1.本发明涉及温度敏感性雄性不育植物的制造方法、和由此获得的该植物。另外,本发明涉及使用所述植物制造杂种种子的制造方法、和由此获得的该种子。
背景技术:
2.杂交种(杂种)中,可获得与亲本品种相比生长发育旺盛、收量、品质高这样的杂种优势(杂交优势)。进而,在能够利用病害抵抗性基因群等来积累有用的性状等方面,杂种育种的优点大。在蔬菜、花卉、玉米等中,一般被利用的几乎全部是杂种品种。另外,中国、美国的稻类作物中杂种品种也广泛普及。
3.但是,杂种品种由于是限于一代的品种,因此每次种子生产都需要大量的交配操作,现状是为了交配所花费的劳动力和时间、成本成了瓶颈。一般来说作物的品种间的交配需要将一个品种制成缺损花粉的状态,使其附着另一品种的花粉。现状是为了使花粉缺损,基本采取以下的任一方法:手工操作摘去雄蕊(去雄操作);或者事先培育出特殊的雄性不育系统。例如,在番茄、茄子等中没有雄性不育系统,因而在杂种种子生产中进行通过手工操作的去雄交配。如果能够利用雄性不育系统,则可以期待削减去雄操作。
4.利用了雄性不育系统的杂种种子的主要生产法被称为三系法,需要作为父本的育性恢复系统、成为母本的雄性不育系统、和其维持系统3个系统(参照图1)。对所使用的优良品种,雄性不育、和用于使其具有抵消雄性不育的育性恢复的性状的特别的育种是必要的,不仅繁琐而且也花时间。不仅如此,能够利用的细胞质雄性不育因子和与其配合的育性恢复因子仅有有限的组合,因为需要通过交配育种导入这些因子,所以花费劳动力和时间。进而,成为利用广泛的遗传资源的妨碍。
5.于是,为了仅用2个系统制作杂种种子(为了进行二系法),主要研究了如图2所示那样的条件雄性不育系统的利用、转基因系统的利用等(专利文献1~3、非专利文献1~3)。
6.例如,仅在特定的环境条件下变成雄性不育的系统中,有温度敏感性雄性不育。其能够通过温度条件来控制形成正常的花粉而结实的状态,还是形成由于花粉形成不全而变成雄性不育的状态。因此,仅通过温度条件就能够简便地切换自交和异交,具有系统的维持与杂交种的育成两方在一个系统中进行这样的大优点。进而,如果能够高准确度地控制花粉形成,则扩展到可能实现利用蜂等传粉昆虫的交配等,可期待采种的进一步效率化。因此,要求能够在广泛的作物种中建立温度敏感性雄性不育系统,简便且短时期地进行杂种育种的方法。
7.作为这样的温度敏感性雄性不育系统,在稻中报告了图3所示的、以
“レイメイ”
作为原品种的“水稻中间母本农12号”(pl12),并被利用(专利文献1、非专利文献1)。然而,参与该突变体的表型的责任基因尚未被鉴定。因此,在
“レイメイ”
以外的品种、稻以外的植物种中制作该温度敏感性雄性不育系统、进而进行二系法制作杂种种子是困难的。
8.现有技术文献
9.专利文献
10.专利文献1:日本特开平02-207722号公报
11.专利文献2:日本特表2017-535294号公报
12.专利文献3:日本特表2018-535672号公报
13.非专利文献
14.非专利文献1:yamaguchi et al.1997,breeding science,vol.47,371-373
15.非专利文献2:chueasiri et al.2014,plos one,vol.9(9),e106386
16.非专利文献3:pitnjam et al.2008,planta,vol.228,813-822
技术实现要素:
17.发明要解决的课题
18.本发明是鉴于上述课题而做出的,目的是鉴定“pl12”中的与温度敏感性雄性不育性状有关的责任基因,提供以该基因作为靶标的条件型雄性不育植物的制造方法。
19.用于解决课题的手段
20.本发明者们为了实现上述目的,首先进行了精细定位。但是,不能比第7染色体的54cm附近的约1.8mb进一步缩窄范围。于是,进行了原品种
“レイメイ”
与“pl12”的全基因组序列分析,将两者进行了比较。其结果判明了,在“pl12”的所述约1.8mb的区域中有约150kb的缺失。
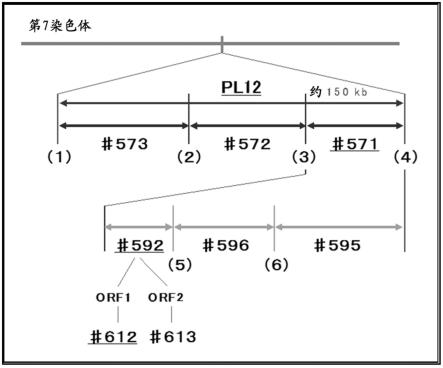
21.接着,如图4所示,将约150kb的缺失区域大致3等分,通过基因组编辑法制成使这3个区域分别缺失的系统。其结果得知其中1个显示与“pl12”同样的表型,候选区域被缩小。进一步同样地,将所述候选区域分3份,制成使3个区域分别缺失的系统,成功地将存在责任突变的范围压缩到约10kb。
22.然后,根据数据库的信息,预想这10kb的区域中存在2个基因(图4所示的orf1和orf2),因而通过基因组编辑分别破坏了各个基因功能。其结果终于表明,通过orf1的缺失,从而发挥温度敏感性雄性不育的性状,因而该基因是“pl12”中的与温度敏感性雄性不育性状有关的责任基因(以下也称为“温度敏感性雄性不育基因”)。
23.进而,通过基因组编辑抑制了该基因的功能,结果发现,在作为双子叶植物的拟南芥和番茄中也得到同样的温度敏感性雄性不育的性状,从而完成了本发明。因此,本发明是提供以下内容的发明。
24.即,本发明涉及温度敏感性雄性不育植物的制造方法、和由此得到的该植物。另外,本发明提供利用所述植物的杂种种子的制造方法、和由此得到的该种子,更具体地提供以下<1>~<4>。
25.<1>温度敏感性雄性不育植物的制造方法,
26.该方法包括人工地抑制植物的、选自由下述(a)~(d)组成的组中的至少一种基因的功能的工序,
27.(a)编码由序列号1~22中的任一项所记载的氨基酸序列组成的蛋白质的基因
28.(b)编码由在序列号1~22中的任一项所记载的氨基酸序列中替换、缺失、添加和/或插入了1个或多个氨基酸的氨基酸序列组成的蛋白质的基因
29.(c)编码与序列号1~22中的任一项所记载的氨基酸序列具有60%以上的同源性的氨基酸序列的基因
30.(d)包含与由编码序列号1~22中的任一项所记载的氨基酸序列的核苷酸序列组成的dna在严格条件下杂交的dna的基因。
31.<2>温度敏感性雄性不育植物,其中,选自由下述(a)~(d)组成的组中的至少一种基因的功能被人工地抑制,
32.(a)编码由序列号1~22中的任一项所记载的氨基酸序列组成的蛋白质的基因
