1.本发明涉及生物技术领域,特别是涉及一种米黑根毛霉脂肪酶突变体及提高米黑根毛霉脂肪酶活性和/或甲醇耐受性的方法,以及米黑根毛霉脂肪酶突变体在催化油脂制备生物柴油中的应用。
背景技术:
2.脂肪酶,又称三酯酰甘油酰基水解酶(ec 3.1.1.3),属于α/β水解酶超家族中的一员。由于对硝基苯磷酸二乙酯对脂肪酶有抑制作用,因此脂肪酶属于丝氨酸水解酶。脂肪酶能在油水界面水解三酰基甘油生成脂肪酸、甘油和单、二酰基甘油,并能逆转非水介质中的反应,是生物体进行脂质代谢必不可少的水解酶,也是一种在非水溶剂中进行脂质可逆反应的酶,在食品加工、污水处理、洗涤剂和生物柴油等领域具有很大的应用空间。
3.脂肪酶能催化酯化和转酯化反应,利用脂肪酶催化油脂制备生物柴油是生物柴油工业发展的一个重要方向。酶法生产生物柴油具有对原料要求低,反应条件温和,游离脂肪酸可以被脂肪酶直接酯化,且副产物甘油分离简单等优点,降低了生产工艺要求和分离成本。但是,由于游离脂肪酶在反应时容易聚集,回收和重复利用困难,甲醇对酶的失活效应等因素,限制了其在工业规模生产生物柴油中的应用。目前用于催化合成生物柴油的微生物脂肪酶有南极假丝酵母脂肪酶,皱褶假丝酵母脂肪酶和米黑根毛霉脂肪酶等,通常采用分步流加甲醇的方法降低甲醇的失活影响,反应时间36h催化效率一般为70%~100%。
4.米黑根毛霉脂肪酶(rhizomucor miehei lipase,rml)是一种具有优良的sn-1立体特异性的弱碱性脂肪酶,催化中心由ser144-asp203-his257组成。在有水的情况下,液体米黑根毛霉脂肪酶可用于生产生物柴油,但是低活性和甲醇对酶的变性效果增加了米黑根毛霉脂肪酶的生产成本,从而限制了它在生物柴油制备中的大规模应用。因此,设计一种具有高活性和高甲醇耐受性的突变型米黑根毛霉脂肪酶具有重要意义。
5.米黑根毛霉脂肪酶由前导肽和成熟肽组成,前导肽作为分子内分子伴侣介导成熟肽的折叠,从而形成具有生物学功能的蛋白。n-糖基化是毕赤酵母表达最常见的一种翻译后修饰。在蛋白质的功能域中添加一个大的聚糖可以显著改变其结构,从而改变蛋白的功能。n-聚糖的大小,亲水性或仅仅是他们存在的特定位置均可以改变糖蛋白的行为,使它们更容易溶解或更稳定,保护它们不被蛋白质水解。以上发生的一切,都源于n-聚糖对蛋白质二级结构和物理化学性质的影响。β-sheet是重要的二级结构,由两条或多条几乎完全伸展的肽链平行排列,通过链间的氢键交联而形成的。在β-sheet结构中引入n-糖链,聚糖的亲水性会改变周围环境,从而影响脂肪酶的生化性质。
技术实现要素:
6.针对上述领域中的缺陷,本发明提供一种米黑根毛霉脂肪酶突变体及提高米黑根毛霉脂肪酶活性和/或甲醇耐受性的方法,通过在米黑根毛霉脂肪酶的成熟肽β-sheet二级
结构中引入n-糖链,获得高活性高甲醇耐受性的米黑根毛霉脂肪酶突变体,得到的米黑根毛霉脂肪酶突变体能高效催化废油脂一步甲醇法转化为生物柴油。
7.为了达到上述目的,本发明采用的技术方案为:
8.一种米黑根毛霉脂肪酶突变体,所述的米黑根毛霉脂肪酶突变体的氨基酸序列上引入有n-糖基化位点,所述的n-糖基化位点上连接有n-糖链。
9.所述的米黑根毛霉脂肪酶突变体的氨基酸序列上引入有n-糖基化位点是通过以下方法获得的:将如seq id no.1所示的氨基酸序列上的第167位由p突变为n、第242位由l突变n、第267位由r突变为n或第300位由p突变为n。
