglec15抗体。这可以通过选择pi在正常范围内的抗体或通过突变带电表面残基来实现。
[0121]
在另一方面,本技术提供编码本技术抗体重链和/或轻链可变区或cdr的核酸分子。核酸可以存在于完整细胞、细胞裂解液中或处于部分纯化或基本纯化的形式。当通过标准技术从其他细胞组分或者其他污染物例如其他细胞核酸或蛋白中纯化出来时,核酸是“分离的”或“呈处于基本纯化的状态”。本技术的核酸可以为例如dna或rna,且可以包含或可以不包含内含子序列。在优选实施方式中,核酸是cdna分子。
[0122]
本技术的核酸可以使用标准的分子生物学技术获得。对于由杂交瘤(例如,由携带人免疫球蛋白基因的转基因小鼠制备的杂交瘤,下文会进一步描述)表达的抗体,编码杂交瘤制备的抗体的轻链和重链的cdna可以通过标准pcr扩增或cdna克隆技术获得。对于(例如使用噬菌体展示技术)从免疫球蛋白基因库获得的抗体,可以从基因库中回收编码这类抗体的核酸。
technology.methods in enzymology 185,academic press,san diego,calif.(1990))中有过描述。优选的用于哺乳动物宿主细胞表达的调控序列包括在哺乳动物细胞中引导高水平蛋白表达的病毒元件,例如得自巨细胞病毒(cmv)、猿猴病毒40(sv40)、腺病毒如腺病毒主要晚期启动子(admlp)和多瘤病毒的启动子和/或增强子。或者,可以使用非病毒调控序列,例如泛素启动子或β-珠蛋白启动子。另外,调控元件由不同来源的序列构成,例如srα启动子系统,其包含来自sv40早期启动子的序列和人t细胞白血病i型病毒的长末端重复(takebe et al.,(1988)mol.cell.biol.8:466-472)。选择与使用的表达宿主细胞兼容的表达载体和表达控制序列。
[0131]
抗体轻链基因和抗体重链基因可以插入到同一或不同的表达载体中。在优选实施方式中,通过将可变区插入到已经编码所需同种型的重链恒定区和轻链恒定区的表达载体中来构建任意抗体同种型的全长抗体基因,从而vh部分与载体中的ch部分可操作地连接,v
l
部分与载体中的c
l
部分可操作地连接。此外或者另外地,重组表达载体可以编码促进抗体链从宿主细胞分泌的信号肽。抗体链基因可以克隆到载体中,进而信号肽在框内连接到抗体链基因的氨基端。信号肽可以是免疫球蛋白信号肽或异源信号肽(即,来自非免疫球蛋白的信号肽)。
[0132]
除抗体链基因和调控序列外,本技术的重组表达载体可以携带其他序列,例如,在宿主细胞中调控载体复制的序列(例如,复制起始点)和可选择标记物基因。可选择标记物基因促进已导入载体的宿主细胞的选择(参见例如,美国专利4,399,216;4,634,665和5,179,017)。例如,通常而言,可选择标记物基因赋予已导入载体的宿主细胞对药物如g418、潮霉素、或甲氨喋呤的抗性。优选的可选择标记物基因包括二氢叶酸还原酶(dhfr)基因(用于具有甲氨喋呤选择/扩增的dhfr宿主细胞)和neo基因(用于g418选择)。
[0133]
对于轻链和重链的表达,编码重链和轻链的表达载体通过标准技术转染到宿主细胞中。多种形式的术语“转染”意在包括多种将外源dna导入原核或真核宿主细胞的常用技术,例如,电穿孔、磷酸钙沉淀、deae-葡聚糖转染等。尽管在原核或真核宿主细胞中表达本技术抗体在理论上是可行的,在真核细胞中表达抗体是最优选的,最优选在哺乳动物宿主细胞,因为这些真核细胞,特别是哺乳动物细胞,比原核细胞更可能组装并分泌出适当折叠且有免疫活性的抗体。
[0134]
优选的用于表达本技术重组抗体的哺乳动物宿主细胞包括中国仓鼠卵巢(cho细胞)(包括与dhfr可选择标记物一起使用的dhfr-cho细胞,dhfr-cho细胞在urlaub and chasin,(1980)proc.natl.acad.sci.usa 77:4216-4220中有过描述,dhfr可选择标记物在例如r.j.kaufman and p.a.sharp(1982)j.mol.biol.159:601-621中有过描述)、nso骨髓瘤细胞、cos细胞和sp2细胞。特别在使用nso骨髓瘤细胞时,另一优选的表达系统是gs基因表达系统,记载于wo 87/04462、wo 89/01036和ep 338,841。当编码抗体基因的重组表达载体导入哺乳动物宿主细胞时,通过培养宿主细胞一段足以在宿主细胞中表达抗体的时间,或优选将抗体分泌在宿主细胞生长的培养液中,来制备抗体。使用标准的蛋白纯化方法从培养基中回收抗体。
[0135]
本技术的抗体可以与治疗剂结合,形成例如抗体-药物偶联物(adc)的免疫偶联物。合适的治疗剂包括细胞毒素、烷化剂、dna小沟结合剂、dna嵌入剂、dna交联剂、组蛋白去乙酰化酶抑制剂、核输出抑制剂、蛋白酶体抑制剂、拓扑异构酶i或ii抑制剂、热激蛋白抑制
剂、酪氨酸激酶抑制剂、抗生素和抗有丝分裂剂。在adc中,抗体和治疗剂优选地通过可切割的接头结合,例如肽类接头、二硫类接头或腙类接头。更优选地,接头是肽类接头,例如val-cit、ala-val、val-ala-val、lys-lys、ala-asn-val、val-leu-lys、ala-ala-asn、cit-cit、val-lys、lys、cit、ser或glu。adc可以按照美国专利7,087,600;6,989,452;和7,129,261;pct公开wo 02/096910;wo 07/038,658;wo 07/051,081;wo 07/059,404;wo 08/083,312;和wo 08/103,693;美国专利公开20060024317;20060004081;和20060247295中的描述进行制备。