33.(b)编码由在序列号1~22中的任一项所记载的氨基酸序列中替换、缺失、添加和/或插入了1个或多个氨基酸的氨基酸序列组成的蛋白质的基因
34.(c)编码与序列号1~22中的任一项所记载的氨基酸序列具有60%以上的同源性的氨基酸序列的基因
35.(d)包含与由编码序列号1~22中的任一项所记载的氨基酸序列的核苷酸序列组成的dna在严格条件杂交的dna的基因。
36.<3>杂种种子的制造方法,该方法包括
37.使在限制温度下栽培、制成了雄性不育的<2>所述的温度敏感性雄性不育植物,与任意的植物进行异花传粉的工序,和
38.由所述温度敏感性雄性不育植物回收种子的工序。
39.<4>杂种种子,其母本是以在限制温度下栽培、制成了雄性不育的<2>所述的温度敏感性雄性不育植物,其父本是任意的植物。
40.发明的效果
41.根据本发明,能够制造温度敏感性不育植物。特别是除了抑制本发明的温度敏感性雄性不育基因的功能以外,不需要特别的育种,就能够制造温度敏感性不育植物。
42.另外,如图6和7所示,由本发明的温度敏感性雄性不育基因所编码的蛋白质的氨基酸序列高度保守。由于该基因在作为最原始的被子植物的无油樟目(amborellales)中也存在,因而可知是至少在被子植物中普遍存在的基因。因此,认为原则上可以在所有农作物中利用本基因。这样,在本发明的方法中,所应用的系统不受限定,因而能够在各种植物种的各种品种/系统中,通过抑制该基因的功能而制成雄性不育系统。
43.并且,通过以所得的温度敏感性雄性不育植物作为母本,与作为父本的其他系统一起在限制温度下栽培,从而能够简便地获得这2个系统杂交而成的杂种种子。
附图说明
44.图1是显示作为杂种种子的制造方法使用的三系法的概要的图。
45.图2是显示作为杂种种子的制造方法使用的二系法的概要的图。“α”表示通过温度等环境条件而形成雄性不育的系统,“β”表示任意的品种/系统。
46.图3是显示“pl12”的表型的照片。显示容许温度(28℃)和限制温度(33℃)下的开花期的颖花的立体图像(上)和花药的染色图像(下)。
47.图4是显示“pl12”的责任因子鉴定的情况的概要图。首先,在(1)~(6)的部位设计了指导rna。利用以温度敏感性雄性不育系统“pl12”的基因组的缺失区域作为起始,通过crispr/cas9使2处双链切割之间缺失的方法,制成了具有部分缺失的系统。将其表型与“pl12”同样的候选区域依次收窄。因为已知缺失(3)至(4)之间的#571的系统具有温度敏感性雄性不育的性状,所以将其内部细分化而进行分析,将候选压缩至(3)至(5)之间。该区域
内有orf1和orf2的2个开放式阅读框,导入了分别使各个orf缺损这样的突变。此外,图中,系统名带有下划线的是具有温度敏感性雄性不育的性状的系统。
48.图5是显示图4所示的、具有部分缺失的各系统在容许温度(28℃)和限制温度(33℃)下的开花期的颖花的立体图像(各代表例)的照片。图中,带有下划线的#571、#592、#612的系统显示温度敏感性雄性不育的性状。
49.图6显示将由本发明的温度敏感性雄性不育基因所编码的氨基酸序列在各植物种间进行比较而得的排比。以高等植物的数据库的信息为基础,预测orf而翻译成氨基酸序列进行比较。使用qiagen社clc软件进行了序列分析。
50.图7显示将由本发明的温度敏感性雄性不育基因所编码的氨基酸序列在稻科作物间进行比较而得的排比。以稻科作物的数据库的信息为基础,预测orf而翻译成氨基酸序列进行比较。使用qiagen社clc软件进行了序列分析。
51.图8是显示稻的基因组编辑系统等的表型(代表例)的照片。“pl12”在28℃正常形成花粉而形成黄色且充实的花药,与此相对,在33℃花粉不发达,花药的形态、颜色异常。标准品种“日本晴”无论温度如何都正常形成花粉。通过基因组编辑而在“日本晴”使该基因缺损了的系统与“pl12”同样地显示温度敏感性雄性不育的表型。
52.图9是显示稻的基因组编辑系统等的外观(代表例)的照片。将通过稻品种“日本晴”的基因组编辑而基因功能缺失了的系统在28℃(左)和33℃(右)进行了栽培。在28℃正常结实而穗垂下来,与此相对,在33℃由于不结实而形成穗立着的状态。
53.图10是显示“日本晴”和“北陆193号”各自的基因组编辑系统等的外观(代表例)的照片。在“北陆193号”来源的基因组编辑系统中,也与“日本晴”来源的基因组编辑系统同样地,在通常条件下形成花粉而花药发达,在高温条件下由于花粉形成不全而变成雄性不育。
54.图11是显示由(1)
“オオナリ”
、(2)“北陆193号”来源的基因组编辑系统、(3)对高温下栽培的“北陆193号”来源的基因组编辑系统施与
“オオナリ”
的花粉而得的个体分别纯化基因组dna,分析本发明的温度敏感性雄性不育基因的突变插入部位的碱基序列的结果的图。如下所示,在(3)中,形成了(1)与(2)的基因组序列的杂合子,显示通过(1)与(2)的交配获得了杂种。(1)acgctcttgcggaaga(序列号34)野生型、(2)acgctctttgcggaaga(序列号35)单碱基t插入、(3)acgctctt(t/g)(g/c)(c/g)g(g/a)a(a/g)(g/a)(序列号36)单碱基t插入/野生型。
55.图12是显示拟南芥的基因组编辑系统的外观(代表例)的照片。将通过拟南芥的基因组编辑而基因功能缺失了的系统在21℃(上)和27℃(下)进行了栽培。在21℃正常结实且荚果的长度也与野生型相比无变化,与此相对,在27℃不结实且荚果不伸长。
56.图13是显示将番茄的基因组编辑系统与野生型一起在高温条件(昼30℃/夜28℃)栽培,判定所得的果实内是否形成种子的结果的照片。图中,上半部分是植物个体形成了果实的样子的照片,下半部分是将果实切成一半的状态的照片。将有种子的果实用虚线包围起来而显示,将没有种子的果实用实线包围起来而显示。图中“tms2”是指本发明的温度敏感性雄性不育基因(参照序列号43和44)。认为在具有6碱基缺失突变的系统中发生该部位的2氨基酸的缺失(参照序列号45和46),此外形成具有功能的蛋白质,在种子形成方面与野生型未见差异。相对地,具有在推定通过移码而蛋白质功能缺损的5碱基缺失突变(参照序列号47和48)的系统中,显示在高温条件不能形成种子。
