用于控制嵌合抗原受体t细胞激活/抑制的连接臂及其应用
1.本案为分案申请,其母案申请日为2018年8月9日,申请号为201810900635.5,发明名称为“用于控制嵌合抗原受体t细胞激活/抑制的连接臂及其应用”。
技术领域
2.本发明属于细胞免疫治疗领域,涉及一种可”开关”模式调控嵌合抗原受体t细胞(car-t)的技术及应用方法。
背景技术:
3.car-t技术已经成为细胞免疫治疗的关键性技术,在诸如肿瘤、自身免疫病、以及抗病毒治疗上有着广泛的应用前景。正常生理中,t细胞识别肿瘤细胞首先需要mhc-i类分子提呈肿瘤细胞表面特异标记作为第一信号,进而需要cd28等共刺激作为第二信号,才能够有效激活t细胞对肿瘤细胞进行杀灭。此条件往往无法有效激活免疫t细胞,从而极易造成肿瘤细胞的免疫逃逸。car-t细胞技术是将能识别某种肿瘤抗原的抗体与cd3ζ链或者fcεriγ的胞内部分结合形成融合基因,再通过基因转导的方法转染患者自身的t细胞,使其表达嵌合抗原受体(car)。患者的t细胞被转导后,将生成大量的肿瘤特异性t细胞,即car-t细胞,来对抗肿瘤细胞。
4.第一代car由识别肿瘤表面抗原的单链抗体和免疫受体酪氨酸活化序列(cd3ζ链或fcεriγ)组成。第一代car由于缺少t细胞活化的共刺激信号,无法很有效的模拟内源性t细胞激活过程,因此只能引起短暂的t细胞增殖和较低水平的细胞因子分泌,不能提供长期持久的t细胞扩增和抗肿瘤效应。为了更有效的模拟内源性t细胞的激活过程,随之开发的第二和第三代car在设计上引入了共刺激分子的信号序列,比如cd28和4-1bb。这种设计的car-t细胞在体内的增值性,存活时间,细胞因子释放能力以及细胞毒性都有了很大提高,可以产生体内持久且有效的肿瘤细胞清除效果。
5.尽管car-t技术已经在临床取得了很好的疗效,但可控性成为其主要应用问题。car-t细胞在体内一旦与靶细胞接触,就会引起明显的细胞毒效果及大量的细胞因子分泌,表现为高热、高烧、神经紊乱等副反应现象,严重时将危及生命。如何有效的对car-t细胞进行激活和关闭的调控,是该领域亟需解决的关键问题。现有的研究主要集中在以下三方面:(1)自杀基因的引入。通过自杀基因的引入,可以很有效的对car-t进行关闭。现有的自杀基因包括hsv-tk基因,caspase9基因等,通过将这些基因于car基因一同引入t细胞,构建可小分子诱导凋亡的car-t细胞,当t细胞反应过度时,通过诱导剂的加入,将即时杀灭这些过度活化的car-t细胞,减少毒副作用(brown et al.,2014);(2)开关连接物的引入。通过设计中间连接臂,使car-t细胞识别连接臂的一端,而连接臂的另一端识别靶细胞(cartellieri et al.,2016;rodgers et al.,2016;tamada et al.,2012;urbanska et al.,2012)。这种设计下的car-t细胞,其激活将依赖与连接臂的加入,并成剂量依赖性性,可以很有效的通过中间连接臂对car-t细胞进行激活方面的调控;(3)抗体的引入。用特异性抗体封闭特定细胞因子,比如il-6和il-1,在一些临床案例中可以很有效的对抗car-t细胞带来的细胞因
子释放综合征(cytokine release syndrome,crs)。
6.在实际应用中,理想的car-t细胞调控模式应该包括有效性、即时性、和可逆性。上述的引入自杀基因的方法,虽然可以有效的对car-t细胞进行关闭,但是不可逆的细胞损耗也会对治疗效果带来不可避免的损伤,因此限制了其临床应用;另一方面,如上所述的开关连接物的引入,虽然可以很有效的控制car-t细胞的激活,但是car-t细胞的关闭则依赖于开关化合物的代谢。在有靶细胞存在的情况下,开关化合物会优先结合靶细胞,进而延迟其在体内的代谢,而基于抗体的开关化合物本身也有较长的半衰期。因此,此方法虽然可以很有效的控制car-t细胞激活,但是不能对car-t细胞进行即时有效的关闭作用;相比于上述两种方法,基于抗体的控制方法在临床上更受青睐。然而car-t细胞所导致的的crs效应为多因子共同释放所引起,因此单一控制一种因子的作用并未能达到系统性的crs抑制效果。综上,尽管已有的研究可以一定程度的解决car-t的控制问题,但都仅对其一方面进行了改进(关闭或激活),仍未能达到对car-t细胞进行有效、即时且可逆的控制效果。而目前也缺少能有效对car-t细胞达到上述调节效果的技术理论方法。
