1.本公开涉及将在菌体内产生的蛋白分泌到菌体外的蛋白的分泌生产率提高了的修饰蓝细菌、修饰蓝细菌的制造方法以及蛋白的制造方法。
背景技术:
2.在从化学工业到农渔畜牧业的广泛的产业领域中均要求不依赖于化石燃料且环境低负荷的物质生产方式。使用了微生物的物质生产由于能够在常温常压的环境下进行,并且随着近年来的基因操作技术的发展而变得能够生产广泛的化合物种类,因此作为满足上述要求的生产系统而受到关注。其中,由于蓝细菌及藻类等光合作用微生物将光作为能量源而能够将空气中的二氧化碳(co2)用作碳原料,因此作为碳中和的下一代的物质生产系统而特别受到期待。
3.例如,作为使用蓝细菌生产的物质,报告有乙醇(非专利文献1)、异丁醇(非专利文献2)、烷烃类(专利文献1)、脂肪酸(专利文献2)以及蛋白(非专利文献3)等。
4.另外,例如在非专利文献4中还公开了使蓝细菌表达蛋白的方法。
5.现有技术文献
6.专利文献
7.专利文献1:日本特开2015-109828号公报
8.专利文献2:日本专利第6341676号公报
9.非专利文献
10.非专利文献1:jason dexter and pengcheng fu,“metabolic engineering of cyanobacteria for ethanol production”,energy&environmental science,royal society of chemistry,2009,vol.2,pp.857-864
11.非专利文献2:shota atsumi et al.,“direct photosynthetic recycling of carbon dioxide to isobutyraldehyde”,nature biotechnology,nature publishing group,2009,vol.27,pp.1177-1180
12.非专利文献3:jie zhou et al.,“discovery of a super-strong promoter enable efficient production of heterologous proteins in cyanobacteria”,scientific reports,nature research,2014,vol.4,article no.4500
13.非专利文献4:andrew h.ng et al.,“fine-tuning of photoautotrophic synechocystis sp.strain pcc 6803”,applied and environmental microbiology,american society for microbiology,2015,vol.81,pp.6857-6863
技术实现要素:
14.发明所要解决的课题
15.但是,在上述现有技术中,使用蓝细菌难以高效地生产蛋白。
16.因此,本公开提供蛋白的分泌生产率提高了的修饰蓝细菌、修饰蓝细菌的制造方
法、以及使用了修饰蓝细菌的蛋白的制造方法。
17.用于解决课题的手段
18.本公开的一个方式的修饰蓝细菌表达使外膜的蛋白透过性提高的通道蛋白。
19.发明效果
20.根据本公开的修饰蓝细菌及修饰蓝细菌的制造方法,能够提供蛋白的分泌生产率提高了的修饰蓝细菌。另外,根据本公开的蛋白的制造方法,能够高效地制造蛋白。
附图说明
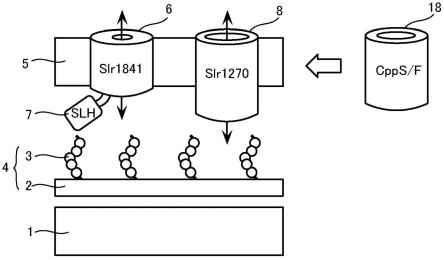
21.图1是示意性地示出了蓝细菌的细胞表层的图。
22.图2是实施例1及比较例1的修饰蓝细菌在培养上清液中的蛋白量(n=3、误差条=sd)的图表。
具体实施方式
23.(成为本公开基础的见解)
24.蓝细菌(cyanobacteria,也称为蓝色细菌或蓝藻)是真细菌的一组,通过光合作用将水分解而产生氧,通过得到的能量将空气中的co2固定。需要说明的是,蓝细菌根据种类的不同,也可以固定空气中的氮(n2)。另外,作为蓝细菌的特性,已知生长快、光利用效率高,并且由于与其他藻类种类相比,基因操作更容易,因此在光合作用微生物中,关于蓝细菌的利用进行了活跃的研究开发。如上所述,作为使用了蓝细菌的物质生产的例子,报告了乙醇(非专利文献1)、异丁醇(非专利文献2)、烷烃类(专利文献1)以及脂肪酸(专利文献2)等燃料的生产。
25.另外,还进行了有关使用蓝细菌生产成为生物的营养源的物质的研究开发。例如,蛋白只能在生物中合成,因此认为作为简便且高效地生产蛋白的生物种类中的1种,利用光能和大气中的co2的光合作用微生物是有用的。
26.例如,在非专利文献3中记载了使用了蓝细菌集胞藻(synechocystis sp.)pcc 6803的蛋白的生产方法。在该文献中,公开了用于使乙烯合成酶的基因的转录高效率地活化的启动子的碱基序列和用于增强翻译的核糖体结合序列。
27.另外,非专利文献4中公开了在将集胞藻pcc 6803作为宿主表达重组蛋白的情况下,在该菌株所保持的质粒上的ns-pcc2区域插入编码蛋白的基因的方法。并报告了通过使用该方法,与在染色体dna(deoxyribonucleic acid,脱氧核糖核酸)上插入该基因的情况相比,重组蛋白的表达效率提高至约14倍。
