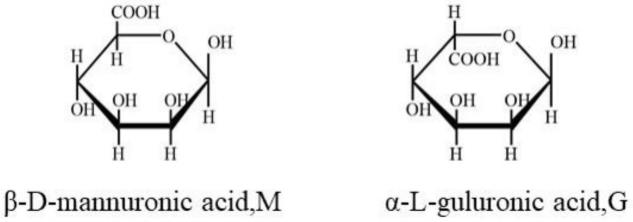
重组甘露糖醛酸c-5差向异构酶及其编码基因和制备方法与应用
技术领域
1.本发明涉及重组甘露糖醛酸c-5差向异构酶及其编码基因和制备方法与应用,属于基因工程领域。
背景技术:
2.褐藻多糖是由β-d-甘露糖醛酸(mannuronate,m)和它的c-5差向异构体α-l-古罗糖醛酸(guluronate,g)两种单体组成的线性聚合物。在褐藻多糖中m与g的存在形式有3种:聚甘露糖醛酸片段(poly-mannuronate,pm)、聚古罗糖醛酸片段(poly-guluronate,pg)和甘露糖醛酸-古罗糖醛酸混合嵌段(mg block)。如下结构式所示:
[0003][0004]
不同来源的褐藻多糖不仅分子间m/g比值不同,m、g的排列顺序也不同,结构上的差异导致褐藻多糖的凝胶强度和持水力等理化性质也存在较大差异。研究发现,褐藻多糖中m/g值越大,其凝胶强度越小,而其持水力越强。褐藻多糖中的m/g比决定其生物活性,如富含m的褐藻多糖和寡糖具有抗肿瘤、免疫调节等作用,而富含g的褐藻多糖具有抗艾滋病等重要应用价值。目前在食品、医药等多个领域,都有对不同性质(不同m/g比值)的褐藻多糖的需求,尤其是对高g含量的褐藻多糖需求较大,这是由于大多数藻类的m/g值为1.2-1.6,因而对m/g比值较低的0.5-0.7的褐藻多糖的需求量更大,目前主要依赖于从g含量高的褐藻中进行提取获得,m/g值为0.4的褐藻多糖仅存在于海带(laminariahyperborea)和螺纹雷松藻(lessoniatrabeculata)的叶柄中,而整个海带植物中褐藻多糖的m/g值约为1.0,从中进行叶柄的分离耗时且费力。目前自然界中g含量高的褐藻不仅存在储量低的不足,而且也会随着各种环境条件发生变化,因而不能提供稳定的高g含量的褐藻多糖,极大地限制了其在各个领域的应用。针对此,开发有效的方法对褐藻多糖中的m构型进行异构成为获得高g含量褐藻多糖的可能途径,化学方法可以实现m到g的异构,但是通过化学方法进行异构化反应存在一些弊端,针对多糖底物异构化过程比较困难,通常需要较高的反应温度,会导致多糖的解聚,同时由于化学试剂的引入,后期还需要进行繁杂耗时的分离纯化工作。
[0005]
研究发现自然界中存在可以将β-d-甘露糖醛酸(mannuronate,m)转化为它的c-5差向异构体α-l-古罗糖醛酸(guluronate,g)的酶,这类酶被命名为甘露糖醛酸c-5差向异构酶(mannuronan c-5epimerase),一般来源于海藻、假单胞菌属以及棕色固氮菌属。在大肠杆菌中异源表达的来源于褐藻中的甘露糖醛酸c-5差向异构酶manc5-e,分子量75kd,具
有非常低的体外异构酶活性。作用于m/g为1.5和1.7的褐藻多糖,可分别将其m/g值降至1.3、1.5,g百分含量提高约3%。来源于棕色固氮菌(azotobactervinelandii)的甘露糖醛酸c-5差向异构酶alge,分子量为103kd,可实现多聚物m到g的催化转化,产物中g构型处于随机分布的状态,发挥作用时依赖于ca
2
的存在。同样是来源于该菌株的甘露糖醛酸c-5差向异构酶alg2e,其重组表达菌株可以对g起始含量为0-45%的褐藻胶变位至g含量达70%。然而该酶对mg交替结构变位能力相对较弱,该酶的活力也会受到钙离子的影响,同时该酶还具有一定的裂解酶活性。该菌株中的alge7甘露糖醛酸c-5差向异构酶同时具有差向异构和裂解两种活性,在降解富含m的褐藻多糖时,可产生聚合度4到7的寡糖。研究发现该酶的异构酶活性和裂解酶活性在同一活性位点。棕色固氮菌中的另一个甘露糖醛酸c-5差向异构酶algg是一个定位于周质的蛋白,分子量58kd,其差向异构酶活性可以不依赖于ca