技术实现要素:
7.为了解决上述问题,本发明提供一种用于控制car-t激活/抑制的连接臂。
8.本发明提供的一种用于控制car-t激活/抑制的连接臂,其一端含有可识别靶细胞的靶头分子,另一端含有可被特异性car-t细胞所识别的具有生物正交性的部分,中间用可被生物正交断裂的化学基团偶联。
9.在本发明的一个实施方式中,所述的靶头分子可识别靶细胞。优选的,所述的可识别靶细胞的靶头分子为具有细胞靶向性的大分子或小分子化合物。更优选的,所述靶头分子包括但不限于叶酸,抗体,抗体片段,以及具有肿瘤靶向性的多糖,肽等物质。α-叶酸受体表达在约50%的肿瘤细胞中,而尤其高表达在乳腺、肺、子宫以及脑癌相关细胞中,但是较少表达在正常组织中。而基于肿瘤标志物的筛查,例如针对乳腺癌细胞的特异性her2抗体及其片段;结直肠癌、头颈鳞癌等细胞的egfr的抗体及抗体片段;以及针对b细胞表面特异性cd19,cd20的抗体及其片段等。
10.在本发明的一个实施方案中,所述连接臂含有可被特异性car-t细胞所识别的具有生物正交性的部分,并与car-t细胞的胞外区域形成特异性正交对。异硫氰酸荧光素(fluorescein isothiocyanate,fitc)及其特异性抗体已被证明在生物体内有良好的特异性和正交性,此外,生物素/亲和素或生物素抗体,叠氮/叠氮特异性抗体或炔类化合物,硝基苯/硝基苯特异性抗体,或其余在生物体系内具有良好特异性和正交性的反应对均可作为连接臂分子与t细胞的特异性识别区。所述部分能特异性的被所构建t细胞的胞外区正交识别,而所述的特异性t细胞也可以编码所识别部分的抗体或其余亲和性片段。所述部分包含但不限于fitc,生物素,叠氮,硝基苯的小分子以及小肽、蛋白、核酸或多糖的生物大分子。对应的,所述t细胞包括但也不限于特异性识别fitc或其它生物正交性部分的抗体,抗体片段,以及其它具有特异性亲和力的分子。
11.在本发明一个实施方案中,所述的可被断裂的化学基团为任何可人为控制诱导产生断裂的化学基团。可被断裂的化学基团存在多种原理的断裂反应,其可被不同波长的紫外、红外、蓝光等激发断裂;也可被小分子、酸碱ph改变、温度改变等诱导断裂。也可被超声、
核磁、放射线等诱导断裂。如邻硝基苄基酯,其在紫外光365nm下,基团内两分子间化学键会发生断裂,进而分离断裂基团两边的化学基团。虽然本发明使用可被断裂的化学基团为邻硝基苄基酯,但本发明所述的可断裂的化学基团原理包括但不限于可紫外断裂、红外断裂、蓝光照射断裂、以及小分子断裂等;所述具体可断裂化学基团包括但也不限于紫外光断裂的邻硝基苄基酯,四嗪类诱导断裂的反式环辛烯等。所述具体可断裂化学基团包括但也不限于紫外光断裂的邻硝基苄基酯,四嗪类诱导断裂的反式环辛烯等。
12.本发明还提供含有所述的用于控制car-t激活/抑制的连接臂的治疗药物。
13.本发明还提供所述的用于控制car-t激活/抑制的连接臂在制备治疗疾病药物中的用途。
14.本发明还提供一种控制car-t激活/抑制的方法,其为通过所述的连接臂偶联car-t细胞和靶细胞,进而对car-t细胞进行激活;通过可控的断裂反应,使car-t细胞与靶细胞进行即时有效的断裂,进而抑制car-t细胞的副作用。
