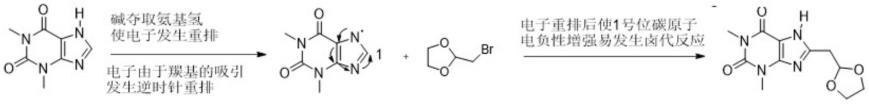
基于il-15和il-15r
α
sushi结构域的免疫细胞因子
1.分案说明
2.本技术是申请日为2012年6月22日、申请号为201810233665.5、发明名称为“基于il-15和il-15rαsushi结构域的免疫细胞因子”的发明专利申请的进一步分案申请。而申请号为201810233665.5的发明专利申请是申请日为2012年6月22日、国家申请号为201280037114.2、发明名称为“基于il-15和il-15rαsushi结构域的免疫细胞因子”的发明专利申请的分案申请。
3.本分案的国际专利申请要求2011年6月24日提交的欧洲专利申请11358005.4的优先权,其通过引用合并于此。
发明领域
4.本发明涉及新的免疫细胞因子(immunocytokine)及其作为药物,特别是用于治疗癌症的药物的用途。
背景技术:
5.免疫疗法在医学中指一系列基于调节免疫系统以实现预防和/或治疗目标的原理的治疗策略。
6.在过去几年中,免疫疗法已被用于治疗或预防一些病变,特别是癌症。由于用于产生单克隆抗体的细胞融合技术的发展,研究人员已经生产了大量单克隆抗体。此后,开发了产生单克隆抗体的其它技术,包括产生人单克隆抗体的b细胞杂交瘤技术和ebv杂交瘤技术。
7.单克隆抗体(mab)可以开发为靶向几乎任何抗原决定簇。特异性识别和结合特定细胞/分子的性质促进mab开发为用于各种疾病状态的诊断剂和治疗剂。dna重组技术已用于产生嵌合抗体或人源化抗体,以适应其给药于人类。目前,对于癌症、传染病、免疫性疾病等的治疗,一些单克隆抗体是市销且可获得的,比如比如
8.单克隆抗体是定向的分子,能够定位于特定区域(细胞、组织),例如肿瘤组织之内。在靶向称为肿瘤抗原的肿瘤部位中的特定分子的尝试中,该性质也导致产生结合到各种物质(有效载荷,payload)的mab的开发。此类物质(有效载荷)可以为毒素、药物、放射性核素、前药化合物等。这些连接中许多涉及反应性部分(有效载荷)与给定抗体制品的化学结合,其可能是缓慢复杂的并经历变化的过程(us 4,671,958)。
9.在这些新分子中,免疫细胞因子是特别令人感兴趣的。所述免疫细胞因子相当于包括抗体和细胞因子的融合蛋白。这些蛋白质同时保持抗原结合力和细胞因子活性。
10.细胞因子是一类在细胞通信中广泛使用的信号传导蛋白和糖蛋白,如激素和神经递质。虽然激素是由特定器官分泌到血液中,并且神经递质与神经活性有关,但是细胞因子为在其来源和用途方面更多样化类型的化合物。它们由多种造血细胞和非造血细胞类型产生,并可对邻近细胞或整个生物体都有作用,有时强烈地依赖于其它化学品的存在。细胞因子家族主要由质量为8至30kda之间的较小的水溶性蛋白和糖蛋白组成。细胞因子对于天然
免疫应答和适应性免疫应答的功能都重要。通常由已经遇到病原体的免疫细胞分泌,以作为活化和募集更多免疫细胞并增加系统对病原体反应的方式。然而,除了它们在免疫系统的发育和功能化中的作用,细胞因子还参与胚胎发生期间的一些发育过程。
11.在细胞因子中,白细胞介素15(il-15)是一种具有与il-2类似的结构的细胞因子,白细胞介素15是被病毒感染或被识别为非自身或虚弱的(debilitated)细胞间接刺激之后由单核吞噬细胞(和一些其它细胞)分泌的。该细胞因子诱导自然杀伤细胞的细胞增殖;自然杀伤细胞是主要作用为杀死病毒感染的细胞的固有先天免疫系统的细胞。由该基因编码的蛋白是调节t细胞和天然杀伤细胞活化和增殖的细胞因子。
12.因此,基于il-15的免疫细胞因子的构建对于肿瘤特异性抗体的肿瘤靶向益处与白细胞介素15的免疫调节作用的组合有特别的意义。已经获得特别地使用白细胞介素-2(il-2)的一些免疫细胞因子,并且在2期肿瘤学临床试验中证实了非常令人感兴趣和令人鼓舞的结果。这些融合蛋白的一些实施例描述在一些专利申请(us 5,645,835、ep 0,305,967、wo 86/01533、ep 0,439,095和wo 85/00974)中说明。
13.因此,已经在hek-293中产生了基于白细胞介素-15的免疫细胞因子,并在国际专利申请pct wo 2007/128563和kaspar等的(cancer research,vol.67(10),p:4940-4948,2007)中公开。
14.然而,本发明人证实此类基于白细胞介素15的免疫细胞因子具有非常有限的白细胞介素15活性,并且在cho细胞中特别地非常难于制备,产率低且具有许多蛋白污染物。
15.因此,仍然需要可以在免疫疗法中使用的基于白细胞介素15的免疫细胞因子。
16.发明简述
17.本发明涉及免疫细胞因子,所述免疫细胞因子包括:
18.a)缀合物,和
19.b)直接地或间接地共价连接至所述缀合物的抗体或所述抗体的片段,
20.其中,所述缀合物包括:
21.(i)包括白细胞介素15或所述白细胞介素15的衍生物的氨基酸序列的多肽,和
22.(ii)包括il-15rα或所述il-15rα的衍生物的sushi结构域的氨基酸序列的多肽。
23.在第二个方面,本发明涉及编码如上所述免疫细胞因子的核酸。
24.在第三个方面,本发明提供包括如上所述核酸的载体。
25.在第四个方面,本发明涉及一种用之前描述的多核苷酸或载体遗传工程化的宿主细胞。本发明还涉及一种制备宿主细胞的方法,所述遗传工程化的宿主细胞表达根据本发明的免疫细胞因子,所述方法包括下述步骤:(i)体外或间接体内引入如上所述核酸或载体到宿主细胞中,(ii)在体外或间接体内培养遗传工程化获得的重组宿主细胞,和(iii)可选地,选择表达和/或分泌所述免疫细胞因子的细胞。
26.在优选的实施方式中,所述遗传工程化的宿主细胞是动物细胞,优选cho细胞。
27.在第五个方面,本发明提供一种药物组合物,其包括最终与药学上可接受的媒介物结合的如上所述的免疫细胞因子、编码所述免疫细胞因子的核酸、或包括所述核酸的核酸载体。
28.在优选的实施方式中,所述组合物包括另外的治疗剂,优选抗癌剂。
29.在第六个方面,本发明涉及如之前描述的药物组合物,用于治疗受试者中的癌症。
30.在第七个方面,本发明涉及产品,包含:
31.(i)如上所述的免疫细胞因子,编码所述的免疫细胞因子的核酸序列,或包括这样的核酸序列的载体,和
32.(ii)治疗剂,优选抗癌剂,
33.作为用于治疗受试者中的癌症的同时、单独或连续使用的组合制剂。
34.在第八个方面,本发明涉及一种用于治疗受试者中的癌症的方法,包括给药如之前描述的药物组合物给所述受试者的步骤。
35.在最后的方面,本发明涉及用于治疗癌症的方法,包括向需要它的受试者同时、单独或顺序地给药治疗有效量的如下物质的步骤:
36.(i)如上所述的免疫细胞因子、编码所述的免疫细胞因子的核酸序列,或包括这样的核酸序列的载体,和
37.(ii)治疗剂,优选抗癌剂。
附图说明
38.图1显示与il15相比较,il15-抗-cd20免疫细胞因子的活性。
39.图2显示与il15相比较,il15-抗-o-乙酰化的cd20免疫细胞因子的活性。
40.图3分别显示il15-抗-cd20免疫细胞因子、抗-o-乙酰化的gd2免疫细胞因子与抗-her2 il-15免疫细胞因子的cd20、o-乙酰化的gd2和her-2结合活性。
41.图4显示与抗-cd20抗体(利妥昔单抗)相比较,il15抗-cd20免疫细胞因子的il-15rα结合活性。
42.图5分别显示il15抗-cd20免疫细胞因子、抗-o-乙酰化的gd2免疫细胞因子和抗her2 rli免疫细胞因子的cd20、o-乙酰化的gd2和her-2结合活性。
43.图6显示rli抗-cd20免疫细胞因子和il15抗-o-乙酰化的gd2免疫细胞因子的il15rα结合活性。
44.图7显示与il15相比较,rli抗-cd20免疫细胞因子的活性。
45.图8显示与il15相比较,rli抗-o-乙酰化的gd2免疫细胞因子的活性。
46.图9显示与抗-o乙酰化的gd2的抗体相比较,抗-o-乙酰化的gd2免疫细胞因子的抗转移活性。
47.图10显示抗-cd20免疫细胞因子在raji模型中的抗肿瘤活性。
48.图11显示与il15相比较,il-15抗-her2免疫细胞因子的活性。
49.图12显示与il15相比较,rli抗-her2免疫细胞因子的活性。
50.详细说明
51.本发明是基于本发明人发现,虽然包括白细胞介素15的免疫细胞因子的产生导致超过90%的白细胞介素15活性的丧失,但是基于rli的免疫细胞因子的产生导致新的il15免疫细胞因子,其对αβγ和βγ免疫细胞呈现出强的生物活性,很大程度上优于基于il-15的免疫细胞因子。
52.出人意料地,与单独的rli或scfv片段抗体相比较,具有全igg单克隆抗体的基于rli免疫细胞因子呈现出对βγ免疫细胞改善的生物学效力。在免疫抑制环境中,这个出人意料地获得的对βγ免疫细胞的活性在关于nk细胞和t淋巴细胞的活化/再活化方面可能很
重要。
