红颜草莓硝酸盐转运蛋白基因fanrt1.1及其应用
技术领域
1.本发明涉及植物基因技术领域,具体涉及红颜草莓硝酸盐转运蛋白基因fanrt1.1及其应用。
背景技术:
2.nrt1.1是唯一被证实的双亲和硝酸盐转运体和假定的硝酸盐受体。拟南芥atnrt1.1是 nrt1家族第一个被发现且功能研究最为清楚的硝酸盐转运蛋白。1993年,tsay首次通过硝态氮类似物氯酸盐的筛选,获得了第一个植物硝酸盐运输蛋白基因chl1(atnrt1.1)突变体;克隆该基因并在爪蟾卵母细胞中异源表达,研究发现,该基因为编码硝酸盐诱导的低亲和运输蛋白基因,并且在细胞低ph时表达量增加,而在铵盐处理时表达量降低。chl1同样具有高亲和硝酸盐运输蛋白的功能特征,后续研究发现,chl1对硝酸盐的亲和力是由该蛋白第101位上thr的磷酸化水平来调控的,磷酸化的chl1对硝酸盐具有高亲和性,去磷酸化后chl1对硝酸盐具有低亲和性。受外界环境中no
3-浓度影响atnrt1.1进行磷酸化的调节,蛋白激酶cipk23可响应低浓度no3-而使atnrt1.1的thr101位点发生磷酸化,用来抑制atnrt1.1的低亲和性no
3-转运活性;而cipk8则正向调控atnrt1.1的低亲和性转运活性。
3.nrt1.1作为硝酸盐受体已被现有研究证实,在许多生物中,nrt1.1硝酸盐受体通常参与营养物质的运输和感知。nrt1.1是硝酸盐受体的证据是通过一个nrt1.1突变体获得的,该突变体阻止硝酸盐摄取,但保留了响应硝酸盐反应的基因表达诱导。nrt1.1可以激活拟南芥根中调控硝酸盐响应基因所需的多种硝酸盐传感和信号机制。nrt1.1也会影响根系发育和形态建成、生长素运输、种子休眠、开花时间和气孔运动等多种生物过程。对nrt1.1基因的研究在拟南芥中已经展开详细研究,在其他植物中的研究也在开展。单子叶植物和真子叶植物的nrt1.1基因数量不同,大多数真子叶植物只有一个nrt1.1基因,而禾本科植物通常有3 到4个nrt1.1成员。现有报道表明atnrt1.1(atnpf6.3)在玉米中的两个假定的同源基因 zmnpf6.4和zmnpf6.6对硝酸盐和氯化物表现出不同的底物亲和力,这表明玉米中中nrt1.1 家族的不同成员之间可能存在功能差异。osnrt1.1a(osnpf6.3)在功能上偏离了以往报道的植物nrt1.1基因,其作用是上调硝酸盐和铵态氮利用相关基因以及花相关基因的表达,参与调控n利用和开花,促进了水稻高产和早熟。osnrt1.1b(osnpf6.5)是atnrt1.1的功能同源物,参与了硝酸盐的利用,该基因的单一多态性导致了亚洲栽培水稻(oryzasativa)籼稻亚种和粳稻亚种间nue的差异。结球白菜中bcnrt1.1基因在地上部分生长点和地下根尖表达,参与侧根发育,受磷酸激酶调控并参与初期硝酸盐反应,可以调控bcnrt2.1表达。
4.鉴于上述缺陷,本发明创作者经过长时间的研究和实践终于获得了本发明。
技术实现要素:
5.本发明的目的在于解决如何对草莓硝酸盐转运蛋白相关基因及其表达与相应模
式进行研究的问题,提供了红颜草莓硝酸盐转运蛋白基因fanrt1.1及其应用。
6.为了实现上述目的,本发明公开了红颜草莓硝酸盐转运蛋白基因fanrt1.1,核苷酸序列如seq id no.1所示。