15.本发明通过将生物正交断裂技术引入到car-t的设计和应用中,合成含有可正交断裂基团的双功能连接臂,用以提高car-t细胞控制中的灵活性和有效性,进而实现car-t细胞可“开关”控制。设计的双功能连接臂其一端含有靶头分子可以识别靶细胞,比如抗体分子或叶酸等小分子靶头;另一端设计含有具有生物正交性的基团用于被特异性car-t细胞所识别,比如异硫氰酸荧光素(fitc)或者生物素;中间用可被生物正交断裂的化学基团偶联,比如可紫外断裂的基团或可小分子断裂的基团。如此,car-t的激活将依赖于连接分子的加入,car-t的关闭也可被特定的化学反应所终止。
16.相比于其它方法,本发明的优点可体现在如下中的一个或几个:
17.调节具有即时、高效、且灵活的特点,可以实现car-t细胞从激活到静息状态的快速且灵活的转换;
18.对car-t细胞的抑制具有系统性和高效性。直接分离car-t细胞与靶细胞,进而对car-t细胞接触产生的激活效应和分泌的细胞因子进行系统性的抑制。
19.调节具有可逆性,不损害或杀灭已有的car-t细胞,可以实现car-t细胞从激活到关闭再到激活的灵活转变,不影响治疗的整体作用;
20.借由即时、高效、可逆的特点,可实现对car-t细胞的“开关”型理想调控模式。
附图说明
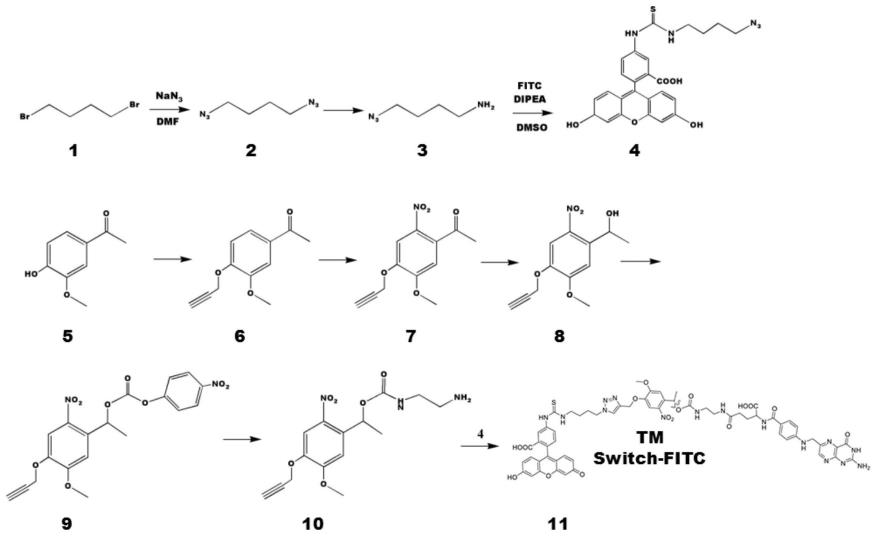
21.图1所示为folate-nbe-fitc合成路线图。
22.图2所示为folate-nbe-fitc靶向性验证。a.fr阳性细胞株筛选;b.folate-nbe-fitc靶向fr高表达细胞株验证。
23.图3所示为anti-fitc car-t细胞构建。a.第二代car-t慢病毒载体构建;b.car结构表达验证;c.car-t特异性识别folate-nbe-fitc验证。
24.图4所示为folate-nbe-fitc细胞水平上断裂效果验证。
25.图5所示为folate-nbe-fitc细胞水平上断裂条件优化。a.断裂效率随时间的条件优化,上为kb细胞,下为car-t细胞;b.台盼蓝法检测光照时间对细胞活力影响。
26.图6所示为folate-nbe-fitc介导的car-t细胞与kb细胞的结合与解离。a.结合与解离的流式细胞术结果;b.双阳性细胞簇的荧光图像及明场图像。
27.图7所示为folate-nbe-fitc介导的car-t细胞激活。a.car-t细胞可被folate-nbe-fitc剂量依赖性激活;b.car-t细胞激活表面标记物检测;c.folate-nbe-fitc介导的car-t细胞ifn-γ(左)以及il-2(右)释放检测;d.folate-nbe-fitc介导的car-t细胞聚集检测。
28.图8所示为folate-nbe-fitc介导的car-t细胞体外活性调控。a.模型设计流程图;b.0-31小时细胞毒效应曲线图;c.0-31小时il-2累积量趋势图;d.0-31小时ifn-γ累积量趋势图;e.folate-nbe-fitc加入/断裂介导的car-t细胞聚集效应检测。
29.图9所示为弥散瘤小鼠中folate-nbe-fitc介导的car-t细胞体内抗肿瘤效应。
30.图10所示为弥散瘤小鼠中folate-nbe-fitc介导的car-t细胞体内抗肿瘤效应。a.模型设计流程图;b.小鼠生物发光成像结果;c.小鼠荷瘤体积随时间趋势变化图;d.紫外透皮断裂folate-nbe-fitc分子验证。
具体实施方式
31.以下实施例用于说明本发明,但不用来限制本发明的范围。
32.实施例1
33.合成了基于fitc、叶酸、以及邻硝基苄基酯的可控断裂的双功能连接臂(folate-nbe-fitc),其主要包括三部分:(1)fitc作为连接配体,与特异性识别fitc的car-t细胞结合。fitc作为荧光分子,已经证明在体内有很好的生物正交性以及生物兼容性;(2)叶酸作为靶头部分,靶向叶酸高表达肿瘤细胞。现有的结果表明,α型叶酸受体充分表达在约50%的肿瘤细胞上,而很少表达在正常组织中,因此叶酸分子可作为理想的靶头用于模型的验证;(3)紫外光可控断裂的邻硝基苄基酯(ortho-nitrobenzylic ester group,nbe)。该基团可在生物兼容的紫外光照条件下(uv365nm),发生迅速且高效的断裂,方便模型的验证。
