一种基于lpp引导序列靶向呈递和高效表达外源抗原的重组沙门菌构建方法
技术领域
1.本发明属于动物细菌基因工程技术领域,具体通过质粒改造引入lpp引导序列以提高沙门菌中外源抗原表达水平和免疫原性的一种构建方法。
背景技术:
2.猪链球菌(streptococcus suis)是一种重要的人畜共患病原体,是猪链球菌病的主要病原。它能够导致多种疾病包括脑膜炎、关节炎和败血症等,不仅给养猪生产业造成巨大的经济损失,也给相关从业人员的健康造成了巨大的威胁。根据其荚膜多糖不同可分为33种血清型。猪链球菌2型是临床分离率最高和致病力最强的血清型,猪链球菌9型和7型在世界范围内也引起广泛关注。saoa蛋白几乎在所有猪链球菌血清型中均有分布的高度保守具有免疫原性的抗原蛋白。我们的前期研究证实递送猪链球菌saoa蛋白的重组减毒猪霍乱沙门菌可有效保护小鼠免受猪链球菌多种血清型的感染。细菌感染仍然是导致人类和动物死亡的主要原因之一。由于耐药菌的增加和快速传播,抗生素治疗细菌性疾病的有效性受到了挑战。因此,疫苗被认为是后抗生素时代应对细菌性疾病最直接有效的策略。目前针对细菌的商品化疫苗主要包括亚单位疫苗、灭活苗和弱毒苗。这些疫苗目前都存在着一定的不足。亚单位疫苗的成本相对较高且交叉保护效果不够理想;灭活苗会使其丢失大部分抗原表位、免疫保护持续时间短,保护性不足;弱毒苗具有毒力返强的风险,存在很大的安全隐患。细菌性疾病亚单位疫苗由于其生产工艺简单、成本低及安全性高等优势,成为细菌疫苗的重要发展方向。此类疫苗主要由原核表达系统(如大肠杆菌)大规模生产,表达产物通常无法进行正确的加工和折叠,影响表达蛋白的构象导致免疫原性较差。除了安全性外,评价一个疫苗的另一个重要指标就是其免疫原性。因此如何提高细菌性疾病亚单位疫苗的免疫原性成为首要解决的问题。
3.与传统疫苗相比,omvs作为细菌疫苗存在着诸多优势:(1)免疫原性好:omvs为典型的纳米颗粒结构,有利于进入淋巴管并被抗原递呈细胞高效摄取,另外omvs包含大量天然构象的外膜抗原,可有效刺激机体的体液和细胞免疫反应;(2)有效的天然佐剂:omvs含有lps可作为佐剂,可诱导宿主产生高效的免疫应答;(3)靶向改造:omvs可通过基因工程手段被定向改造,将目标抗原包裹入囊泡腔内或者镶嵌至外膜上,然后递呈至宿主细胞。大量研究证明omvs无论是作为病原菌自身疫苗还是作为疫苗递呈载体构建重组二联疫苗均具有非常大的应用潜力。
4.外源抗原在菌体和omvs中的表达量和表达位置是影响疫苗保护效力的关键因素。作为载体所递送的外源抗原表达量低,会导致诱导针对外源病原的免疫保护效力不足。因此,外源抗原表达量低是诱导高免疫保护效力和疫苗生产一线的大忌。原则上,外源抗原表达在菌体和omvs表面是诱导抗原特异性免疫应答的最佳策略,可以增强抗原与b细胞受体的结合。然而,抗原靶向表达在菌体和omvs表面仍然具有一定的挑战性,通常需要外源抗原和细菌外膜蛋白的嵌合表达模式。相对而言,外源抗原表达在菌体和omvs内腔中则相对容
易。
5.研究已经证实,细菌可以通过脂蛋白转运途径将脂蛋白转运至菌体的表面,进而呈递到omvs的外膜。可利用基因工程手段将外源抗原与脂蛋白引导序列融合表达,以脂蛋白转运机制为理论基础创建omvs呈递系统,构建omvs递送外源抗原的二联疫苗。因此lpp引导目的抗原靶向表达在omvs膜表面的功能对于提高疫苗株omvs表达外源抗原具有重要应用潜力。
技术实现要素:
6.针对现有技术中存在的不足,本发明的目的是提供一种基于lpp引导序列靶向呈递和高效表达外源抗原的重组沙门菌构建方法。
7.本发明的目的是通过以下技术方案来实现的,一种基于lpp引导序列靶向呈递和高效表达外源抗原的重组沙门菌的重组表达质粒ps-lpp-saoa,所述重组表达质粒ps-lpp-saoa包括大肠杆菌lpp引导序列,载体为ps-saoa质粒。
8.一种基于lpp引导序列靶向呈递和高效表达外源抗原的重组沙门菌构建方法,所述方法步骤如下:
9.s1:构建重组表达质粒ps-lpp-saoa;
10.s2:利用重组表达质粒ps-lpp-saoa构建表达saoa蛋白的重组猪霍乱沙门菌载体rsc0016(ps-lpp-saoa)株。
11.优选的,所述s1的具体步骤如下:以大肠杆菌lpp引导序列(63bp)为模板,设计合成lpp引导序列;利用同源重组方法将大肠杆菌lpp引导序列(63bp)引入到ps-saoa质粒中,形成重组表达质粒ps-lpp-saoa。
12.优选的,所述s2的具体步骤如下:在猪霍乱沙门菌载体菌株rsc0016中引入ps-lpp-saoa质粒,形成表达saoa蛋白的重组猪霍乱沙门菌载体rsc0016(ps-lpp-saoa)株。
13.利用western blot、免疫荧光、流式细胞术、亚细胞定位分析和蛋白酶k酶解实验等方法检测saoa蛋白在ps-saoa和ps-lpp-saoa质粒中的表达量和定位情况。
14.检测表达saoa蛋白的沙门菌omvs针对小鼠的免疫应答水平和对小鼠的免疫保护效力。
15.本发明具有以下有益效果:本发明利用同源重组方法将大肠杆菌lpp引导序列引入到本实验室前期构建的ps-saoa质粒中,形成重组表达质粒ps-lpp-saoa,在猪霍乱沙门菌载体菌株rsc0016中引入ps-lpp-saoa质粒,形成重组猪霍乱沙门菌rsc0016(ps-lpp-saoa)。外源抗原saoa在该菌株及其omvs中可以高效表达和靶向呈递,免疫小鼠后可产生强大的体液免疫和细胞免疫,并产生高水平的细胞因子,保护小鼠免受猪链球菌致死剂量的攻击。本发明为无佐剂omvs疫苗平台提供了一种有前景的通用策略,具有较大的市场应用潜力和潜在的经济效益。
附图说明
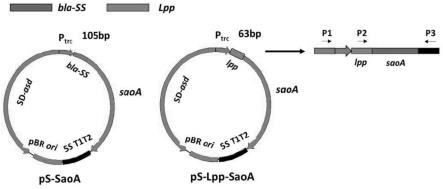
16.图1 ps-lpp-saoa质粒的构建示意图。
17.图2 ps-lpp-saoa质粒的pcr鉴定。
18.图3 saoa蛋白在χ7213(ps-lpp-saoa)和χ7213(ps-saoa)中的表达量分析。
19.图4 saoa蛋白在rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)中的表达量分析。
20.图5 saoa蛋白在χ9241(ps-lpp-saoa)和χ9241(ps-saoa)中的表达量分析。
21.图6免疫荧光法检测saoa蛋白的定位情况。
22.图7流式细胞术检测saoa蛋白的定位情况。
23.图8亚细胞定位检测saoa蛋白的定位情况。
24.图9蛋白酶k酶解实验检测saoa蛋白的在菌体中的定位情况。
25.图10蛋白酶k酶解实验检测saoa蛋白的在omvs中的定位情况。
26.图11saoa在菌体及其omvs中的脂化检测。
27.图12各免疫组针对saoa的特异性igg抗体滴度检测。
28.图13各免疫组针对saoa的特异性igg2a抗体滴度检测。
29.图14各免疫组针对saoa的特异性igg1抗体滴度检测。
30.图15各免疫组针对saoa的igg2a/igg1比值分析。
31.图16细胞免疫应答检测示意图。
32.图17各免疫组小鼠血清中il-4水平测定。
33.图18各免疫组小鼠血清中ifn-γ水平测定。
34.图19各免疫组中小鼠ifn-γ/il-4比值分析。
35.图20各免疫组小鼠血清和脾脏组织中il-17a水平测定。
36.图21各免疫组小鼠脾脏组织中il-4和ifn-γ的elispot测定。
37.图22各免疫组小鼠血液中细菌载量的检测。
38.图23各免疫组小鼠血液中细菌载量的检测。
39.图24动物保护试验中8
×
ld
50
攻毒小鼠存活率的测定。
40.图25动物保护试验中16
×
ld
50
攻毒小鼠存活率的测定。
具体实施方式
41.实施1 ps-lpp-saoa质粒和重组菌株的构建
42.1.1 ps-lpp-saoa质粒与鉴定