[0136]
另一方面,本公开涉及双特异性分子,其包含一个或多个本技术抗体与至少一个其他功能性分子例如另一个肽或蛋白(例如,另一抗体或受体的配体)相连,以生成与至少两个不同的结合位点或靶分子结合的双特异性分子。因此,如本文所用的,“双特异性分子”包括有三种或更多种特异性的分子。
[0137]
双特异性分子可以以多种不同形式和尺寸出现。在尺寸谱的一端,双特异性分子保持传统抗体形式,除其具有两条结合臂且各臂具有不同特异性,而非具有两条相同特异性的结合臂外。在另一个极端的是,由两个经肽链连接的单链抗体片段(scfv)构成的双特异性分子,即所谓的bs(scfv)2构建体。中间尺寸的双特异性分子包括由肽类接头连接的两个不同f(ab)片段。这些和其他形式的双特异性分子可以通过基因改造、体细胞杂交或化学方法进行制备。参见,例如kufer et al,同上;cao and suresh,bioconjugate chemistry,9(6),635-644(1998);和van spriel et al.,immunology today,21(8),391-397(2000),以及其中引用的文献。
[0138]
本文还提供了优先侵染并杀灭癌细胞的溶瘤病毒。本技术的抗体可以与溶瘤病毒一起使用。或者,编码本技术抗体的溶瘤病毒可以引入到人体中。
[0139]
本文还提供包含siglec15 scfv的嵌合抗原受体(car),该siglec15 scfv包含本文所述的cdr和重/轻链可变区。
[0140]
siglec15 car可以包含(a)含siglec15 scfv的胞外抗原结合域;(b)跨膜结构域;和(c)胞内信号转导结构域。
[0141]
car可以在胞外抗原结合域的n-端含有指导新生受体进入内质网的信号肽、以及在胞外抗原结合域的n-端的使受体更容易结合的铰链肽。car优选地在胞内信号传导结构域包含主要胞内信号传导结构域和一个或多个共刺激信号传导结构域。主要使用的和最有效的主要胞内信号传导域是包含itam的cd3-ζ胞质结构域,其磷酸化会引起t细胞活化。共刺激信号传导结构域可以衍生自共刺激蛋白,例如cd28、cd137和ox40。
[0142]
car还可以添加增强t细胞扩增、持久性和抗肿瘤活性的因子,例如细胞因子、和共刺激配体。
[0143]
还提供基因修饰的免疫效应细胞,其包含本文提供的car。在一些实施方式中,免疫效应细胞是t细胞、nk细胞、外周血单核细胞(pbmc)、造血干细胞、多能干细胞、或胚胎干细胞。在一些实施方式中,免疫效应细胞是t细胞。
[0144]
在另一方面,本技术提供一种药物组合物,其可以包含与药学上可接受的载体配制在一起的本技术的一种或多种抗体或其抗原结合部分、双特异性分子、car-t细胞、溶瘤病毒、免疫偶联物、核酸分子、表达载体、或宿主细胞。当组合物中含有多于一种的抗体(或其抗原结合部分、双特异性分子、car-t细胞、溶瘤病毒、免疫偶联物、核酸分子、表达载体、
或宿主细胞)时,抗体或其抗原结合部分、双特异性分子、car-t细胞、溶瘤病毒、免疫偶联物、核酸分子、表达载体、或宿主细胞可以单独给药。组合物可以任选地包含一种或多种其他药学上的活性成分,例如另一抗体或药物,例如抗肿瘤药物。
[0145]
药物组合物可以包含任何数量的赋形剂。可以使用的赋形剂包括载体、表面活性剂、增稠或乳化剂、固体粘合剂、分散或混悬剂、增溶剂、染色剂、矫味剂、包衣、崩解剂、润滑剂、甜味剂、防腐剂、等渗剂及其组合。在gennaro,ed.,remington:the science and practice of pharmacy,20th ed.(lippincott williams&wilkins 2003)中,对合适赋形剂的选择和使用有所讲解,其公开内容通过引用的方式并入本文。
[0146]
优选地,药物组合物适合于静脉内、肌内、皮下、肠道外、脊柱或表皮给药(例如通过注射或输注)。基于给药途径的不同,有效成分可以包在材料中,以保护其免受酸和其他可能使其失活的自然条件的影响。本文所用的术语“肠道外给药”是指非肠内和非局部外用的给药给药方式,通常通过注射进行,包括但不限于静脉内、肌内、动脉内、鞘内、囊内、眶内、心内、皮内、腹膜内、经气管、皮下、表皮下、关节内、囊膜下、蛛网膜下腔、椎管内、硬膜外和胸骨内注射和输注。或者,本技术的抗体可以通过非肠道外的途径给药,例如外用、表皮或粘膜给药,例如鼻内、经口、阴道、直肠、舌下、或局部外用。
[0147]
药物组合物可以为无菌水溶液或分散液的形式。它们也可以配制在微乳剂、脂质体或其他适于高药物浓度的有序结构中。
[0148]
可以与载体材料一起制备成单剂量型的有效成分的量将随着治疗主体和特定给药方式而变,且通常是产生疗效的组合物的量。基本而言,以百分比计,该量是与药学上可接受载体组合在一起的约0.01%-约99%,优选为约0.1%-约70%,最优选为约1%-约30%的有效成分。
[0149]
调整给药方案以提供最佳的所需的应答(例如,治疗应答)。例如,可以施用快速灌注剂,可以随时间推移施用多个分剂量,或者剂量可以随治疗情况的危急程度成比例降低或提高。特别有利的是,为方便施用和剂量均匀,配置剂量单位型的肠道外组合物。本文所用的剂量单位型是指物理上不连续的单位,适于治疗主体的单次给药;各单位包含计算出来与所需药学载体一起产生所需疗效的预定量的有效成分。或者,抗体可以以缓释剂施用,这种情况下所需的给药频率降低。
[0150]
对于组合物的给药,剂量范围可以为约0.0001-100mg/kg。