57.图14是显示由分别来源于稻、番茄和拟南芥的温度敏感性雄性不育基因所编码的氨基酸序列、和后述的实施例中制作的基因组编辑个体中的突变位置的概略图。图中(i)和(ii)表示稻中的突变导入部位(用crispr/cas9导入了双链切割的位置),(iii)表示拟南芥中的突变导入部位,(iv)和(v)表示番茄中的突变导入部位。
具体实施方式
58.(温度敏感性雄性不育植物的制造方法)
59.如后述的实施例所示,本发明者们搞清楚了与作为温度敏感性雄性不育系统的“水稻中间母本农12号”(pl12)的表型相关的责任基因。进而,通过在野生型的稻、拟南芥和番茄利用基因组编辑法抑制该基因的功能,从而成功地对这些植物赋予了温度敏感性雄性不育性状。因此,本发明的温度敏感性雄性不育植物的制造方法的特征在于,包括人工地抑制所述基因(温度敏感性雄性不育基因)的功能的工序,更具体地提供以下发明。
60.温度敏感性雄性不育植物的制造方法,该方法包括人工地抑制植物的、选自由下述(a)~(d)组成的组中的至少一种基因的功能的工序,
61.(a)编码由序列号1~22中的任一项所记载的氨基酸序列组成的蛋白质的基因
62.(b)编码由在序列号1~22中的任一项所记载的氨基酸序列中替换、缺失、添加和/或插入了1个或多个氨基酸的氨基酸序列组成的蛋白质的基因
63.(c)编码与序列号1~22中的任一项所记载的氨基酸序列具有60%以上的同源性的氨基酸序列的基因
64.(d)包含与由编码序列号1~22中的任一项所记载的氨基酸序列的核苷酸序列组成的dna在严格条件下杂交的dna的基因。
65.在本发明中,所谓“温度敏感性雄性不育”是指在后述的容许温度条件下进行了栽培时正常结实,与此相对,在后述的限制温度条件下进行了栽培时变为雄性不育的性状。
66.在本发明中,作为成为赋予温度敏感性雄性不育性状的对象的“植物”,只要是具有后述的温度敏感性雄性不育基因的植物就不特别限制,可列举例如,包括单子叶植物(例如,稻、大麦、小麦、高粱、玉米等稻科植物、洋葱、葱等石蒜科植物)和双子叶植物(例如,拟南芥、白菜、欧洲油菜(菜籽)、野生甘蓝(甘蓝、花椰菜、青花菜等的原种)等十字花科植物、番茄、马铃薯等茄科植物、莴苣、向日葵等菊科植物、大豆等豆科植物、黄瓜等葫芦科植物、胡萝卜等伞形科植物、苹果等葫芦科植物、香蕉等芭蕉科植物、无油樟科植物(无油樟(amborella trichopoda)))的被子植物、裸子植物、苔藓类植物、蕨类植物。进而,可以是这些植物的基因重组体、基因组编辑体(例如,除草剂耐性作物、害虫耐性作物、病害耐性作物、风味提高作物、保存性提高作物、收量提高作物)。
67.作为在本发明中成为功能抑制的对象的“温度敏感性雄性不育基因”,编码各植物种来源的典型的氨基酸序列的基因的例子如下述表1所示。
68.表1
[0069][0070]
此外,在自然界中核苷酸序列突变也是可以发生的。并且,伴随突变所编码的氨基酸也可以变化。因此,本发明的温度敏感性雄性不育基因只要能够通过抑制其功能而赋予温度敏感性雄性不育的性状,就也包括编码由在序列号1~22中的任一项所记载的氨基酸序列中替换、缺失、添加和/或插入了1个或多个氨基酸的氨基酸序列组成的蛋白质的基因。
[0071]
这里所谓“多个”,通常为50个氨基酸以内,优选为45个氨基酸以内,更优选为40个氨基酸以内,进一步优选为35个氨基酸以内,更优选为30个氨基酸以内,进一步优选为25个氨基酸以内,更优选为20个氨基酸以内,进一步优选为15个氨基酸以内,更优选为10个氨基
酸以内(例如,9个氨基酸以内,8个氨基酸以内,7个氨基酸以内,6个氨基酸以内),特别优选为数个氨基酸以内(例如,5个氨基酸以内,4个氨基酸以内,3个氨基酸以内,2个氨基酸以内)。
[0072]
进而,在现在的技术水准下,只要是本领域技术人员,就能够在得到了特定的基因的情况下,利用该基因的核苷酸序列信息,从同种或其他植物鉴定其同源基因。作为用于鉴定同源基因的方法,可列举例如,杂交技术(southern,e.m.,j.mol.biol.,98:503,1975)、聚合酶链反应(pcr)技术(saiki,r.k.,et al.science,230:1350-1354,1985、saiki,r.k.et al.science,239:487-491,1988)。为了鉴定同源基因,通常在严格条件下进行杂交反应。作为严格的杂交条件,可以例示6m尿素、0.4%sds、0.5
×
ssc的条件或与此同等严格性的杂交条件。如果使用严格性更高的条件,例如6m尿素、0.4%sds、0.1
×
ssc的条件,则可以期待同源性更高的基因的分离。本发明的温度敏感性雄性不育基因只要能够通过抑制其功能而赋予温度敏感性雄性不育的性状,就也包括与由编码序列号1~22中的任一项所记载的氨基酸序列的核苷酸序列组成的dna在严格条件下杂交的dna。
[0073]
由所鉴定的同源基因编码的蛋白质通常与由所述特定的基因编码的蛋白质具有高的同源性(高的类似性)、优选为高的同一性。这里所谓“高的”是指至少40%以上、更优选为50%以上、进一步优选为60%以上、更优选为70%以上、进一步优选为80%以上、更优选为85%以上(例如,90%以上、95%以上、96%以上、97%以上、98%以上、99%以上)。本发明的温度敏感性雄性不育基因只要能够通过抑制其功能而赋予温度敏感性雄性不育的性状,就也包括编码与序列号1~22中的任一项所记载的氨基酸序列具有60%以上的同源性(类似性)或40%以上的同一性的氨基酸序列的基因。此外,序列号14所记载的氨基酸序列相对于序列号1所记载的氨基酸序列的同源性为65%(同一性为45%),序列号14所记载的氨基酸序列相对于序列号9或44所记载的氨基酸序列的同源性为73%(同一性为55%)。
[0074]
