1.本发明属于生物技术领域,具体涉及一种培育花青素含量降低和开花时间推迟的转基因植物的方法。
背景技术:
2.植物开花标志着从营养阶段转换到生殖阶段,在植物后代繁衍和生态适应过程中发挥着重要作用;其中作物开花的早晚对作物的高产、稳产、品质和区域分布具有重要影响。育种家们将适宜的开花时间作为关键农艺性状之一,一直致力于获得具有理想开花时间的优异种质。对植物开花时间的解析一直是植物分子遗传学、进化生物学、生态学和作物育种学等多个学科的研究热点。目前,豆科牧草开花时间调控的分子机理还尚不清晰,有待进一步研究。
3.花青素是广泛存在于植物中的水溶性黄酮类化合物,是构成花瓣和果实颜色的主要色素之一,在植物生理上发挥重要作用;同时花青素也是天然的植物抗氧化剂,能够保护植物和人体免受自由基等有害物质的损伤,对人类的健康具有诸多益处。
技术实现要素:
4.本发明的目的为促进植物花青素合成。
5.本发明首先保护eaf2蛋白的应用,可为a1)-a6)中的至少一种:
6.a1)调控花青素合成;
7.a2)培育花青素含量改变的转基因植物;
8.a3)调控植物开花时间;
9.a4)培育开花时间改变的转基因植物;
10.a5)调控植物开花节数;
11.a6)培育开花节数改变的转基因植物。
12.上述应用中,所述eaf2蛋白可为如下a1)或a2)或a3)或a4):
13.a1)氨基酸序列是seq id no:2所示的蛋白质;
14.a2)在seq id no:2所示的蛋白质的n端或/和c端连接标签得到的融合蛋白质;
15.a3)将a1)或a2)所示的蛋白质经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与植物花青素合成、开花节数和/或开花时间相关的蛋白质;
16.a4)与a1)或a2)所示的蛋白质具有80%或80%以上同一性,来源于蒺藜苜蓿且与植物花青素合成、开花节数和/或开花时间相关的蛋白质。
17.其中,seq id no:2由140个氨基酸残基组成。
18.为了使蛋白质便于纯化和检测,可在由seq id no:2所示的氨基酸序列组成的eaf2蛋白的氨基末端或羧基末端连接上如表1所示的标签。
19.表1.标签的序列
20.标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tag ii8wshpqfekc-myc10eqkliseedl
21.上述eaf2蛋白可人工合成,也可先合成其编码基因,再进行生物表达得到。
22.上述eaf2蛋白的编码基因可通过将seq id no:1所示的dna序列在其5’末端和/或3’末端连上表1所示的标签的编码序列得到。
23.本发明还保护编码上述任一所述eaf2蛋白的核酸分子的应用,可为a1)-a6)中的至少一种:
24.a1)调控花青素合成;
25.a2)培育花青素含量改变的转基因植物;
26.a3)调控植物开花时间;
27.a4)培育开花时间改变的转基因植物;
28.a5)调控植物开花节数;
29.a6)培育开花节数改变的转基因植物。
30.上述应用中,所述核酸分子可为如下b1)或b2)或b3)或b4)所示的dna分子:
31.b1)编码区是seq id no:1所示的dna分子;
32.b2)核苷酸序列是seq id no:1所示的dna分子;
33.b3)与b1)或b2)限定的核苷酸序列具有75%或75%以上同一性,来源于蒺藜苜蓿且编码上述任一所述eaf2蛋白的dna分子;
34.b4)在严格条件下与b1)或b2)限定的核苷酸序列杂交,来源于蒺藜苜蓿且编码上述任一所述eaf2蛋白的dna分子。
35.其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
36.其中,seq id no:1由423个核苷酸组成,seq id no:1所示的核苷酸编码seqid no:2所示的氨基酸序列。