53.更出人意料地,虽然白细胞介素15免疫细胞因子使在免疫球蛋白和白细胞介素15部分之间必须存在接头,以被活化;但是本发明的免疫细胞因子在其各自的免疫球蛋白和细胞因子部分之间有或者没有任何接头的情况下,呈现类似的白细胞介素15活性。在融合蛋白免疫原性、限制绞链区产生新的抗原表位和免疫原性方面及在具有有限裂解形式的产物收率方面,该接头区域的不必存在可提供有力的证明。
54.更出人意料地,本发明的免疫细胞因子是il-15超激动剂(super agonist),与单独的rli相比较,呈现出增强的活性(即10至100倍)。
55.而且,本发明人在cho细胞中获得了本发明的免疫细胞因子的产物的良好收率,收率超过90%。这是出人意料的,因为在cho细胞的相同细胞中白细胞介素15免疫细胞因子的产生非常难。
56.由于免疫细胞因子传统上具有有限的血清半衰期,并且由于免疫细胞因子相关的肿瘤定位率是产生强抗肿瘤效力的关键问题,基于rli的免疫细胞因子的特定生物活性允许以非常低的浓度活化免疫细胞,代表该领域的一个重要创新步骤,可以改善这类生物学化合物在癌症患者中的功效。
57.最后,本发明的免疫细胞因子的强的活性能够预示该免疫细胞因子的实际治疗用途,其应当以2.5~1mg/kg受试者或更小的剂量,并且甚至以0.1mg/kg或更小的剂量注射给药。实际上,白细胞介素15免疫细胞因子的低活性,比如在国际专利申请wo 2007/128563中公开的那种,不具有任何治疗用途(即,在小鼠肿瘤模型中,获得治疗效果需要超过20μg免疫细胞因子的剂量,每日注射四次,表明获得一些治疗效果需要超过5mg/kg免疫细胞因子的剂量)。
58.因此,本发明的一个方面涉及免疫细胞因子,包括∶
59.a)缀合物,和
60.b)直接地或间接地共价连接至所述缀合物的抗体或所述抗体的片段,
61.其中,所述缀合物包括:
62.(i)包括白细胞介素15或其衍生物的氨基酸序列的多肽,和
63.(ii)包括il-15rα或其衍生物的sushi结构域的氨基酸序列的多肽。
64.术语“免疫细胞因子”指包括直接地或间接地共价连接至细胞因子或其衍生物的抗体或它的片段的分子。所述抗体和所述细胞因子可以通过接头肽连接。
65.本发明的免疫细胞因子的缀合物
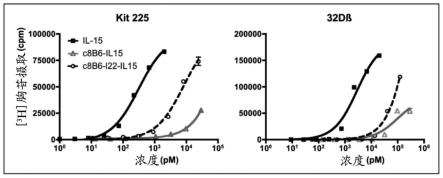
66.术语“白细胞介素15”为本领域它的一般含义,并指具有类似于il-2结构的细胞因子(grabstein等,science,vol.264(5161),p:965-968,1994)。该细胞因子也称为il-15、il15或mgc9721。该细胞因子和il-2共享许多生物学活性,发现它们结合常见血细胞生成素受体亚基。因此,它们可竞争相同的受体,负调节彼此的活性。已确定il-15调节t和自然杀伤细胞活化和增殖,并且cd8 记忆细胞的数量显示受该细胞因子和il2之间的平衡的控制。可以通过测定其对kit225细胞系的增殖诱导来测量il-15活性(hori等,blood,vol.70(4),p:1069-72,1987),如实施例中公开。
67.所述il-15或它的衍生物对于kit225细胞系的增殖诱导具有人白细胞介素15活性的至少10%,优选至少25%,更优选至少50%。
68.所述白细胞介素15为哺乳动物白细胞介素15,优选灵长类白细胞介素15,更优选人白细胞介素15。
69.本领域技术人员可简单地鉴定哺乳动物白细胞介素15。作为实例,可以引用来自野猪(登录号abf82250)、来自褐家鼠(登录号np_037261)、来自小家鼠(登录号np_032383)、来自黄牛(登录号np_776515)、来自家兔(登录号np_001075685)、来自绵羊(登录号np_001009734)、来自家猫(登录号np_001009207)、来自食蟹猴(登录号baa19149)、来自智人(登录号np_000576)、来自恒河猴(登录号np_001038196)、来自豚鼠(登录号np_001166300)或来自绿猴(chlorocebus sabaeus)(登录号aci289)的白细胞介素15。
70.如本文使用的术语“哺乳动物白细胞介素15”指共有序列seq id no.:1。
71.本领域技术人员可简单地鉴定灵长类白细胞介素15。作为实例,可以引用来自野猪(登录号abf82250)、来自家兔(登录号np_001075685)、来自食蟹猴(登录号baa19149)、来自智人(登录号np_000576)、来自恒河猴(登录号np_001038196)或来自绿猴(登录号aci289)的白细胞介素15。
72.如本文使用的术语“灵长类白细胞介素15”指共有序列seq id no.:2。
73.本领域技术人员可简单地鉴定人白细胞介素15,人白细胞介素15指氨基酸序列seq id no.:3。
74.如本文使用的术语“白细胞介素15衍生物”指具有与选自由seq id no.:1、seq id no.:2和seq id no.:3组成的组中的氨基酸序列至少92.5%(即,相当于约10个氨基酸取代)的同一性百分数的氨基酸序列,优选地至少96%(即相当于约5个氨基酸取代),更优选地至少98.5%(即相当于约2个氨基酸取代)或至少99%(即相当于约1个氨基酸取代)。本领域技术人员可基于其个人的知识和本专利申请的教导简单地鉴定这样的衍生物。作为这样的衍生物的实例,可以引用在国际专利申请pct wo 2009/135031中描述的那些。也应当理解天然氨基酸可以被化学修饰的氨基酸所替代。通常,这样的化学修饰的氨基酸提高了多肽半衰期。
75.如本文使用的两个氨基酸序列之间“同一性百分数”指由将进行比较的两个序列之间相同氨基酸的百分数,从所述序列的最佳比对获得,该百分数是纯粹统计学意义的,并且这两个序列之间的差异随机分布于所述氨基酸序列中。如本文使用的“最佳比对”或“最优的比对”指确定的同一性百分数(见下文)最高的比对。两个氨基酸序列之间的序列比较通常是通过比较之前已经根据最佳比对来比较的这些序列来实现的;该比较是在比较节段(segment)上实现的,以便鉴定和比较局部区域相似性。除了手动方式之外,可通过使用smith和waterman开发的总体同源性算法(ad.app.math.,vol.2,p:482,1981),可通过使用neddleman和wunsch开发的局部同源性算法(j.mol.biol.,vol.48,p:443,1970),pearson和lipman开发的相似性方法(proc.natl.acd.sci.usa,vol.85,p:2444,1988),通过使用这类算法的计算机软件(在wisconsin genetics software package,genetics computer group,575science dr.,madison,wi usa中的gap、bestfit、blast p、blast n、fasta、tfasta),通过使用muscle多重比对算法(edgar,robert c.,nucleic acids research,vol.32,p:1792,2004),实现用于进行比较的最佳序列比对。为了得到最优局部比对,可以优选地使用具有blosum 62矩阵的blast软件。通过比较最佳比对的这两个序列来确定的两个氨基酸序列之间的同一性百分数,所述氨基酸序列能够涵盖相对于参见序列的添加或缺
失,以便得到这两个序列之间的最佳比对。通过确定这两个序列之间相同位点的数目,用该数值除以比较位点的总数,得到的结果乘以100以得到这两个序列之间的同一性百分数,而计算同一性百分数。
76.优选地,白细胞介素15衍生物为il-15激动剂或超激动剂。在本领域技术人员可以简单地鉴定il-15-激动剂或超激动剂。作为il-15激动剂或超激动剂的实例,可以引用在国际专利申请wo 2005/085282或zhu等的(j.immunol.,vol.183(6),p:3598-607,2009)中公开的那些。
77.更优选地,所述il-15激动剂或超激动剂选自包括l45d、l45e、s51d、l52d、n72d、n72e、n72a、n72s n72y和n72p的组中/由l45d、l45e、s51d、l52d、n72d、n72e、n72a、n72s n72y和n72p组成的组中(参照人il-15的序列,seq id no.:3)。
78.如本文使用的术语“il-15rα的sushi结构域”具有本领域中其一般含义,指在il-15rα的信号肽之后第一个半胱氨酸残基(c1)开始,在所述信号肽之后第四个半胱氨酸残基(c4)终止的结构域。所述sushi结构域相应于il-15rα的细胞外区域的一部分对于它结合il-15是必需的(wei等,j.immunol,vol.167(1),p:277-282,2001)。
79.所述il-15rα的sushi结构域或所述il-15rα的sushi结构域的衍生物具有人il-15rα的sushi结构域对人白细胞介素-15的结合活性的至少10%,优选至少25%,更优选至少50%。可以通过wei等公开的方法(上述,2001)简单地确定所述结合活性。
80.所述il-15rα的sushi结构域为哺乳动物il-15rα的sushi结构域,优选灵长类il-15rα的sushi结构域,更优选人il-15rα的sushi结构域。