序列的同源性可以利用blast的程序(altschul et al.j.mol.biol.,215:403-410,1990)来确定。该程序基于karlin和altschul的算法blast(proc.natl.acad.sci.usa,87:2264-2268,1990,proc.natl.acad.sci.usa,90:5873-5877,1993)。例如,在通过blast分析氨基酸序列时,参数为例如score=50、wordlength=3。另外,在使用gapped blast程序分析氨基酸序列时,可以如altschul等(nucleic acids res.25:3389-3402,1997)所记载那样进行。在使用blast和gapped blast程序时,使用各程序的默认参数。这些分析方法的具体手法是公知的。
[0075]
本发明的“温度敏感性雄性不育基因的功能的人工抑制”包括该功能的完全抑制(阻遏)和部分抑制的双方。另外,除了温度敏感性雄性不育基因的表达的人工抑制之外,还包括由温度敏感性雄性不育基因所编码的蛋白质的活性的人工抑制。并且,该人工抑制可以通过例如,在温度敏感性雄性不育基因的编码区域、非编码区域、转录控制区域(启动子区域)等导入突变来进行。
[0076]
在本发明中,作为被导入温度敏感性雄性不育基因中的突变,只要抑制该基因的功能就不特别限制,可列举例如,核苷酸的替换、缺失、添加、和/或插入,优选为无义突变(nonsense mutation)、移码突变、无效突变(null mutation)。另外,即使作为被导入温度敏感性雄性不育基因的突变的个数,只要抑制该基因的功能就不特别限制,可以是1个,另外也可以是多个(例如,2个、3个以下、5个以下、10个以下、20个以下、30个以下、40个以下、
prokaryotic and vertebrate adaptive immune systems,science,doi:10.1126/science.aaf8729,(2016))等指导rna与蛋白质的复合体的方法。
[0084]
作为物理诱变法,可列举例如,重离子束(hib)照射、快中子射线照射、γ射线照射、紫外线照射(参照hayashi等、cyclotrons and their applications、2007年、第18回国际会议、237~239页,和kazama等、plant biotechnology、2008年、25卷、第113~117页)。
[0085]
作为使用化学诱变剂的方法,可列举例如,用化学诱变剂处理种子等的方法(参照zwar和chandler、planta、1995年、197卷、39~48页等)。作为化学诱变剂不特别限制,可列举甲磺酸乙酯(ems)、n-乙基-n-亚硝基脲(enu)、n-甲基-n-亚硝基脲(mnu)、叠氮化钠、亚硫酸氢钠、羟胺(
ヒドリキシルアミン
)、n-甲基-n
’‑
硝基-n-硝基胍(mnng)、n-甲基-n
’‑
亚硝基胍(ntg)、o-甲基羟胺、亚硝酸、甲酸和核苷酸类似物。
[0086]
作为将转座子等导入基因组dna的方法,可列举例如,将t
os
17等转座子、t-dna等插入植物的基因组dna的方法(参照kumar等、trends plant sci.、2001年、6卷、3号、127~134页,和tamara等、trends in plant science、1999年、4卷、3号、90~96页)。
[0087]
对于通过以上的方法导入了突变的植物,可以通过公知的方法确认在温度敏感性雄性不育基因中导入了突变。作为该公知的方法,可列举例如,dna测序法(新一代测序法等)、pcr法、使用微阵列的分析法、dna印迹法、rna印迹法。根据这些方法,可以通过比较突变导入前后的该基因的序列或长度来判断在温度敏感性雄性不育基因中是否导入了突变。另外,如果通过利用rna印迹法、rt-pcr法、蛋白质印迹法、elisa法、利用微阵列的分析法等,在转录控制区域等中导入了突变的植物中确认了温度敏感性雄性不育基因的转录产物或翻译产物的表达量降低,则也可以确认该植物是在温度敏感性雄性不育基因导入了突变的植物。
[0088]
另外,作为确认在温度敏感性雄性不育基因中导入了突变的其他方法,可列举tilling(定向诱导基因组局部突变技术、targeting induced local lesions in genomes)(参照slade等、transgenic res.、2005年、14卷、109~115页,和comai等、plant j.、2004年、37卷、778~786页)。特别是在使用前述的重离子束照射、化学诱变剂等在植物基因组中导入非选择性突变的情况下,将温度敏感性雄性不育基因或其一部分用pcr扩增之后,通过所述tilling等筛选该扩增产物中具有突变的个体。
[0089]
另外,使通过上述的方法被导入了突变的植物与野生型的植物交配,进行回交,能够除去在目的基因以外的基因中导入的突变。
[0090]
通过在温度敏感性雄性不育基因导入突变而该基因的功能被抑制了的植物有时是温度敏感性雄性不育基因的杂合子(heterozygote)。在这样的情况下,例如,通过将该杂合子彼此交配而获得f1植物体,从而由该f1植物体筛选具有被导入了该突变的温度敏感性雄性不育基因的纯合子(homozygote)。该情况下,“是具有被导入了该突变的温度敏感性雄性不育基因的纯合子的植物”不仅包括具有2个具有彼此相同的突变的温度敏感性雄性不育基因的等位基因(allele)的植物,还包括具有编码具有第1突变而活性被抑制了的蛋白质的第1温度敏感性雄性不育基因、和编码具有第2突变而活性被抑制了的蛋白质的第2温度敏感性雄性不育基因的植物。
[0091]
在本发明中,作为用于人工地抑制温度敏感性雄性不育基因的功能的方法,除了上述的突变导入之外,可列举使用编码与温度敏感性雄性不育基因的转录产物互补的
dsrna(双链rna、例如sirna)的dna的方法、使用编码与温度敏感性雄性不育基因的转录产物互补的反义rna的dna(反义dna)的方法、使用编码具有特异性地使温度敏感性雄性不育基因的转录产物开裂的核糖体活性的rna的dna的方法(核糖体法)这样的以温度敏感性雄性不育基因的转录产物作为靶标的方法。
[0092]