37.本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码所述eaf2蛋白的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的所述eaf2蛋白的核苷酸序列75%或者更高同一性的核苷酸,只要编码所述eaf2蛋白,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
38.这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码seq id no:2所示的氨基酸序列组成的eaf2蛋白的核苷酸序列具有75%或更高,或80%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。
39.编码所述eaf2蛋白的核酸分子具体可为编码所述eaf2蛋白的基因,命名为eaf2基因。
40.上述任一所述的应用中,所述调控植物花青素合成可为促进植物花青素合成或抑制植物花青素合成。
41.上述任一所述的应用中,所述“培育花青素含量改变的转基因植物”可为培育花青素含量增加的转基因植物或培育花青素含量降低的转基因植物。
42.上述任一所述的应用中,所述“调控植物开花时间”可为植物开花时间提前或植物开花时间推迟。
43.上述任一所述的应用中,所述“培育开花时间改变的转基因植物”可为培育开花时间提前的转基因植物或培育开花时间推迟的转基因植物。
44.上述任一所述的应用中,所述“调控植物开花节数”可为植物开花节数增加或植物开花节数减少。
45.上述任一所述的应用中,所述“培育开花节数改变的转基因植物”可为培育开花节数增加的转基因植物或培育开花节数减少的转基因植物。
46.上述任一所述的应用中,所述植物可为如下c1)至c6)中的任一种:c1)双子叶植物;c2)单子叶植物;c3)豆科植物;c4)蒺藜苜蓿;c5)蒺藜苜蓿r108;c6)突变体eaf2。
47.本发明还保护h1)或h2)。
48.h1)植物育种方法甲,包括如下步骤:增加植物中上述任一所述eaf2蛋白的表达量和/或活性,从而降低花青素含量、开花节数增加和/或开花时间推迟。
49.h2)植物育种方法乙,包括如下步骤:降低植物中上述任一所述eaf2蛋白的表达量和/或活性,从而提高花青素含量、开花节数减少和/或开花时间提前。
50.上述方法中,所述植物可为如下c1)至c6)中的任一种:c1)双子叶植物;c2)单子叶植物;c3)豆科植物;c4)蒺藜苜蓿;c5)蒺藜苜蓿r108;c6)突变体eaf2。
51.本发明还保护一种培育转基因植物甲的方法,可包括如下步骤:提高出发植物甲中上述任一所述eaf2蛋白的表达量和/或活性,得到转基因植物甲;与出发植物甲相比,转基因植物甲的花青素含量降低、开花节数增加和/或开花时间推迟。
52.上述方法中,所述“提高出发植物甲中上述任一所述eaf2蛋白的表达量和/或活性”可通过多拷贝、改变启动子、调控因子、转基因等本领域熟知的方法,达到提高eaf2蛋白的表达量和/或活性的效果。
53.上述方法中,所述“提高出发植物甲中上述任一所述eaf2蛋白的表达量和/或活性”可通过向出发植物甲中导入编码eaf2蛋白的核酸分子实现。
54.上述方法中,编码所述eaf2蛋白的核酸分子可为如下b1)或b2)或b3)或b4)所示的dna分子:
55.b1)编码区是seq id no:1所示的dna分子;
56.b2)核苷酸序列是seq id no:1所示的dna分子;
57.b3)与b1)或b2)限定的核苷酸序列具有75%或75%以上同一性,来源于蒺藜苜蓿且编码上述任一所述eaf2蛋白的dna分子;
58.b4)在严格条件下与b1)或b2)限定的核苷酸序列杂交,来源于蒺藜苜蓿且编码上述任一所述eaf2蛋白的dna分子。
59.其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
60.其中,seq id no:1由423个核苷酸组成,seq id no:1所示的核苷酸编码seq id no:2所示的氨基酸序列。