10.本发明的第二个目的是提供一种提高米黑根毛霉脂肪酶活性和/或甲醇耐受性的方法,在米黑根毛霉脂肪酶成熟肽的β-sheet二级结构中引入n-糖链。所述引入n-糖链是通过将表达载体ppic9k-rml/wt上的米黑根毛霉脂肪酶相应基因进行点突变,从而实现在米黑根毛霉脂肪酶的氨基酸序列中引入n-糖基化位点,真核体系表达所述脂肪酶引入n-糖链。
11.具体步骤如下:在如seq id no.1所示的氨基酸序列上引入n-糖基化位点,所述的n-糖基化位点上连接有n-糖链。
12.优选地,所述的在如seq id no.1所示的氨基酸序列上引入n-糖基化位点是将seq id no.1所示的氨基酸序列上的第167位由p突变为n、第242位由l突变n、第267位由r突变为n或第300位由p突变为n。
13.所述的米黑根毛霉脂肪酶(即具体实施例中的米黑根毛霉脂肪酶wt)的氨基酸序列如下所示:
14.mvpikrqsnstvdslpplipsrtsapssspsttdpeapamsrngplpsdvetkygmalnatsypdsvvqamsidggiraatsqeineltyyttlsansycrtvipgatwdcihcdatedlkiiktwstliydtnamvargdsektiyivfrgsssirnwiadltfvpvsyppvsgtkvhkgfldsygevqnelvatvldqfkqypsykvavtghslggatallcalglyqreeglsssnlflytqgqprvgdpafanyvvstgipyrrtvnerdivphlppaafgflhageeywitdnspetvqvctsdletsdcsnsivpftsvldhlsyfgintglct(如seq id no.1所示)。
15.n-糖基化特征序列为asn-xaa-ser/thr(xaa是除了脯氨酸外的任意氨基酸)。根据n-糖基化特征序列,对米黑根毛霉脂肪酶成熟肽β-sheet结构中的氨基酸进行突变,建构n-糖基化位点。
16.上述方法得到四个高活性高甲醇耐受性的米黑根毛霉脂肪酶突变体在真核体系中表达,获得突变酶p167n(其氨基酸序列如seq id no.12所示,且在n-糖基化位点(
167
nvs
169
)上连接有n-糖链),l242n(其氨基酸序列如seq id no.13所示,且在n-糖基化位点(
242
nyt
244
)上连接有n-糖链),r267n(其氨基酸序列如seq id no.14所示,且在n-糖基化位点(
267
nrt
269
)上连接有n-糖链)和p300n(其氨基酸序列如seq id no.15所示,且在n-糖基化位点(
300
net
302
)上连接有n-糖链)。
17.优选地,获得的米黑根毛霉脂肪酶突变体p167n,l242n,r267n和p300n酶活分别是1846.12
±
58.56u/mg,974.86
±
18.88u/mg,766.25
±
31.04u/mg和667.59
±
36.70u/mg。与改造前的米黑根毛霉脂肪酶wt相比,活性分别提高了33.90,17.90,14.07和12.26倍。
18.优选地,当在60%的甲醇溶液中孵育1h后,获得的突变酶p167n,l242n,r267n和p300n的残余活力由改造前的残余活性19.10%提高到60%以上,其中突变酶p167n,p300n,
r267n的残余活力均高于85%。
19.本发明的第三个目的是提供了上述的米黑根毛霉脂肪酶突变体在催化油脂制备生物柴油中的应用。