81.本领域技术人员可简单地鉴定哺乳动物il-15rα的sushi结构域。作为实例,可以引用来自褐家鼠(登录号xp_002728555)、来自小家鼠(登录号edl08026)、来自黄牛(登录号xp_002692113)、来自家兔(登录号xp_002723298)、来自食蟹猴(登录号aci42785)、来自豚尾猴(登录号aci42783)、来自智人(登录号cai41081)、来自恒河猴(登录号np_001166315)、苏门达腊猩猩(登录号xp_002820541)、白颈白眉猴(登录号aci42784)、普通狨(登录号xp_002750073)或豚鼠(登录号np_001166314)的il-15rα的sushi结构域。
82.如本文使用的术语“哺乳动物il-15rα的sushi结构域”指共有序列seq id no.:4。
83.优选地,包括哺乳动物il-15rα的sushi结构域的氨基酸序列的多肽指共有序列seq id no.:5。
84.本领域技术人员可简单地鉴定灵长类il-15rα的sushi结构域。作为实例,可以引用来自家兔、来自食蟹猴、来自豚尾猴、来自智人、来自恒河猴、苏门答腊猩猩、白颈白眉猴或普通狨的il-15rα的sushi结构域。
85.如本文使用,术语“灵长类il-15rα的sushi结构域”指共有序列seq id no.:6。
86.优选地,包括灵长类il-15rα的sushi结构域的氨基酸序列的多肽指共有序列seq id no.:7。
87.本领域技术人员可简单地鉴定人il-15rα的sushi结构域,并且所述人il-15rα的sushi结构域是指氨基酸序列seq id no.:8。
88.优选地,包括人il-15rα的sushi结构域的氨基酸序列的多肽指seq id no.:9。
89.如本文使用,术语“il-15rα的sushi结构域的衍生物”指具有与选自由seq id no.:4、seq id no.:5、seq id no.:6、seq id no.:7、seq id no.:8和seq id no.:9组成的
组中的氨基酸序列至少92%(即相当于约5个氨基酸取代)的同一性百分数的氨基酸序列,优选地至少96%(即相当于约2个氨基酸取代),更优选地至少98%(即相当于约1个氨基酸取代)。这样的衍生物包括l-15rα的sushi结构域的四个半胱氨酸残基,可以由本领域技术人员基于他/她的常识和本专利申请的教导简单地鉴定。还应当理解天然氨基酸可以被化学修饰的氨基酸所替代。通常,这样的化学修饰的氨基酸能够提高多肽半衰期。
90.根据优选的实施方式,缀合物包括(ii)含有il-15rα或其衍生物的sushi和hinge结构域的氨基酸序列的多肽。
91.所述il-15rα铰链结构域定义为开始于sushi结构域后的第一个氨基残基且终止于糖基化的第一个潜在位点前的最后一个氨基酸残基的氨基酸序列。在人il-15rα中,所述绞链区的氨基酸序列由十四个氨基酸组成,所述十四个氨基酸位于该il-15rα的sushi结构域之后的相对于所述sushi结构域的c末端位置,即所述il-15rα绞链区开始于所述(c4)半胱氨酸残基后的第一个氨基酸且终止于第十四个氨基酸(以标准的“从n-末端到c-末端”方向的计数)。
92.所述il-15rα的sushi和铰链结构域为哺乳动物il-15rα的sushi和铰链结构域,优选灵长类il-15rα的sushi和铰链结构域,更优选人il-15rα的sushi和铰链结构域。
93.本领域技术人员可简单地鉴定哺乳动物il-15rα的sushi和铰链结构域的氨基酸序列可以。如本文使用,术语“哺乳动物il-15rα的sushi和铰链结构域”指共有序列seq id no.:10。
94.本领域技术人员可简单地鉴定灵长类il-15rα的sushi和铰链结构域的氨基酸序列。如本文使用,术语“灵长类il-15rα的sushi和铰链结构域”指共有序列seq id no.:11。
95.本领域技术人员可简单地鉴定人il-15rα的sushi和铰链结构域的氨基酸序列。如本文使用,术语“人il-15rα的sushi和铰链结构域”指共有序列seq id no.:12。
96.如本文使用,术语“il-15rα的sushi和铰链结构域的衍生物”指具有与选自由seq id no.::10、seq id no.:11和seq id no.:12组成的组中的氨基酸序列至少93%(即相当于约5个氨基酸取代)的同一性百分数的氨基酸序列,优选地至少97%(即相当于约2个氨基酸取代),更优选地至少98%(即相当于约1个氨基酸取代)。这样的衍生物包括l-15rα的sushi结构域的四个半胱氨酸残基,并可以由本领域技术人员基于其常识和本专利申请的教导简单地鉴定。还应当理解天然氨基酸可以被化学修饰的氨基酸所替代。通常,这样的化学修饰的氨基酸能够提高多肽半衰期。
97.缀合物的多肽i)和多肽ii)都可以是非共价连接的,例如在美国专利8,124,084b2中公开的复合物中。可通过提供合适量的多肽i),提供合适量的多肽ii),在合适的ph和离子状态下混合这两种多肽足以允许复合物(即缀合物)形成的持续时间,并可选择地浓缩或纯化所述复合物,简单地获得所述缀合物或复合物。例如,根据标准方法使用肽合成仪;通过在细胞或细胞提取物中分别表达每种多肽,然后分离和纯化所述多肽,形成所述复合物(即缀合物)的多肽。可选择地,通过在相同细胞或细胞提取物中表达多肽i)和多肽ii),然后,例如使用色谱技术,例如具有针对淋巴因子部分、淋巴因子受体部分或复合物的抗体的亲和色谱,分离和纯化该复合物,可形成本发明的治疗性多肽复合物。
98.也可以使用双官能蛋白质偶联剂或在融合蛋白中共价连接缀合物的多肽i)和多肽ii)。
99.本领域技术人员熟知双官能蛋白质偶联剂,例如使用他们的方法,包括例如n-琥珀酰亚胺基(2-吡啶基二硫代)丙酸酯(spdp)、琥珀酰亚胺基(n-马来酰亚胺基甲基)环己烷-1-羧酸酯、亚氨基硫烷(it)、亚氨酸酯的双官能衍生物(盐酸二甲基亚氨酸酯)、活性酯类(例如辛二酸二琥珀酸亚胺酯)、醛类(例如戊二醛)、双-叠氮化合物(例如双(对-叠氮基苯甲酰)己二胺)、双-重氮衍生物(例如双-(对重氮苯甲酰)己二胺)、二异氰酸酯(例如甲苯2,6-二异氰酸酯)和双活性氟化合物(例如1,5-二氟-2,4-二硝基苯)。
100.术语“融合蛋白”指通过最初编码单独的蛋白质的两种或更多种基因连接产生的蛋白质。其也称为嵌合蛋白。该融合基因的翻译产生具有衍生自每种原始蛋白的功能特性的单一多肽。通过用于生物学研究或治疗的dna重组技术人工产生重组融合蛋白。重组融合蛋白是通过融合基因的遗传工程化产生的蛋白质。这通常包括从编码第一个蛋白质的cdna序列除去终止密码子,然后通过连接pcr或重叠延伸pcr在框架中附加第二个蛋白质的cdna序列。然后,由细胞将dna序列表达为单个蛋白质。该蛋白质可以工程化为包括两种原始蛋白质的全序列或任一个序列的仅一部分。
101.在优选的实施方式中,缀合物为融合蛋白。
102.白细胞介素15或其衍生物的氨基酸序列可以位于相对于il-15rα的sushi结构域或所述il-15rα的sushi结构域的衍生物的氨基酸序列的c-末端或n-末端位置。优选地,白细胞介素15或其衍生物的氨基酸序列位于相对于il-15rα的sushi结构域或所述il-15rα的sushi结构域的衍生物的氨基酸序列的c-末端位置。
103.白细胞介素15或其衍生物的氨基酸序列与il-15rα的sushi结构域或所述il-15rα的sushi结构域的衍生物的氨基酸序列可以被第一“接头”氨基酸序列隔开。所述第一“接头”氨基酸序列可以具有足够保证融合蛋白形成合适的二级和三级结构的长度。
104.所述第一接头氨基酸序列的长度可以变化,而不会显著地影响融合蛋白的生物活性。通常,第一接头氨基酸序列包括至少一个,但少于30个氨基酸,例如2~30个氨基酸,优选10~30个氨基酸,更优选15~30个氨基酸,进一步更优选15~25个氨基酸,最优选18~22个氨基酸的接头。
105.优选的接头氨基酸序列为允许缀合物采用合适构象的那些(即允许通过il-15rβ/γ信号通路的合适的信号传导活动的构象)。
106.最合适的第一接头氨基酸序列(1)将采用柔性延伸构象,(2)不会显示出发展为有序二级结构的倾向,所述二级结构可与融合蛋白的功能域相互作用,和(3)具有最小的疏水性或带电特性,所述疏水性或带电特性可促进与功能蛋白结构域的相互作用。
107.优选地,所述第一接头氨基酸序列包括选自由gly(g)、asn(n)、ser(s)、thr(t)、ala(a)、leu(l)和gln(q)组成的组中,最优选地选自由gly(g)、asn(n)和ser(s)组成的组中的接近中性的氨基酸。
108.美国专利no.:5,073,627和5,108,910中描述了接头序列的实例。
109.更特别地适用于本发明的示例性柔性接头包括seq id no:13(sggsggggsgggsggggslq)、seq id no.:14(sggsggggsgggsggggsgg)或seq id no.:15(sgggsggggsggggsgggslq)的序列编码的那些。