61.上述方法中,所述“向出发植物甲中导入编码eaf2蛋白的核酸分子”具体可通过向出发植物甲导入含有上述任一所述核酸分子的重组载体实现。所述“含有上述任一所述核酸分子的重组载体”具体可为重组质粒35s::eaf2。
62.所述重组质粒35s::eaf2的构建过程如下:
63.(1)以pmdc83载体为模板,采用引物caccgfp-f:5
′-
caccatgggtaaaggagaacttttca-3
′
和引物ha-r 6*gly:5
′-
gccaccccctccgccaccggcataatcaggcacatcg-3
′
组成的引物对进行pcr扩增,得到约800bp的gfp片段。
64.(2)以seq id no:1所示的dna分子为模板,采用引物6*gly-nfy-b4-f:5
′-
ggtggcggagggggtggcatggctgaaacggaggag-3
′
和引物mtnf-yb4-r:5
′-
tcattggtcatcatcaacaccatc-3
′
组成的引物对进行pcr扩增,得到约420bp的nf-yb4片段。
65.(3)将gfp片段和nf-yb4片段混合后作为模板,采用引物caccgfp-f和引物mtnf-yb4-r组成的引物对进行pcr扩增,得到约1200bp的gfp-nf-yb4片段。
66.(4)将gfp-nf-yb4片段和载体pentr/d-top0(invitrogen)进行bp反应,得到中间载体pentr/d-gfp-nf-yb4。
67.(5)将中间载体pentr/d-gfp-nf-yb4和pmdc83载体进行lr反应,得到重组质粒35s::eaf2。
68.上述任一所述出发植物甲可为如下c1)、c2)、c3)、c4)和c6)中的任一种;c1)双子叶植物;c2)单子叶植物;c3)豆科植物;c4)蒺藜苜蓿;c6)突变体eaf2。
69.当出发植物甲为突变体eaf2时,所述转基因植物甲具体可为实施例中的互补株35s::eaf2/eaf2-2、35s::eaf2/eaf2-6或35s::eaf2/eaf2-8。
70.本发明还保护一种培育转基因植物乙的方法,可包括如下步骤:降低出发植物乙中上述任一所述eaf2蛋白的表达量和/或活性,得到转基因植物乙;与出发植物乙相比,转基因植物乙的花青素含量提高、开花节数减少和/或开花时间提前。
71.上述方法中,所述“降低出发植物乙中上述任一所述eaf2蛋白的表达量和/或活性”可通过dna插入、rna干扰、同源重组、基因定点编辑等本领域熟知的方法,达到抑制所述eaf2蛋白的表达量或活性的目的。
72.上述方法中,所述“降低出发植物乙中上述任一所述eaf2蛋白的表达量和/或活性”具体可通过将dna插入eaf2基因的外显子上获得。
73.上述任一所述出发植物乙可为c1)、c2)、c3)、c4)和c5)中的任一种;c1)双子叶植物;c2)单子叶植物;c3)豆科植物;c4)蒺藜苜蓿;c5)蒺藜苜蓿r108。
74.所述转基因植物乙具体可为突变体eaf2。
75.实验证明,向突变体eaf2中导入重组质粒35s::eaf2,得到互补株;与突变体eaf2相比,互补株的叶片中花青素含量显著降低,开花节数增加,开花时间推迟。由此可见,eaf2蛋白及其编码基因可以调控花青素合成、开花节数和开花时间。本发明具有重要的应用价值。
附图说明
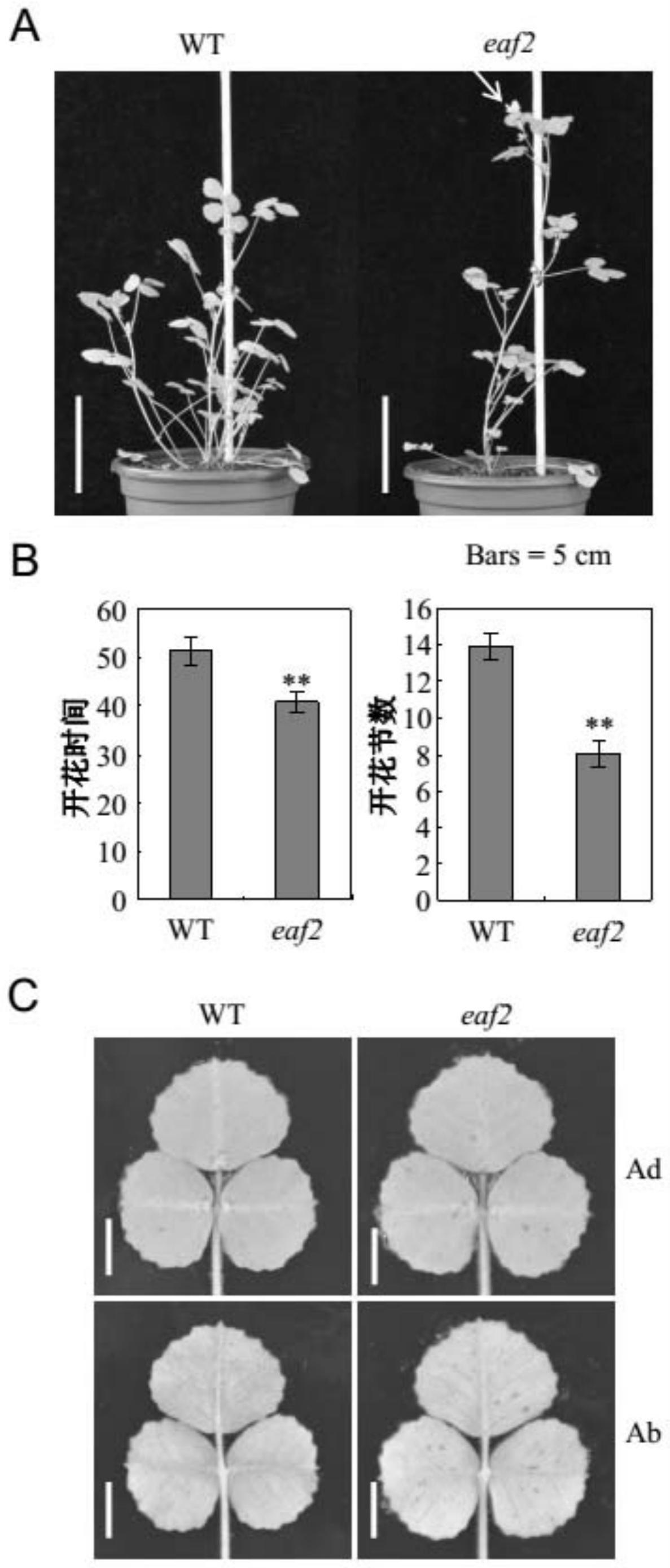
76.图1为蒺藜苜蓿r108和突变体eaf2的开花表型、开花时间和开花节数的统计结果及叶片表型。
77.图2为蒺藜苜蓿r108和突变体eaf2的花青素含量检测结果。
78.图3为互补株中eaf2基因的表达量检测结果。
79.图4为互补株的开花表型鉴定、开花节数统计结果和叶片表型鉴定。
80.图5为互补株的花青素含量检测结果。
具体实施方式
81.以下的实施例便于更好地理解本发明,但并不限定本发明。
82.下述实施例中的实验方法,如无特殊说明,均为常规方法。
83.下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂商店购买得到的。
84.以下实施例中的定量试验,均设置三次重复实验,结果取平均值。