2
,然而添加钙离子也会适度刺激酶活性。比较特殊的是,该酶的产物只有mg片段,而且该酶的异构化活性会被锌离子阻断。来源于铜绿假单胞菌(pseudomonas aeruginosa)的甘露糖醛酸c-5差向异构酶也定位于胞质外,可以将最小单元为9碳糖的褐藻胶寡糖进行m到g的催化变位,且酶反应过程同样不需要钙离子的参与,最终产物古洛糖醛酸单体的含量可达到75%。利用酶进行差向异构反应,具有方法简单、易控制,后期分离容易的优点,且酶作为一种生物制剂,不会带来化学试剂产生的不利影响。然而目前这类酶的相关报道较少,而且由于酶的活性较低,还处于实验室阶段,实际将其应用于褐藻多糖g含量的调控尚属空白。
技术实现要素:
[0006]
本发明提供了2个来源于门多萨假单胞菌(pseudomonas mendocina)甘露糖醛酸c-5差向异构酶突变体,和野生型相比,突变体的裂解酶活性大幅降低甚至丧失,主要保留了异构酶活性,而且异构酶活性还有不同程度的提高,可应用于多糖的异构化反应,本发明所提供的异构酶突变体,分子量小,易于表达,研究发现可将底物的g含量从32%提高至67%。
[0007]
本发明的第一个目的是提供两个来源于门多萨假单胞菌(pseudomonas mendocina)的甘露糖醛酸c-5差向异构酶pmc5a的突变体及其编码基因pmc5a
238n-s
和pmc5a
348s-n
基因。
[0008]
本发明的第二个目的是提供含有所述的甘露糖醛酸c-5差向异构酶pmc5a突变基因重组表达质粒和重组基因工程菌株。
[0009]
本发明的第三个目的是提供一种制备甘露糖醛酸c-5差向异构酶pmc5a突变体的方法。
[0010]
本发明的第四个目的是提供一种甘露糖醛酸c-5差向异构酶pmc5a突变体在差向甘露糖醛酸c-5键中的应用。
[0011]
本发明所提供的重组甘露糖醛酸c-5差向异构酶基因突变体pmc5a
238n-s
和pmc5a
348s-n
,是由来源于海胆肠胃中分离纯化的门多萨假单胞菌pseudomonas mendocina的甘露糖醛酸c-5差向异构酶pmc5a突变而来;
[0012]
所述pmc5a
238n-s
的核苷酸序列具有如下1a)~3a)特征中的一种、两种或两种以上:
[0013]
1a)具有seq id no.1的脱氧核糖核酸(dna)序列;
[0014]
2a)编码seq id no.2氨基酸序列的脱氧核糖核酸(dna)序列;
[0015]
3a)对seq id no.1的脱氧核糖核酸(dna)序列除第712-714位点外,进行一个、两个或两个以上核苷酸取代和/或缺失和/或添加而得到的编码仅具有甘露糖醛酸c-5差向异构酶活性的核苷酸序列;
[0016]
所述pmc5a
348s-n
的核苷酸序列具有如下1b)~3b)特征中的一种、两种或两种以上:
[0017]
1b)具有seq id no.3的脱氧核糖核酸序列;
[0018]
2b)编码seq id no.4氨基酸序列的脱氧核糖核酸序列;
[0019]
3b)对seq id no.3的脱氧核糖核酸序列除第1042-1044位进行一个、两个或两个以上核苷酸取代和/或缺失和/或添加而得到的编码仅具有甘露糖醛酸c-5差向异构酶活性的核苷酸序列。
[0020]
本发明还提供了重组甘露糖醛酸c-5差向异构酶的氨基酸序列。
[0021]
pmc5a
238n-s
编码的氨基酸序列具有如下1c)~3c)特征中的一种、两种或两种以上:
[0022]
1c)具有seq id no.2的氨基酸序列;
[0023]
2c)具有如seq id no.2从氨基端开始的第1-475位氨基酸残基序列;
[0024]
3c)对seq id no.2所示的氨基端序列除第238位外,进行一个、两个或两个以上氨基酸取代和/或缺失和/或添加而形成仅具有甘露糖醛酸c-5差向异构酶活性氨基酸序列。
[0025]
pmc5a
348s-n
编码的氨基酸序列具有如下1d)~3d)特征中的一种、两种或两种以上:
[0026]
1d)具有seq id no.4所示的氨基酸序列;
[0027]
2d)具有如seq id no.4从氨基端开始的第1-475位氨基酸残基序列;
[0028]
3d)对seq id no.4所示的氨基酸序列除第348位外,进行一个、两个或两个以上氨基酸取代和/或缺失和/或添加而形成的仅具有甘露糖醛酸c-5差向异构酶活性的氨基酸序列。
[0029]
本发明提供了一种含有重组甘露糖醛酸c-5差向异构酶突变体基因的重组表达质粒。
[0030]
本发明提供了一种含有重组甘露糖醛酸c-5差向异构酶突变体基因的重组基因工程菌株。
[0031]
本发明提供了一种重组甘露糖醛酸c-5差向异构酶的制备方法,是将重组甘露糖醛酸c-5差向异构酶突变基因克隆入重组表达载体,将重组表达载体导入宿主细胞,获得重组甘露糖醛酸c-5差向异构酶。
[0032]
进一步地,所述的重组表达载体包括大肠杆菌表达载体、酵母表达载体、枯草杆菌表达载体、乳酸菌表达载体、链霉菌表达载体、噬菌体载体、丝状真菌表达载体、植物表达载体、昆虫表达载体、或哺乳动物细胞表达载体中的一种、两种或两种以上。
[0033]
进一步地,所述宿主细胞包括大肠杆菌宿主细胞、酵母菌宿主细胞、枯草杆菌宿主细胞、乳酸菌宿主细胞、放线菌宿主细胞、丝状真菌宿主细胞、昆虫细胞或哺乳动物细胞。
[0034]
所述大肠杆菌宿主细胞包括escherichia coli bl21、escherichia coli jm109或escherichia coli dh5α;所述乳酸菌宿主细胞包括lactic acid bacteria cocc101;所述酵母菌宿主细胞包括saccharomyces cerevisiae、pichiapastoris或kluyveromyceslactis;所述枯草杆菌宿主细胞包括bacillus subtilis r25或bacillus subtilis9920;所述放线菌宿主细胞包括streptomyces spp.;所述丝状真菌宿主细胞包括trichodermaviride、trichodermareesei、aspergillusniger或aspergillusnidulans;所
述昆虫细胞包括bombyxmori或antharaea eucalypti;所述哺乳动物细胞包括中国仓鼠卵巢细胞cho、幼小仓鼠肾脏细胞bhk或中国仓鼠肺细胞chl。
[0035]
本发明的甘露糖醛酸c-5差向异构酶pmc5a突变体的基因序列是通过pcr技术从门多萨假单胞菌中克隆得到。突变体基因编码区长1428bp,包含多个结构域,pmc5a
238n-s
核苷酸序列如seq id no 1所示。pmc5a
348s-n
核苷酸序列如seq id no 3所示,pmc5a突变体编码467个氨基酸和一个终止密码子,pmc5a
238n-s
氨基酸序列如seq id no 2所示,pmc5a
348s-n
氨基酸序列如seq id no 4所示。
[0036]
本发明提供了重组甘露糖醛酸c-5差向异构酶在差向甘露糖醛酸中的应用。