110.本发明的免疫细胞因子的抗体
111.术语“抗体”指免疫球蛋白分子,其相当于包括通过二硫键互连的两个相同的重
(h)链(全长时约50~70个kda)和两个相同的轻(l)链(全长时约25kda)的四个多肽链的四聚体。轻链分为κ和λ。重链分为γ、μ、α、δ或ε,并分别限定抗体的同种型为igg、igm、iga、igd和ige。每个重链由n-末端重链可变区(本文缩写为hcvr)和重链恒定区组成。对于igg、igd和iga,重链恒定区由三个结构域(ch1、ch2和ch3)组成;对于igm和ige,其由4个结构域(ch1、ch2、ch3和ch4)组成。每个轻链由n-末端轻链可变区(本文缩写为lcvr)和轻链恒定区组成。轻链恒定区由一个结构域cl组成。hcvr和lcvr区域可以进一步分为称为互补决定区(cdr)的高可变性区域、称为构架区(fr)的更保守的散布区域。每个hcvr和lcvr由按以下顺序从氨基末端至羧基末端排列的三个cdr和四个fr组成:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4。根据已知的惯例,对各个结构域的氨基酸进行归属。抗体结合特定抗原的功能能力取决于各个轻链/重链对的可变区,且主要由cdr确定。
112.如本文使用,术语“抗体”指单克隆抗体本身。单克隆抗体可以是人抗体、嵌合抗体和/或人源化抗体。
113.有利地,术语抗体指igg,例如igg1、igg2(igg2a或igg2b)、igg3和igg4。优选地,术语抗体指igg1或igg2,更优选igg2a。
[0114]“嵌合抗体”指由鼠科动物免疫球蛋白的可变区和人免疫球蛋白的恒定区组成的抗体。该改变仅包含用鼠科动物恒定区取代人抗体的恒定区,从而产生可具有对于药物用途可接受的足够低的免疫原性的人/鼠科动物嵌合体。已经报道了用于产生这样的嵌合抗体的大量方法,从而构成本领域技术人员的常识的一部分(参见,例如美国专利no.:5,225,539)。
[0115]“人源化抗体”指通过改变具有非人互补决定区(cdr)的抗体的序列,部分或完全由衍生自人抗体种系的氨基酸序列组成的抗体。通过本领域目前已知的技术进行该抗体可变区以及最终cdr的人源化。作为实例,英国专利申请gb 2188638a和美国专利no.:5,585,089公开产生重组抗体的方法,其中抗体被取代的唯一部分是互补决定区或“cdr”。cdr移植技术已用于产生由鼠科动物cdr和人可变区框架和恒定区组成的抗体(参见,例如riechmann等,nature,vol.332,p:323-327,1988)。这些抗体保留fc依赖性效应子功能所必需的、但是不太可能引起针对该抗体的免疫应答的人恒定区。作为实例,可变区的框架区被相应的人框架区取代,保持非人cdr的基本完整,或者甚至用衍生自人基因组的序列替换cdr(参见例如专利申请us2006/25885)。在为了响应于人免疫系统而改变其免疫系统的遗传修饰的小鼠中完全地产生人类抗体。如上述,使用所述抗体的免疫特异性片段,包括代表单链形态的片段,对于用在本发明方法中是足够的。
[0116]
人源化抗体还指包括人框架、至少一个来自非人抗体的cdr的抗体,其中存在的任何恒定区与人免疫球蛋白恒定区基本上相同,即至少约85或90%、优选至少95%相同。因此,可能除了cdr,人源化抗体的所有部分都与一个或多个天然人免疫球蛋白序列的相应部分基本上相同。例如,人源化的免疫球蛋白通常不涵盖嵌合体小鼠可变区/人恒定区抗体。作为实例,可以如下进行人源化免疫球蛋白的设计:当氨基酸属于下述类别时,将被使用的人免疫球蛋白(受体免疫球蛋白)的框架氨基酸被来自提供cdr的非人免疫球蛋白(供体免疫球蛋白)的框架氨基酸替换:(a)在所述位置的受体免疫球蛋白的人框架区中的氨基酸对于人免疫球蛋白是不常见的,而在所述位置的供体免疫球蛋白中的相应氨基酸对于人免疫球蛋白是常见的;(b)氨基酸的位置紧邻cdr的一个;或(c)在三维免疫球蛋白模型中,框架
氨基酸的任一个侧链原子在cdr氨基酸的任一个原子的约5~6埃(中心-至中心)之内(queen等,proc.natl.acad.sci.usa,vol.88,p:2869,1991)。当在该位置受体免疫球蛋白的人框架区中的氨基酸和供体免疫球蛋白中的相应氨基酸的每个对于人免疫球蛋白不常见时,在该位置这样的氨基酸被对于人免疫球蛋白典型的氨基酸替换。
[0117]
如本文使用的术语“抗体片段”指能够与其抗体对应物的相同的抗原反应的抗体片段。本领域技术人员可简单地鉴定这样的片段,并且本发明所涵盖,作为举例,包括f
ab
片段(例如通过木瓜蛋白酶消化)、f
ab
'片段(例如通过胃蛋白酶消化和部分还原)、f(
ab
')2片段(例如通过胃蛋白酶消化)、f
acb
(例如通过纤溶酶消化)、fd(例如,通过胃蛋白酶消化、部分还原和重聚合)、以及scfv(单链fv;例如通过分子生物学技术)片段。
[0118]
可以通过如本领域已知的和/或如本文描述的酶裂解、合成或重组技术产生这样的片段。也可以使用抗体基因以各种截短的形式产生抗体,所述抗体基因中已经在天然终止位点的上游引入了一个或多个终止密码子。例如,可设计编码f(
ab
')2重链部分的组合基因包含编码重链的ch1结构域和/或绞链区的dna序列。抗体的各个部分可以通过常规技术化学结合在一起,或者可以使用遗传工程化技术制备成连接蛋白。
[0119]
优选地,所述抗体片段为scfv片段。
[0120]
在优选的实施方式中,所述抗体或其片段针对与肿瘤新血管形成或肿瘤细胞外基质有关的抗原,或者针对肿瘤抗原。
[0121]
如本文使用,“与肿瘤新血管形成有关的抗原”指肿瘤中存在的新合成的血管表达的抗原。
[0122]
作为这样的抗原的实例,可以引用纤连蛋白的eda和edb结构域、endosalin/tem1、endoglin/105、psma或b7-h4。
[0123]
如本文使用,“与肿瘤细胞外基质有关的抗原”指肿瘤中存在的细胞外基质中表达的抗原。
[0124]
作为这样的抗原的实例,可以引用层粘连蛋白-332的g45片段(rousselle等,cancer research,vol.68(8),p:2885-94,2008)。
[0125]
如本文使用的“肿瘤抗原”指肿瘤细胞中产生的抗原物质。本领域技术人员已知许多肿瘤抗原,作为非限制性实例,可以引用cd-20、cea、egfr、gd2、epcam、muc1、psma、cd-19、gd3、gm1、caix、o-乙酰化的gd2或her2。
[0126]
cd-20为在前b细胞早期发育期间表达并保持直到浆细胞分化的非糖基化的磷蛋白。特别地,cd20分子可以调节细胞周期启动和分化所需的激活过程中的步骤,并通常在肿瘤性(“肿瘤”)b细胞中以非常高水平表达。根据定义,cd20同时存在于“正常”b细胞以及“恶性”b细胞中。因此,cd20表面抗原具有作为“靶向”b细胞淋巴瘤的候选物的潜力。
[0127]
关于针对cd-20的抗体,可以引用利妥昔单抗(美国专利no.:5,736,137);钇-[90]-标记的2b8鼠科动物抗体,称为“y2b8”或“替伊莫单抗”(美国专利no.:5,736,137);鼠科动物igg2a“bi”,也称为“托西莫单抗”,可选择地用
131
i标记以产生“131
i-bi”抗体(碘131托西莫单抗,)(美国专利no.:5,595,721);和人源化的2h7;奥法单抗(ofatumumab),针对cd20humax-cd20上的新抗原决定簇的全人源化的igg1(国际专利申请pct wo 2004/035607)。它们中,利妥昔单抗、替伊莫单抗和托西莫单抗取得销售许可,用于治疗特异性淋巴瘤,奥法单抗取得销售许可,用于治疗特异性白血病。
[0128]
cea(癌胚抗原)糖蛋白是一种参与细胞粘附的肿瘤标记物。
[0129]
关于针对cea的抗体,可以引用阿西莫单抗(immunomedics)。
[0130]
erbb受体在上皮细胞、间叶细胞和神经元来源的各种组织中表达。在正常情况下,erbb受体的活化受到其配体的时空表达的调控,所述配体是生长因子egf家族的成员。配体结合到erbb受体诱导受体同源二聚体和异源二聚体的形成和内在激酶结构域的活化,导致在胞质尾区之内特定酪氨酸激酶残基的磷酸化。这些磷酸化残基充当各种蛋白质的停泊位点,所述蛋白质的募集导致细胞内信号通路的激活。在erbb受体中,已知egfr和her2在调控细胞增殖和分化中起关键作用。一旦结合细胞外生长因子,它们具有强烈的与其它her受体组装成同源二聚体和/或异源二聚体的倾向,这导致各种形式的信号转导途径激活,导致细胞凋亡、存活或细胞增殖。
[0131]
关于针对egfr的抗体,可以引用人源化的单克隆抗体425,也称为马妥珠单抗(matuzumab,hmab 425,美国专利no.:5,558,864;ep 0531472),嵌合单克隆抗体225(cmab 225),也称为西妥昔单抗(美国专利no.:7,060,808)及人的完全抗-egfr抗体帕尼单抗(美国专利no.:6,235,883)。它们中,证实西妥昔单抗和帕木单抗在体内抑制人结肠直肠肿瘤,并且都取得销售许可。