[0151]“治疗有效剂量”的本技术siglec15抗体或其抗原结合部分、或双特异性分子、car-t细胞、溶瘤病毒、免疫偶联物,优选地引起疾病症状严重程度的降低、疾病无症状期频率和持久度的增加、或预防疾病折磨造成的损伤或残疾。例如,对于荷瘤受试者的治疗,与未接受治疗的受试者相比,优选“治疗有效剂量”对肿瘤生长的抑制至少为约20%、更优选为至少约40%,甚至更优选为至少约60%,更优选至少为约80%。治疗有效量的治疗抗体可以在受试者中减小肿瘤尺寸,或者减轻受试者的症状,受试者通常为人,或者可以是其他哺乳动物。
[0152]
药物组合物可以是控释剂型,包括植入体、经皮贴剂和微胶囊递送系统。可以使用生物可降解、生物相容的聚合物,例如乙烯-醋酸乙烯、聚酸酐、聚乙醇酸、胶原蛋白、聚原酸酯和聚乳酸。参见,例如,sustained and controlled release drug delivery systems.j.r.robinson,ed.,marcel dekker,inc.,new york,1978。
[0153]
药学组合物可以经医学设备来给药,例如(1)无针皮下注射设备(例如,美国专利5,399,163;5,383,851;5,312,335;5,064,413;4,941,880;4,790,824和4,596,556);(2)微量输液泵(美国专利4,487,603);(3)经皮给药设备(美国专利4,486,194);(4)推注设备(美国专利4,447,233和4,447,224);和(5)渗透设备(美国专利4,439,196和4,475,196),上述文献的公开内容通过引用的方式并入本文。
[0154]
在某些实施方式中,可以配制本技术的单克隆抗体或其抗原结合部位、双特异性分子、或表达这些的核酸分子或表达载体,以确保在体内的合适分布。例如,为确保本技术的治疗抗体可以穿越血脑屏障,它们可以配制在脂质体中,其还可以额外地包含靶向功能基团,以增强对特定细胞或器官的选择性输送。参见,例如美国专利4,522,811;5,374,548;5,416,016;和5,399,331;v.v.ranade(1989)j.clin.pharmacol.29:685;umezawa et al.,(1988)biochem.biophys.res.commun.153:1038;bloeman et al.,(1995)febs lett.357:140;m.owais et al.,(1995)antimicrob.agents chemother.39:180;briscoe et al.,(1995)am.j.physiol.1233:134;schreier et al.,(1994)j.biol.chem.269:9090;keinanen and laukkanen(1994)febs lett.346:123;和killion and fidler(1994)immunomethods 4:273。
[0155]
本技术的组合物有许多体外和体内用途,涉及例如癌症和骨质疏松的治疗。该抗体可以施用于人受试者,例如在体内抑制肿瘤生长或骨质流失。
[0156]
鉴于本技术的siglec15抗体或其抗原结合部分的逆转siglec15介导的t细胞抑制和抑制癌细胞的增殖和存活的能力,本技术提供用于抑制受试者体内肿瘤细胞生长的方法,包括向受试者施用本技术的组合物,从而使得受试者体内的肿瘤生长受到抑制。可以通过本技术组合物治疗的肿瘤的非限制性实例,包括,但不仅限于,非小细胞型肺癌、卵巢癌、黑色素瘤、结直肠癌、乳癌(包括三阴性乳癌)、头颈部鳞状细胞癌、子宫内膜癌和鳞状细胞癌。此外,可以使用本技术的抗体来抑制复发或难治黑色素瘤的生长。
[0157]
在另一个方面,本技术提供一种用于抑制骨流失或提高骨密度的方法,包括向受试者施用治疗有效剂量的本技术的抗体或其抗原结合部分。
[0158]
在另一个方面,本技术提供一种联合治疗方法,其中本技术的siglec15抗体或其抗原结合部分、或双特异性分子、car-t细胞、溶瘤病毒、免疫偶联物与一个或多个对抑制受试者中肿瘤生长有效的其他抗体共同施用。在一个实施方式中,本技术提供一种用于抑制受试者中肿瘤生长的方法,包括向受试者施用siglec15抗体(或其抗原结合部分、或car-t细胞、溶瘤病毒、免疫偶联物)和一个或多个其他抗体,例如vista抗体、lag-3抗体、pd-l1抗体和pd-1抗体和/或ctla-4抗体。在某些实施例中,受试者是人。
[0159]
siglec15信号通路阻断还可以与标准肿瘤治疗相结合。例如,siglec15信号阻断可以与ctla-4和/或lag-3和/或pd-1阻断以及化疗方案相结合。例如,化疗试剂可以与siglec15抗体相结合,其可以为细胞毒试剂。例如,将表阿霉素、奥沙利铂和5-fu施用给接受siglec15疗法的患者。
[0160]
可选地,siglec15抗体与一个或多个其他抗体(例如,ctla-4抗体和/或lag-3抗体和/或pd-1抗体)的联合使用还可以与免疫原剂例如癌细胞、纯化的肿瘤抗原(包括重组蛋白、肽类和碳水化合物)和转染有编码免疫刺激因子的基因的细胞相结合(he et al.,(2004)j.immunol.173:4919-28)。可以使用的肿瘤疫苗的非限制性实例包括黑色素瘤抗原
specificity,
″