43.以大肠杆菌lpp引导序列(63bp)为模板,设计合成lpp引导序列。利用同源重组方法将大肠杆菌lpp引导序列(63bp)引入到本实验室前期构建的ps-saoa质粒中,形成重组表达质粒ps-lpp-saoa,ps-lpp-saoa质粒构建示意图如图1所示。随后将质粒转化至工程菌大肠杆菌χ7213中,挑取克隆株,通过pcr和测序鉴定,鉴定均正确克隆株为阳性克隆株,命名为χ7213(ps-lpp-saoa)。pcr结果如图2所示,内部引物(p2/p3)从ps-saoa质粒中未扩增出片段,外部引物(p1/p3)从ps-saoa质粒中扩增出的片段比从ps-lpp-saoa质粒中扩增出的片段稍大,证实ps-lpp-saoa质粒初步构建成功。随后的测序结果也正确(未显示),证实ps-lpp-saoa质粒构建成功。
44.1.2重组猪霍乱沙门菌载体rsc0016(ps-lpp-saoa)与重组鼠伤寒沙门菌载体χ9241(ps-lpp-saoa)的构建
45.随后向猪霍乱沙门菌载体rsc0016与鼠伤寒沙门菌载体χ9241中引入ps-lpp-saoa质粒,成功构建表达saoa蛋白的重组猪霍乱沙门菌载体rsc0016(ps-lpp-saoa)株和重组鼠伤寒沙门菌载体χ9241(ps-lpp-saoa)株。同时将不含有lpp引导序列的ps-saoa原核表达质
粒引入到相同的菌株中,形成χ7213(ps-saoa)、rsc0016(ps-saoa)和χ9241(ps-saoa)作为对照株。挑取转化后的克隆子进行pcr鉴定,经pcr鉴定正确后的阳性菌株冻存于-70℃冰箱备用。实施2saoa蛋白在ps-saoa和ps-lpp-saoa质粒中的表达量分析和定位检测
46.2.1western blot方法检测saoa蛋白在ps-saoa和ps-lpp-saoa中的表达水平
47.于-70℃冰箱取出冻存的χ7213(ps-lpp-saoa)、rsc0016(ps-lpp-saoa)和χ9241(ps-lpp-saoa)、χ7213(ps-saoa)、rsc0016(ps-saoa)和χ9241(ps-saoa)在lb培养基上划线复苏。随后挑取上述单菌落在lb培养基中37℃静置培养过夜,过夜培养物以1:100的比例接种于lb液体培养基中,摇菌至od
600
=0.6时加入终浓度1mm iptg,在37℃诱导3h、12,000
×
g离心10min收集菌体。随后沸水中煮样10min后离心取上清进行sds-page蛋白电泳,利用anti-saoa抗体作为一抗,hrp标记的羊抗兔抗体作为二抗,进行westren blot实验,随后用image j分析saoa蛋白的浓度,蛋白浓度越高,表明saoa蛋白表达量越高。western blot分析结果显示,χ7213(ps-lpp-saoa)和χ7213(ps-saoa)中均可检测到saoa蛋白,但在空质粒对照菌株χ7213(pya3493)中检测不到(图3)。χ7213(ps-saoa)菌株中检测的saoa蛋白分子量(~38kda)大于χ7213(ps-lpp-saoa)中检测的saoa分子量(~37kda)。证实lpp引导序列成功引入到ps-saoa质粒中取代了bla-ss分泌信号。值得注意的是,χ7213(ps-lpp-saoa)中saoa蛋白的表达量显著上调,大概上调3.5倍左右。随后又检测了saoa在rsc0016(ps-lpp-saoa)和χ9241(ps-lpp-saoa)表达水平,结果与预期的一样,saoa在rsc0016(ps-lpp-saoa)(图4)和χ9241(ps-lpp-saoa)(图5)表达水平均显著增加,与χ7213(ps-lpp-saoa)中得到的结果一致。以上结果表明,ps-lpp-saoa质粒构建良好,在不同的菌株中saoa的表达量均高于ps-saoa质粒。
48.2.2免疫荧光和流式细胞术检测saoa蛋白在rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)中的定位情况