20.优选地,以废油脂为原料,米黑根毛霉脂肪酶突变体为催化剂,一次性加入甲醇(醇:油摩尔比=3:1)反应24h后,米黑根毛霉脂肪酶突变体p167n,l242n,r267n和p300n的生物柴油转化率由改造前的8.44%分别提高到72.68%,64.07%,68.42%和63.99%。
21.与现有技术相比,本发明提供的米黑根毛霉脂肪酶突变体,具有如下优势:
22.(1)米黑根毛霉脂肪酶突变体为高活性高甲醇耐受性的米黑根毛霉脂肪酶突变体;
23.(2)突变体可以高效利用废油脂一步甲醇法制备生物柴油。
附图说明
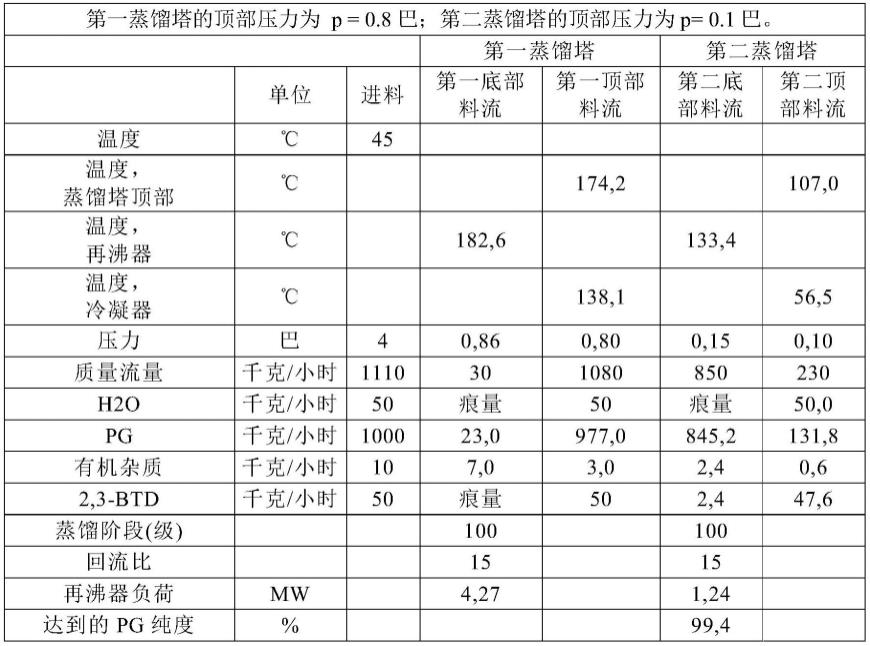
24.图1是质粒ppic9k-rml/wt的质粒图谱;
25.图2是米黑根毛霉脂肪酶突变体的sds-page电泳图;
26.图3是米黑根毛霉脂肪酶突变体的酶活分析图;
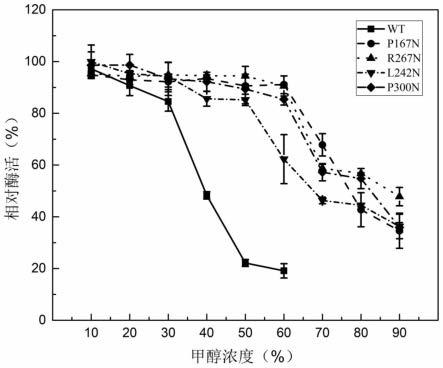
27.图4是米黑根毛霉脂肪酶突变体的甲醇耐受性图;
28.图5是米黑根毛霉脂肪酶突变体在50%甲醇浓度下的稳定性图;
29.图6是米黑根毛霉脂肪酶突变体的fame转化图。
具体实施方式
30.下面结合具体实施例对本发明作进一步解释,但应当注意的是,以下实施例仅用以解释本发明,而不能用来限制本发明,所有与本发明相同或相近的技术方案均在本发明的保护范围之内。若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段,所用原料为市售商品。
31.实施例1:米黑根毛霉脂肪酶突变体p167n
32.1.米黑根毛霉脂肪酶突变体p167n表达菌株的构建
33.(1)以一种米黑根毛霉脂肪酶wt的质粒ppic9k-rml/wt为模板,该模板由本发明人先前构建,ppic9k-rml/wt的质粒图谱如图1所示,其核苷酸序列如seq id no.16所示。
34.(2)在米黑根毛霉脂肪酶wt成熟肽β-sheet二级结构的
167
pvs
169
序列上进行突变,突变为
167
nvs
169
,引入新的n-糖基化位点。
35.(3)以质粒ppic9k-rml/wt为模板,以突变引物对n167f/n167r作为扩增引物,根据全式金定点突变试剂盒说明书进行定点突变,突变引物序列见表1。在扩增得到的pcr产物中加入1μl dpni酶,于37℃恒温培养箱反应1h,反应结束后取10μl酶切后的pcr产物加入到100μl刚融化的dmt感受态细胞,冰上孵育30min,然后42℃水浴45sec,立即置于冰上孵育2-3min,随后加入250μl的soc无抗培养基于37℃,200rpm复苏1h后涂布于lb固体平板(1
‰
amp抗性),37℃倒置过夜培养;用20μl无抗lb培养基稀释挑选的单克隆,取1μl菌液进行菌液pcr反应。
36.菌液pcr反应体系:
[0037][0038]
菌液pcr反应程序:
[0039][0040]
菌液pcr引物是通用引物α-factor(5
’‑
tactattgccagcattgctgc-3’,如seq id no.2所示)和3’aox(5
’‑
ggcaaatggcattctgacat-3’,如seq id no.3所示)。菌液pcr产物用0.8%的琼脂糖凝胶电泳检测,对有目的条带的单克隆进一步送上海生工测序。
[0041]
表1突变引物序列
[0042][0043]
(4)对测序成功的米黑根毛霉脂肪酶突变体p167n(即含有突变质粒ppic9k-rml/p167n的菌株)于lb液体培养基(1