28.然而,在上述现有技术中,为了回收在pcc 6803株的菌体内所产生的蛋白,需要将pcc 6803株的菌体破碎。因此,蛋白的回收很费功夫。另外,由于菌体内存在各种物质,因此有时需要除去这些物质以生成目标蛋白。因此,蛋白的回收率降低。另外,每次生产蛋白时,都需要准备新的菌株,因此费功夫,生产成本也增加。如此,在上述的现有技术中,使用了蓝细菌的蛋白的生产效率仍处于低水平,期望开发出生产效率更高的技术。
29.因此,本发明人发现了,通过使植物的叶绿体来源的外膜通道蛋白在蓝细菌的外膜中表达,外膜的蛋白透过性提高。由此发现了,每单位蓝细胞的培养液或每个细胞所分泌生产的蛋白量增大。
30.因此,根据本公开,能够提供蛋白的分泌生产率提高了的修饰蓝细菌。另外,若对本公开的修饰蓝细菌进行培养,则还能够高效地制造蛋白。
31.(本公开的概要)
32.本公开的一个方式的概要如下。
33.本公开的一个方式的修饰蓝细菌表达使外膜的蛋白透过性提高的通道蛋白。
34.由此,外膜的蛋白透过性提高。因此,根据本公开的一个方式的修饰蓝细菌,能够提供蛋白的分泌生产率提高了的蓝细菌。
35.例如,在本公开的一个方式的修饰蓝细菌中,所述通道蛋白可以是由序列号1所示的氨基酸序列构成的cpps、由序列号2所示的氨基酸序列构成的cppf、或者氨基酸序列的50%以上与这些中的任一个通道蛋白相同的蛋白。
36.由此,在修饰蓝细菌中,表达作为使外膜的蛋白透过性提高的通道蛋白的cpps(序列号1)或cppf(序列号2)、或具有与这些中的任一个通道蛋白具有同等的功能的蛋白。因此,根据本公开的一个方式的修饰蓝细菌,外膜的蛋白透过性提高。
37.例如,在本公开的一个方式的修饰蓝细菌中,可以导入有编码所述通道蛋白的基因。
38.由此,表达使外膜的蛋白透过性提高的通道蛋白。因此,根据本公开的一个方式的修饰蓝细菌,外膜的蛋白透过性提高,因此能够提供蛋白的分泌生产率提高了的蓝细菌。
39.例如,在本公开的一个方式的修饰蓝细菌中,所述基因可以是叶绿体来源的基因。例如,在本公开的一个方式的修饰蓝细菌中,所述基因可以是由序列号3所示的碱基序列构成的cpps、由序列号4所示的碱基序列构成的cppf、或者碱基序列的50%以上与这些中的任一个基因相同的基因。
40.由此,在修饰蓝细菌中,导入有编码上述的序列号3和序列号4所示的任一个通道蛋白的基因或与这些中的任一个基因的碱基序列的50%以上相同的基因。因此,在修饰蓝细菌中,表达具有使外膜中的蛋白透过性提高的功能的蛋白或与该蛋白具有同等功能的蛋白。因此,在本公开的一个方式的修饰蓝细菌中,外膜的蛋白透过性提高。
41.另外,本公开的一个方式的修饰蓝细菌的制造方法包含使通道蛋白表达的步骤,所述通道蛋白使外膜的蛋白透过性提高。
42.由此,能够表达使蓝细菌的外膜的蛋白透过性提高的通道蛋白。因此,在菌体内产生的蛋白变得容易从该通道蛋白透过而分泌到外膜之外(即菌体之外)。因此,根据本公开的一个方式的修饰蓝细菌的制造方法,能够制造外膜的蛋白透过性提高了的修饰蓝细菌。
43.另外,本公开的一个方式的蛋白的制造方法包含培养上述的任一个修饰蓝细菌的步骤。
44.由此,在修饰蓝细菌的菌体(也称为细胞)内产生的蛋白被分泌到菌体外(即培养液中),因此能够回收分泌到培养液中的蛋白。因此,无需为了回收蛋白而收集菌体并进行破碎。另外,在回收蛋白后,还可以重复使用菌体。另外,在修饰蓝细菌的培养中,不需要特别的营养,仅通过给予光、水、空气以及微量的无机物就能够简便地进行培养。因此,根据本公开的一个方式的蛋白的制造方法,能够高效地制造蛋白。
45.以下,参照附图对实施方式具体地进行说明。
46.需要说明的是,以下说明的实施方式均表示总括性或具体的例子。在以下的实施
方式中示出的数值、材料、步骤、步骤的顺序等是一个例子,并不是为了限定本公开。另外,关于以下实施方式中的构成要素中的在表示最上位概念的独立权利要求中没有记载的构成要素,作为任意的构成要素进行说明。
47.另外,各图不一定是严格地进行图示的图。在各图中,对实质上相同的构成标注相同的附图标记,有时省略或简化重复的说明。
48.另外,下文中,数值范围并不是仅表示严格意义,还包括实质上同等的范围,例如对蛋白的量(例如数或浓度等)或其范围进行测量等。
49.另外,在本说明书中,菌体和细胞均表示1个蓝细菌的个体。
50.(实施方式)
51.[1.定义]
[0052]
本说明书中,碱基序列和氨基酸序列的同源性通过blast(basic local alignment search tool,基于局部比对算法的搜索工具)算法来计算。具体而言,通过用在ncbi(national center for biotechnology information,美国国立生物技术信息中心)(https://blast.ncbi.nlm.nih.gov/blast.cgi)的网页中可利用的blast程序进行成对(pair-wise)分析来计算。关于蓝细菌和植物的基因以及蛋白的信息例如在上述ncbi数据库及cyanobase(http://genome.microbedb.jp/cyanobase/)中有公开。可以从这些数据库中获得目标蛋白的氨基酸序列和编码这些蛋白的基因的碱基序列。