[0132]
关于针对her2的抗体,可以引用小鼠抗体4d5的重组人源化形式(美国专利no.:5,677,171),称为humab4d5-8、rhumab her2、曲妥珠单抗或(美国专利no.:5,821,337)。该抗体在1998年取得销售许可,用于治疗患有转移性乳腺癌的患者,所述患者的肿瘤过表达erbb2蛋白质。
[0133]
gd2是一种在神经外胚层来源的肿瘤,包括成神经细胞瘤和黑素瘤上表达的双唾液酸神经节苷脂。
[0134]
关于针对gd2的抗体,可以引用已用于治疗成神经细胞瘤的鼠科动物igg3单克隆抗体3f8,或鼠科动物igg3单克隆抗体8b6,单克隆抗体3f8为gd2具体的o-乙酰化形式(国际专利申请pct wo 2008/043777)。
[0135]
优选地,所述抗体针对cd-20(例如,在美国专利no.:5,736,137中公开的利妥昔单抗)、o-乙酰化的gd2(例如,在国际专利申请pct wo2008/043777中公开的一种)或her2(例如在美国专利no.:5,821,337中公开的曲妥珠单抗或)。
[0136]
可以使用双官能蛋白质偶联剂或在融合蛋白中共价连接缀合物与抗体或所述抗体的片段。
[0137]
本领域技术人员熟知双官能蛋白质偶联剂方法,并且之前已经公开。作为实例,本领域技术人员可以使用在till等(proc.natl.acad,u.s.a.,vol.86(6),p:1987-91,1989)公开的方法。
[0138]
在优选的实施方式中,所述免疫细胞因子为融合蛋白。
[0139]
在另一个优选的实施方式中,免疫细胞因子为复合物,优选包括多肽i)和多肽ii)之间的缀合物的复合物,其中,所述多肽i)或多肽ii)融合到抗体或所述抗体的片段。
[0140]
所述多肽i)、多肽ii)或缀合物可以在相对于抗体或所述抗体的片段的氨基酸序列的c-末端或n-末端。
[0141]
优选地,缀合物为融合蛋白,所述缀合物的氨基酸序列在相对于抗体或所述抗体
片段的氨基酸序列的c末端,最优选在相对于抗体或所述抗体的片段的至少一个重链恒定区的氨基酸序列的c末端。
[0142]
缀合物的氨基酸序列和抗体或所述抗体的片段的氨基酸序列可以被或者不被第二“接头”氨基酸序列隔开。
[0143]
在特别的实施方式中,本发明的免疫细胞因子为融合蛋白,其中缀合物和抗体或抗体的片段没有被任何接头隔开。
[0144]
实际上,本发明人出人意料地确定本发明的免疫细胞因子在免疫球蛋白和细胞因子部分之间不需任何接头而具有活性。
[0145]
关于第一接头氨基酸序列,所述第二“接头”氨基酸序列可以具有足以确保融合蛋白形成合适的二级结构和三级结构的长度。
[0146]
第一接头氨基酸序列的长度可以变化,而不会显著地影响融合蛋白的生物活性。通常,第一接头氨基酸序列包括至少一个,但少于30个氨基酸,例如2~30个氨基酸,优选10~30个氨基酸,更优选15~30个氨基酸,最优选15~25个氨基酸的接头。
[0147]
关于第一接头氨基酸序列,最合适的第二接头氨基酸序列(1)将采用柔性延伸构象,(2)不会显示出发展为有序二级结构的倾向,所述二级结构可与融合蛋白的功能域相互作用,和(3)具有最小的疏水性或带电特性,所述疏水性或带电特性可促进与功能蛋白结构域的相互作用。
[0148]
优选地,所述第二接头氨基酸序列包括选自由gly(g)、asn(n)、ser(s)、thr(t)、ala(a)、leu(l)和gln(q)组成的组中,最优选由gly(g)、asn(n)和ser(s)组成的组中的接近中性的氨基酸。
[0149]
作为适于本发明的第二接头氨基酸序列的实例,可以引用序列seq id no.:16(sggggsggggsggggsggggsg)或seq id no.:17(aagggsgggsggggsggggsaa)。
[0150]
核苷酸、载体和重组宿主细胞
[0151]
在第二个方面,本发明涉及编码如上述的免疫细胞因子,优选相应于融合蛋白的免疫细胞因子的核酸。
[0152]
所述核酸对应于rna或dna,优选dna。
[0153]
根据优选的实施方式,编码本发明的免疫细胞因子的核酸有效地连接到基因表达序列,所述基因表达序列引导在原核细胞或真核细胞内、优选真核细胞之内的核酸表达。“基因表达序列”为任何调控核苷酸序列,例如启动子序列或启动子-增强子组合,其促进它有效地连接的免疫细胞因子核酸的有效转录和翻译。基因表达序列可以为例如哺乳动物或病毒启动子,例如组成型或诱导型启动子。
[0154]
组成型哺乳动物启动子包括但不限于用于下述基因的启动子:次黄嘌呤磷酸核糖转移酶(hptr)、腺苷脱氨酶、丙酮酸激酶、β-肌动蛋白启动子、肌肉肌酸激酶启动子、人延伸因子启动子及其它组成型启动子。真核细胞中的组成性起作用的示例性病毒启动子包括,例如来自猴病毒(例如sv40)、乳头状瘤病毒、腺病毒、人类免疫缺陷性病毒(hiv)、巨细胞病毒(cmv)、劳斯氏肉瘤病毒(rsv)、乙型肝炎病毒(hbv)、莫洛尼氏白血病毒及其它逆转录病毒的长末端重复(ltr)和单纯性疱疹病毒的胸苷激酶启动子。本领域普通技术人员已知其它组成型启动子。
[0155]
用作本发明的基因表达序列的启动子还包括诱导型启动子。诱导型启动子在诱导
剂的存在下表达。例如,在某些金属离子的存在下,启动子中的金属硫基蛋白(metallothione)被诱导,以促进转录和翻译。本领域普通技术人员已知其它诱导型启动子。
[0156]
通常,如需要,基因表达序列应当包括分别参与转录和翻译的启动的5'非转录和5'非翻译序列,例如tata盒、加帽序列、caat序列等。特别地,这样的5'非转录序列包括启动子区域,所述启动子区域包括用于有效地结合核酸的转录控制的启动子序列。如需要,基因表达序列可选择地包括增强子序列或上游激活序列。如本文使用,当编码本发明的免疫细胞因子的核酸序列和基因表达序列共价连接而使得本发明的免疫细胞因子编码序列的表达或转录和/或翻译处于基因表达序列的影响或控制下时,编码本发明的免疫细胞因子的核酸序列和基因表达序列被认为是“有效地连接的”。
[0157]
如果5'基因表达序列中启动子诱导导致本发明的免疫细胞因子转录并且如果两个dna序列之间的连接的性质不会(1)导致引入移码突变的引入、(2)干扰启动子区域引导本发明的免疫细胞因子转录的能力或(3)干扰相应的rna转录产物翻译至蛋白质中的能力,则认为有效地连接两个dna序列。因此,如果基因表达序列能够影响该核酸序列的转录而使产生的转录产物翻译至期望多肽中,则基因表达序列可有效地连接至编码本发明的免疫细胞因子的核酸序列。
[0158]
有利地,所述核酸序列包括内含子,因为前体mrna分子通常被证实提高重组分子的产品收率。可以提出任何内含子序列,作为实例,可以引用在zago等(biotechnol.appl.biochem.,vol.52(pt 3),p:191-8,2009)和campos-da-paz等(mol.biotechnol.,vol.39(2),p:155-8,2008)中公开的那些。
[0159]
编码本发明的免疫细胞因子的核酸可以在体内单独递送或与载体组合递送。
[0160]
在第三个方面,本发明涉及包括如上所述的核酸的载体。
[0161]
最广义地讲,“载体”为能够促进编码本发明的免疫细胞因子的核酸转移到细胞中的任何赋形剂。优选地,载体将核酸转运到细胞,相对于在不存在载体时导致的降解程度,减少了降解。通常,本发明中有用的载体包括但不限于质粒、粘粒、噬菌粒(phagmid)、附加体、人工染色体、病毒、已经通过插入或引入免疫细胞因子核酸序列操作的源自病毒或细菌源的其它赋形剂。
[0162]
质粒载体是优选类型的载体,在本领域已经进行了广泛的描述,并且本领域技术人员熟知。参见例如sanbrook等,“molecular cloning:a laboratory manual”第二版,cold spring harbor laboratory press,1989。质粒的非限制性实例包括pbr322、puc18、pucl9、prc/cmv、sv40和pbluescript、以及本领域普通技术人员熟知的其它质粒。另外,可以使用限制性内切酶和连接反应以除去和加入dna的特定片段来定制设计质粒。
[0163]
优选地,核酸载体可以包括在细菌和哺乳动物细胞中均有活性的可选标记。
[0164]
在第四个方面,本发明涉及用之前描述的核酸或载体遗传工程化的宿主细胞。
[0165]
如本文使用,术语“遗传工程化的宿主细胞”涉及已经用之前描述的核酸或载体转导、转化或转染的宿主细胞。
[0166]
作为合适的宿主细胞的代表性实例,可以引用细菌细胞,比如大肠杆菌、比如酵母菌的真菌细胞、例如sf9的昆虫细胞、例如cho或cos的动物细胞、植物细胞等。根据本文教导,认为合适的宿主的选择被在本领域技术人员的范围之内。
[0167]
优选地,遗传地工程化的宿主细胞为动物细胞,并且最优cho-s细胞(invitrogen,编号no.:11619-012)。