‰
amp抗性)过夜培养,用质粒小提试剂盒提取质粒,然后用胶回收试剂盒对1%的琼脂糖凝胶进行胶回收,胶回收产物于0.8%的琼脂糖凝胶电泳验证,得到突变质粒ppic9k-rml/p167n,用限制性内切酶saci对验证正确的突变质粒ppic9k-rml/p167n酶切线性化,然后利用电转化方法将线性化后的突变质粒转化至毕赤酵母gs115感受态细胞,得到含有突变质粒ppic9k-rml/p167n的毕赤酵母gs115。
[0044]
(5)毕赤酵母gs115感受态细胞的制备:
[0045]
挑取ypd平板上保藏的毕赤酵母gs115于5ml ypd液体培养基中,30℃,200rpm活化22h至od600=0.8-1.0;取1%的接种量于50ml ypd培养基中30℃,200rpm培养16h,于4℃冰箱预冷30min左右;4℃,4000rpm离心5min,弃上清;用50ml冰预冷的灭菌的ddh2o重悬,4℃,4000rpm离心5min,弃上清;用25ml冰预冷的灭菌的ddh2o重悬,4℃,4000rpm离心5min,弃上清;用2.5ml冰预冷的1m山梨醇溶液重悬,4℃,4000rpm离心5min,弃上清;用200-300μl冰预冷的1m山梨醇溶液重悬,每100μl分装于1.5ml的离心管,存储于-80℃备用。
[0046]
(6)重组质粒电转化毕赤酵母gs115感受态细胞:
[0047]
取10μl用限制性内切酶saci酶切得到的线性化突变质粒ppic9k-rml/p167n于100μl刚融化的毕赤酵母gs115感受态细胞中,轻轻混匀,转移到冰预冷的电转化杯,冰上静止5min;然后在电转仪进行电转,条件是:1500v,200ω,0.2cm,电转完成后立即添加1ml冰预冷的1m山梨醇溶液,然后于30℃恒温培养箱孵育2-3h,取300μl涂布于md固体平板,30℃倒置培养2-3天。
[0048]
(7)挑取md平板上的转化子于三丁酸甘油酯平板筛选阳性转化子(即为含有突变质粒ppic9k-rml/p167n的毕赤酵母gs115),筛选得到的阳性转化子进行保菌。
[0049]
2.米黑根毛霉脂肪酶突变体p167n的表达
[0050]
挑取步骤1(7)中筛选得到的阳性转化子于5ml ypd液体培养基中,30℃,200rpm过夜活化22h至od600=0.8左右,取1%接种于50ml bmgy培养基中30℃,200rpm培养40h,4℃,4000rpm离心5min,弃上清,将菌体转接于50ml bmmy培养基中30℃,200rpm培养,每隔24h流加甲醇至终浓度为0.5%,总共诱导120h;最后将诱导得到的菌液于4℃,8000rpm离心10min,上清即为表达的粗酶液。
[0051]
3.米黑根毛霉脂肪酶突变体p167n的纯化
[0052]
将步骤2的粗酶液通过10kda膜(millipore,usa)超滤浓缩,4℃,4000rpm离心30min。然后将浓缩液加载到经binding buffer(20mm磷酸缓冲液,20-40mm咪唑,ph 7.4)平衡的histrap
tm hp柱上;目的蛋白用elution buffer(20mm磷酸缓冲液,400mm咪唑,ph 7.4)梯度洗脱,将洗脱的组分合并后继续浓缩,浓缩的目的蛋白(即突变酶p167n,其氨基酸序列如seq id no.12所示)储存于-20℃备用。
[0053]
4.米黑根毛霉脂肪酶突变体p167n的糖基化验证
[0054]