在本发明中,温度敏感性雄性不育基因功能的人工抑制可以根据上述的方法等,对各种植物体、种子或植物细胞进行。植物细胞中除了植物来源的培养细胞之外,也包含植物体中的细胞。进一步包含各种形态的植物来源的细胞,例如,悬浮培养细胞、原生质体、叶的切片、愈伤组织、未成熟胚、花粉等。
[0093]
另外,在本发明中,可以将编码上述的部位特异性核酸酶、融合蛋白质或指导rna与蛋白质的复合体的dna、编码转座子的dna、编码双链rna的dna、编码反义rna的dna、编码具有核糖体活性的rna的dna等以插入到载体中的形态导入植物的细胞。
[0094]
作为被插入用于人工地抑制温度敏感性雄性不育基因的功能的所述dna的载体,只要能够在植物细胞内表达插入基因,就不特别限制,可以含有用于恒常或诱导地表达所述dna的启动子。作为用于恒常地表达的启动子,可列举例如,稻的泛素启动子、花椰菜花叶病毒的35s启动子、稻的肌动蛋白启动子、玉米的泛素启动子等。另外,作为用于诱导地表达的启动子,可列举例如,已知由于丝状菌/细菌/病毒的感染或侵入、低温、高温、干燥、紫外线照射、特定的化合物的散布等外因而表达的启动子等。进而,作为用于表达作为本发明所涉及的dna编码指导rna、sirna等短rna的dna的启动子,适合使用u6启动子、poliii系的启动子。
[0095]
作为向植物细胞导入所述dna或插入有该dna的载体等的方法,可以使用例如,粒子枪法、粒子轰击法、农杆菌介导的方法(农杆菌法)、聚乙二醇法、电穿孔法(electroporation)等本领域技术人员公知的各种方法。
[0096]
此外,即使不采用dna的形态,将上述的部位特异性核酸酶、融合蛋白质、转座子作为蛋白质导入植物细胞,将上述的指导rna、双链rna、反义rna、具有核糖体活性的rna作为rna导入植物细胞,也能够导入突变。
[0097]
另外,通过由上述的方法等而基因的功能被人工地抑制了的植物细胞再生植物体,从而能够获得温度敏感性雄性不育植物。
[0098]
例如,关于在稻中制成转化植物体的方法,通过聚乙二醇对原生质体进行基因导入、使植物体再生的方法(datta,s.k.in gene transfer to plants(potrykus i and spangenberg eds.)pp66-74,1995),通过电脉冲对原生质体进行基因导入、使植物体再生的方法(toki et al.plant physiol.100,1503-1507,1992),通过粒子枪法对细胞直接导入基因、使植物体再生的方法(christou et al.bio/technology,9:957-962,1991),以及介由农杆菌导入基因、使植物体再生的方法(hiei et al.plant j.6:271-282,1994)等几种技术已经确立,在本发明的技术领域中广泛被使用。
[0099]
对于拟南芥,可列举浸花(floral-dip)法(clough sj&bent af,plant j16:735-743,1998)、akama等(akama et al.plant cell reports 12:7-11,1992)的方法,在本发明中可以适合使用这些方法。
[0100]
另外,作为对于麦制成转化植物体的方法,可以列举tingay等(tingay s.et al.plant j.11:1369-1376,1997)、murray等(murray f et al.plant cell report 22:
397-402,2004)、和travalla等(travalla s et al.plant cell report 23:780-789,2005)所记载的方法。
[0101]
作为再生高粱植物体的方法适合使用例如,通过农杆菌法或粒子枪法对未成熟胚或愈伤组织进行基因导入而使植物体再生的方法、使用通过超声波导入了基因的花粉进行传粉的方法(j.a.able et al.,in vitro cell.dev.biol.37:341-348,2001、a.m.casas et al.,proc.natl.acad.sci.usa 90:11212-11216,1993、v.girijashankar et al.,plant cell rep 24:513-522,2005、j.m.jeoung et al.,hereditas 137:20-28,2002、v girijashankar et al.,plant cell rep 24(9):513-522,2005、zuo-yu zhao et al.,plant molecular biology 44:789-798,2000、s.gurel et al.,plant cell rep 28(3):429-444,2009、zy zhao,methods mol biol,343:233-244,2006、ak shrawat and h lorz,plant biotechnol j,4(6):575-603,2006、d syamala and p devi indian j exp biol,41(12):1482-1486,2003、z gao et al.,plant biotechnol j,3(6):591-599,2005)。
[0102]
对于玉米,可列举shillito等(bio/technology,7:581,1989)所记载的方法、gorden-kamm等(plant cell 2:603,1990)所记载的方法。
[0103]
对于番茄,可列举matsukura等(j.exp.bot.,44:1837-1845,1993)所记载的方法。
[0104]
对于大豆,可列举专利公报(美国专利第5,416,011号)所记载的方法。
[0105]
对于马铃薯,可列举visser等(theor.appl.genet,78:594,1989)所记载的方法。
[0106]
另外,对于其他植物,也可以使用tabei等(田部井丰编、《形質転換
プロトコール
[植物編](转化方案[植物篇])》、株式会社化学同人、2012年9月20日出版)所记载的方法等,进行对转化和植物体的再生。