[0168]
中国仓鼠卵巢(cho)细胞经常用于制备生物制品,例如重组蛋白质、抗体、肽体(peptibody)和受体配体的生物制药工业中。经常使用cho细胞的原因之一是这些细胞具有用于生物制品生产的广泛的安全性记录。认为这是很好地表征的细胞系,因此,所需的安全试验在某些方面(例如,逆转录病毒安全性)可比其它细胞类型所需要较不严格。然而,白细胞介素15的产生非常难,尤其在该细胞中。
[0169]
出人意料地,本发明人证实本发明的免疫细胞因子在该细胞中很好地产生,得到的免疫细胞因子还具有非常好的纯度和活性。
[0170]
通过本领域技术人员熟知的方法进行之前描述的核酸或载体引入到宿主细胞中,所述方法例如磷酸钙转染、deae-葡聚糖介导的转染或电穿孔。
[0171]
本发明还涉及一种产生表达根据本发明的免疫细胞因子的遗传工程化的宿主细胞的方法,所述方法包括步骤:(i)在体外或间接体内引入如上所述核酸或载体到宿主细胞中,(ii)在体外或间接体内培养遗传工程化获得的重组宿主细胞,和(iii),可选择地,选择表达和/或分泌所述免疫细胞因子的细胞。这样的重组宿主细胞可用于产生本发明的免疫细胞因子。
[0172]
包括本发明的免疫细胞因子的药物组合物
[0173]
本发明的进一步目的涉及药物组合物,包括最终与药学上可接受的媒介物结合的如上述的免疫细胞因子、编码其的核酸、或包括所述核酸的载体。
[0174]
表述“药学上可接受的”指这样的分子实体和组合物:其是生理耐受的,并且当给药于人时通常不产生过敏反应或类似的不良反应,例如反胃、头晕等。优选地,如本文使用,表述“药学上可接受的”指联邦或州政府的管理机构批准的或在美国药典或其它普遍认可的用于动物、更特别是人类的药典中列出的。
[0175]
术语“媒介物”指与化合物一起给药的溶剂、助剂、辅料或赋形剂。这样的药物媒介物可以为无菌液体,比如水和油,包括石油、动物、植物或合成来源的油,比如花生油、大豆油、矿物油、芝麻油等。
[0176]
药物组合物包括“有效量”的本发明免疫细胞因子,其中有效量足以抑制癌细胞的生长,优选地足够诱导肿瘤生长的消退。用于给药的剂量可作为多种参数的函数而改变,特别是作为使用的给药模式、相关病理学模式或可选地期望的治疗持续时间的函数。自然地,药物组合物的形式、给药途径、剂量和方案自然取决于将被治疗的病症、疾病的严重性、受试者的年龄、体重和性别等。下面提供的有效剂量的范围不旨在限制本发明,并代表优选的剂量范围。然而,可为单独的受试者调整优选的剂量,如本领域技术人员理解和可确定,而无需过度实验。
[0177]
鉴于本发明的免疫细胞因子的显著功效,本领域技术人员可以计划使用非常少的剂量来治疗受试者。作为非限制性实例,本发明的免疫细胞因子可以以2.5mg/kg或1mg/kg受试者或更低的剂量,优选0.5mg/kg或更低或0.25mg/kg或更低的剂量,更优选0.1mg/kg或更低的剂量通过注射给药。
[0178]
作为实例,可以配制本发明的药物组合物用于局部、口服、鼻内、眼内、静脉内、肌内或皮下给药等。优选地,药物组合物包含对于旨在被注射的剂型药学上可接受的赋形剂。
特别地,这些可以为等渗的、无菌的盐溶液(磷酸二氢钠或磷酸氢二钠,氯化钠、氯化钾、氯化钙或氯化镁等,或这些盐的混合物),或干的,尤其是冷冻干燥的组合物,其根据情形加入无菌水或生理盐水时,允许构成可注射的溶液。在e.w.martin的“remington's pharmaceutical sciences”中描述了合适的药物媒介物。
[0179]
本发明的免疫细胞因子、编码其的核苷酸或核酸载体可以溶解在缓冲液或水中,或者包含在乳液、微乳液、水凝胶(例如plga-peg-plga三嵌段共聚物类水凝胶)、微球、纳米球、微粒、纳米颗粒(例如,聚(乳酸-共-乙醇酸)微粒(例如,聚乳酸(pla);聚(丙交酯-共-乙醇酸)(plga);聚谷氨酸酯微球、纳米球、微粒或纳米颗粒)、脂质体或其它盖仑剂型中。在所有情形中,制剂必须是无菌的和注射可接受程度的液体。其必须在制备和贮存条件下稳定,并且必须防止例如细菌和真菌的微生物的污染作用。
[0180]
呈游离碱或药理学可接受的盐的活性化合物的溶液可在与例如羟丙基纤维素的表面活性剂合适地混合的水中制备。
[0181]
分散体也可以在甘油、液体聚乙二醇、其混合物和油类中制备。在平常的储存和使用条件下,这些制剂包含防腐剂以防止微生物生长。
[0182]
根据本发明的免疫细胞因子可以以中性或盐形式配制到组合物中。药学可接收盐包括酸加成盐(与蛋白质的游离氨基形成),用例如盐酸或磷酸的无机酸或例如乙酸、草酸、酒石酸、扁桃酸等的有机酸形成所述酸加成盐。由游离羧基形成的盐也可以来源于无机碱,例如氢氧化钠、氢氧化钾、氢氧化铵、氢氧化钙、或氢氧化铁等,以及有机碱例如异丙胺、三甲胺、组氨酸、普鲁卡因等。
[0183]
所述媒介物也可以是溶剂或分散介质,包含例如水、乙醇、多元醇(例如甘油、丙二醇和液体聚乙二醇等)、它们合适的混合物及植物油。本发明的免疫细胞因子也可以被修饰,例如被聚乙二醇化,以提高其生物可分布性(biodisponibility)。当本发明的免疫细胞因子具有核酸形式时,所述媒介物也可以为载体,例如病毒(例如mva、raav、慢病毒等)。
[0184]
例如,可以通过使用包衣,例如卵磷脂,在分散体的情况下通过维持所需的粒径和通过使用表面活性剂,可以保持适当的流动性。通过多种抗菌剂和抗真菌剂,例如对羟基苯甲酸酯类、氯丁醇、苯酚、山梨酸、硫柳汞等,可以实现防止微生物的作用。在许多情况下,优选地包括等渗剂,例如糖或氯化钠。
[0185]
通过在组合物中使用延迟吸收的试剂,例如单硬脂酸铝、明胶、多元醇、增加半衰期的共价和非共价配方(covalent and non covalent formulations),可以导致可注射组合物的延长吸收。
[0186]
包括水解和变性的肽不稳定或降解有许多原因。疏水性相互作用可引起分子集合到一起(即聚集)。可以加入稳定剂以减少或防止这样的问题。
[0187]
稳定剂包括环状糊精及其衍生物(参见美国专利no.:5,730,969)。也可以加入合适的防腐剂,例如蔗糖、甘露醇、山梨醇、海藻糖、葡聚糖和甘油,以稳定最终制剂。选自离子和非离子表面活性剂、d-葡萄糖、d-半乳糖、d-木糖、d-半乳糖醛酸、海藻糖、葡聚糖、羟乙基淀粉、及其混合物的稳定剂可以加入到所述制剂中。加入碱金属盐或氯化镁可稳定肽。也可通过使肽接触选自由葡聚糖、硫酸软骨素、淀粉、糖原、糊精和海藻酸盐的糖类来稳定所述肽。可以加入的其它糖包括单糖、二糖、糖醇、及其混合物(例如,葡萄糖、甘露糖、半乳糖、果糖、蔗糖、麦芽糖、乳糖、甘露醇、木糖醇)。多元醇可稳定肽,且是可水混溶的或水溶性的。合
适的多元醇可以是多羟基醇、单糖和二糖,包括甘露醇、甘油、乙二醇、丙二醇、三甲基乙二醇、乙烯基吡咯烷酮、葡萄糖、乙烯基吡咯烷酮、葡萄糖、果糖、阿拉伯糖、甘露糖、麦芽糖、蔗糖及其聚合物。各种辅料也可稳定肽,包括血清白蛋白、氨基酸、肝素、脂肪酸和磷脂、表面活性剂、金属、多元醇、还原剂、金属螯合剂、聚乙烯吡咯烷酮、水解明胶和硫酸铵。
[0188]
细胞因子疗法的前景实际上源自这些新细胞因子的鉴定,但是甚至更重要地,所述领域极大地受益于临床前期数据的不断增长的量,所述数据有说服力地证实了协同和/或新的生物效应,其可以通过使用具有互补的免疫-刺激能力的细胞因子的一些组合的使用获得。与基于rli的免疫细胞因子组合的潜在治疗活性剂包括例如化疗剂、抗血管生成剂或免疫调节剂。
[0189]
在优选的实施方式中,本发明的组合物可以包括其它治疗活性剂,例如化疗剂、抗血管生成剂或免疫调节剂。
[0190]
对于化疗剂,已经证实其治疗作用可以部分地通过对于免疫应答的间接作用所调节,所述间接作用是通过诱导免疫原性细胞死亡、使免疫抑制环境保持平衡、使原发性大肿瘤减瘤去容积(debulk)并且然后促进免疫攻击,或通过诱导短暂的淋巴细胞减少然后诱导体内平衡的淋巴细胞增殖(homeostatic lymphoproliferation)来进行。本领域技术人员已知它们的许多种,作为可以与本发明的免疫细胞因子组合的化疗剂的实例,可以引用氟达拉滨、吉西他滨、卡培他滨、甲氨蝶呤、紫杉酚、泰索帝、巯嘌呤、硫鸟嘌呤、羟基脲、阿糖胞苷、环磷酰胺、异环磷酰胺、亚硝基脲、例如顺铂、卡铂和奥沙利铂的铂复合物、丝裂霉素、达卡巴嗪、丙卡巴肼(procarbizine)、依托泊苷、替尼泊苷、喜树碱类、博来霉素、多柔比星、伊达比星、柔红霉素、更生霉素、普卡霉素、米托蒽醌、左旋天冬酰胺酶、多柔比星、epimbicm、5-氟尿嘧啶、例如多西他赛和紫杉醇的紫杉烷、亚叶酸、左旋咪唑、依立替康、雌莫司汀、依托泊苷、氮芥、bcnu、例如卡莫司汀和洛莫司的汀亚硝基脲、例如长春碱、长春新碱和长春瑞滨的长春花生物碱、甲磺酸伊马替尼(imatimb mesylate)、六甲蜜胺、托泊替康、激酶抑制剂、磷酸酶抑制剂、atp酶抑制剂、酪氨酸磷酸化抑制剂(tyrphostin)、蛋白酶抑制剂、抑制剂除莠霉素a(herbimycm a)、染料木素、erbstatin和熏草菌素a。