(1)将纯化后的突变酶p167n(即米黑根毛霉脂肪酶突变体p167n,由米黑根毛霉脂肪酶wt的如seq id no.1所示氨基酸序列的第167位由p突变为n,其氨基酸序列如seq id no.12所示)用n-糖苷酶pngase f酶切,然后跑sds-page电泳,检测目的条带的变化。
[0055]
(2)pngase f酶切方法:
[0056]
将步骤3储存的10μg突变酶p167n和2μl 10
×
glycoprotein变性缓冲液混合,添加ddh2o至反应体系为10μl,100℃沸水煮10min。变性后的突变酶p167n,添加2μl 10
×
g7 buffer,2μl 10%np40,1μl的pngase f酶,加5μl ddh2o至反应总体系为20μl,37℃酶切1小时。sds-page检测上清的糖基化情况。
[0057]
(3)酶切后的突变酶p167n样品用15%(w/v)丙烯酰胺凝胶进行sds-page分析。酶切后的突变酶p167n样品和6
×
protein buffer的体积比为6:1,混合后于100℃沸水中煮5min,12000rpm离心5min后取20μl上样,120v跑1h左右。蛋白胶在考马斯亮蓝g-250染色2-3h,染色结束后用双蒸馏水清洗,然后在脱色液中过夜脱色,脱色液需要定期更换。脱色后,
将凝胶置于fusion fx(vilber lourmat)成像仪中观察,使用fusioncapt(advance)拍照。
[0058]
(4)sds-page电泳结果如图2所示,米黑根毛霉脂肪酶突变体p167n在没有经糖苷酶pngase f酶切处理前,分子量大小约为55kda左右,远高于米黑根毛霉脂肪酶wt的分子量(~40kda)。将突变酶p167n酶切处理后,有两条带,一条与米黑根毛霉脂肪酶wt的分子量大小一致,约为40kda,一条与pngase f酶的大小一致,约为35kda,说明构建的米黑根毛霉脂肪酶突变体p167n发生了n-糖基化修饰。
[0059]
5.米黑根毛霉脂肪酶突变体p167n的酶活分析
[0060]
为研究米黑根毛霉脂肪酶成熟肽上β-sheet二级结构中的n-糖基化修饰对脂肪酶活性的影响,采用对硝基棕榈酸苯酯(pnpp)法对获得的突变酶p167n进行酶活力的测定,以米黑根毛霉脂肪酶wt(无糖基化修饰的酶)为对照,结果如图3所示。在米黑根毛霉脂肪酶wt成熟肽的β-sheet结构中引入n-糖链后,突变酶p167n的酶活是1846.12
±
58.56u/mg。与改造前的米黑根毛霉脂肪酶wt相比,活性提高了33.91倍。这说明在米黑根毛霉脂肪酶成熟肽的β-sheet结构中引入n-糖链后,有利于提高rml的水解活性。
[0061]
6.米黑根毛霉脂肪酶突变体p167n的甲醇耐受性分析
[0062]
米黑根毛霉脂肪酶因其良好的甲醇耐受性,广泛应用于生物柴油的制备,因此提高活性和甲醇耐受性能扩大其在生物柴油行业的应用。实验考察了突变酶p167n在10-90%甲醇浓度中的稳定性,结果如图4和5所示。当在60%的甲醇溶液中孵育1h后,获得的突变酶p167n的残余活力高达91.12%,远高于改造前的米黑根毛霉脂肪酶wt的19.10%。当甲醇浓度上升至90%时,突变酶p167n的残余酶活力高达34.6%。继续将突变酶p167n在50%的甲醇溶液中孵育一段时间,结果如图5所示。孵育2.5h后,突变酶p167n的残余酶活高达92.62%,突变酶p167n的活性几乎没有损失,远高于米黑根毛霉脂肪酶wt(21.30%),结果说明在米黑根毛霉脂肪酶成熟肽的β-sheet结构中引入n-糖链后,大大提高了酶对甲醇的耐受性。