49.前期研究表明,外源抗原可以与bla-ss引导序列或lpp引导序列以融合蛋白的形式分别合成到细菌的周质和表面。为验证这一结果,我们用免疫荧光和流式细胞术分析saoa蛋白是否能表达在rsc0016(ps-lpp-saoa)的表面。用相同的方法将rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)进行培养,iptg诱导后收集菌体,随后1ml pbs中重悬。随后分别加入saoa抗体和fitc标记的二抗,每次抗体孵育结束后均用含1%bsa的pbs清洗3次,最终样品用2%的多聚甲醛固定15min,同时设置空质粒菌株作为对照。最后洗涤后,样品用荧光显微镜和流式细胞仪分别检测菌体表面的荧光强度。空白对照细胞的平均荧光强度设置为1,rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)细胞的相对荧光强度。免疫荧光结果如图6所示,在rsc0016(ps-lpp-saoa)的表面可以观察到明显的荧光信号。而rsc0016(ps-saoa)、rsc0016(pya3493)及空白对照均未观察到明显的荧光信号。随后流式细胞术也检测了各菌株的荧光强度,结果显示rsc0016(ps-saoa)的平均荧光强度(mfi)水平与rsc0016(pya3493)和空白对照相似,而rsc0016(ps-lpp-saoa)的平均荧光强度约为rsc0016(pya3493)的20倍(图7)。证实saoa蛋白可以表达在rsc0016(ps-lpp-saoa)的表面。
50.2.3亚细胞定位检测saoa蛋白在rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)中的定位情况
51.用相同的方法将rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)进行培养,iptg诱导后,分别提取rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)的分泌蛋白、外膜蛋白、周质蛋白
和胞质蛋白。westren blot检测saoa蛋白在菌体不同组分中的表达情况。western blot分析结果显示,在rsc0016(ps-saoa)菌株的分泌蛋白、周质蛋白和胞质蛋白可以检测到saoa蛋白的表达,而在外膜蛋白中没有检测到saoa蛋白。不出所料,在rsc0016(ps-lpp-saoa)外膜蛋白组分中检测到了saoa蛋白的表达,提示lpp-saoa嵌合体位于rsc0016(ps-lpp-saoa)的表面。结果如图8。
52.2.4蛋白酶k酶解实验检测saoa蛋白在rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)及其omvs中的定位情况
53.将准备好的rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)菌株及其omvs样品在37℃下用15μg/ml或20μg/ml蛋白酶k(pk)处理20分钟。通过western blot分析检测pk处理对saoa蛋白条带情况的影响。然后使用image j软件对光密度进行量化,确定saoa在菌体和omvs上的定位。western blot结果显示rsc0016(ps-saoa)在pk处理后没有显著影响saoa的蛋白条带密度,表明saoa没有在rsc0016(ps-saoa)的表面表达。结果显示rsc0016(ps-lpp-saoa)的saoa条带强度进行pk处理后降低了约40%,表明lpp-saoa融合抗原至少部分是暴露在rsc0016(ps-lpp-saoa)的表面。结果如图9。
54.随后,我们分析了saoa在omvs中的定位情况。结果如图10所示,大约44%的saoa表达在rsc0016(ps-lpp-saoa)omvs的表面,而rsc0016(ps-saoa)中的saoa蛋白均表达在其omvs的内腔中。