85.蒺藜苜蓿r108由the nobel foundation(网址为:https://www.nobelprize.org/the-nobel-prize-organisation/the-nobel-foundat ion/)提供。
86.pmdc83载体和根癌农杆菌agl1均由中国农业科学院生物技术所(即申请人处)提供,公众可以从申请人处获得。
87.yep液体培养基:将蛋白胨10g、酵母提取物10g和氯化钠5g用适量蒸馏水溶解,然后用蒸馏水定容至1l,121℃高压灭菌15min。
88.愈伤诱导液体培养基:将大量元素母液100ml、微量元素母液1ml、有机元素母液1ml、铁盐母液20ml、肌醇100mg、蔗糖30g、生长素4mg和细胞分裂素0.5mg用适量蒸馏水溶解,然后用蒸馏水定容至1l,调节ph值至5.8,121℃高压灭菌15min。
89.愈伤诱导固体培养基:将大量元素母液100ml、微量元素母液1ml、有机元素母液1ml、铁盐母液20ml、肌醇100mg、蔗糖30g、生长素4mg、细胞分裂素0.5mg、头孢200mg、特美汀250mg、潮霉素b 5mg和phytagel 3.2g用适量蒸馏水溶解,然后用蒸馏水定容至1l,调节ph值至5.8,121℃高压灭菌15min。
90.分化培养基:将大量元素母液100ml、微量元素母液1ml、有机元素母液1ml、铁盐母液20ml、肌醇100mg、蔗糖20g、头孢200mg、特美汀250mg、潮霉素b 5mg和phytagel 3.2g用适量蒸馏水溶解,然后用蒸馏水定容至1l,调节ph值至5.8,121℃高压灭菌15min。
91.生根培养基:将2.215g murashige&skoog basal medium with vitamins(phytotechnology laboratories公司的产品,货号为16b0519138a)、蔗糖15g和琼脂粉7g用适量蒸馏水溶解,然后用蒸馏水定容至1l,调节ph值至5.8,121℃高压灭菌15min。
92.铁盐母液:将乙二胺四乙酸二钠37.3mg和七水合硫酸亚铁27.8mg用适量蒸馏水溶解,然后用蒸馏水定容至1l。
93.大量元素母液:将七水合硫酸镁1.85g、硝酸钾28.3g、硫酸铵4.63g、二水合氯化钙1.66g和磷酸二氢钾4g用适量蒸馏水溶解,然后用蒸馏水定容至1l。
94.微量元素母液:将一水合硫酸锰1g、硼酸500mg、七水合硫酸锌100mg、碘化钾100mg、二水合钼酸纳10mg、五水合硫酸铜20mg和六水合氯化钴10mg用适量蒸馏水溶解,然
后用蒸馏水定容至1l。
95.有机元素母液:将烟酸500mg、盐酸硫胺素500mg和盐酸吡哆醇500mg用适量蒸馏水溶解,然后用蒸馏水定容至1l。
96.突变体eaf2由蒺藜苜蓿tnt1插入突变体库(网址为:https://medicago-mutant.noble.org/mutant/index.php)提供,突变体eaf2在蒺藜苜蓿tnt1插入突变体库中的编号为nf16632。根据蒺藜苜蓿tnt1插入突变体库记载的信息,突变体eaf2为tnt1插入eaf2基因的第一个外显子(eaf2基因包含3个外显子和两个内含子)上获得的蒺藜苜蓿突变体。
97.实施例1、eaf2基因的克隆
98.一、突变体eaf2的表型分析
99.取30粒突变体eaf2种子或蒺藜苜蓿r108种子,4℃春化处理7d,然后种植至营养土中,光暗交替培养(16h光照/8h黑暗),观察表型并统计。
100.光暗交替培养40d时,部分突变体eaf2和蒺藜苜蓿r108的生长状态见图1中a(wt为蒺藜苜蓿r108,eaf2为突变体eaf2)。结果表明,光暗交替培养40d时,突变体eaf2均已开花,蒺藜苜蓿r108均未开花。
101.统计各株蒺藜苜蓿的开花时间和节数(即第一朵花完全开放时蒺藜苜蓿的生长天数和节数),然后取平均值。结果见图1中b(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,**表示具有极显著性差异,p<0.01)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2的开花时间显著提前,开花节数显著减少。