[0063]
7.米黑根毛霉脂肪酶突变体p167n的生物柴油制备
[0064]
如图6所示,以废弃的大豆油(5g)为原料,突变酶p167n为催化剂(添加量为5ml),一次性加入甲醇(醇油摩尔比3:1)反应24h后,突变酶p167n的生物柴油(fame)转化率得到明显提高,是72.68%,远高于米黑根毛霉脂肪酶wt的生物柴油转化率8.45%;继续反应至48h后,突变酶p167n的转化率是79.06%,远高于米黑根毛霉脂肪酶wt的fame转化率,这说明在米黑根毛霉脂肪酶成熟肽的β-sheet结构中引入n-糖链后,可以高效的将废弃的食用油转化为生物柴油,这不仅提高了反应效率,缩短了反应时间,还使分步流加甲醇工艺简化为一步甲醇流加工艺。
[0065]
实施例2:米黑根毛霉脂肪酶突变体l242n
[0066]
1.米黑根毛霉脂肪酶突变体l242n表达菌株的构建
[0067]
构建方法同实施例1,突变酶l242n(即米黑根毛霉脂肪酶突变体l242n)在米黑根毛霉脂肪酶wt成熟肽β-sheet二级结构的
240
lflytq 245
序列上进行突变,突变为序列
240
lfnytq 245
,引入新的n-糖基化位点。突变引物为表1所示的引物对n242f/n242r,构建得到的突变质粒为ppic9k-rml/l242n。按照实施例1步骤(4)-步骤(7)获得阳性转化子即含有突变质粒ppic9k-rml/l242n的毕赤酵母gs115。
[0068]
2.米黑根毛霉脂肪酶突变体l242n的表达,方法同实施例1。
[0069]
3.米黑根毛霉脂肪酶突变体l242n(即突变酶l242n,由米黑根毛霉脂肪酶wt的如seq id no.1所示氨基酸序列的第242位由l突变为n,其氨基酸序列如seq id no.13所示)的纯化,方法同实施例1。
[0070]
4.米黑根毛霉脂肪酶突变体l242n的糖基化验证,方法同实施例1。
[0071]
sds-page电泳结果如图2所示,米黑根毛霉脂肪酶突变体l242n在没有经糖苷酶pngase f酶切处理前,分子量大小约为55kda左右,远高于米黑根毛霉脂肪酶wt的分子量(~40kda)。将突变酶l242n酶切处理后,有两条带,一条与米黑根毛霉脂肪酶wt的分子量大小一致,约为40kda,一条与pngase f酶的大小一致,约为35kda,说明构建的米黑根毛霉脂肪酶突变体l242n发生了n-糖基化修饰。
[0072]
5.米黑根毛霉脂肪酶突变体l242n的酶活分析,方法同实施例1。
[0073]
如图3所示,突变酶l242n的酶活是974.86
±
18.88u/mg。与改造前的米黑根毛霉脂肪酶wt相比,活性提高了17.90倍。
[0074]
6.米黑根毛霉脂肪酶突变体l242n的甲醇耐受性分析,方法同实施例1。
[0075]
如图4和5所示,当在60%的甲醇溶液中孵育1h后,获得的突变酶l242n的残余活力高达62.27%,远高于对照组wt的19.10%。当甲醇浓度上升至90%时,突变酶l242n的残余酶活力高达36.26%。继续将突变酶l242n在50%的甲醇溶液中孵育一段时间,结果如图5所示。孵育2.5h后,突变酶l242n的残余酶活高达67.48%,远高于米黑根毛霉脂肪酶wt的残余活性21.30%。
[0076]
7.米黑根毛霉脂肪酶突变体l242n的生物柴油制备
[0077]