55.2.5 saoa在rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)及其omvs中的脂化检测
56.将准备好的rsc0016(ps-lpp-saoa)和rsc0016(ps-saoa)全细胞裂解物及其omvs蛋白样品稀释在pbs中,随后加入终浓度为1%的冷tritonx-114(去垢剂),4℃振荡孵育1h,然后将溶液在37℃作用10min,通过离心将亲水相和疏水相分开。接着用氯仿/甲醇法沉淀两相中蛋白进行sds-page蛋白电泳。通过western blot检测saoa在亲水相和疏水相中的表达,判断以lpp作为引导序列saoa蛋白的脂化情况。结果如图11所示,rsc0016(ps-saoa)中的saoa蛋白在水相中被检测到,而rsc0016(ps-lpp-saoa)中的saoa蛋白则一半以上保留在疏水相中。与全细胞裂解物的结果一致,rsc0016(ps-saoa)omvs样品中的大部分saoa蛋白处于水相中,而rsc0016(ps-lpp-saoa)omvs样品中超过一半的saoa蛋白在疏水相中。证实saoa在rsc0016(ps-lpp-saoa)及其omvs中可以发生脂化。
57.实施3表达saoa蛋白的沙门菌omvs针对小鼠的免疫应答水平检测
58.3.1 rsc0016(ps-lpp-saoa)omvs和rsc0016(ps-saoa)omvs体液免疫应答能力的检测
59.6周龄balb/c小鼠腹腔免疫3周后用相同免疫途径和剂量加强免疫一次,在首免5周后采集小鼠的血液,分离血清后,通过elisa测定血清中针对saoa蛋白的特异性igg、igg1和igg2a抗体滴度。结果显示rsc0016(ps-lpp-saoa)omvs免疫小鼠后产生针对saoa的特异性igg抗体滴度显著高于rsc0016(ps-saoa)omvs、rsc0016(pya3493)omvs rsaoa和rsaoa佐剂组(图12)。此外,与其他3个免疫组相比,在rsc0016(ps-lpp-saoa)omvs免疫小鼠中,针对saoa的特异性igg2a(图13)和igg1(图14)抗体滴度也显著增高。现有研究表明,igg2a与th1免疫反应相关,而igg1与小鼠th2免疫反应相关。如图15所示,3个omvs免疫组中的igg2a/igg1比值均显著高于用佐剂组。上述数据表明,相比rsc0016(ps-saoa)omvs、rsc0016(pya3493)omvs rsaoa和rsaoa佐剂组,rsc0016(ps-lpp-saoa)omvs可以诱导更强的体液免
疫应答。omvs候选疫苗免疫可诱导更加平衡的th1/th2免疫反应,而佐剂免疫组则更偏向于th2免疫反应。
60.3.2 rsc0016(ps-lpp-saoa)omvs和rsc0016(ps-saoa)omvs细胞免疫应答能力的检测
61.在小鼠模型中我们进一步探索了由酯化omvs诱导的外源抗原特异性细胞免疫反应。在加强免疫后6小时采集小鼠的血液,分离血清后,按照说明书使用小鼠细胞因子检测试剂盒检测血清样本中的细胞因子(il-4、ifn-γ和il-17a)。在加强免疫后7天,从免疫小鼠中采集脾组织用于il-4和ifn-γ的elispot试验和脾脏中il-17a测定,检测示意图如图16所示。用rsc0016(ps-lpp-saoa)omvs免疫的小鼠血清中il-4和ifn-γ细胞因子水平显著高于rsc0016(ps-saoa)omvs、rsc0016(pya3493)omvs rsaoa和rsaoa佐剂组(图17和图18)。通常,ifn-γ与th1免疫反应相关,而il-4与小鼠th2免疫反应相关。如图19所示,3个omvs免疫组中的ifn-γ/il-4比值均显著高于用佐剂组。此结果与抗体检测的结果一致。表明omvs候选疫苗免疫可诱导更加平衡的th1/th2免疫反应。与其他4个免疫组相比,rsc0016(ps-lpp-saoa)omvs免疫小鼠的血清和脾脏组织中il-17a水平显著升高(图20)。elispot结果表明,与其他免疫组相比,rsc0016(ps-lpp-saoa)omvs免疫的小鼠诱导了更多的il-4和ifn-γ分泌t细胞,与血清中观察到的结果一致(图21)。上述结果表明,rsc0016(ps-lpp-saoa)omvs免疫小鼠可诱导最高水平的il-4、ifn-γ和il-17a。