102.部分蒺藜苜蓿复叶的正面和背面表型见图1中c(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,ad为正面,ab为反面)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2复叶的正面和背面均具有更多的红色斑点。
103.二、叶片花青素含量检测
104.1、标准曲线的制备
105.(1)取矢车菊素3-o-葡萄糖苷氯化物标准品(sigma-aldrich公司的产品,货号为52976),用水稀释,得到不同浓度的标准品溶液。
106.(2)用紫外可见分光光度计分别检测530nm处步骤(1)得到的标准品溶液的吸光度值(即a530值)。
107.(3)完成步骤(2)后,分别以标准品溶液的浓度为横坐标,相应的a530值为纵坐标,绘制标准曲线。
108.标准曲线为:y=0.024x-0.0009;r2=0.9988。其中x为a530值。
109.2、花青素含量的检测
110.实验重复三次取平均值,每次重复的步骤如下:
111.(1)从生长状态基本一致的待测蒺藜苜蓿(蒺藜苜蓿r108或突变体eaf2)植株上取刚展开的成熟复叶(约50mg),称重(记为m);然后将叶片液氮冷冻20min,研磨成粉末;之后分别加入500μl盐酸/甲醇溶液(由999μl甲醇和1μl盐酸混合而成),振荡混匀,4℃黑暗旋转过夜,获得待测蒺藜苜蓿的提取液。
112.(2)取待测蒺藜苜蓿的提取液,2500g离心15min,获得待测蒺藜苜蓿的上清液。
113.(3)用紫外可见分光光度计检测530nm处待测蒺藜苜蓿的上清液的吸光度值(即
a530值)。
114.(4)将步骤(3)得到的吸光度值代入标准曲线,得到待测蒺藜苜蓿的上清液中花青素的含量;进一步得到待测蒺藜苜蓿叶片中花青素的含量。
115.检测结果见图2(wt为蒺藜苜蓿r108,eaf2为突变体eaf2;数据为平均值
±
标准差,*表示具有显著性差异,p<0.05)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2叶片中花青素含量显著增加。
116.三、eaf2基因的克隆
117.1、提取蒺藜苜蓿r108植株上取刚展开的成熟复叶的总rna,然后反转录,得到蒺藜苜蓿r108的cdna。
118.2、完成步骤1后,以蒺藜苜蓿r108的cdna为模板,采用引物mtnf-yb4-f:5
′-
atggctgaaacggaggag-3
′
和引物mtnf-yb4-r:5
′-
tcattggtcatcatcaacaccatc-3
′
组成的引物对进行扩增,得到423bp的pcr扩增产物。
119.3、将步骤2得到的pcr扩增产物进行测序。
120.测序结果表明,pcr扩增产物中含有seq id no:1所示的dna分子。seq id no:1所示的dna分子即eaf2基因,编码seq id no:2所示的eaf2蛋白。
121.实施例2、互补株的获得、表型鉴定及花青素含量检测
122.一、重组质粒35s::eaf2的构建
123.1、以pmdc83载体为模板,采用引物caccgfp-f:5
′-
caccatgggtaaaggagaacttttca-3
′
和引物ha-r 6*gly:5
′-
gccaccccctccgccaccggcataatcaggcacatcg-3
′
组成的引物对进行pcr扩增,得到约800bp的gfp片段。
124.2、以实施例1步骤三中2得到的pcr扩增产物为模板,采用引物6*gly-nfy-b4-f:5
′-
ggtggcggagggggtggcatggctgaaacggaggag-3
′
和引物mtnf-yb4-r组成的引物对进行pcr扩增,得到约420bp的nf-yb4片段。