34.如图1所示,取200ml圆底烧瓶,加入1,4-二溴丁烷(21.6g,10mmol,1eq),加入50mldmf,再加入叠氮钠|(16.5g,21mmol,2.1eq溶于25ml水中)。然后将反应转移到80℃反应20小时。反应结束后使用200ml盐水稀释并用正己烷萃取(200ml
×
3),然后有机相使用无水硫酸钠干燥。蒸干溶剂得到2(1,4-二叠氮丁烷)(13.72g 98%)。1h nmr(400mhz,cdcl3)δ3.30(t,j=6.8hz,2h),2.74(t,j=6.7hz,2h),1.70
–
1.60(m,2h),1.53(dt,j=14.2,7.0hz,2h).
35.取200ml圆底烧瓶加入1,4-二叠氮丁烷(4.2g,30mmol,1eq),加入1m hcl(60ml),乙醚(20ml),乙酸乙酯(20ml),然后冷却到0℃在一个小时时间段内缓慢加入三苯基膦(7.86g,30mmol,1eq)。然后将反应转移到21℃搅拌20小时。反应结束后分离水相与有机相,水相使用乙醚洗涤(100ml
×
3)以除去生成的三苯氧磷,然后水相使用新配置的氢氧化钠水溶液调节ph=13,然后使用二氯甲烷萃取(200ml
×
3),然后有机相使用无水硫酸镁干燥,蒸发旋干得4-叠氮丁胺淡黄色油状液体3(2.74g,24mmol,80%)。1h nmr(400mhz,cdcl3)δ3.29(s,2h),1.64(s,2h).
36.取25ml的圆底烧瓶,加入异硫氰荧光素(100mg,0.25mmol,1eq)5mldmso,,4-叠氮丁胺(34.2mg,0.3mmol,1.2eq)dipea(64.63mg,0.5mmol,2eq)。然后室温搅拌6小时,tlc监测,反应结束后旋干dmso,然后通过柱层析得到纯净的化合物亮黄色固体4(40%)
37.取50ml烧瓶称4-甲氧基-3-羟基苯乙酮(3.32g,20mmol,1eq),加入溶剂dmf 15ml,
加入碳酸钾(2.76g,20mmol,1eq)溴丙炔(2.38g,20mmol,1eq)然后室温反应1小时。反应结束后蒸干溶剂旋干然后用二氯甲烷萃取(200ml
×
3),有机相使用无水硫酸钠干燥,蒸发旋干的白色固体收率6(4.0g 98%)
38.取100ml圆底烧瓶,加入10ml的浓硝酸,然后冰浴下加入5ml乙酸酐并搅拌30min.向该溶液中缓慢加入溶于2.5ml乙酸酐中的6(2.17g,12mmol)。将反应再搅拌5小时,然后倒入200毫升冰水中。然后使用二氯甲烷萃取3次,所得有机溶液用无水硫酸钠干燥,真空浓缩。用乙醇重结晶,得到产物7(1.63g,61%)。
39.取50ml烧瓶,称取7(1.2g,4.9mmol)加入thf(10ml)溶液中和甲醇(20ml),然后将反应转移到冰浴,缓慢将四氢硼酸钠(1.2g,34mmol)加入到上述溶液中搅拌3小时。反应结束后真空浓缩除去溶剂,然后加入h2o(50ml)和2m加入hcl(5ml),溶液用氯仿萃取氯(3
×
100ml)。有机相使用无水硫酸钠干燥,减压下除去溶剂得到(99%)淡黄色固体8。
40.取100ml的圆底烧瓶,称取化合物8(1.25g,5mmol),然后加入吡啶(5ml),转移至冰浴下加入氯甲酸4-硝基苯酯(1.2g,6mmol)。然后将反应转移到室温并搅拌持续6小时,然后用nahco3(10ml)淬灭。该反应混合物用乙酸乙酯萃取,洗涤有机层用盐水(10ml)洗涤,用mgso4干燥,真空浓缩。然后通过柱层析硅胶纯化,得到9(1.35g,65%)。