[0037]
进一步地,在差向甘露糖醛酸c-5键,获得不同结构的褐藻胶中的应用。
[0038]
本发明从大肠杆菌重组表达获得的重组甘露糖醛酸c-5差向异构酶突变体,突变体pmc5a
238n-s
虽还同时具有差向异构和裂解两种功能,但是相较于未突变之前,裂解酶活性大幅降低,异构酶活性部分提高。而突变体pmc5a
348s-n
只具有差向异构酶活性,且突变后该活性得到了明显提高,可将海藻多糖底物g含量提高至67%,且表达量提高约1倍。本发明所述重组甘露糖醛酸c-5差向异构酶,能高效的将褐藻多糖中的β-d-甘露糖醛酸(m)转化为α-l-古罗糖醛酸(g),且基本不发生降解作用。本发明提供的甘露糖醛酸c-5差向异构酶可广泛应用于食品、医药等领域。
附图说明
[0039]
图1为甘露糖醛酸c-5差向异构酶pmc5a突变体表达及纯化的sds-page图;图中,泳道m:蛋白分子量标准,泳道1:pmc5a,泳道:2:突变体pmc5a
238n-s
,泳道:3:突变体pmc5a
348s-n
。
[0040]
图2为甘露糖醛酸c-5差向异构酶pmc5a突变体的酶活性比较。
具体实施方式
[0041]
下述非限定性实施例可以使本领域的普通技术人员更全面地理解本发明,但不以任何方式限制本发明。
[0042]
实施例1甘露糖醛酸c-5差向异构酶pmc5a突变体全长基因克隆
[0043]
参照基因组dna纯化试剂盒(thermo,lot 00105781)操作步骤提取门多萨假单胞菌的基因组dna。根据甘露糖醛酸c-5差向异构酶pmc5a基因序列进行比对分析后,设计两对引物如下:
[0044]
seq id no.5:
[0045]
238n-s-f:5
’‑
acgtggtgatcaagggcagcacctaccgcgacaacatcgtctacggc-3’;seq id no.6:
[0046]
238n-s-r:5
’‑
gccgtagacgatgttgtcgcggtaggtgctgcccttgatcaccacgt-3’;seq id no.7:
[0047]
348s-n-f:5
’‑
acggcattcgcatccgcaacagcgagaacattcgcctctacgacaac-3’;seq id no.8:
[0048]
348s-n-r:5
’‑
gttgtcgtagaggcgaatgttctcgctgttgcggatgcgaatgccgt-3’。以提取的门多萨假单胞菌的基因组dna为模板,扩增编码双功能酶成熟蛋白的基因序列(不包括信号肽基因)。pcr反应条件为:94℃2min,1个循环;94℃30s,55℃30s,72℃7min,30个循环;72
℃10min,1个循环。pcr产物进行琼脂糖凝胶电泳分析后,对目的基因进行切胶回收,经双酶切的方法连接到原核表达载体pet21a上后测序。
[0049]
实施例2甘露糖醛酸c-5差向异构酶突变体基因序列分析
[0050]
测序结果采用genbank数据库中的basic local alignment search tool(blast)分析,dnaman软件进行多序列比对,vector nti分析序列信息。
[0051]
获得的两种pmc5a突变体基因(分别命名为pmc5a
238n-s
和pmc5a
348s-n
)编码区长1428bp,pmc5a
238n-s
和pmc5a
348s-n
核苷酸序列分别如seq id no.1和seq id no.3所示。pmc5a突变体编码467个氨基酸和一个终止密码子,pmc5a
238n-s
和pmc5a