[0053]
蓝细菌也被称为蓝藻或蓝色细菌,是通过叶绿素捕集光能,用得到的能量将水电解而产生氧,同时进行光合作用的原核生物的一组。蓝细菌富有多样性,例如在细胞形状方面,有集胞藻pcc 6803那样的单细胞性的种类和鱼腥藻(anabaena sp.)pcc 7120那样的多细胞相连而成的丝状性的种类。在生长环境方面,也有细长嗜热聚球藻(thermosynechococcus elongatus)那样的嗜热性的种类、细长聚球藻(synechococcus elongatus)那样的海洋性的种类、集胞藻属(synechocystis)那样的淡水性的种类。另外,还可举出许多如铜绿微囊藻(microcystis aeruginosa)那样具有气体囊泡并产生毒素的种类、以及不具有类囊体而在细胞膜中具有被称为聚光天线即藻胆体的蛋白的类囊体蓝藻(gloeobacter violaceus)那样具有独自的特征的种类。
[0054]
图1是示意性地示出了蓝细菌的细胞表层的图。如图1所示,蓝细菌的细胞表层从内侧起依次由细胞膜(也称为内膜1)、肽聚糖2、以及形成细胞最外层的脂质膜即外膜5构成。在肽聚糖2上共价键合有由葡糖胺和甘露糖胺等构成的糖链3,另外在这些共价键型的糖链3上键合有丙酮酸。在本说明书中,将包括肽聚糖2和共价键型的糖链3在内地称为细胞壁4。通过细胞壁4的糖链3与后述的离子通道蛋白6的结合域7结合,外膜5将细胞壁4覆盖。另外,细胞膜(即内膜1)与外膜5之间的间隙存在被称为胞外质(periplasm)、参与蛋白的分解或立体结构的形成、脂质或核酸的分解、或者细胞外的营养素的摄入等的各种酶。
[0055]
通道蛋白是指形成用于使规定的物质从脂质膜(例如外膜5)的内侧向外侧、或者从外侧向内侧选择性地透过的路径(即通道)的膜蛋白。在大肠杆菌和沙门氏菌等通常的异养革兰氏阴性细菌的外膜中,大量存在用于使糖和氨基酸等较低分子量的营养素从外膜的外侧向内侧选择性地透过而摄入细胞内的、被称为porin的通道蛋白(非专利文献5:nikaido,2003,microbiol.mol.biol.rev.,67(4):593-656)。另一方面,在蓝细菌的外膜5中不存在porin,取而代之,外膜5中大量地存在仅使无机离子选择性地透过的离子通道蛋
白。该离子通道蛋白占外膜5的总蛋白的约80%(非专利文献6:kowata et al.,2017,j.bacteriol.,199(19):e00371-17)。因此,在蓝细菌中,只要不使用基因导入等技术较大地改变外膜5的性质,蛋白这样的高分子量的物质是难以透过外膜5而扩散到细胞外(即外膜5之外)的。
[0056]
植物的叶绿体在约15~20亿年前起源于原始的真核细胞的细胞内共生的蓝细菌,通过之后的进化而变化为叶绿体(非专利文献7:ponce-toledo et al.,2017,curr.biol.,27(3):386-391)。作为被认为是最原始的植物的单细胞藻类的灰色藻所保持的叶绿体具有肽聚糖,残留有与蓝细菌非常相似的表层结构。另一方面,比单细胞藻类更为进化的种子植物的叶绿体中不存在肽聚糖。另外,蓝细菌的外膜蛋白大多在进化的过程中,在叶绿体的诞生初期从叶绿体的外膜失去了。因此,上文所述的灰色藻的叶绿体的外膜蛋白与蓝细菌的外膜蛋白的结构大不相同。例如,在蓝细菌的外膜5中大量含有slr1841等使无机物透过的离子通道蛋白6。该离子通道蛋白6占外膜5的总蛋白的约80%。另一方面,在灰色藻的叶绿体的外膜中,外膜中大量含有被命名为cpps和cppf的使有机物透过的通道蛋白(以下也称为有机物通道蛋白18)。该有机物通道蛋白18占灰色藻的叶绿体的外膜的总蛋白的80%以上(非专利文献8:kojima et al.,2016,j.biol.chem.,291:20198-20209)。cpps及cppf是具有选择性地使较高分子量的有机物(例如蛋白等生物分子)透过的通道功能的通道蛋白,被认为作为将植物细胞中的叶绿体的内部与植物细胞的细胞质连接的物质输送路径而发挥功能。cpps及cppf广泛地分布于灰色藻类中。另一方面,在细菌中,仅在属于浮霉菌(planctomycetes)门的细菌中存在cpps和cppf的类似蛋白。需要说明的是,蓝细菌不保持有cpps和cppf以及它们的类似蛋白(参照非专利文献8)。
[0057]
在蓝细菌中,通过光合作用而被固定的co2经过多阶段的酶反应而被转换为各种氨基酸。将它们作为原料,在蓝细菌的细胞质内合成蛋白。在这些蛋白中,既存在在细胞质内发挥功能的蛋白,也存在从细胞质被输送至胞外质而在胞外质内发挥功能的蛋白。但是,迄今为止在蓝细菌中尚没有报告过将蛋白主动分泌到细胞外的情况。
[0058]
由于蓝细菌具有较高的光合作用能力,因此不一定需要将有机物作为营养成分从外部摄入。因此,如图1的有机物通道蛋白8(例如slr1270)那样,蓝细菌在外膜5仅具有非常微少的使有机物透过的通道蛋白。例如,在集胞藻pcc 6803中,使有机物透过的有机物通道蛋白8(例如slr1270)仅存在外膜5的总蛋白量的约4%。另一方面,蓝细菌为了高效地将生长所需的无机离子类摄入到细胞内,如图1的离子通道蛋白6(例如slr1841等)那样,在外膜5中具有大量仅使无机离子类透过的离子通道。例如,在集胞藻pcc 6803中,使无机离子透过的离子通道蛋白6占外膜5的总蛋白量的约80%。