[0107]
(温度敏感性雄性不育植物、及其利用)
[0108]
通过上述的方法等,能够获得本发明的温度敏感性雄性不育基因的功能被人工地抑制了的温度敏感性雄性不育植物。因此,本发明提供温度敏感性雄性不育植物,其中
[0109]
选自由下述(a)~(d)组成的组中的至少一种基因的功能被人工地抑制,
[0110]
(a)编码由序列号1~22中的任一项所记载的氨基酸序列组成的蛋白质的基因
[0111]
(b)编码由在序列号1~22中的任一项所记载的氨基酸序列中替换、缺失、添加和/或插入了1个或多个氨基酸的氨基酸序列组成的蛋白质的基因
[0112]
(c)编码与序列号1~22中的任一项所记载的氨基酸序列具有60%以上的同源性的氨基酸序列的基因
[0113]
(d)包含与由编码序列号1~22中的任一项所记载的氨基酸序列的核苷酸序列组成的dna在严格条件杂交的dna的基因。
[0114]
此外,关于温度敏感性雄性不育基因、其功能的人工抑制、通过该抑制而被赋予温度敏感性雄性不育的植物等如上所述,但作为本发明的温度敏感性雄性不育植物,优选为除了“pl12”(水稻中间母本农12号)以外的植物,更优选为除了本发明的温度敏感性雄性不育基因的功能被人工地抑制了的
“レイメイ”
以外的植物。
[0115]
另外,如果一旦得到了温度敏感性雄性不育基因的功能被人工地抑制了的植物体,则能够由该植物体通过有性生殖或无性生殖获得后代。进而,能够由该植物体、其后代或克隆获得繁殖材料(例如,种子、切穗、植株、愈伤组织、原生质体等),以它们为基础量产该植物体。因此,本发明包括温度敏感性雄性不育植物的后代和克隆、以及它们的繁殖材
料。此外,作为繁殖材料,可列举例如,种子、株、愈伤组织、原生质体。
[0116]
另外,本发明的温度敏感性雄性不育植物可以在如图2所示的利用通过2系法的杂种种子的制造法中利用。因此,本发明还提供杂种种子的制造方法,该方法包括
[0117]
使在限制温度下栽培、制成了雄性不育的权利要求2所述的温度敏感性雄性不育植物,与任意的植物进行异花传粉的工序,和
[0118]
由所述温度敏感性雄性不育植物回收种子的工序。
[0119]
在本发明的杂种种子的制造中,使用将本发明的温度敏感性雄性不育植物在限制温度下栽培、处于雄性不育状态的植物。
[0120]
所谓“限制温度”,是指高于本发明的温度敏感性雄性不育植物能够形成花粉的栽培温度(容许温度)的温度。该限制温度和容许温度根据植物的种类等,只要是本领域技术人员就能够适当调整。
[0121]
另外,限制温度下的栽培根据植物的种类等使用公知的栽培方法进行即可。另外,作为其栽培期间只要包含从花芽形成(成花)到花粉形成的期间就不特别限制。此外,所谓“花芽”是进行营养生长起来的生长点分化成进行生殖生长的生长点而得的、即花的原基。所述形成期间根据植物的种类、其品种/系统、栽培条件等不同而不同,只要是本领域技术人员就能够通过公知的手法(例如,目视)判断。另外,该期间中的限制温度下的栽培既可以是连续的(例如,不分昼夜在限制温度下栽培),另外也可以是间歇的(例如,仅白天在限制温度下栽培)。
[0122]
作为与本发明的温度敏感性雄性不育植物交配的“任意的植物”,只要是维持着雄性的育性的植物就不特别限制,可列举例如,与本发明的温度敏感性雄性不育植物同种的、不同的系统或品种。
[0123]
本发明的温度敏感性雄性不育植物与任意的植物的“异花传粉”、和由此形成的“种子的回收”,只要是本领域技术人员,就能够使用适应植物的种类等的公知的方法适当进行。
[0124]
另外,这样得到的种子成为以本发明的温度敏感性雄性不育植物作为母本(母系)、以任意的植物作为父本(父系)的一代杂种(f1)的种子、所谓杂种种子。因此,在本发明中还提供杂种种子,
[0125]
其母本是在限制温度下栽培、制成了雄性不育的本发明的温度敏感性雄性不育植物,其父本是任意的植物。
[0126]
实施例
[0127]
以下,基于实施例更具体地说明本发明,但本发明不限于以下实施例。另外,本实施例除了后述的关于番茄的实施例5以外,使用以下所示的材料和方法进行。
[0128]
(品种、系统)
[0129]
关于稻,使用了标准品种“日本晴”、温度敏感性雄性不育系统水稻“中间母本农12号”(pl12)及其原品种
“レイメイ”
。另外,也使用了稻的实用多产品种“北陆193号”(印度型稻)。关于拟南芥,使用了标准生态型columbia。
[0130]
(植物的栽培方法)
[0131]
稻在其通常的栽培条件(昼28℃/夜22℃)下进行了盆栽培。短日照条件下诱导成花,在昼28℃/夜22℃或昼33℃/夜22℃继续栽培。出穗时观察花粉的形成,同时确认种子育
性。
[0132]
拟南芥在设定为21℃连续光照的条件的生长箱中进行了栽培。抽苔后在21℃或27℃继续栽培,观察了花粉形成、结实的样子。
[0133]
(稻的基因组编辑)
[0134]
利用了在植物中一般使用的crispr/cas9法。更具体地,将表达指导rna、cas9、和作为筛选标志物的潮霉素抵抗性基因的载体,通过农杆菌法,导入稻品种“日本晴”的愈伤组织或“北陆193号”的未成熟胚中。此外,该载体以ppzp202为基础,能够将指导rna和cas9分别在稻的u6启动子和稻的泛素启动子的控制下表达。接着,由用潮霉素抵抗性筛选而得的愈伤组织获得再分化植物,由其叶制备基因组dna,确认了基因组编辑部分的碱基序列。选定具有单碱基插入等密码子的阅读框移动的突变的突变体。在这些突变体中,不能表达该基因发挥功能的蛋白质,因而成为基因缺损的状态。
[0135]
然后,将稻品种“pl12”、以及“日本晴”、“北陆193号”和它们的基因组编辑系统在温室内在通常的栽培条件(昼28℃/夜22℃)进行盆栽培。在短日照条件下诱导成花,在昼28℃/夜22℃或高温条件(对于“日本晴”为昼33℃/夜22℃、对于“北陆193号”为昼35℃/夜25℃)继续栽培,出穗时观察花粉的形成。
[0136]
(拟南芥的基因组编辑)
[0137]