62.实施4表达saoa蛋白的沙门菌omvs针对小鼠的免疫保护效力检测
63.4.1 rsc0016(ps-lpp-saoa)omvs和rsc0016(ps-saoa)omvs免疫小鼠攻毒后细菌载量的检测
64.为了进一步评估小鼠免疫后攻毒菌在小鼠血液和脑组织中的定植能力,我们在加强免疫后2周攻毒了2
×
ld
50
的猪链球菌2型,感染24小时后对各免疫组的小鼠进行安乐死。血液样品从心脏中获得,脑组织称重后匀浆,随后将组织中的细菌梯度稀释平板计数。结果显示,与pbs组相比,rsc0016(ps-lpp-saoa)omvs、rsc0016(ps-saoa)omvs、rsc0016(pya3493)omvs rsaoa和rsaoa佐剂组的小鼠血液和脑组织中细菌载量显著降低(图22和图23)。然而,与其他免疫组相比,rsc0016(ps-lpp-saoa)omvs免疫组的小鼠血液和脑组织中细菌载量显著减少,表明rsc0016(ps-lpp-saoa)omvs免疫组具有最高的细菌清除能力。
65.4.2 rsc0016(ps-lpp-saoa)omvs和rsc0016(ps-saoa)omvs免疫小鼠攻毒后保护效率的检测
66.在加强免疫后2周攻毒致死剂量(8
×
ld
50
和16
×
ld
50
)的猪链球菌2型,攻毒后连续14天记录小鼠的存活率。结果如图24所示,所有pbs组的小鼠在受到8
×
ld
50
猪链球菌2型攻击后3天内全部死亡。显着的典型临床症状,包括跛行、嗜睡、颤抖、眼睛肿胀和毛发粗糙等。rsaoa佐剂组可提供70%的保护率并且表现出轻微的临床症状。rsc0016(ps-saoa)omvs和rsc0016(pya3493)omvs rsaoa免疫组可使小鼠的存活率提高到80%。值得注意的是,rsc0016(ps-lpp-saoa)omvs免疫组对小鼠的保护率为100%,且没有一只小鼠出现任何典型的临床症状。随后进一步评估了针对16
×
ld
50
高致死剂量猪链球菌2型的保护效率rsc0016(ps-lpp-saoa)omvs免疫组可提供90%保护效率,而rsc0016(ps-saoa)omvs、rsc0016(pya3493)omvs rsaoa免疫组和rsaoa佐剂组免疫后在相同的攻击剂量下分别提供了50%、60%和40%的保护效率(图25)。上述结果证实了rsc0016(ps-lpp-saoa)omvs可以
在小鼠感染模型中保护小鼠免受猪链球菌致死剂量的攻击。
67.综上所述,本发明在猪霍乱沙门菌载体菌株rsc0016中引入ps-lpp-saoa质粒,成功构建表达saoa蛋白的重组猪霍乱沙门菌rsc0016(ps-lpp-saoa)。外源抗原saoa在该菌株及其omvs中可以高效表达和靶向呈递,免疫小鼠后可产生强大的体液免疫和细胞免疫,并产生高水平的细胞因子,保护小鼠免受猪链球菌致死剂量的攻击。本发明为无佐剂omvs疫苗平台提供了一种有前景的通用策略,具有较大的市场应用潜力和潜在的经济效益。
68.最后应说明的是:以上所述的各实施例仅用于说明本发明的技术方案,而非对其限制;尽管参照前述实施例对本发明进行了详细的说明,本领域的普通技术人员应当理解:其依然可以对前述实施例所记载的技术方案进行修改,或者对其中部分或全部技术特征进行等同替换;而这些修改或替换,并不使相应技术方案的本质脱离本发明各实施例技术方案的范围。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。