[0059]
如此,认为由于在蓝细菌中,外膜5中的使蛋白等有机物透过的通道非常少,因此难以将在菌体内产生的蛋白主动地分泌到菌体外。
[0060]
[2.修饰蓝细菌]
[0061]
接着,参照图1对本实施方式的修饰蓝细菌31进行说明。
[0062]
本实施方式的修饰蓝细菌表达使外膜5的蛋白透过性提高的通道蛋白(所谓的有机物通道蛋白18)。由此,外膜5的蛋白透过性提高,因此将在菌体内产生的蛋白分泌到菌体外的蛋白的分泌生产率提高。另外,由于不需要将菌体破碎而回收蛋白,因此即使在回收蛋白后,也能够重复使用修饰蓝细菌而产生蛋白。另外,在本说明书中,将修饰蓝细菌在菌体
内制造蛋白称为产生,将所产生的蛋白分泌到菌体外的情况称为分泌生产。
[0063]
作为本实施方式的修饰蓝细菌的亲本微生物的、使外膜5的蛋白透过性提高之前的蓝细菌(以下称为亲本细菌)的种类没有特别限制,可以是任何种类的蓝细菌。例如,亲本蓝细菌可以是集胞藻(synechocystis)属、聚球藻(synechococcus)属、鱼腥藻(anabaena)属、或嗜热聚球藻(thermosynechococcus)属,其中,可以是集胞藻(synechocystis sp.)pcc6803、聚球藻(synechococcus sp)pcc 7942或嗜热聚球藻(thermosynechococcus elongatus)bp-1。
[0064]
需要说明的是,在本说明书中,使有机物通道蛋白18在蓝细菌的外膜5中表达是指将编码有机物通道蛋白18的基因插入到蓝细菌的染色体dna或质粒中,经过该基因的转录和翻译而合成的有机物通道蛋白18被输送到外膜5,在蓝细菌的外膜5中表现出使蛋白选择性地透过的通道功能。该基因的插入和表达的方法只要是通常使用的方法就没有特别限定,并不受用于转录活化的启动子的碱基序列和用于翻译的核糖体结合序列、以及用于向外膜5输送的信号序列的种类的限制。
[0065]
在本实施方式中,在蓝细菌的外膜5中表达的有机物通道蛋白18可以是叶绿体来源的外膜通道蛋白。该有机物通道蛋白18例如可以是灰色藻cyanophora paradoxa(蓝色奇异矽藻,以下也称为c.paradoxa)的cpps(序列号1)或cppf(序列号2)等。另外,有机物通道蛋白18也可以是氨基酸序列的50%以上与cpps或cppf相同的蛋白。需要说明的是,氨基酸序列的50%以上与cpps或cppf相同的蛋白不限于叶绿体来源的蛋白,例如也可以是细菌等微生物来源的cpps或cppf的类似蛋白。
[0066]
由此,在修饰蓝细菌中,表达作为使外膜5的蛋白透过性提高的有机物通道蛋白18的cpps(序列号1)或cppf(序列号2)、或者与它们中的任一个有机物通道蛋白18具有同等的功能的蛋白。因此,本实施方式的修饰蓝细菌的外膜5的蛋白透过性提高。
[0067]
通常认为,如果蛋白的氨基酸序列的30%以上相同,则蛋白的立体结构的同源性高,因此与该蛋白具有同等功能的可能性高。因此,作为有机物通道蛋白18,例如可以是由与上述序列号1和序列号2所示的蛋白中的任一个的氨基酸序列具有40%以上、优选50%以上、更优选60%以上、进一步优选70%以上、更进一步优选80%以上、再进一步优选90%以上的同源性的氨基酸序列构成、且具有使外膜5的蛋白透过性提高的功能的蛋白或多肽。
[0068]
另外,在本实施方式中,修饰蓝细菌也可以导入有编码有机物通道蛋白18的基因。由此,在修饰蓝细菌中,表达使外膜5的蛋白透过性提高的有机物通道蛋白18。
[0069]
上述基因例如可以是叶绿体来源的基因。编码叶绿体来源的有机物通道蛋白18的基因例如可以是灰色藻cyanophora paradoxa的cpps(序列号3)或cppf(序列号4)。另外,有机物通道蛋白18也可以是碱基序列的50%以上与这些中的任一个基因相同的基因。由此,在修饰蓝细菌中,导入有编码上述序列号3和序列号4所示的任一个有机物通道蛋白18的基因或与这些中的任一个基因的碱基序列的50%以上相同的基因。因此,表达具有使外膜5中的蛋白透过性提高的功能的蛋白或与该蛋白具有同等功能的蛋白。由此,在修饰蓝细菌中,外膜5的蛋白透过性提高。
[0070]
另外,作为编码有机物通道蛋白18的基因,并不限于叶绿体来源的基因。作为编码有机物通道蛋白18的基因,例如可以是由与上述基因cpps(序列号3)和cppf(序列号4)中的任一个碱基序列具有40%以上、优选50%以上、更优选60%以上、进一步优选70%以上、更
进一步优选80%以上、再进一步优选90%以上的同源性的碱基序列构成、且编码具有使外膜5的蛋白透过性提高的功能的蛋白或多肽的基因。
[0071]
[3.修饰蓝细菌的制造方法]
[0072]
接着,对本实施方式的修饰蓝细菌的制造方法进行说明。修饰蓝细菌的制造方法包含使有机物通道蛋白18表达的步骤,所述有机物通道蛋白18使外膜的蛋白透过性提高。
[0073]
在本实施方式中,使外膜5的蛋白透过性提高的有机物通道蛋白18例如是叶绿体来源的通道蛋白,具体而言,可以是由序列号1所示的氨基酸序列构成的cpps、或者由序列号2所示的氨基酸序列构成的cppf。另外,有机物通道蛋白18也可以是氨基酸序列的50%以上与这些中的任一个蛋白相同的蛋白。