[0191]
对于抗血管生成剂,已经证实其对免疫系统具脱靶效应,因此可促进肿瘤免疫应答。作为可以与本发明的免疫细胞因子组合的抗血管生成剂的实例,可以引用经由其酪氨酸激酶靶向血管内皮生长因子受体(vegfr)的药物,比如索拉非尼、舒尼替尼和帕唑帕尼,或雷帕霉素(mtor)的哺乳动物靶点,例如西罗莫司(temsirolimus)和依维莫司。
[0192]
对于可以与本发明的免疫细胞因子组合的免疫调节剂,可以引用细胞因子(il-2、il-7、il-15、il-12、il18、il-21、gm-csf、g-csf、ifnα
……
)、趋化因子/抗血管生成细胞因子(ip10、mig、sdf-1、rantes
……
)、tlr激动剂和免疫调节抗体(抗-ctla4、抗-pd1、抗-tgfb、抗-cd40的激动剂
……
)。
[0193]
治疗方法和用途
[0194]
在另外的方面,本发明涉及用于治疗受试者中的癌症的如之前描述的药物组合物,优选包括如之前描述的免疫细胞因子的药物组合物。
[0195]
如本文使用,术语“受试者”指哺乳动物,比如啮齿类动物、猫科动物、犬科动物或灵长类动物,最优选人类。
[0196]
在另一个方面,本发明涉及产品,包括:
[0197]
(i)如上所述的免疫细胞因子,编码所述的免疫细胞因子的核酸序列,或包括这样的核酸序列的载体,和
[0198]
(ii)治疗剂,优选抗癌剂,
[0199]
以作为用于治疗受试者中的癌症的同时、单独或连续使用的组合制剂。
[0200]
在另一个的方面,本发明涉及用于治疗受试者中癌症的方法,其包括给药如之前描述的药物组合物给所述受试者的步骤。
[0201]
在最后的方面,本发明涉及用于治疗癌症的方法,包括向需要它的受试者同时、单独或顺序地给药治疗有效量的下列物质的步骤:
[0202]
(i)如上所述的免疫细胞因子、编码所述的免疫细胞因子的核酸序列,或包括这样的核酸序列的载体,和
[0203]
(ii)治疗剂,优选抗癌剂。
[0204]
在本发明的上下文中,如本文使用的术语“治疗”或“处理”指逆转、减缓、抑制发展、或防止应用该术语的病症或疾病或这样的病症或疾病一个或多个症状、抑制所述病症或疾病或其症状的发展、或预防所述病症或疾病或其症状。如本文使用的术语“治疗癌症”指抑制癌细胞的生长。优选地,这样的治疗也引起肿瘤生长的肿瘤生长的消退,即可测量的肿瘤尺寸减小。最优选地,这样的治疗引起肿瘤完全消退。
[0205]
在下文中,参照氨基酸序列、核酸序列和实施例更详细地描述本发明。然而,本发明不旨在受实施例的细节的限制。相反,本发明适于包括在本文实施例中未明确提及、但本领域技术人员无需过度努力可发现的细节的任何实施方式。
实施例
[0206]
1)基于白细胞介素15的免疫细胞因子的构建
[0207]
抗-cd20(利妥昔单抗)和抗-o-乙酰化的gd2免疫细胞因子的构建
[0208]
编码抗-cd20嵌合igg轻链和抗-o-乙酰化的gd2嵌合igg轻链的表达质粒分别由watier博士(法国universit
é‑
rabelais de tours)和birkle博士(法国universit
é
de nantes,u892,inserm)友好地提供。设计每种抗体的嵌合igg重链序列在有或者没有22个氨基酸的接头(seq id no.:16)的情况下在3
’‑
末端融合至il15(seq id no.:3,其中在位点93的氨基酸为k)。通过geneart在pcdna3.1质粒中合成和克隆这些核苷酸序列。在专利申请ep 2,076,542 a1和cerato等(hybridoma,vol.16(4),p:307-16,1997)中公开了抗-o-乙酰化的gd2抗体(8b6)的轻链和重链的全序列。在专利us 5,736,137(anderson等,如称为“c2b8”的抗体)和reff等(blood,vol.83(2),p:435-45,1994)中公开了抗-cd20抗体(2b8)的轻链和重链的全序列。
[0209]
质粒dna制备和转染试剂
[0210]
由polyscience获得40kda的直链pei。通过加热在水中溶解pei,用naoh中和,并通过透过0.22μm过滤器过滤来灭菌,制备1mg/ml的储备溶液。将储备溶液等分并在﹣20℃储存。
[0211]
使用质粒纯化试剂盒,按照制造商的方案(macherey-nagel)纯化用于转染的质粒dna,并透过0.22μm过滤器过滤来灭菌。
[0212]
免疫细胞因子的制备和纯化
[0213]
1-在悬浮液中瞬时转染∶
[0214]
将常规维持的cho-s(invitrogen)细胞以1
×
106细胞/ml的密度接种在powercho2培养基(lonza)中,并且在具有5%co2的振荡培养箱(100rpm)中于37℃下培养过夜。为了转染,然后将细胞在cd-cho培养基(invitrogen)中稀释至2
×
106细胞/ml。以10%的培养体积,使用150mm的nacl制备转染复合物。将表达构建的dna(培养体积2.5mg/l,使用1:2的比的编码重链的质粒与编码轻链的质粒)与在nacl中稀释的pei(10mg/l的最终培养体积)混合,并在室温下培养10分钟,之后加到所述培养基中。在振荡培养箱(130rpm)中37℃培养细胞5小时,然后用powercho2培养基使培养体积翻倍。转染后5天,收集上清液。
[0215]
2-在粘附细胞上稳定的转染
[0216]
在添加左旋谷氨酰胺、10%的fcs和青霉素(100单位/ml)/链霉素(100μg/ml)的dmem中培养cho-k1细胞(atcc no.:ccl-61),并按照制造商建议,使用lipofectamine 2000试剂(invitrogen)用每种载体转染。对于抗-o-乙酰化的gd2 ick和抗-cd20 ick,通过分别用包含遗传霉素和潮霉素(0.5mg/ml)或blasticin和潮霉素(5μg/ml和100μg/ml)有限稀释而选择克隆。对于双官能蛋白质产生,通过elisa测定各个克隆的培养物上清液。对于ick的产生,使选择的克隆在25%dmem培养基和75%aim培养基(invitrogen)中扩增。然后,将细胞保持在100%的aim中,收集上清液,并每2天替换,共10天。
[0217]
3-上清液纯化:
[0218]
在4℃,以3000rpm离心收集的上清液20分钟,用naoh平衡ph至7.8,并透过0.22μm的过滤器过滤。根据制造商的说明,通过亲和色谱,使用a蛋白柱(ge)纯化条件培养基。用50kda amicon装置(millipore)浓缩纯化的蛋白质。在该步骤期间,pbs代替洗脱缓冲液。最后,通过elisa测定纯化的蛋白质,并在280nm测量吸光度。通过电泳评价纯度。
[0219]
4-通过elisa检测免疫球蛋白部分
[0220]
在4℃,用在pbs中稀释至1,5μg/ml的100μl的山羊抗-人抗体(up892370,interchim)涂布maxisorp平底微量滴定板(nunc)h,然后在37℃,用200μl的阻断缓冲液(在pbs中的1%bsa 0.1%吐温20)阻断培养板1小时。然后,用洗涤缓冲液(在pbs中0.1%吐温20)洗涤培养板3次,加入在在阻断缓冲液中稀释的样品,并在37℃下培养30分钟(100μl),在洗涤3次之后,加入1:10000稀释的过氧化物酶缀合的山羊抗-人igg1(109-036-003,jackson),并在37℃下培养30分钟。使用tmb底物(interchim)测定蛋白水平,并在450nm读取培养板。使用纯化的利妥昔单抗(roche)在培养板上产生标准曲线。
[0221]
5-通过elisa检测细胞因子部分
[0222]
在4℃,用在碳酸盐中稀释至2μg/ml的100μg的抗-il15 b-e29(diaclone)涂布maxisorp平底微量滴定板(nunc)16小时。然后,在37℃,用200μl的阻断缓冲液(在pbs中1%的bsa)阻断培养板1小时。然后,洗涤缓冲液(在pbs中0.05%吐温20)洗涤培养板3次。加入在tbs 0.05%bsa中稀释的样品,并在37℃培养1小时30分钟(100μl)。在洗涤3次之后,加入稀释至200ng/ml的生物素化的抗-il15抗体bam 247(r&dsystem),并在37℃下孵育1小时30分钟。洗涤培养板3次,并加入以1:1000稀释的过氧化物酶缀合的链霉亲和素。使用tmb底物(interchim)测定蛋白水平,并在450nm读取培养板。使用il-15(peprotech)在培养板上产生标准曲线。
[0223]
结果显示获得的免疫细胞因子制剂包括许多蛋白质污染物(即,等于或超过
25%)。为了减少这些蛋白质污染物,使两种抗-o-乙酰化的gd2/白细胞介素15免疫细胞因子进行另一轮a蛋白琼脂糖纯化。
[0224]
在这第二轮a蛋白琼脂糖纯化之后,ick c8b6-l22-il15和c8b6-il15的纯度分别为70和90%。
[0225]
免疫细胞因子的增殖活性
[0226]