与所述稻同样地,使用crispr/cas9法进行基因组编辑。与所述稻基因导入方法不同。拟南芥的转化使用一般使用的浸花(floral-dip)法。
[0138]
温度敏感性雄性不育系统水稻“pl12”从品种
“レイメイ”
的利用γ射线照射的突变集团中筛选培育出。该系统在通常的栽培温度(28℃程度)下正常形成花粉而结实,但高温(33℃程度)变成花粉形成不全而看不到结实(参照图3)。另外,叶等营养器官、雌蕊未见异常,可知敏感性期是出穗前20天前后的数天之间。由遗传学的分析显示,该温度敏感性雄性不育的性状是由隐性的单因子引起的,责任突变位于第7染色体上(参照非专利文献1)。于是如下所述,尝试了温度敏感性雄性不育性状的责任突变的鉴定。
[0139]
(实施例1)
[0140]
为了鉴定温度敏感性雄性不育性状的责任突变,进行了精细定位,但不能比54cm附近的约1.8mb再缩小范围。于是,进行原品种的
“レイメイ”
与“pl12”的全基因组序列分析,将两者进行了比较。其结果判明,“pl12”第7染色体的该部分有约150kb的缺失。
[0141]
接着,通过利用使用了2个指导rna的crispr/cas9法,在基因组dna的2处导入双链切割,使之间的dna片段缺失的方法,制成部分缺失的系列。更具体地,如图4所示,以将约150kb的缺失区域大致3等分的方式,制成具有(1)~(2)、(2)~(3)、(3)~(4)的范围的部分缺失的系统。此外,为了使(1)~(2)、(2)~(3)、和(3)~(4)缺失,以序列号23和24、24和25、以及25和26作为目标序列,通过crispr/cas9分别产生2处双链切割。
[0142]
其结果如图5所示,可知其中1个(#571)与“pl12”显示同样的表型,将候选区域缩小到了(3)~(4)的范围。
[0143]
进而同样地,在#571的缺失区域的内部设计3个部分缺失。更具体地,如图4所示,以将(3)~(4)的区域大致分3份的方式,制成具有(3)~(5)、(5)~(6)、(6)~(4)的范围的部分缺失的系统。此外,为了使(3)~(5)、(5)~(6)、和(6)~(4)缺失,以序列号25和27、27和28、以及28和26作为目标序列,通过crispr/cas9分别产生2处双链切割。
[0144]
其结果如图5所示,由#592显示温度敏感性雄性不育的性状,考虑在(3)至(5)的约10kb的范围有责任突变。由数据库的信息预想,该区域存在2个基因(图4所示的orf1和orf2)。
[0145]
因此,在各自的编码区域内设计指导rna,通过导入移码突变尝试了基因功能的破坏。此外,将由orf1编码的氨基酸序列示于序列号1,将由orf2编码的氨基酸序列示于序列号29。将对orf1的指导rna中的目标序列示于序列号30或31。将对orf2的指导rna中的目标序列示于序列号32。另外,将所得的突变的例子对于orf1示于表2。
[0146]
表2
[0147][0148]
其结果如图5所示,#612的系统显示温度敏感性雄性不育的性状。进而,虽未图示,但确认了不限于图5所示的突变株的代表例,只要发生移码突变,就显示温度敏感性雄性不育的性状,判明了orf1的基因功能缺失是“pl12”的责任突变。
[0149]
该基因在msu数据库中以loc_os07g26794的id预测为基因,在rap-db中以os07g0482700的id预测为基因,但却是功能未知的新基因。该基因在生物中广泛保守,特别在植物种中保守性高。将预想的氨基酸序列在高等植物中的比较示于图6。稻科作物之间序列的保守性更高(参照图7)。另外,将这样新鉴定的温度敏感性雄性不育基因命名为tms2。
[0150]
(实施例2)
[0151]
将缺损了所述基因的基因组编辑系统,与基因组编辑前的标准品种“日本晴”、温度敏感性雄性不育系统“pl12”进行比较,在容许温度和限制温度下栽培,观察了颖花的样子。其结果如图8所示,在标准品种“日本晴”中不依赖于温度而正常地形成了花粉。另一方面,温度敏感性雄性不育系统“pl12”在28℃正常地形成花粉而形成黄色且充实的花药,与此相对,在33℃花粉不发达,花药的形态、颜色变得异常。并且,通过基因组编辑而在“日本晴”中缺损了所述基因的系统与“pl12”同样地显示温度敏感性雄性不育的表型。
[0152]
进而,将稻品种“日本晴”的通过基因组编辑而基因功能缺失了的系统在28℃和33℃进行了栽培。其结果如图9所示,在28℃正常结实而穗垂下,与此相对,在33℃由于不结实而形成穗立着的状态。
[0153]
(实施例3)
[0154]
使用稻的实用多产品种“北陆193号”(印度型稻),与实施例2(“日本晴”(日本型稻))同样地进行基因组编辑,使温度敏感性雄性不育基因缺损。此外,“北陆193号”的转化参照樋江井祐弘“agrobacterium tumefaciens
によるイネ
形質転換方法
に
関
する
研究”2014年名古屋大学学位论文进行。然后,将这样得到的“北陆193号”来源的基因组编辑系统
在设定为通常条件(28℃)或高温条件(35℃)的温室栽培,观察了所形成的花药。
[0155]
其结果如图10所示,确认了在“北陆193号”来源的基因组编辑系统中,也与“日本晴”来源的基因组编辑系统同样地,在通常条件下形成花粉而花药发达,但在高温条件下由于花粉形成不全而变成雄性不育。
[0156]
另外,在开花期对“北陆193号”来源的基因组编辑系统授予其他多产品种
“オオナリ”
的花粉,使结成的种子发芽而进行了栽培。然后,从对(1)
“オオナリ”
、(2)“北陆193号”来源的基因组编辑系统、和(3)高温下栽培的“北陆193号”来源的基因组编辑系统授予
“オオナリ”
的花粉而得的个体各自的叶分别纯化基因组dna,分析温度敏感性雄性不育基因的基因组编辑部位(突变插入部位)的碱基序列。
[0157]
其结果如图11所示,在(3)的个体中形成了(1)与(2)的基因组序列的杂合子,显示通过(1)与(2)的交配得到了杂种。由此确认了即使在将基因组编辑系统在高温栽培的情况下,雌蕊也正常发挥功能,基因组编辑系统能够实际在交配中利用。
[0158]
(实施例4)
[0159]