nature,256:495-497(1975)中提到的方法与骨髓瘤细胞融合。之后将融合的“杂交细胞”在dmem/20%fcs/hat培养基中于96孔细胞板铺板。融合后的7-10天,在显微镜下观察存活的杂交瘤群落。在两周后,使用重组猴siglec15-his蛋白,对各孔的上清液进行间接elisa。选出分泌与猴siglec15-his蛋白结合的抗体的阳性杂交瘤并转移到24孔板中。之后进一步测试这些杂交瘤群落阻断人siglec15-lrrc4c结合的活性。通过有限稀释,对产生猴siglec15高特异结合性和siglec15-lrrc4c阻断活性的抗体的杂交瘤群落进行亚克隆,以确保细胞系的单克隆源性,之后纯化单克隆抗体。简单而言,使用5-10倍柱体积的pbs缓冲液冲洗蛋白a琼脂糖色谱柱(bestchrom(shanghai)biosciences,cat#aa0273)。杂交瘤单克隆的细胞上清液流经柱子,然后用pbs缓冲液冲洗柱子,直至蛋白吸光度达到基线。用洗脱缓冲液(0.1m甘氨酸-hcl,ph 2.7)洗脱柱子,并立即收集到含有中性缓冲液(1m tris-hcl,ph 9.0)的1.5ml管中。将含有免疫球蛋白的部分混合,并于4℃在pbs中透析过夜。随后,如下所示对纯化单克隆抗体的功能活性进行体外表征。
[0172]
实施例2.使用biacore表面等离子体共振对siglec15单克隆抗体的结合亲和力确定
[0173]
用biacore t200系统(ge healthcare,pittsburgh,pa,usa)对实施例1中产生的纯化siglec15单克隆抗体(mab)进行结合亲和力和结合动力学的表征。
[0174]
简单地说,使用biacore提供的标准胺偶联试剂盒(ge healthcare,pittsburgh,pa,usa),将山羊抗人igg(ge healthcare,cat#br100839,人抗体捕获试剂盒)经伯胺共价连接到cm5芯片(羧甲基右旋糖包被芯片,ge healthcare,cat#br-1005-30)。生物传感器表面未反应的基团用乙醇胺封闭。然后使纯化的本技术siglec15抗体以13.3nm的浓度,以及siglec15抗体对照基准物(ch5g12,也称为bm,参见美国专利公开us20190202912a1,重链和轻链的氨基酸序列分别为seq id nos:24和25)以13.3nm,分别流经芯片,流速为10μl/min。然后,将梯度稀释的重组人siglec15-his蛋白(内部制备,氨基酸序列为seq id no:20)或猴siglec15-his蛋白(内部制备,氨基酸序列为seq id no:21),起始浓度为80nm,在hbs-ep
缓冲液中2倍梯度稀释,以30μl/min的流速分别流到芯片上。抗原-抗体结合动力学观测2分钟,解离动力学观测10分钟。用biacore评估软件将结合与解离曲线拟合到1∶1 langmuir结合模型中。确定kd、ka、和kd值,并总结在以下表2中。
[0175]
表2.siglec15抗体的结合亲和力
[0176][0177]
本技术的所有siglec15抗体均与人siglec15和猴siglec15特异性结合,且与对照基准物相比,具有相当或更高的结合亲和力。抗体a2a1d2f1、a2a5c7e8、a1e10g7h9和a1d1b7h9显示出最高的结合亲和力。
[0178]
实施例3.siglec15抗体的siglec15结合活性
[0179]
通过捕获elisa,流式细胞术(facs)和间接elisa进一步测试了本公开的抗体与siglec15的结合活性。
[0180]
3.1捕获elisa
[0181]
简单来说,用100μl 2μg/ml f(ab
′
)2片段特异的亲和纯化山羊抗人igg(jackson immuno research,cat#109-005-097)的pbs溶液包被96孔板,37℃ 2小时。板用洗涤缓冲液(pbs 0.05%吐温-20,pbst)冲洗,再于4℃用200μl封闭缓冲液(含5%脱脂奶粉的pbst)封闭过夜。冲洗板4次,分别与100μl梯度稀释的本技术siglec15抗体、对照基准物和higg(静脉注射用人免疫球蛋白(ph 4),华兰生物工程有限公司)(起始浓度为66.7nm,用含2.5%w/v脱脂奶粉的pbst 5倍稀释)于37℃孵育40分钟,然后再次冲洗板四次。含有捕获抗体的板与100μl生物素标记的人siglec15-his蛋白(内部制备,seq id no:20,145ng/ml,于含2.5%脱脂奶粉的pbst中),于37℃孵育40分钟,冲洗4次,与链霉亲和素结合的hrp(1∶10000稀释于pbst缓冲液,jackson immuno research,cat#016-030-084,100μl/孔),于37℃孵育40分钟。在终洗后。板与100μl/孔elisa底物tmb(innoreagents,cat#tmb-s-002)孵育。在室温4分钟后用50μl/孔1m h2so4中止反应,在酶标仪中对各孔的吸光度进行读数,使用双波长模式,450nm用于tmb,630nm作为参照波长,用od(450-630)值和抗体浓度做图。使用graphpad prism软件对数据进行分析,得出ec
50
值。
[0182]
3.2基于细胞的结合facs
[0183]
采用内部制备的在细胞膜上稳定表达全长人siglec15(uniprot#q6zmc9,氨基酸残基met1-pro328)的人siglec15-2d3-1e1细胞,经流式细胞术(facs)检测siglec15抗体与
细胞表面siglec15的结合活性。按照lipofectamine 3000转染试剂(thermo fisher)的说明书,通过用ecori和xbai位点之间插入人siglec15编码序列的pcmv-t-p质粒转染hek-293细胞(atcc#crl-1573),制备出人siglec15-2d3-1e1细胞。从细胞培养瓶中收集人siglec15-2d3-1e1细胞,洗涤两次,在含2%v/v胎牛血清的磷酸盐缓冲液(pbs)(facs缓冲液)中重悬。96孔板中每孔2