测试获得的免疫细胞因子的白细胞介素-15增殖活性。通过[3h]胸苷结合测量kit225和32dβ细胞对ick的增殖反应。将细胞保持在培养基中3天,洗涤两次,并使kit 225和32dβ在不含细胞因子的培养基分别挨饿24小时或4小时。然后,按104细胞/孔以100μl接种在多孔培养板中,并在加入递增浓度的样品的培养基中培养48小时。人ril-15和rli用作校准。用0.5μci/孔的[3h]胸苷脉冲细胞16小时,收集到玻璃纤维过滤器上,并测量细胞关联的放射性。
[0227]
图1显示用递增浓度的ril-15(
■
)、c8b6-il15(δ)和c8b6-l22-il15(
○
)培养的kit 225和32dβ细胞结合的[3h]胸苷。
[0228]
图2显示用递增浓度的ril-15和c2b8-l22-il15(
○
)培养的kit 225和32dβ细胞结合的[3h]胸苷。
[0229]
结果显示在免疫细胞因子的情况中,il-15的生物活性显著降低,表明il-15与单克隆抗体的缀合诱发活性丧失。而且,当两个部分之间不存在接头的情况下,该丧失更重要。应当注意该活性丧失在βγ的情况下更严重。
[0230]
免疫细胞因子的结合活性
[0231]
通过流式细胞仪,分别评价抗-cd20和抗-o-乙酰化的gd2的ick对肿瘤细胞raji和imr32的特异性结合。在kit225上测定ick结合效应细胞上的il-15受体的能力。用pe-缀合的山羊抗-人igg mab(pn im0550,beckman coulter)或偶联至pe-链霉亲和素(sigma-aldrich)的生物素化的小鼠抗-il15抗体(bam247,r&d system)显示涂布在靶细胞上的ick。在4℃,用每个ick培养靶细胞(1
×
105)1小时,洗涤,然后在4℃用pe-缀合物培养1小时。最后,在facscalibur(becton dickinson)上最终分析洗涤的细胞。
[0232]
图3显示在表达cd20的raji细胞上和在表达o-乙酰化的gd2的imr32细胞上ick抗-cd20(c2b8-l22-il15)和抗-o-乙酰化的gd2(c8b6-il15和c8b6-l22-il15)的流式细胞计数评价。首先,用ick培养细胞,然后对于抗-cd20用pe-缀合的山羊抗-人igg mab培养,或者对于抗-cd20和抗-gd2分别用生物素化的抗-il15 pe-缀合的链霉亲和素培养。最后,在facscalibur上分析样品。比较raji细胞上的ick与抗-cd20 mab利妥昔单抗(mabthera,roche)。
[0233]
图4显示在表达il15r的kit 225细胞上ick抗-cd20(c2b8-l22-il15)和抗-o-乙酰化的gd2(c8b6-il15和c8b6-l22-il15)的流式细胞计数评价。首先,用ick培养细胞,然后用pe-缀合的山羊抗-人igg mab培养。最后,在facscalibur上分析样品。比较ick与抗-cd20 mab利妥昔单抗(mabthera,roche)。
[0234]
结果显示不同的免疫细胞因子结合il-15受体以及它们各自的肿瘤抗原靶。
[0235]
因此,这些免疫细胞因子中白细胞介素15活性丧失不是白细胞介素15结合其特异性抗体的丧失的结果。然而,似乎是已存在的结合不允许诱导正常细胞增殖。
[0236]
基于rli-的免疫细胞因子的构建
laboratories批准的,且根据inserm institute and the french department of agriculture的条例和标准维护。
[0254]
通过尾静脉注射在200μl的dmem(ph 7.4)中的1
×
105nxs2 nb肿瘤细胞诱发试验性肝转移瘤。在接种肿瘤细胞之后1天开始处理,并包括在第1、4、7和11天4次i.p.注射80pmol的c8b6-rli2或c8b6。在移植之后25天处死小鼠,通过湿肝重量评价肝肿瘤负荷。
[0255]
图9显示c8b6-rli2对nxs2肝转移的功效。在第1、4、7和11天i.p.给药c8b6(12μg)或c8b6-rli(16μg)。左:图表代表各组的平均值(n=5);条,sem。右:肝的代表性图像,箭头指示一些转移。
[0256]
结果显示接受ick的小鼠保持没有肝转移。因此,与c8b6相反,ick可以根除nxs2肝转移的发展,表明rli结合单克隆抗体显著地增加其抗肿瘤效力。
[0257]
在raji模型中,抗-cd20-rli2的抗肿瘤效力
[0258]
在添加10%胎牛血清、2mm l-谷氨酰胺的rpmi1640培养基中培养人raji b细胞。
[0259]
scid cb-17小鼠,8周龄,购自charles river breeding laboratories。将小鼠保持在使用微隔离器装置的高压处理笼的单独设备中的无特定病原体条件下,供应辐射的固体食物和无菌水。
[0260]
对于接种,收集处于对数期的raji细胞,洗涤并以2.5
×
106细胞/0.1ml再悬浮在磷酸盐缓冲液(pbs)中,之后静脉注射到小鼠中,接着在植入之后用免疫细胞因子ip处理3次/周(开始于第5天),处理3周。小鼠接受等摩尔量的处理,除了“免疫细胞因子”和“利妥昔单抗 rli”组接受一半剂量。每日监测小鼠后肢瘫痪的存在,在此情形下将其处死,并对评价为死亡。
[0261]
图10显示iv注射raji细胞(n=5)并且对j5-j7-j9、j12-j14-j16、j19-j21-j23用pbs(
■
)、rli(
▲
;2μg)、利妥昔单抗(
◆
;12μg)、利妥昔单抗 rli(
●
;6μg 1μg)、抗cd20-rli(
▼
;8μg)处理的cb17 scid小鼠的kaplan-meier存活率分析。
[0262]
结果显示在用rli或利妥昔单抗处理的raji小鼠中获得的存活率百分数相似,相对于pbs对照,分别将负载肿瘤小鼠的50%存活率从20天延长至27天和28天。
[0263]
结果显示对于“利妥昔单抗 rli”组,负载肿瘤小鼠的存活率百分数进一步增长一半,该增长显著不同于在rli组中得到的增长(p《0.01或更小)。
[0264]
最终并且出人意料地,结果显示在试验结束时(50天),用抗-cd20免疫细胞因子处理全部中止了肿瘤发展,而没有小鼠死亡。
[0265]
3)其它免疫细胞因子的构建
[0266]
抗-her2neu(全igg和scfv)rli和il15免疫细胞因子的构建
[0267]
编码抗-her2鼠科动物4d5 igg轻链、抗-her2鼠科动物igg 4d5重链和抗-her2scfv的序列均由donda博士(瑞士biochemistry institute lausanne)友好地提供。如前所述,基于抗-her2neu的轻链(seq id no.:18)和重链(seq id no.:19)构建抗-her2neu il15-免疫细胞因子和rli-免疫细胞因子。对于这些构造,设计在具有编码嵌合igg重链序列的框架中的β2微球蛋白前导序列的序列在有或者没有22个氨基酸的接头(seq id no.:16)的情况下在3'-末端融合至il15(分别为id no.:20和21)和rli(分别为seq id no.:22和23)。
[0268]
进一步设计和产生相应于编码在有或者没有22个氨基酸的接头(seq id no.:16)
15和曲妥珠单抗或抗-her2 scfv片段融合的白细胞介素-15增殖活性。
[0286]
图11显示用递增浓度的曲妥珠单抗-l22-il-15(
○
)、曲妥珠单抗-il-15(
△
)和ril-15(
■
)培养的kit 225和32dβ细胞结合的[3h]胸苷。
[0287]
图12显示用递增浓度的曲妥珠单抗-rli(
△
)、rli(
■
)和ril-15(
◆
)培养的kit 225和32dβ细胞结合的[3h]胸苷。
[0288]
结果显示在免疫细胞因子的情况下,il-15对αβγ细胞的生物活性显著降低,表明il-15与单克隆抗体的缀合诱发活性丧失。而且,当两个部分之间不存在接头下,该丧失更重要。在βγ细胞上,在有或者没有接头的情况下,缀合完全消除了il-15的生物活性。
[0289]
在除了il15免疫细胞因子的rli-衍生的免疫细胞因子的情况下,结果显示il-15的生物活性得到保持,表明rli与单克隆抗体的缀合出人意料地允许该il-15活性的保持。而且,该令人感兴趣的作用并不需要在rli和单克隆抗体之间的任何第二接头。
[0290]
更出人意料地,结果显示在βγ情况下,与自由il-15相比,rli-衍生的曲妥珠单抗免疫细胞因子呈现显著的生物活性增加(增加约10至100倍)。
[0291]
结果进一步表明,在除了il15免疫细胞因子的rli-衍生的scfv免疫细胞因子的情况下,il-15的生物活性也得到保持,表明rli与scfv片段的结合出人意料地允许该il-15活性的保持(未示出数据)。此外,该令人感兴趣的作用并不需要在rli和scfv片段之间的任何第二接头(未示出数据)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。