41.取干燥的50ml的圆底烧瓶,加入乙二胺(180mg,3mmol)和dipea(623ul,4mmol)然后称取去9(832mg,2mmol)溶于5ml二氯甲烷中。在室内搅拌后温度过夜,将反应混合物真空浓缩至干燥。使用硅胶色谱法(etoac/己烷=1/1)纯化残余物得到黄色油状物10(573mg,84%)
42.取干燥的50ml圆底烧瓶,称取叶酸(300mg,0.68mmol,1eq)氮气保护下加入干燥的dmso溶剂,然后加入nhs(86mg,0.748mmol,1.1eq)dcc(154.22mg,0.748mmol,1.1eq)反应在条件下搅拌4小时。反应结束后过滤,除去生成的dcu,然后向滤液中加入10(229.16mg,0.68mmol,1eq)氮气保护下避光搅拌28h。反应结束后将反应液逐滴滴入实现配置好的冷却到0℃的乙醚/丙酮=7/3的溶液中,析出橙黄色沉淀,然后过滤并使用二氯甲烷洗涤三次以除去未反应的10,再用蒸馏水洗涤两次,然后干燥。干燥完全以后放入圆底烧瓶中,加入干燥的dmso10ml溶解,然后加入1(343.4mg,0.68mmol,1eq),五水硫酸铜(17mg,0.068mmol,0.1eq)抗坏血酸钠(53.85mg,0.272,0.4eq),氮气保护下反应两天。反应结束后将将反应液逐滴滴入实现配置好的冷却到0度的乙醚/丙酮=7/3的溶液中,使用二氯甲烷洗涤三次以除去未反应的1,再用蒸馏水洗涤两次,然后通过hplc纯化,得11。
43.对folate-nbe-fitc的靶向性进行了验证。首先用流式细胞术,筛选了叶酸受体(fr)高表达肿瘤细胞株kb和skov3,以及fr低表达细胞系a549和4t1(图2-a)。随后将folate-nbe-fitc与细胞共孵育,清洗后用识别fitc分子的apc荧光标记二抗再进行标记,通过流式细胞术,可以检测到folate-nbe-fitc特异性的识别了fr高表达肿瘤细胞株,而很少,甚至不结合fr低表达细胞株(图2-b),证明了folate-nbe-fitc具有靶向性。另一方面,也证明了在folate-nbe-fitc分子结合靶细胞后,fitc分子暴露在外,可以结合抗体以及car-t细胞。
44.实施例2
45.构建了识别fitc分子的car-t细胞(anti-fitc-car-t)。通过将识别fitc分子的单链抗体基因序列与cd8铰链区和跨膜区,4-1bb以及cd3ζ区融合(car结构),并将构建好的基
因片段构建入慢病毒载体中(图3-a)。并通过包装出来的慢病毒载体感染人cd3 原代t细胞,使cd3 t细胞表达car结构,进而得到特异性识别fitc分子且能被其特异性激活的car-t细胞。我们用apc标记的anti-mouse-fab抗体对细胞表面的car结构表达进行了检测(图3-b),并通过小分子与anti-mouse-fab-apc抗体双标记的方法,确认了anti-fitc-scfv在t细胞表面得到了正确的折叠,成功制备了特异性识别folate-nbe-fitc的car-t细胞(图3-c)。
46.对folate-nbe-fitc在car-t细胞和靶细胞水平上的可断裂性进行了验证,确认其可以通过365nm的紫外光照,在kb细胞(图4-a)以及car-t细胞(图4-b)上进行有效的断裂。
47.通过对folate-nbe-fitc在细胞水平的断裂条件上进行优化,得到了在细胞水平上断裂连接臂用以关闭car-t细胞活性的最佳条件。通过用不同光强度,不同光照时间在car-t细胞与kb细胞上进行光照断裂条件的摸索(图5),可以确认在低紫外功率下(毫瓦级别),短时间内(car-t细胞10分钟,kb细胞20分钟),就可在细胞水平上实现folate-nbe-fitc快速且高效的断裂,且反应具有生物兼容性。