[0074]
在使有机物通道蛋白18表达的步骤中,首先,编码使外膜5的蛋白透过性提高的有机物通道蛋白18的基因被插入到蓝细菌的染色体dna或质粒中。然后,经过该基因的转录和翻译而合成的有机物通道蛋白18被输送到外膜5,在蓝细菌的外膜5中表现出通道功能。其中,基因的插入及表达的方法只要是通常使用的方法就没有特别限定,并不受用于转录活化的启动子的碱基序列及用于翻译的核糖体结合序列、以及用于向外膜5输送的信号序列的种类的限制。
[0075]
通过以上的步骤,可以表达使外膜5的蛋白透过性提高的通道蛋白,从而制造修饰蓝细菌。
[0076]
由此,通过本实施方式的制造方法制造的修饰蓝细菌在外膜5中表达使蛋白透过性提高的有机物通道蛋白18,因此外膜5的蛋白透过性提高。因此,在菌体内产生的蛋白变得容易从该有机物通道蛋白18透过而分泌到外膜5之外(即菌体之外)。因此,根据本实施方式的修饰蓝细菌的制造方法,能够提供蛋白的分泌生产率提高了的修饰蓝细菌。
[0077]
另外,在通过本实施方式的制造方法制造的修饰蓝细菌中,如上所述,在菌体内产生的蛋白扩散到菌体外,因此不需要为了回收蛋白而破碎菌体。例如,只要在适当的条件下培养修饰蓝细菌,接着回收分泌到培养液中的蛋白即可,因此还能够一边培养修饰蓝细菌一边回收培养液中的蛋白。因此,如果使用通过本制造方法得到的修饰蓝细菌,则能够实施高效的微生物蛋白生产。因此,根据本实施方式的修饰蓝细菌的制造方法,能够提供在回收蛋白后还能够重复使用的利用效率高的修饰蓝细菌。
[0078]
需要说明的是,通过本实施方式的修饰蓝细菌的制造方法制造的修饰蓝细菌将肽酶或磷酸酶等主要是原本存在于胞外质中的蛋白组分泌到细胞外。在本实施方式中,例如,通过对编码如存在于胞外质中的蛋白组那样原本在蓝细菌的细胞内产生的蛋白的基因进行修饰,替换为编码其他蛋白的基因,还能够使修饰蓝细菌产生所希望的蛋白。因此,根据本实施方式的修饰蓝细菌的制造方法,还能够提供能够简便且高效地产生所希望的蛋白的修饰蓝细菌。
[0079]
[4.蛋白的制造方法]
[0080]
接着,对本实施方式的蛋白的制造方法进行说明。本实施方式的蛋白的制造方法包含培养上述修饰蓝细菌的步骤。
[0081]
蓝细菌的培养通常可以基于使用了bg-11培养基(参照表2)的液体培养或其变通方法来实施。因此,修饰蓝细菌的培养也可以同样地实施。另外,作为用于制造蛋白的蓝细菌的培养期间,只要是能够在菌体充分增殖了的条件下以高浓度蓄积蛋白的方式进行的期
间即可,例如可以是1~3天,也可以是4~7天。另外,培养方法例如可以是通气搅拌培养或振荡培养。
[0082]
通过在上述条件下进行培养,修饰蓝细菌在菌体内产生蛋白,并将该蛋白分泌到培养液中。回收被分泌到培养液中的蛋白时,可以通过对培养液进行过滤或离心分离等,从培养液中除去细胞(所谓的菌体)等固体成分来回收培养上清液。根据本实施方式的蛋白的制造方法,由于蛋白被分泌到修饰蓝细菌的细胞外,因此不需要为了回收蛋白而破碎细胞。因此,蛋白回收后能够重复使用留下的修饰蓝细菌来进行蛋白的制造。
[0083]
需要说明的是,分泌到培养液中的蛋白的回收方法并不限于上述的例子,也可以一边培养修饰蓝细菌一边回收培养液中的蛋白。例如,也可以通过使用使蛋白透过的透过膜来回收从透过膜透过的蛋白。在该情况下,也可以将从透过膜透过的蛋白作为营养源,培养乳酸菌等有用微生物。这样,能够一边培养修饰蓝细菌一边回收培养液中的蛋白,因此不需要从培养液中除去修饰蓝细菌的菌体的处理。因此,能够更简便且高效地制造蛋白。
[0084]
另外,由于不需要从培养液中回收菌体的处理和菌体的破碎处理,因此能够降低修饰蓝细菌受到的损伤和应激。因此,修饰蓝细菌的蛋白分泌生产率不易降低,能够更长期地使用修饰蓝细菌。
[0085]
如以上那样,根据本实施方式的使用了修饰蓝细菌的蛋白的制造方法,能够简便且高效地得到用于制造食品成分原料或化合物的酶、医疗用领域中的诊断用酶或治疗用酶、或者农渔畜产领域中的饲料用酶等。
[0086]
实施例
[0087]
以下,用实施例对本公开的修饰蓝细菌、修饰蓝细菌的制造方法以及蛋白的制造方法具体地进行说明,但本公开并不仅仅限定于以下的实施例。
[0088]
在本实施例中,作为提高蓝细菌的外膜的蛋白透过性的方法,使灰色藻cyanophora paradoxa所保持的叶绿体的外膜通道蛋白cpps(序列号1)在蓝细菌的外膜中表达。然后,进行表达了外膜通道蛋白cpps(序列号1)的修饰蓝细菌的蛋白分泌生产率的测定和所分泌的蛋白的鉴定。另外,本实施例中使用的蓝细菌种类是集胞藻pcc 6803(以下简称为“蓝细菌”)。
[0089]
(1)cpps基因表达盒的构建
[0090]
按照以下的步骤制作cpps基因、用于cpps基因的表达控制的启动子区域(pl22)、蓝细菌中的外膜转移信号序列(slr0042-signal)、和作为基因导入的标记的卡那霉素抗性标记基因(kmr)连接而成的基因盒。
[0091]