7.这种红颜草莓硝酸盐转运蛋白基因fanrt1.1所编码的产物的氨基酸序列如seq idno.2所示。
8.本发明还公开了上述红颜草莓硝酸盐转运蛋白基因fanrt1.1通过基因fanrt1.1的不同响应模式在筛选种质材料、培育优质品种和应对非生物胁迫中的应用。
9.与现有技术比较本发明的有益效果在于:本发明通过以n11f和n11r为引物,红颜草莓根的cdna为模板,用lataq酶(takara)体系扩增nrt1.1基因,根据已经获得的红颜草莓fanrt1.1的基因序列,从八倍体栽培草莓卡姆罗莎(camarose)的基因组序列中找到 fanrt1.1同源序列上游的大约2000bp的非编码核苷酸序列,设计带酶切位点的特异性引物,并对这种红颜草莓fanrt1.1进行响应模式分析,根据不同的响应模式可将其应用到筛选种质材料、培育优质品种和应对非生物胁迫中,anrt1.1是硝酸盐信号基因,一种硝酸盐转运蛋白,国内外在草莓中对该基因未有研究,该基因在草莓中首次被发现,具有硝酸盐转运功能,通过建立草莓毛状根诱导体系,上调氮代谢相关基因表达,促进根系硝酸盐的吸收,提高草莓果实硝酸盐含量,促进果实成熟,对提高草莓品质和产量具有重大意义。
附图说明
10.图1为fanrt1.1的电泳检测图;
11.图2为红颜草莓与其它植物nrt1.1间的系统发育关系;
12.图3为fanrt1.1蛋白结构分析;
13.图4为fanrt1.1启动子的电泳检测图;
14.图5为fanrt1.1的时空表达模式;
15.图6为fanrt1.1对不同氮形态的响应模式;
16.图7为fanrt1.1对硝态氮短期诱导的响应模式;
17.图8为fanrt1.1对硝态氮长期处理的响应模式;
18.图9为fanrt1.1对激素的响应模式;
19.图10为fanrt1.1对其他非生物胁迫的响应模式;
20.图11为fanrt1.1亚细胞定位;
21.图12为naa诱导的fanrt1.1组织定位分析。
具体实施方式
22.以下结合附图,对本发明上述的和另外的技术特征和优点作更详细的说明。
23.一、红颜草莓硝酸盐转运蛋白基因fanrt1.1的基因和启动子的克隆与分析。
24.本发明利用软件fpcr和bioxm2.6,根据红颜草莓转录组数据和ncbi比对八倍体草莓基因组数据,设计特异性引物。以n11f和n11r为引物,红颜草莓根的cdna为模板,用 lataq酶(takara)体系扩增nrt1.1基因。
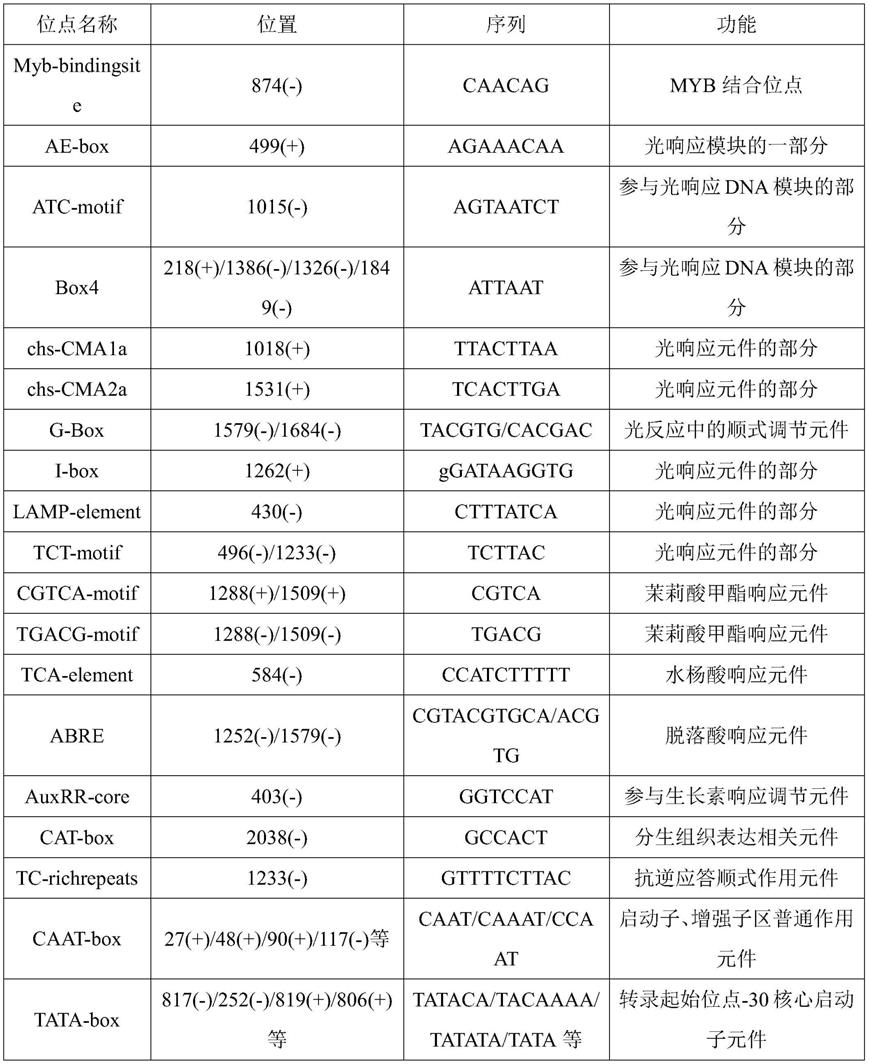
25.将pcr扩增产物进行1%的琼脂糖凝胶电泳检测,切取目的长度条带的胶块,采用天根琼脂糖凝胶dna回收试剂盒(tiangelmidipurificationkit),回收目的片段。将胶回收
的目的片段与pmdtm19-t载体连接,转化大肠杆菌(e.coli)dh5α感受态细胞,amp抗性培养皿上筛选,菌落pcr反应检测。挑取含有目的片段的菌落,重新摇菌,按天根的质粒小提试剂盒(tianprepminiplasmidkit)方法提取质粒,送上海生工生物工程有限公司测序,将菌液和质粒-80℃保存备用。根据草莓转录组数据设计fanrt1.1基因的上下游引物,进行pcr扩增,电泳检测结果如图1所示,经测序发现红颜草莓fanrt1.1的cdna克隆共得到1799bp 的序列。通过与红颜草莓转录组的比对发现fanrt1.1位于草莓基因组内的第5号染色体上。据nrt家族基因的系统发育分析可以发现fanrt1.1(fanpf6.3.3)与拟南芥atnrt1.1 (atnpf6.3)聚类更近,表明红颜草莓克隆到的这个nrt家族基因命名合理,属于 nrt1/ptr(npf)家族。