348s-n
的氨基酸序列分别如seq id no.2和seq id no.4所示。
[0052]
实施例3 pmc5a突变体基因在大肠杆菌中的重组表达及纯化
[0053]
为了便于基因的重组表达,在设计的上下游引物中分别引入ndei和xhoi酶切位点。将pcr产物pmc5a突变体和表达载体pet21a分别用ndei和xhoi进行双酶切,酶切产物经清洁回收后,用t4dna连接酶连接(连接体系:(5μlt4dna ligase 0.5μl,10
×
t4dna ligase buffer 0.5μl,pet21a 2μl,pcr产物2μl),连接条件:室温过夜连接。)。取5μl连接产物转化e.coli top10感受态细胞,涂布在含100μg/ml氨苄青霉素的固体luria-bertani培养基上,37℃培养12-16h。挑取单克隆,使用简并引物进行菌落pcr验证,将扩增正确的单克隆接入含有100μg/ml氨苄青霉素的液体luria-bertani培养基中培养,提取质粒;使用内切酶ndei和xhoi对提取的质粒进行双酶切,结果正确的重组质粒送华大基因测序。测序结果表明,在pet21a的ndei和xhoi酶切位点之间插入了seq id no 1或seq id no 3所示的突变体基因,且插入方向正确,证明重组质粒构建成功。
[0054]
将重组质粒转化e.coli bl21(de3),对其进行诱导表达及纯化。用聚丙烯酰胺凝胶电泳检测突变体的表达及纯化情况,结果如图1所示,纯化后的突变体在电泳胶上呈单一条带,且位置与预测的分子量相吻合。突变体pmc5a
238n-s
与原蛋白表达量相当,突变体pmc5a
348s-n
表达量提高约1倍。
[0055]
实施例4重组甘露糖醛酸c-5差向异构酶突变体裂解酶活性测定
[0056]
以1000μl 0.5%(w/v)的褐藻胶为底物,加入500μl重组酶pmc5a突变体(pmc5a
238n-s
或pmc5a
348s-n
),反应30min,采用3,5-二硝基水杨酸(dns)法测定其活性。酶活力单位定义为:每分钟释放1μmol还原糖(以甘露糖计)所需的酶量为一个酶活力单位(u)。蛋白浓度使用碧云天bca蛋白浓度测定试剂盒进行测定。
[0057]
如图2所示,在最适温度和最适ph条件下,按标准方法测得pmc5a作为裂解酶的比活为60u/mg;pmc5a
238n-s
裂解酶的比活为6.3u/mg,pmc5a
348s-n
的裂解酶比活为0。
[0058]
实施例5重组甘露糖醛酸c-5差向异构酶突变体的异构酶活性测定
[0059]
将0.5%(w/v)海藻多糖分别与突变体按2:1(体积比)的比例混合后,在30℃条件下反应2h,通过高分辨核磁共振对其产物进行分析。如表1所示,两种突变体(pmc5a
238n-s
或pmc5a
348s-n
)作为差向异构酶可以引入g残基,且突变体pmc5a
238n-s
还同时保留较低的海藻酸钠裂解酶活性。pmc5a可将底物的g含量从32%提高至47%。pmc5a
238n-s
可将底物的g含量从32%提高至50%,pmc5a
348s-n
可将底物的g含量从32%提高至67%。
[0060]
表1.重组甘露糖醛酸c-5差向异构酶突变体的异构酶活性测定
[0061] fgfm
海藻多糖0.320.68海藻多糖 pmc5a0.470.53海藻多糖 pmc5a
238n-s
0.500.50海藻多糖 pmc5a
348s-n
0.670.33
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