首先,以蓝细菌的染色体dna为模板,使用表1中记载的引物slr0042-fw(序列号5)和slr0042-rv(序列号6)通过pcr(聚合酶链反应)法进行扩增,得到slr0042基因。接着,以集胞藻(synechocystis)ly07株(非专利文献9:yao et al.,acs synth.biol.,2016,5(3):207-212)的染色体dna为模板,使用表1中记载的引物slr2030-fw(序列号7)和pl22-rv(序列号8)的组以及kmr-fw(序列号9)和slr2031-rv(序列号10)的组通过pcr法进行扩增,得到pl22和kmr。其中,ly07株中,它们插入染色体上的slr2030-slr2031基因中而存在,因此使用上述4个引物通过pcr法进行扩增时,以在pl22的5’末端侧连接有slr2030基因片段的形式被扩增,以在kmr的3’末端侧连接有slr2031基因片段的形式被扩增。接着,以上述步骤中得到的slr0042基因、pl22和kmr的混合溶液为模板,使用表1中记载的4个引物(序列号7~
10)通过pcr法进行扩增,由此得到从5’末端侧依次连接有slr2030基因片段、pl22、slr0042基因、kmr和slr2031基因片段的基因盒(slr2030-2031::slr0042-kmr盒)。使用in-fusion pcr克隆法(注册商标),将slr2030-2031::slr0042-kmr盒插入到puc19质粒中,得到puc19-slr0042质粒。
[0092]
与上述步骤并行地,使用smart cdna文库合成试剂盒(clontech)由灰色藻c.paradoxa nies-547制备总cdna。以该cdna为模板,使用表1中记载的引物cpps-fw(序列号11)和cpps-rv(序列号12)通过pcr法进行扩增,得到cpps基因。使用in-fusion pcr克隆法(注册商标),将cpps基因插入到puc19-slr0042质粒中,得到puc19-cpps质粒。其中,通过该步骤,cpps基因以与sr0042基因的外膜转移信号序列的3’末端侧连接的形式被插入,除sr0042基因的外膜转移信号序列以外的区域以插入替换为cpps的编码区域的形式被除去。
[0093]
表1
[0094]
引物名碱基序列序列号slr0042-fw5
′‑
tagatataatgtgtggtaaattagaggaattcatcgca-3
′
5slr0042-rv5
′‑
cgctcttccgatctgatttaaaatgtgaacgtcgtccgtag-3
′
6slr2030-fw5
′‑
cagtgaattcgagctcggtaataaccgttgtcccttttgtttcatcg-3
′
7pl22-rv5
′‑
tgttagtgagccctgctgttagctcccagtatctctatcactgat-3
′
8kmr-fw5
′‑
acagcagggctcactaacagttttagagctagaaatagcaagttaaaataa-3
′
9slr2031-rv5
′‑
gattacgccaagcttgcatggggaacaagctgaatctgggcatc-3
′
10cpps-fw5
′‑
tccctggttccgctggggctatggccccggagtgcaccgt-3
′
11cpps-rv5
′‑
atttgatgcctggctctagtttacagaagctcgatctcga-3
′
12psba1-fw5
′‑
cagtgaattcgagctcggtatatagcgttgcagtccctgg-3
′
13tetr-rv5
′‑
tagcaggtactgcaaacgctatgccgctcccggtgttcgc-3
′
14tetr-fw5
′‑
ttagaactggtaaacgatacttactgatcgataacgagcg-3
′
15psba1-rv5
′‑
gattacgccaagcttgcatgaccgcggtcacttcataacc-3
′
16
[0095]
(2)pl22的启动子活性控制盒(tetr)的构建
[0096]
上述pl22的启动子活性通过借助tetr抑制因子的控制,仅在脱氢四环素(atc)的存在下被诱导。因此,需要将用于pl22的活性控制的tetr基因导入到修饰蓝细菌中。
[0097]
首先,以ly07株的染色体dna为模板,使用表1中记载的引物psba1-fw(序列号13)和tetr-rv(序列号14)组、以及tetr-fw(序列号15)和psba1-rv(序列号16)的组通过pcr法进行扩增,得到tetr基因和作为基因导入的标记的大观霉素抗性标记基因(spcr)。其中,ly07株中,它们插入到染色体上的psba1基因中而存在,因此使用上述引物通过pcr法进行扩增时,以在tetr基因的5’末端侧连接有psba1基因的上游侧片段的形式被扩增,以在spcr的3’末端侧连接有psba1基因的下游侧片段的形式被扩增。接着,以tetr基因和spcr的混合溶液为模板,使用表1中记载的引物psba1-fw(序列号13)和psba1-rv(序列号14)通过pcr法进行扩增,由此得到从5’末端侧起依次连接有psba1基因上游侧片段、tetr、spcr、psba1基因下游侧片段的基因盒(psba1::tetr盒)。使用in-fusion pcr克隆法(注册商标),将psba1::tetr盒插入到puc19质粒中,得到puc19-tetr质粒。