26.克隆到的红颜草莓nrt1.1与其它植物nrt1.1氨基酸序列进行ncbi (nationalcenterforbiotechnologyinformation)的blast比对分析,并利用mega7.0软件进行数据的聚类比较,采用邻接法构建系统进化树,bootstrap方法重抽样进行1000次,进行聚类分析。
27.利用软件或网站进行生物信息学分析,主要包括:同源序列分析,氨基酸序列分析,跨膜结构域分析,蛋白质二级结构分析等。运用dnaman5.2软件进行序列比对分析;利用 ncbi和bioxm2.6进行序列比对和开放阅读框分析;利用pi/mwtool分析基因序列等电点与分子量;利用tmhmmserverv.2.0工具分析氨基酸序列跨膜区域;利用signalp4.1分析氨基酸序列信号肽;利用smart工具分析蛋白结构域;利用sopma工具进行蛋白质二级结构分析;利用ncbi进行物种同源性比对。
28.克隆到的fanrt1.1与其它植物nrt1.1氨基酸序列进行系统进化分析(如图2所示)发现,与红颜草莓最相近的物种有森林草莓(fragariavesca)和月季(rosachinensis),其次有扁桃(prunusdulcis)、甜樱桃(prunusavium)、毛桃(prunuspersica)、梅花(prunusmume)、苹果(malusdomestica)和白梨(pyrusxbretschneideri),较远的有欧洲栓皮栎(quercussuber)、核桃(juglansregia)、河岸葡萄(vitisriparia)、可可树(theobromacacao)、大麻(cannabissativa)、木豆(cajanuscajan)和拟南芥。虽然不同物种间有明显的变异性,但保守结构域也较多,可见克隆的fanrt1.1在进化过程中保持了一定的同源性。
29.生物信息学分析发现:fanrt1.1含1767bp的完整开放阅读框,编码588个氨基酸;所编码的氨基酸序列等电点为9.08,蛋白质分子量为64617.47;预测有12跨膜螺旋(tmhs),跨膜螺旋中的预测氨基酸数量(aas)为252.29324;氨基酸序列中没有信号肽;fanrt1.1 蛋白结构域预测(如图3)结果显示跨膜区域为46-68、73-92、99-121、146-168、189-211、 215-237、337-359、374-396、417-436、456-478、499-521和541-563,复杂程度较低的区域为575-584;fanrt1.1的蛋白质二级结构分发现α-螺旋占39.63%,延伸链占20.07%,β-转角占8.50%,无规卷曲占31.80%。