×
105细胞,在冰上,与100μl梯度稀释于facs缓冲液的siglec15抗体或对照(起始浓度为66.67nm,5倍梯度稀释)孵育40分钟。细胞用facs缓冲液洗涤两次,并加入100μl r-藻红蛋白亲和纯化fcγ片段特异的山羊抗人igg(1∶1000稀释于facs缓冲液,jackson immunoresearch,cat#109-115-098)。4℃避光孵育40分钟后,细胞洗3次并用facs缓冲液重悬。使用becton dickinson facs canto ii-hts设备进行荧光测定。采用graphpad prism软件进行数据分析,得出ec
50
值。
[0184]
3.3 间接elisa
[0185]
检测siglec15抗体与猴或小鼠siglec15蛋白的交叉反应。简单来说,96孔微孔板用100μl 2μg/ml猴siglec15-his蛋白(内部制备,氨基酸序列为seq id no:21)或2μg/ml小鼠siglec15-his蛋白(内部制备,氨基酸序列为seq id no:22)的碳酸盐/碳酸氢盐缓冲液(ph 9.6)37℃包被2小时。elisa板用清洗缓冲液(pbs 0.05%吐温-20,pbst)洗涤一次,然后用200μl/孔封闭缓冲液(含5%w/v脱脂奶粉的pbst)4℃封闭过夜。板洗涤4次,并与100μl梯度稀释的本技术siglec15抗体或对照(起始浓度66.7nm,5倍梯度稀释于含2.5%脱脂奶粉的pbst中)37℃孵育40分钟。elisa板再冲洗4次,与过氧化物酶亲和纯化f(ab
′
)2片段特异的山羊抗人igg(1∶5000稀释于pbst缓冲液,jackson immunoresearch,cat#109-035-097,100μl/孔)37℃孵育40分钟。终洗后,板用100μl/孔tmb(innoreagent,cat#tmb-s-002)孵育。室温4分钟后,用50μl/孔1m h2so4中止反应,在酶标仪中对各孔的吸光度进行读数,使用双波长模式,450nm用于tmb,630nm作为参照波长,然后用od(450-630)值和抗体浓度做图。使用graphpad prism软件对数据进行分析,得出ec
50
值。
[0186]
三组检测的结果如图1a-1c到4a-4c所示。
[0187]
从图1a-1c可以看出,除a1c8c6h1以外的所有本技术抗体均与人siglec15特异性结合,与对照基准物相比,有更低的ec
50
和更高的bmax(最大结合)。
[0188]
如图2a-2c所示,与对照基准物相比,siglec15抗体a1d1b7h9、a1d11a7h10、a1e10g7h9、a2a1d2f1、a2g4c8g7和a2h5f1a1更高效地(更低的ec
50
)、以更高的bmax与细胞表面人siglec15结合。
[0189]
图3a-3c示出,大多数本技术的抗体与猴siglec15蛋白结合,且结合活性与对照基准物相似。然而,只有少数的本技术抗体显示出相当或更高的小鼠siglec15结合活性,如图4a-4c所示。例如,与对照基准物相比,抗体a1e10g7h9和a2a1d2f1示出更高的bmax。
[0190]
实施例4.siglec15抗体对siglec15-lrrc4c或siglec15-对照基准物结合的阻断活性
[0191]
4.1配体阻断elisa
[0192]
在竞争elisa实验中测定本技术siglec15抗体阻断siglec15-lrrc4c结合的活性。lrrc4c是siglec15的配体,可以由肿瘤细胞表达(wo2018057735)。简而言之,用100μl 2μg/ml于碳酸盐/碳酸氢盐缓冲液中的人siglec15-fc蛋白(内部制备,氨基酸序列为seq id no:19)包被96孔微孔板,37℃ 2小时。板用洗涤缓冲液(pbs 0.05%吐温-20,pbst缓冲液)
冲洗一次,用含5%脱脂奶粉的pbst4℃封闭过夜。板用洗涤缓冲液冲洗4次。
[0193]
将梯度稀释于含有2.5%w/v脱脂牛奶的pbst中的siglec15抗体或对照(起始浓度为66.67nm,5倍梯度稀释)加至siglec15-fc结合的板中,每孔100μl,与人siglec15-fc蛋白于37℃孵育40分钟。板用洗涤缓冲液再洗涤4次,添加100μl/孔290ng/ml生物素标记的人lrrc4c-fc蛋白(内部制备,seq id no:23),于37℃孵育40分钟。用清洗缓冲液洗板。然后,向板加入100μl/孔链霉亲和素结合的hrp(1∶5000稀释于pbst缓冲液,jackson immunoresearch,cat#016-030-084),于37℃孵育40分钟。用清洗缓冲液洗板。最后,加入tmb,用1m h2so4终止反应,在酶标仪中对各孔的吸光度进行读数,使用双波长模式,450nm用于tmb,630nm作为参照波长,用od(450-630)值和抗体浓度做图。使用graphpad prism软件对数据进行分析,得出ic
50
值。
[0194]
4.2对照基准物阻断elisa
[0195]