[0098]
(3)cpps基因表达盒和tetr盒的导入
[0099]
将通过上述步骤得到的puc19-cpps质粒1μg与蓝细菌培养液(菌体浓度od730=0.5左右)混合,通过自然转化将质粒导入到细胞内。使转化后的细胞在含有30μg/ml卡那霉
素的bg-11琼脂培养基上生长,由此进行筛选。在所筛选的细胞中,染色体上的slr2030-2031基因与puc19-cpps质粒上的slr2030基因片段区域和slr2031基因片段区域之间发生同源重组。由此,得到在slr2030-2031基因区域插入有cpps基因表达盒的synechocystis cpps株。其中,所使用的bg-11培养基的组成如表2所示。
[0100]
表2
[0101]
成分含量(mg/l)edta-na1柠檬酸铁铵6nano31500mgso475k2hpo439cacl228.6h3bo42.8mncl21.8znso40.2cuso40.08na2moo40.02co(no3)20.005tes-koh(ph 7.5)4580
[0102]
接着,将puc19-tetr质粒1μg与集胞藻cpps培养液(菌体浓度od730=0.5左右)混合,通过自然转化将质粒导入到细胞内。使转化后的细胞在含有30μg/ml的卡那霉素和20μg/ml的大观霉素的bg-11琼脂培养基上生长,由此进行筛选,得到集胞藻cpps tetr株。与上述同样地,在该菌株中,在染色体dna上的psba1基因中插入有tetr盒。
[0103]
(4)蛋白的分泌生产率试验
[0104]
将上述(3)中得到的集胞藻cpps tetr株在以下的实施例1和比较例1中记载的条件下分别培养,测定分泌到细胞外的蛋白量(以下也称为分泌蛋白量)。根据培养液中的蛋白量,评价蛋白的分泌生产率。其中,蛋白的分泌生产率是指通过将在细胞内产生的蛋白分泌到细胞外,从而生产蛋白的能力。以下,对具体的方法进行说明。
[0105]
(4-1)修饰株的培养
[0106]
[实施例1]
[0107]
在实施例1中,以初始菌体浓度od730=0.05的方式将集胞藻cpps tetr株接种于含有1μg/ml atc的bg-11培养基中,在光量为100μmol/m2/s、30℃的条件下振荡培养5天。培养独立地进行3次。
[0108]
[比较例1]
[0109]
在比较例1中,使用不含atc的bg-11培养基,除此以外,在与实施例1相同的条件下培养集胞藻cpps tetr株。培养独立地进行3次。
[0110]
(4-2)所分泌生产的蛋白的定量
[0111]
将上述(4-1)的实施例1中得到的培养液在室温下、以2,500g离心分离10分钟,得到培养上清液。使用孔径为0.22μm的膜滤器对得到的培养上清液进行过滤,完全除去集胞
藻cpps tetr株的细胞。通过bca(bicinchoninic acid,二辛可宁酸)法对过滤后的培养上清液中所含的总蛋白量进行定量。对独立培养的3个培养液分别进行该一系列操作,求出实施例1的集胞藻cpps tetr株的分泌到细胞外的蛋白量的平均值和标准偏差。另外,对于在比较例1的条件下培养的集胞藻cpps tetr株,也以同样的条件进行3个培养液的蛋白量的定量,求出3个培养液中的蛋白量的平均值和标准偏差。
[0112]
将结果示于图2。图2是表示实施例1和比较例1的修饰蓝细菌的培养上清液中的蛋白量(n=3、误差条=sd)的图表。
[0113]
如图2所示,在实施例1的atc存在下培养的集胞藻cpps tetr株中,与比较例1的无atc存在下培养的集胞藻cpps tetr株相比,培养上清液中分泌的蛋白量(mg/l)提高了约25倍。
[0114]
虽然省略数据的记载,但测定培养液的吸光度(730nm),算出每1g菌体干燥重量的分泌蛋白量(mg蛋白/g菌体干燥重量),结果在实施例1的条件下培养的集胞藻cpps tetr株的每1g菌体干燥重量的分泌蛋白量(mg蛋白/g菌体干燥重量)与在比较例1的条件下培养的集胞藻cpps tetr株相比,提高了约36倍。
[0115]
(5)所分泌的蛋白的鉴定
[0116]
接着,通过lc-ms/ms鉴定上述(4-2)中得到的培养上清液中所含的分泌蛋白。以下对方法进行说明。
[0117]
(5-1)试样制备
[0118]
加入相对于培养上清液的液量为8倍量的冷丙酮,在20℃下静置2小时后,在20,000g下离心分离15分钟,得到蛋白的沉淀物。在该沉淀物中加入100mm tris ph8.5、0.5%十二烷酸钠(sdod),利用密闭式超声波破碎机溶解蛋白。调整至蛋白浓度为1μg/ml后,添加终浓度为10mm的二硫苏糖醇(dtt),在50℃下静置30分钟。接着,添加终浓度为30mm的碘乙酰胺(iaa),在室温(遮光)下静置30分钟。为了停止iaa的反应,添加终浓度为60mm的半胱氨酸,在室温下静置10分钟。添加胰蛋白酶400ng,在37℃下静置一晚,将蛋白进行肽片段化。加入5%tfa(trifluoroacetic acid,三氟乙酸)后,在室温下以15,000g离心分离10分钟,得到上清液。通过该操作除去sdod。使用c18旋转柱进行脱盐后,利用离心蒸发器使试样干固。然后,加入3%乙腈、0.1%甲酸(formic acid),使用密闭式超声波破碎机将试样溶解。以肽浓度达到200ng/μl的方式进行制备。