30.根据已经获得的红颜草莓fanrt1.1的基因序列,从八倍体栽培草莓卡姆罗莎(camarose) 的基因组序列中找到fanrt1.1同源序列上游的大约2000bp的非编码核苷酸序列,设计带酶切位点的特异性引物。以红颜草莓叶片的dna为模板,以n11pf (cccaagcttaaattaagttctaaacccatgc)和n11pr (ggaattctgccctatatagttttgagcgcc)为引物,进行pcr扩增,克隆fanrt1.1 的启动子。
31.以红颜草莓dna为模板,n11pf和n11pr引物,进行pcr扩增。扩增产物经电泳检测,
发现在2000bp左右出现一条带(图4)。经过克隆转化、测序和序列比对,得到2064bp(不带16bp引物区)的fanrt1.1基因启动子。
32.本发明通过启动子在线分析软件plantcare和place对fanrt1.1启动子调控元件进行预测分析。
33.采用plantcare进行启动子顺式作用元件分析(如表1所示)发现fanrt1.1启动子上存在很多不同功能的顺式作用元件。有重要转录因子myb家族的结合位点myb-bindingsite,有ae-box、atc-motif、g-box和lamp-element等9类与光响应相关的顺式作用元件,有 cgtca-motif、tca-element和abre等激素应答元件(茉莉酸甲酯、水杨酸和脱落酸),也有auxrr-core和cat-box与生长素、分生组织表达相关的元件,有tc-richrepeats的抗逆应答顺式作用元件,还有caat-box和tata-box等启动子核心元件。由此推测,fanrt1.1基因的表达受光照和激素等多种因素调控。
34.表1 fanrt1.1基因启动子plantcare顺式元件分析
[0035][0036]
为了分析一些相关的具体顺式作用元件同时采用place进行分析(如表2)。其中, caatbox1、ccaatbox1、acgtabox、gatabox和gtgantg10等各种数目较多的 box类的元件,具有热休克、叶绿素结合等相关功能;mybcore、mybpzm、myb1at 和mybst1等myb类顺式作用元件,具有mybst1结合位点、水胁迫响应和类黄酮生物合成等相关功能;有12个wrky71os位点,可以结合转录因子wrky71,影响赤霉素信号通路;ltre类的ltrecoreatcor15和ltreatlti78可以响应低温;dre类的 dre2corezmrab17和drecrtcoreat可以响应干旱;cact中的cactftppca1属于磷酸烯醇丙酮酸羧化酶远端顺式调控元件mem1的关键成分;dof中的dofcorezm和 ntbbf1arrolb可以增强细胞质磷酸激酶启动和生长素诱导;abre中的abreratcal 可以ca
2
响应上调;myc中的mycconsensusat可以低温诱导转录;cbf中的cbfhv 参与脱水反应;rootmotif中的rootmotiftapox1属于根特异调控元件。
[0037]
表2 fanrt1.1基因启动子place顺式元件分析
[0038][0039][0040]
二、红颜草莓硝酸盐转运蛋白基因fanrtf2.7的表达模式
[0041]