在竞争elisa检定中测定本技术siglec15抗体阻断对照基准物-人siglec15结合的能力。简而言之,将于pbs中的2μg/ml对照基准物包被于96孔微板,100μl/孔,并于37℃孵育2小时。之后用清洗缓冲液清洗板1次,用含5%w/v脱脂奶粉的pbst于4℃封闭过夜。次日,用生物素标记的人siglec15-fc蛋白(内部制备,seq id no:19,37ng/ml于含2.5%脱脂奶粉的pbst中)稀释本技术的siglec15抗体或对照,起始浓度80nm,4倍梯度稀释,于室温孵育40分钟。冲洗板4次后,将抗体/siglec15-fc混合物加入到对照基准物包被的板中,每孔100μl。37℃孵育40分钟后,板用洗涤缓冲液再洗涤4次。然后向板加入100μl每孔链霉亲和素结合的hrp,37℃孵育40分钟。用清洗缓冲液再次洗板。最后,加入tmb,用1m h2so4终止反应,在酶标仪中对各孔的吸光度进行读数,使用双波长模式,450nm用于tmb,630nm作为参照波长,用od(450-630)值和抗体浓度做图。使用graphpad prism软件对数据进行分析,得出ic
50
值。
[0196]
4.3基于细胞的配体阻断facs
[0197]
采用内部制备的lrrc4c-3f12-1b9细胞,经流式细胞术(facs)检测siglec15抗体阻断siglec15蛋白与细胞表面lrrc4c结合的能力。简而言之,按照lipofectamine 3000转染试剂(thermo fisher)的说明书,用ecori和xbai位点之间插入编码人lrrc4c(uniprot#q9hcj2,氨基酸残基为met1-lle640)的核苷酸序列的pcmv-t-p质粒构建体转染hek-293细胞(atcc#crl-1573)。选出称为lrrc4c-3f12-1b9的稳定细胞用于之后的基于细胞的配体阻断测试。
[0198]
简而言之,用人siglec15-小鼠fc蛋白(内部制备,seq id no:26,8μg/ml nm于facs缓冲液中)稀释本技术的siglec15抗体或对照,起始浓度66.67nm,5倍梯度稀释,室温孵育40分钟。然后,从细胞培养瓶中收集lrrc4c-3f12-1b9细胞,洗涤两次,在含2%v/v胎牛血清的pbs(facs缓冲液)中重悬。然后,96孔板中每孔1
×
105细胞,与100μl抗体/siglec15-小鼠fc混合物,于4℃孵育40分钟。板用facs缓冲液洗涤两次,然后加入100μl/孔r-藻红素亲和纯化f(ab
′
)2片段山羊抗小鼠igg(h l)(1∶1000稀释于facs缓冲液,jackson immunoresearch,cat#115-116-146),4℃避光孵育40分钟。细胞洗涤两次,并用facs缓冲液重悬。使用becton dickinson facs canto ii-hts设备进行荧光测定。采用graphpad prism软件进行数据分析,得出ic50值。
[0199]
三组检测结果如图5a-5c到7a-7c所示。
[0200]
从图5a-5c可以看出,本技术的所有siglec15抗体能够阻断人siglec15-人lrrc4c
细胞/ml。与此同时,梯度稀释的本技术siglec15抗体或对照(4倍稀释于补充10%fbs的rpmi-1640培养液,起始浓度120μg/ml)与人siglec15-fc蛋白(内部制备,seq id no:19,40μg/ml于补充10%fbs的rpmi-1640培养液中)以1∶1体积比混合,于室温孵育30分钟。然后,将50μl含pmbc细胞的培养液和50μl抗体/siglec15-fc混合物加入到cd3抗体结合的板中,于37℃在co2培养箱中孵育3天。然后,100μl facs缓冲液加入到每个孔中,细胞多次抽吸并转移到u型板中。将u型板离心,除去上清液。之后,将细胞与50μl/孔hfcr(1∶10稀释于facs缓冲液,biolegend inc,cat#422302)避光4℃孵育15分钟。于4℃将细胞用抗cd4(biolegend inc.,cat#357410)和抗cd8(biolegend inc.,cat#301066)荧光mab(1∶10稀释于facs缓冲液)染色30分钟。细胞用facs缓冲液(200μl/孔)洗涤两次,在facs缓冲液(200μl/孔)中重悬。使用becton dickinson facs canto ii-hts设备进行荧光测定。采用graphpad prism软件进行数据分析,得出ec
50
值。
[0213]
结果如8a和8b所示。
[0214]
可以看出,抗体a2a5c7e8和a1e10g7h9能够逆转siglec15介导的cd8
t细胞和cd4
t细胞抑制,具有高于对照基准物的ec
50
。尤其是,相较于对照基准物,抗体a2a5c7e8在高剂量时对逆转t细胞抑制更高效,引起更高的cd4
和cd8
t细胞增殖百分比。
[0215]
实施例7.siglec15抗体的测序
[0216]
对两个抗体a2a5c7e8和a1e10g7h9进行测序,得到完整的重链和轻链可变区序列和恒定区序列。重链和轻链可变区的序列id no如表1所示,通过与数据库的序列比对确定重链/轻链的同种型。
[0217]
实施例8.siglec15抗体a2a5c7e8的基因改造工程
[0218]
为避免或降低在例如cdr区中某些氨基酸残基的脱氨基和异构化等可能不利地影响抗体的生产、稳定性、安全性和/或有效性的翻译后修饰,对抗体a2a5c7e8(下文中又称a2a5c7e8-1)的重链cdr2区进一步修饰。得到共两个修饰后的变体,即a2a5c7e8-2和a2a5c7e8-3,其cdr和重/轻链可变区序列id号列于表1。
[0219]
各自包含编码与人igg1重链恒定区(seq id no:17)相连的a2a5c7e8-2或a2a5c7e8-3重链可变区的核苷酸的载体、以及各自包含编码与人κ轻链恒定区(seq id no:18)相连的轻链可变区的核苷酸的载体,以1.1∶1的轻∶重链构建体比,瞬时转染到50ml含1mg/ml pei 293f悬浮细胞培养物中。