如图6所示,以废弃的大豆油(5g)为原料,突变酶l242n为催化剂(添加量为5ml),一次性加入甲醇(醇油摩尔比3:1)反应24h后,突变酶l242n的生物柴油(fame)转化率得到明显提高,是64.07%,远高于米黑根毛霉脂肪酶wt的生物柴油转化率8.45%;继续反应至48h后,突变酶l242n的转化率是67.74%,仍然远高于改造前米黑根毛霉脂肪酶wt的fame转化率。
[0078]
实施例3:米黑根毛霉脂肪酶突变体r267n
[0079]
1.米黑根毛霉脂肪酶突变体r267n表达菌株的构建
[0080]
构建方法同实施例1。突变酶r267n(即米黑根毛霉脂肪酶突变体r267n)在米黑根毛霉脂肪酶wt成熟肽β-sheet二级结构的
266
yrrtvn 271
序列上进行突变,突变为
266
ynrtvn
271
,引入新的n-糖基化位点。突变引物为表1所示的引物对n267f/n267r,构建得到的突变质粒为ppic9k-rml/r267n。按照实施例1步骤(4)-步骤(7)获得阳性转化子即含有突变质粒ppic9k-rml/r267n的毕赤酵母gs115。
[0081]
2.米黑根毛霉脂肪酶突变体r267n的表达,方法同实施例1。
[0082]
3.米黑根毛霉脂肪酶突变体r267n(即突变酶r267n,由米黑根毛霉脂肪酶wt的如seq id no.1所示氨基酸序列的第267位由r突变为n,其氨基酸序列如seq id no.14所示)的纯化,方法同实施例1。
[0083]
4.米黑根毛霉脂肪酶突变体r267n的糖基化验证,方法同实施例1。
[0084]
sds-page电泳结果如图2所示,米黑根毛霉脂肪酶突变体r267n在没有经糖苷酶pngase f酶切处理前,分子量大小约为55kda左右,远高于米黑根毛霉脂肪酶wt的分子量(~40kda)。将突变酶r267n酶切处理后,有两条带,一条与米黑根毛霉脂肪酶wt的分子量大
小一致,约为40kda,一条与pngase f酶的大小一致,约为35kda,说明构建的米黑根毛霉脂肪酶突变体r267n发生了n-糖基化修饰。
[0085]
5.米黑根毛霉脂肪酶突变体r267n的酶活分析,方法同实施例1。
[0086]
如图3所示,在米黑根毛霉脂肪酶wt成熟肽的β-sheet结构中引入n-糖链后,突变酶r267n的酶活是766.25
±
31.05u/mg。与米黑根毛霉脂肪酶wt相比,活性提高了14.07倍。
[0087]
6.米黑根毛霉脂肪酶突变体r267n的甲醇耐受性分析,方法同实施例1。
[0088]
如图4和5所示,当在60%的甲醇溶液中孵育1h后,获得的突变酶r267n的残余活力高达90.53%,远高于对照组wt的19.10%。当甲醇浓度上升至90%时,突变酶r267n的残余酶活力高达47.79%。继续将突变酶r267n在50%的甲醇溶液中孵育一段时间,结果如图5所示。孵育2.5h后,突变酶r267n的残余酶活高达84.94%,远高于wt的残余酶活21.30%,继续孵育至8h,突变酶r267n的残余活力高达63.88%。
[0089]
7.米黑根毛霉脂肪酶突变体r267n的生物柴油制备
[0090]
如图6所示,以废弃的大豆油(5g)为原料,突变酶r267n为催化剂(添加量为5ml),一次性加入甲醇(醇油摩尔比3:1)反应24h后,突变酶r267n的生物柴油(fame)转化率得到明显提高,是64.42%,远高于米黑根毛霉脂肪酶wt的生物柴油转化率8.45%;继续反应至48h后,突变酶r267n的转化率是81.70%,仍然远高于米黑根毛霉脂肪酶wt的fame转化率。