此外,本发明温室中正常生长的红颜草莓植株,取根(全根,root)、茎(短缩茎,stem)、匍匐茎(stolon)、芽(bud)、叶(幼叶和成熟叶混样,leaf)、花(flower)、果实(成熟果, fruit)和瘦果种子(achene),以faact作为内参基因,采用fanrt2.7定量引物,进行基因的表达模式分析。另外取幼根(youngroot,yr)和成熟根(matureroot,mr),幼叶 (youngleaves,yl)、成熟叶(matureleaves,ml)和老叶(oldleaves,ol)对fanrt1.1 基因进一步进行表达分析。
[0042]
取对照(control,n-deficiency,nd)、硝氮(nitrate,ni)、铵氮(ammonium,ai)和
和rw,用于干旱胁迫响应。取14h光/10h暗的盆栽草莓,置于暗下处理48h作为黑暗处理 (d),恢复光照分别在1h和6h采样,记作l1和l6,进行光响应分析。
[0051]
如图9所示,以喷洒清水作为control,喷洒aba、sa和naa可以极显著提高叶片中 fanrt1.1的基因表达;而在果实中,喷洒ga3和naa可以极显著提高fanrt1.1的基因表达,喷洒aba和sa则可以极显著降低fanrt1.1的基因表达;叶片中fanrt1.1响应激素的程度更显著一些。这些结果可以表明,fanrt1.1的基因表达可能受aba、ga3、sa和naa 等激素的诱导。
[0052]
分离拟南芥原生质体,加入含荧光信号超表达载体pcambia1302-fanrt1.1的质粒,进行聚乙二醇(peg)转染,22℃弱光照条件下培养约18h,采用激光共聚焦显微镜进行拍照和亚细胞定位分析。
[0053]
用含有pcambia1391z-profanrt1.1载体的农杆菌侵染拟南芥,具体侵染方法参考 4.1.9.1所述,采用花序侵染法,侵染后收到的种子用hyg抗性筛选,鉴定阳性植株。提取 dna,以n11pf和n11pr为引物,进行pcr扩增鉴定。确定稳定转基因拟南芥材料,进行 gus染色,观测有颜色的组织。
[0054]
其中为了进一步验证gus染色效果,采用无菌播板的拟南芥,分别采用清水(control)、 0.05%赤霉素(ga3)、0.05%萘乙酸(naa)、0.1mm脱落酸(aba)、0.1mm水杨酸(sa)、 0.1mm茉莉酸甲酯(meja)、0.3mnacl、0.8mmcdcl2和0.05mg/lpbcl2喷洒处理,1d后取相应的拟南芥进行gus染色、脱色和显微镜观察记录不同处理条件下的gus染色情况。
[0055]
采用拟南芥原生质体进行亚细胞定位分析(图11),可以发现fanrt1.1定位在质膜上。
[0056]
用含有pcambia1391z-profanrt1.1载体的农杆菌花序法侵染拟南芥,筛选鉴定出t2 代阳性植株,对拟南芥植株进行组织化学染色。一般正常生长的拟南芥所有组织都没有观察到可以被gus染色的部分(如图12a-b);当用0.05%naa喷施处理拟南芥1d后(如图12c-d),可以观察到拟南芥的根系和叶脉可以被gus染色。可以表明,naa诱导条件下fanrt1.1 可以表达,且在根中表达量最高。
[0057]
以上所述仅为本发明的较佳实施例,对本发明而言仅仅是说明性的,而非限制性的。本专业技术人员理解,在本发明权利要求所限定的精神和范围内可对其进行许多改变,修改,甚至等效,但都将落入本发明的保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