对于作为双子叶植物的拟南芥,也尝试了通过使所述基因缺损,而制成温度敏感性雄性不育系统。更具体地,通过crispr/cas9法进行基因组编辑,制成了所述基因不发挥功能的系统。
[0160]
此外,将拟南芥中的目标序列示于序列号33。另外,将所得的突变的例子示于表3。
[0161]
表3
[0162][0163]
然后,将通过拟南芥的基因组编辑而基因功能缺失了系统在21℃和27℃进行了栽培。其结果如图12所示,在21℃正常结实且荚果的长度也与野生型无变化,与此相对,在27℃不结实且荚果不伸长。
[0164]
此外,虽未图示,但与上述的稻同样地确认了,不限于图12所示的突变株的代表例,只要发生移码突变,在拟南芥中就也显示与所述温度敏感性雄性不育有关的性状。
[0165]
另外,与实施例3同样地,在拟南芥中也进行了交配实验。其结果虽未图示,但能够获得种子。因此,与上述稻同样地,温度敏感性雄性不育基因的功能抑制不影响雌蕊的功能,提示在拟南芥中基因组编辑系统也能够在交配中利用。
[0166]
(实施例5)
[0167]
在番茄中进行温度敏感性雄性不育基因的基因组编辑,建立了突变系统。具体地,首先,从ensenmbleplants获得番茄的温度敏感性雄性不育基因的基因组序列信息。此外,将该基因的cdna(野生型)的序列示于序列号43。另外,将由该cdna编码的蛋白质的氨基酸序列示于序列号9或44(在序列号9与44之间显示同一的氨基酸序列)。
[0168]
然后,以所得的基因组序列信息作为对象,通过crisprdirect(http://
crispr.dbcls.jp/)检索crispr/cas的目标序列。结果选定了位于外显子内、在番茄基因组上没有类似序列的下述2个序列作为基因组编辑目标序列。
[0169]
ge51(第3外显子内)
[0170]5’‑
accataggtgagaagtcacgagg-3’(序列号37)
[0171]
ge52(第4外显子内)
[0172]5’‑
ccaggctgtctaccagagaaatg-3’(序列号38)。
[0173]
接着,制作基因组编辑用载体。该载体中使用了植物基因组编辑用基因组编辑载体pegp237-2a-gfp。pegp237-2a-gfp由德岛大学刑部教授分让。载体的详情请参照ueta et al.(2017)rapid breeding of parthenocarpic tomato plants using crispr/cas9.sci rep.7:507。
[0174]
将pegp237-2a-gfp用bsai消化后,进行了纯化。接着,将化学合成的下述各寡聚dna等量混合后,进行热变性处理,然后退火。将所得的双链寡聚dna与经bsai消化的pegp237-2a-gfp混合,插入该载体,进行了连接。然后,导入大肠杆菌,扩增了基因组编辑用载体。
[0175]
#51
[0176]
sl_tms2_grna01f
[0177]5’‑
gattgaccataggtgagaagtcacg-3’(序列号39)
[0178]
sl_tms2_grna01r
[0179]5’‑
aaaccgtgacttctcacgatacca-3’(序列号40)
[0180]
#52
[0181]
sl_tms2_grna02f
[0182]5’‑
gattgcatttctctggtagacagcg-3’(序列号41)
[0183]
sl_tms2_grna02r
[0184]5’‑
aaacggctgtctaccagagaaatg-3’(序列号42)。
[0185]
然后,将这样构建的基因组编辑用载体转化放射型根瘤菌(根癌农杆菌(agrobacterium tumefaciens))gv2260,进一步使用农杆菌法进行番茄品种“microtom”的转化。此外,关于番茄的转化法,请参照sun et al.(2006)a highly efficient transformation protocol for micro-tom,a model cultivar for tomato functional genomics.plant cell physiol.47:426-431。
[0186]
确认这样获得的转化体的目标序列,获得了基因组编辑个体。在转化当代,得不到纯和地具有突变的个体。因此,自花受精采种,从后代中选定突变被固定了的个体。
[0187]
接着,将所得的基因组编辑个体在室温23℃/18℃(昼/夜)14小时日照时长下育苗至开花前。进而移至高温30℃/28℃(昼/夜)14小时日照时长,除去了最初的花。然后,确认所结的果实有无种子。将所得的结果示于图13。
[0188]
如图13所示,在番茄中,也通过温度敏感性雄性不育基因的缺损,从而在通常条件结出种子,与此相对,在高温条件下不形成种子,与稻的情况同样地观察到温度敏感性的表型。番茄与稻同样地是自花传粉植物,自己的花粉附着于雌蕊而形成种子。由此显示了对蔬菜等作物的应用可能性。
[0189]
最后,将由本实施例中使用的各种植物来源的温度敏感性雄性不育基因编码的氨
基酸序列、和在所制作的基因组编辑个体中确认的突变位置归纳示于图14。
[0190]
产业可利用性
[0191]
如以上说明的那样,根据本发明,能够制造温度敏感性不育植物。特别是除了抑制本发明的温度敏感性雄性不育基因的功能之外,不需要特别的育种,就能够制造温度敏感性不育植物。
[0192]
另外,如图6和7所示,由本发明的温度敏感性雄性不育基因编码的蛋白质的氨基酸序列高度保守。因此,在本发明的方法中使用的系统不受限定,能够在各种植物种的各种品种/系统中通过抑制该基因的功能而制成雄性不育系统。例如,仅通过抑制有用的品种的温度敏感性雄性不育基因的功能就能够直接在杂种产生中使用。
[0193]
并且,通过以所得的温度敏感性雄性不育植物作为母本,与作为父本的其他系统一起在限制温度下栽培,从而能够简便地获得这2个系统杂交而得的杂种种子。
[0194]
这样,本发明可期待给农业生产带来大的影响,在该领域是极有用的技术。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。