48.验证了基于可断裂的小分子开关,可以介导car-t细胞与靶细胞的结合与解离。通过将apc标记的car-t细胞与pe标记的kb细胞共孵育,并通过成像流式系统,直观的展现了car-t细胞与kb细胞可以借由folate-nbe-fitc的加入实现结合;并通过光照断裂folate-nbe-fitc,使两种细胞进行了解离(图6-a)。通过对细胞簇进行荧光分析,可以清晰的看到car-t细胞与kb细胞结合,并且借由光照实现解离的图像(图6-b)。
49.实施例3
50.将car-t细胞与kb细胞共孵育,通过加入不同浓度的folate-nbe-fitc,验证了car-t细胞的细胞毒作用可以被folate-nbe-fitc有效的激活,并成剂量依赖效应,ec50约等于85pm(图7-a);t细胞激活表面标记物cd25与cd69也随folate-nbe-fitc的加入而得到明显的增加(图7-b);细胞因子释放水平与细胞毒作用一致,都可以被folate-nbe-fitc明显激活,并呈现剂量依赖效应(图7-c);另外,通过显微镜,可以明显的观察到只有car-t细胞与kb细胞共孵育的实验组呈现了明显的t细胞聚团效应,而其余对照组均无明显现象。此方案证明了folate-nbe-fitc可以介导car-t细胞有效的激活,且具有剂量依赖效应。
51.实施例4
52.从体外的细胞实验中,验证了基于可断裂开关化合物,可以对car-t进行“开关模式”的调节控制。将car-t细胞和靶细胞共孵育,按如图所示的操作流程进行folate-nbe-fitc介导的car-t调控研究(图8-a)。结果表明,0-4小时中,folate-nbe-fitc可以有效的介导car-t细胞杀伤靶细胞;而在第4小时进行断键反应后,成功对car-t细胞进行了“关闭”效应,使得car-t细胞的细胞毒作用得到了明显的抑制;而在第21小时重新添加folate-nbe-fitc后,又成功的再激活了car-t细胞,使其细胞毒作用回复到了正常水平(图8-b)。il-2以及ifn-γ释放水平与细胞毒效应的结果一致(图8-c,d);另一方面,在实验过程中,通过显微镜直接对各实验组进行观察,可以发现car-t细胞的聚团效应也与folate-nbe-fitc的加入-断裂-再加入一致(图8-e)。综上,此结果证明了folate-nbe-fitc分子可以有效的介导的car-t的“开关”模式活性调控。
53.实施例5
54.通过小鼠的弥散瘤模型,验证了car-t细胞在体内的激活可以有效的依赖于folate-nbe-fitc的加入,并呈现剂量依赖效应(图9)。
55.从小鼠的皮下荷瘤模型中,验证了基于可断裂开关化合物,可以对car-t从体内进行“开关模式”的调节控制。具体实验流程如图10-a所示。在皮下荷瘤小鼠的模型实验中,可以观察到,相对于不添加folate-nbe-fitc的对照组(组1),添加folate-nbe-fitc可以有效的对肿瘤生长进行抑制(组2);而通过照射对folate-nbe-fitc进行断裂后,由于car-t细胞的效应受到明显的抑制,导致肿瘤细胞的生长在第19天最后一个监测点达到对照组水平(组3);特别的,对第4组,在断裂后重新补加folate-nbe-fitc,可以明显的观察到car-t细胞的活性得到了再激活,又重新恢复了对肿瘤细胞的抑制效应(图10-b)。通过隔天测量肿瘤细胞的体积,得到的荷瘤体积大小随时间的变化图,可以观察到各组间随folate-nbe-fitc的加入-断裂-再加入的趋势变化与动物成像数据相一致(图10-c)。另一方面,通过体外的透皮实验,验证了uv365nm的紫外光可以穿透表皮,对folate-nbe-fitc进行细胞水平上的断裂,证明了上述调控现象是基于断裂反应而得到的实现(图10-d)。
56.以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明技术原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。