[0220]
实施例9.修饰的a2a5c7e8变体的表征
[0221]
修饰的变体a2a5c7e8-2和a2a5c7e8-3如上所述进行纯化,并按照前述实施例中的方法步骤(有或无修改)以及以下描述的方法步骤,在biacore、捕获elisa、间接elisa、基于细胞的结合facs、竞争elisa以及细胞类功能检测中进行测定。
[0222]
对于测试修饰a2a5c7e8变体与小鼠siglec 15的结合亲和力的biacore测试,使用小鼠siglec15-his蛋白(内部制备,seq id no:22)。
[0223]
在t细胞抑制逆转实验中,将浓度48μg/ml的本技术a2a5c7e8变体或对照与人siglec15-fc蛋白(seq id no:19,20μg/ml于补充10%fbs的rpmi-1640培养基中)以1∶1体积比混合;将浓度120μg/ml的a2a5c7e8变体或对照与人siglec15-fc蛋白(40μg/ml于补充10%fbs的rpmi-1640培养基中)以1∶1体积比混合。
[0224]
biacore测试结果总结在表5-1和5-2中。其他实验的结果如图9-14和15a-15d所
示。
[0225]
表5-1.修饰的a2a5c7e8变体与人和猴siglec15的结合亲和力
[0226][0227]
表5-2.修饰的a2a5c7e8变体与小鼠siglec15的结合亲和力
[0228][0229]
如表5-1和5-2所示,修饰的变体a2a5c7e8-2和a2a5c7e8-3与人siglec15和猴siglec15特异性结合,与对照基准物相比,具有更高的结合亲和力。与对照基准物相比,抗体a2a5c7e8-3还显示出与小鼠siglec15的更高结合亲和力。抗体a2a5c7e8-2显示出与对照基准物相当的小鼠siglec15结合亲和力。
[0230]
修饰的变体a2a5c7e8-2和a2a5c7e8-3可以比对照基准物更高效地、或以更高的bmax与人siglec15结合(如图9和10所示),并比对照基准物更高效地抑制siglec15-lrrc4c结合(如图13所示)。此外,根据图11和12,与对照基准物相比,修饰变体a2a5c7e8-2和a2a5c7e8-3更高效地与猴siglec15以及小鼠siglec15结合。
[0231]
如图14所示,修饰变体a2a5c7e8-2和a2a5c7e8-3能够阻断对照基准物-siglec15的结合,表明修饰变体a2a5c7e8-2和a2a5c7e8-3结合在与对照基准物相似的表位。
[0232]
如图15a-15d所示,在基于细胞的功能检测中,修饰变体a2a5c7e8-2和a2a5c7e8-3逆转了siglec15介导的cd8
t细胞和cd4
t细胞抑制,引起与对照基准物相似或更高的细胞增殖百分比。尤其是,与对照基准物相比,在使用较低剂量人siglec15-fc蛋白时,抗体a2a5c7e8-2和a2a5c7e8-3在逆转cd4
和cd8
t细胞抑制方面更有效力。
[0233]
实施例10.使用杂交瘤技术生成小鼠源siglec15单克隆抗体
[0234]
免疫、杂交瘤融合和筛选
[0235]
小鼠的免疫、杂交瘤融合和筛选按照实施例1的操作(以及以下修改)进行。具体来说,使用正常的小鼠,免疫剂量包含用于初次免疫的50μg重组人siglec15-fc蛋白/小鼠/注
射,以及用于增强免疫的25μg人siglec15-fc蛋白/小鼠/注射。将抗原配制在pbs或生理盐水中,浓度为0.25-0.67mg/ml。以150-200μl的体积,共注射50或25μg抗原。对各个动物进行免疫,之后基于血清效价进行3-4次加强。使用内部制备的人siglec15-his蛋白对融合细胞培养上清液进行间接elisa。挑出分泌与人siglec15-his蛋白结合的抗体的阳性杂交瘤,转移至24孔板中。用这些杂交瘤进行基于细胞的结合facs、捕获elisa、间接elisa和配体阻断elisa。通过有限稀释,对产出有显示人siglec15-his、猴siglec15-his、小鼠siglec15-his高特异结合性以及人siglec15-lrrc4c阻断活性的抗体的杂交瘤克隆进行亚克隆,以确保细胞系的单克隆源性,之后纯化单克隆抗体。
[0236]
实施例11.使用biacore表面等离子体共振技术的小鼠源siglec15单克隆抗体的结合亲和力测定
[0237]
用biacore t200系统(ge healthcare,pittsburgh,pa,usa)对实施例10中生成的纯化小鼠源siglec15单克隆抗体进行结合亲和力和结合动力学表征。
[0238]
简而言之,使用biacore提供的标准胺偶联试剂盒(ge healthcare,pittsburgh,pa,usa),将山羊抗小鼠igg(ge healthcare,cat#br100838,小鼠抗体捕获试剂盒)经伯胺共价连接到cm5芯片(ge healthcare#br100530,羧甲基右旋糖包被的芯片)。生物传感器表面未反应的基团用乙醇胺封闭。纯化的本技术小鼠源siglec15抗体以13.3nm浓度分别流经芯片,流速为10μl/min。然后,梯度稀释的人siglec15-his蛋白(内部制备,氨基酸序列为seq id no:20)、猴siglec15-his蛋白(内部制备,氨基酸序列为seq id no:21)、或小鼠siglec15-his蛋白(内部制备,氨基酸序列为seq id no:22)(起始浓度为80nm,在hbs-ep