[0091]
实施例4:米黑根毛霉脂肪酶突变体p300n
[0092]
1.米黑根毛霉脂肪酶突变体p300n表达菌株的构建
[0093]
构建方法同实施例1。突变酶p300n(即米黑根毛霉脂肪酶突变体p300n)在米黑根毛霉脂肪酶wt成熟肽β-sheet二级结构的
300
petvqvc 306
序列上进行突变,突变为
300
netvqvc 306
,引入新的n-糖基化位点。突变引物为表1所示的引物对n300f/n300r,构建得到的突变质粒为ppic9k-rml/p300n。按照实施例1步骤(4)-步骤(7)获得阳性转化子即含有突变质粒ppic9k-rml/p300n的毕赤酵母gs115。
[0094]
2.米黑根毛霉脂肪酶突变体p300n的表达,方法同实施例1。
[0095]
3.米黑根毛霉脂肪酶突变体p300n(即突变酶p300n,由米黑根毛霉脂肪酶wt的如seq id no.1所示氨基酸序列的第300位由p突变为n,其氨基酸序列如seq id no.15所示)的纯化,方法同实施例1。
[0096]
4.米黑根毛霉脂肪酶突变体p300n的糖基化验证,方法同实施例1。
[0097]
sds-page电泳结果如图2所示,米黑根毛霉脂肪酶突变体p300n在没有经糖苷酶pngase f酶切处理前,分子量大小约为55kda左右,远高于米黑根毛霉脂肪酶wt的分子量(~40kda)。将突变酶p300n酶切处理后,有两条带,一条与米黑根毛霉脂肪酶wt的分子量大小一致,约为40kda,一条与pngase f酶的大小一致,约为35kda,说明构建的米黑根毛霉脂肪酶突变体p300n发生了n-糖基化修饰。
[0098]
5.米黑根毛霉脂肪酶突变体p300n的酶活分析,方法同实施例1。
[0099]
以米黑根毛霉脂肪酶wt(无糖基化修饰的酶)为对照,测定了突变酶p300n的酶活,结果如图3所示。在米黑根毛霉脂肪酶wt成熟肽的β-sheet结构中引入n-糖链后,突变酶p300n的酶活是667.59
±
36.70u/mg。与米黑根毛霉脂肪酶wt相比,活性提高了12.26倍。
[0100]
6.米黑根毛霉脂肪酶突变体p300n的甲醇耐受性分析,方法同实施例1。
[0101]
如图4和5所示,当在60%的甲醇溶液中孵育1h后,获得的突变酶p300n的残余活力
高达85.38%,远高于对照组wt的19.10%。当甲醇浓度上升至90%时,突变酶p300n的残余酶活力高达35.90%。继续将突变酶p300n在50%的甲醇溶液中孵育一段时间,结果如图5所示。孵育2.5h后,突变酶p300n的残余酶活高达79.81%,远高于米黑根毛霉脂肪酶wt的残余酶活力21.30%,结果说明在米黑根毛霉脂肪酶成熟肽的β-sheet结构中引入n-糖链后,大大提高了酶对甲醇的耐受性。
[0102]
7.米黑根毛霉脂肪酶突变体p300n的生物柴油制备
[0103]
如图6所示,以废弃的大豆油(5g)为原料,突变酶p300n为催化剂(添加量为5ml),一次性加入甲醇(醇油摩尔比3:1)反应24h后,突变酶p300n的生物柴油(fame)转化率得到明显提高,是63.99%,远高于米黑根毛霉脂肪酶wt的生物柴油转化率8.45%;继续反应至48h后,突变酶p300n的转化率是83.62%,远高于米黑根毛霉脂肪酶wt的fame转化率。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。