125.3、将gfp片段和nf-yb4片段混合后作为模板,采用引物caccgfp-f和引物mtnf-yb4-r组成的引物对进行pcr扩增,得到约1200bp的gfp-nf-yb4片段。
126.4、将gfp-nf-yb4片段和载体pentr/d-top0(invitrogen)进行bp反应,得到中间载体pentr/d-gfp-nf-yb4。
127.5、将中间载体pentr/d-gfp-nf-yb4和pmdc83载体进行lr反应,得到重组质粒pmdc83-gfp-nf-yb4,记为重组质粒35s::eaf2。
128.二、重组农杆菌的获得
129.将重组质粒35s::eaf2导入根癌农杆菌agl1,得到重组农杆菌,命名为agl1/35s::eaf2。
130.三、互补株的获得
131.1、侵染液的制备
132.(1)将agl1/35s::eaf2单菌落接种于含50mg/ml利福平和50mg/ml卡那霉素的yep液体培养基,28℃、200rpm振荡培养过夜,得到培养菌液1。
133.(2)完成步骤(1)后,将500μl培养菌液1接种于5ml yep液体培养基,再加入5μl浓度为100mg/ml的乙酰丁香酮水溶液,28℃、200rpm振荡培养,得到od
600nm
值为0.8的培养菌液2。
134.(3)完成步骤(2)后,取培养菌液2,3800rpm离心15min,收集菌体。
135.(4)完成步骤(3)后,取菌体,用含100mg/l乙酰丁香酮的愈伤诱导液体培养基重悬,得到od
600nm
值为0.2的侵染液。
136.2、互补株的获得
137.(1)取生长至4周的突变体eaf2的第一片复叶,先用75%(v/v)乙醇水溶液漂洗10s,再用5%(m/v)次氯酸钠溶液浸泡5min,之后在超净台用无菌水洗涤至少5次,最后将叶片切割4-5个切口。
138.(2)完成步骤(1)后,将所述叶片小块置于侵染液,静置15min。
139.(3)完成步骤(2)后,将所述叶片小块转移至愈伤诱导固体培养基,24℃暗培养4周(每隔2周更换一次培养基),得到白色胚性愈伤组织。
140.(4)完成步骤(3)后,将白色胚性愈伤组织转移至分化培养基,24℃光暗交替培养4周(每隔2周更换一次培养基),分化出绿色胚状体。
141.(5)完成步骤(4)后,将绿色胚状体转移至生根培养基,24℃光暗交替培养(每隔2周更换一次培养基),生根长叶后移至蛭石中,再移至营养土中直至成苗,即互补株。
142.共获得10株互补株。
143.3、实时荧光定量检测互补株中eaf2基因的相对表达量
144.待测蒺藜苜蓿分别为10株互补株、蒺藜苜蓿r108和突变体eaf2。
145.(1)分别提取待测蒺藜苜蓿叶片的总rna,然后反转录,得到待测蒺藜苜蓿的cdna。
146.(2)分别以待测蒺藜苜蓿的cdna为模板,实时定量pcr检测eaf2基因的相对表达量(actin基因为内参基因)。
147.反应体系为20μl,由10μl transstart tip green qpcr supermix、0.5μl上游引物水溶液(浓度为10μm)、0.5μl下游引物水溶液(浓度为10μm)、0.5μl待测蒺藜苜蓿的cdna和8.5μl ddh2o组成。
148.transstart tip green qpcr supermix(2
×
)为全式金公司的产品。
149.反应程序为:94℃5min;94℃10s,60℃20s,72℃20s,40个循环。
150.检测eaf2基因的上游引物为5
′-
aggagcttcccaaaactatcg-3
′
,下游引物为5
′-
agattctggcactttcggag-3
′
。
151.检测actin基因的上游引物为5
’-
tcaatgtgcctgccatgtatgt-3’,下游引物为5
’-
actcacaccgtcaccagaatcc-3’。