[0119]
(5-2)lc-ms/ms分析
[0120]
使用lc-ms/ms装置(ultimate 3000rslcnanolc system)在以下的条件下对上述(5-1)中得到的试样实施分析。
[0121]
试样注入量:200ng
[0122]
柱:capcell core mp 75μm
×
250mm
[0123]
溶剂:a溶剂为0.1%甲酸水溶液,b溶剂为0.1%甲酸 80%乙腈
[0124]
梯度程序(gradient program):试样注入4分钟后使b溶剂为8%、27分钟后使b溶剂为44%、28分钟后使b溶剂为80%、34分钟后测定结束(5-3)数据分析
[0125]
得到的数据在以下的条件下进行分析,进行蛋白和肽的鉴定以及定量值的计算。
[0126]
软件:scaffold dia
[0127]
数据库:uniprotkb/swiss prot database(集胞藻pcc 6803)
[0128]
破碎(fragmentation):hcd
[0129]
前体公差(precursor tolerance):8ppm
[0130]
片段公差(fragment tolerance):10ppm
[0131]
数据采集类型(data acquisition type):重叠dia
[0132]
肽长度(peptide length):8-70
[0133]
肽电荷(peptide charge):2-8
[0134]
最大漏切数量(max missed cleavages):1
[0135]
固定修饰(fixed modification):半胱氨酸碘乙酰胺化(carbamidomethylation)
[0136]
肽fdr(peptide fdr):1%以下
[0137]
从所鉴定的蛋白中相对定量值最大的蛋白起依次将10种蛋白示于表3。
[0138]
表3
[0139] 蛋白名uniprot登录id基因名1羧基末端蛋白酶p73458prc2sii0314蛋白q55648sii03143sii0470蛋白q55847sii04704ssr1853蛋白p72639ssr18535sii0319蛋白p74789sii03196slr0581蛋白q55408slr05817二氧化碳-浓缩机制p72761ccmk28蛋白ccmk同源物2q55386slr09249slr0924蛋白q59991slr004210slr0042蛋白p73817sii1951
[0140]
10种蛋白全部分别包含在实施例1和比较例1的培养上清液中。在这些蛋白的全部中,保持有胞外质(指外膜与内膜的间隙)转移信号。根据该结果确认了,在实施例1的修饰株中,通过cpps基因的表达,外膜的蛋白透过性提高,胞外质内的蛋白变得容易从通道蛋白cpps透过而分泌到外膜之外(即菌体外)。因此,显示出本实施方式的修饰蓝细菌的蛋白的分泌生产率大幅提高。
[0141]
(6)考察
[0142]
本实施例中,确认了本公开的修饰蓝细菌将存在于菌体内(在此为胞外质内)的蛋白分泌到菌体外。另外,对修饰蓝细菌所分泌的蛋白进行了鉴定,由此能够通过对编码这些蛋白的基因进行基因修饰而替换为编码其他蛋白的基因。例如,为了代替上述鉴定的蛋白而产生所期望的蛋白,通过对本公开的修饰蓝细菌进行基因修饰并使用修饰蓝细菌能够高效地制造所期望的蛋白。另外,由于蓝细菌的光合作用能力高,因此通过给予光、水、空气和微量的无机物进行培养,能够在需要时简便地得到所需要的蛋白,因此不需要为了制造蛋白而使用复杂的装置。另外,蛋白例如在加工成营养补充剂等时,其功能容易丧失。因此,根据本公开的修饰蓝细菌,能够在维持蛋白的功能的状态下提供蛋白。通过以上的优点,可期待本公开的修饰蓝细菌在各个领域中的应用。
[0143]
以上,基于实施方式对本公开的修饰蓝细菌、修饰蓝细菌的制造方法以及蛋白的制造方法进行了说明,但本公开并不限定于这些实施方式。只要不脱离本公开的主旨,对实
施方式实施本领域技术人员可想到的各种变形而得到的方式、将实施方式中的一部分构成要素组合而构建的其他方式也包含在本公开的范围内。
[0144]
产业上的可利用性
[0145]
根据本公开的修饰蓝细菌、修饰蓝细菌的制造方法、以及使用了修饰蓝细菌的蛋白的制造方法,通过将水、光、空气、以及微量的无机物给予修饰蓝细菌来进行培养,能够高效地得到蛋白。例如,能够得到用于制造食品成分的原料或化合物的酶、医疗用领域中的诊断用酶或治疗用酶、或者农渔畜产领域中的饲料用酶等。
[0146]
符号说明
[0147]1ꢀꢀꢀꢀꢀꢀ
内膜
[0148]2ꢀꢀꢀꢀꢀꢀ
肽聚糖
[0149]3ꢀꢀꢀꢀꢀꢀ
糖链
[0150]4ꢀꢀꢀꢀꢀꢀ
细胞壁
[0151]5ꢀꢀꢀꢀꢀꢀ
外膜
[0152]6ꢀꢀꢀꢀꢀꢀ
离子通道蛋白
[0153]7ꢀꢀꢀꢀꢀꢀ
结合域
[0154]
8、18
ꢀꢀꢀ
有机物通道蛋白
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。