缓冲液中2倍梯度稀释(biacore提供)),以30μl/min的流速流到芯片上。抗原-抗体结合动力学观测2分钟,解离动力学观测10分钟。用biacore评估软件将结合与解离曲线拟合到1∶1 langmuir结合模型中。确定kd、ka、和kd值,并总结在以下表6-1和6-2中。
[0239]
表6-1.小鼠源siglec15抗体与人和猴siglec15的结合亲和力
[0240][0241]
表6-2.小鼠源siglec15抗体与小鼠siglec15的结合亲和力
[0242][0243]
本技术的所有小鼠源siglec15抗体均与人siglec15、猴siglec15、以及小鼠siglec15特异性结合,具有较高的结合亲和力。
[0244]
实施例12.小鼠源siglec15单克隆抗体的结合活性
[0245]
通过捕获elisa、间接elisa和流式细胞术(facs)检测小鼠源siglec15抗体与人siglec15、猴siglec15和小鼠siglec15的结合活性。
[0246]
捕获elisa按照实施例3的操作步骤进行,除用亲和纯化f(ab
′
)2片段特异的山羊抗小鼠igg(jackson immunoresearch,cat#115-005-072)替代亲和纯化f(ab
′
)2片段特异的山羊抗人igg外,100μl/孔。结果如图16所示。
[0247]
基于细胞的facs根据实施例3的操作步骤进行,除96孔板中每孔1.5x105细胞在冰上与100μl facs缓冲液中的梯度稀释siglec15抗体或对照(起始浓度为66.67nm,5倍梯度稀释)孵育40分钟,且向细胞中加入100μl r-藻红蛋白亲和纯化f(ab
′
)2片段山羊抗小鼠igg(h l)(1∶1000稀释于facs缓冲液,jackson immunoresearch,cat#115-116-146)外。结果如图17所示。
[0248]
间接elisa根据实施例3的操作步骤进行,除用过氧化物酶亲和纯化fcγ片段特异的山羊抗小鼠igg(jackson immunoresearch,cat#115-035-071)替代r-藻红蛋白亲和纯化fcγ片段特异的山羊抗人igg外,100μl/孔。结果如图18和19所示。
[0249]
从图16可以看出,本技术的小鼠源siglec15抗体均与人siglec15特异性结合,与对照基准物相比(ch5g12,又称bm,见美国专利公开us20190202912a1,重链和轻链氨基酸序列分别为seq id nos:24和25),具有更高的bmax(最大结合)和更低的ec
50
,表明它们更高效地与人siglec15蛋白结合。根据图17,在facs实验中,本技术的小鼠源siglec15抗体与细胞表面的人siglec15特异性结合,活性略低。
[0250]
图18示出,本技术的小鼠源siglec15抗体与猴siglec15蛋白特异性结合,与对照基准物相比,具有更高的结合活性,而图19显示出,本技术的小鼠源siglec15抗体与小鼠siglec15蛋白特异性结合,与对照基准物相比,结合活性略低。
[0251]
实施例13.小鼠源siglec15抗体对siglec15-lrrc4c或siglec15-对照基准物结合的阻断活性
[0252]
本技术是小鼠源siglec15抗体还在配体阻断elisa和对照基准物阻断elisa中测试,按照上述的操作。结果如图20和21所示。
[0253]
从图20可以看出,本技术的小鼠源siglec15抗体能够阻断人siglec15-人lrrc4c的结合,与对照基准物相比,阻断活性略高。
[0254]
图21示出本技术的小鼠源siglec15抗体能够阻断人siglec15-对照基准物的结合,说明小鼠源siglec15抗体可能结合在与对照基准物相似的表位。
[0255]
实施例14.小鼠源siglec15抗体逆转siglec15介导的t细胞抑制
[0256]
按照前述实施例中的方法步骤(加以修改)以及以下描述的方法步骤,进一步测试本技术小鼠源siglec15抗体逆转siglec15介导的t细胞抑制的生物活性。
[0257]
在t细胞抑制逆转实验中,浓度400μg/ml的本技术小鼠源siglec15抗体或对照与人siglec15-fc蛋白(seq id no:19,40μg/ml于补充10%fbs的rpmi-1640培养基中)以1∶1体积比混合。结果如图22a和22b所示。
[0258]
可以看出,本技术的小鼠源siglec15抗体能够以与对照基准物相近或更低的ec
50
逆转siglec15介导的cd8
t细胞和cd4
t细胞抑制,高剂量的本技术抗体的处理得到高得多的细胞增殖百分比。
[0259]
对本技术的小鼠抗体进行测序,重/轻链可变区的序列id编码总结在表1中。有意思的是,b2d7h7a3c1、b2g12h3e8和b2h2h1a7这三个抗体具有相同的重/轻链cdr序列。
[0260]
尽管本技术已经结合一个或多个实施方式进行了阐述,应当理解的是,本技术不限于那些实施方式,且说明书意在涵盖包括在所附权利要求的宗旨和范围内的所有可选方案、修改和等同物。所有本文中引用的参考文献均通过引用的方式全文并入。
[0261]
本技术中的序列总结如下。
[0262]
[0263]
[0264]
[0265]
[0266][0267]
***
[0268]
尽管已经在本技术的具体优选实施方式中进行了阐述,应当理解的是,由上述段落定义的本发明不限于在上述描述中列出的特定细节,可以在不脱离本发明宗旨和范围的情况下做出很多明显的变形。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。