152.部分检测结果见图3(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,35s::eaf2/eaf2-2、35s::eaf2/eaf2-6和35s::eaf2/eaf2-8为互补株;**表示具有极显著性差异,p<0.01)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2中eaf2基因的表达量显著下降,各个互补株中eaf2基因的表达量均有不同程度的增加,其中3个互补株中eaf2基因的表达量显著增加,将其依次命名为35s::eaf2/eaf2-2、35s::eaf2/eaf2-6和35s::eaf2/eaf2-8。
153.四、互补株的表型鉴定
154.取30粒待测种子(突变体eaf2种子、蒺藜苜蓿r108种子、35s::eaf2/eaf2-2种子、35s::eaf2/eaf2-6种子或35s::eaf2/eaf2-8种子),4℃春化处理7d,然后种植至营养土中,光暗交替培养(16h光照/8h黑暗),观察表型并进行统计。
155.光暗交替培养40d时,35s::eaf2/eaf2-2、突变体eaf2和蒺藜苜蓿r108的生长状态
见图4中a(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,35s::eaf2/eaf2为35s::eaf2/eaf2-2)。结果表明,光暗交替培养40d时,突变体eaf2均已开花,蒺藜苜蓿r108、35s::eaf2/eaf2-2、35s::eaf2/eaf2-6和35s::eaf2/eaf2-8均未开花。
156.统计各株蒺藜苜蓿的开花时间和开花节数(即第一朵花完全开放时蒺藜苜蓿的生长天数和节数),然后取平均值。部分统计结果见图4中b(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,**表示具有极显著性差异,p<0.01)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2的开花节数显著减少,开花时间显著提前;35s::eaf2/eaf2-2、35s::eaf2/eaf2-6、35s::eaf2/eaf2-8和蒺藜苜蓿r108的开花节数和开花时间均无显著差异。
157.部分蒺藜苜蓿复叶的正面和背面表型见图4中c(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,35s::eaf2/eaf2为35s::eaf2/eaf2-2,ad为正面,ab为反面)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2复叶的正面和背面均具有更多的红色斑点;35s::eaf2/eaf2-2、35s::eaf2/eaf2-6、35s::eaf2/eaf2-8和蒺藜苜蓿r108复叶正面和背面的红色斑点数无显著差异。
158.由此可见,互补株(35s::eaf2/eaf2-2、35s::eaf2/eaf2-6、35s::eaf2/eaf2-8)的开花节数和复叶上红色斑点数恢复至与蒺藜苜蓿r108一致。
159.五、互补株叶片中的花青素含量检测
160.按照实施例1中步骤二的方法,检测蒺藜苜蓿r108、突变体eaf2、35s::eaf2/eaf2-2、35s::eaf2/eaf2-6和35s::eaf2/eaf2-8叶片中花青素的含量。
161.部分检测结果见图5(wt为蒺藜苜蓿r108,eaf2为突变体eaf2,35s::eaf2/eaf2为35s::eaf2/eaf2-2,**表示具有极显著性差异,p<0.01)。结果表明,与蒺藜苜蓿r108相比,突变体eaf2叶片中花青素含量显著提高;35s::eaf2/eaf2-2、35s::eaf2/eaf2-6、35s::eaf2/eaf2-8和蒺藜苜蓿r108叶片中花青素含量无显著差异。
162.由此可见,互补株(35s::eaf2/eaf2-2、35s::eaf2/eaf2-6和35s::eaf2/eaf2-8)的花青素含量恢复至与蒺藜苜蓿r108一致。
163.上述结果表明,eaf2蛋白可以调控植物花青素的合成、开花节数和开花时间。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。