红颜草莓硝酸盐转运蛋白基因fanrt2.7及其应用
技术领域
1.本发明涉及植物基因技术领域,具体涉及红颜草莓硝酸盐转运蛋白基因fanrt2.7及其应用。
背景技术:
2.红颜草莓(fragaria x ananassaduch.
‘
benihoppe’)是日本新品种,植株长势强、叶片大、新茎分枝多,株态直立易于栽培管理。红颜草莓又称红颊,是日本静冈县用章姬与幸香杂交育成的早熟栽培品种良种。
3.nrt2.7是一种种子特异表达的高亲和性no
3-的转运蛋白。拟南芥nrt2.7在种子中特别表达,尤其在种子成熟晚期该基因高度表达,定位于生殖器官液泡膜上,负责将no
3-从胞质向液泡运输,进而调控种子中no
3-含量和休眠。据钱瑜等的研究,二穗短柄草、水稻、高粱、大白菜、拟南芥中都有nrt2.7同源基因,但是大豆、杨树、葡萄、玉米中没有nrt2.7同源基因。在多个物种中发现了nrt2.7基因的同源序列存在丢失的情况,可以说明在一些物种中可能nrt2.7的转运功能也已丢失;但是对已有的单、双子叶植物的nrt2.7基因进行分析可以发现其具有非常相似的基因结构,可以表明单、双子叶植物之间可能保留了类似的机制来装载no
3-进入种子和胚胎。另外,现有研究中选取独特的浅褐色种皮黄酮氧化缺失的拟南芥,通过nrt2.7-2等位基因突变体,发现nrt2.7与原花色素的积累和氧化密切相关。
4.硝态氮的吸收和转运是植物氮代谢研究的关键环节。fanrt2.7的研究不仅对于阐明nrt转运蛋白参与草莓氮素代谢调控的分子机理,而且对草莓生产过程中氮素高效利用以及节肥技术栽培,都具有重要的理论研究意义和实际应用价值。
5.鉴于上述缺陷,本发明创作者经过长时间的研究和实践终于获得了本发明。
技术实现要素:
6.本发明的目的在于解决如何阐明明nrt转运蛋白参与草莓氮素代谢调控的分子机理,提高草莓生产过程中氮素利用率以及实现节肥技术栽培的问题,提供了红颜草莓硝酸盐转运蛋白基因fanrt2.7及其应用。
7.为了实现上述目的,本发明公开了红颜草莓硝酸盐转运蛋白基因fanrt2.7,核苷酸序列如seq id no.1所示,这种红颜草莓硝酸盐转运蛋白基因fanrt2.7编码的产物的氨基酸序列如seqidno.2所示。
8.本发明还公开了上述红颜草莓硝酸盐转运蛋白基因fanrt2.7在氮素高效利用、节肥栽培中的应用。
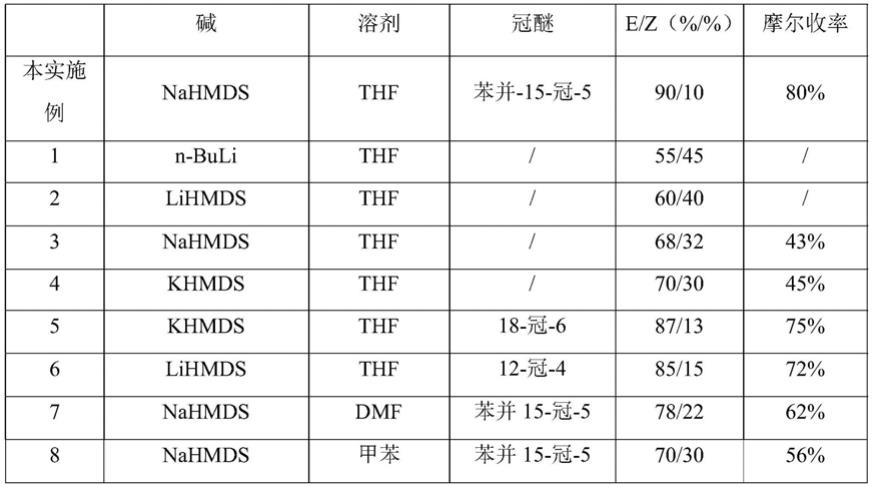
9.与现有技术比较本发明的有益效果在于:本发明以红颜草莓果实cdna为模板,以n27f和n27r为引物,扩增nrt2.7基因,以带酶切位点的特异性引物n27p f和n27p r为引物,克隆fanrt2.7的启动子,对红颜草莓硝酸盐转运蛋白基因fanrtf2.7对不同氮形态、硝态氮、激素和其他非生物胁迫的响应模式分析,并根据响应模式的不同将其应用于筛选种质材料、培育优质品种和应对非生物胁迫,fanrt2.7基因在叶片、瘦果、和花中的表达量较高,
在叶、花、果中响应硝酸盐最显著,fanrt2.7蛋白具有硝酸盐转运的功能,能够提高草莓果实中的硝酸盐含量,并促进草莓果实成熟,对草莓生产过程中氮素高效利用以及节肥技术栽培方面具有较高的应用价值。
附图说明
10.图1为本发明提供的fanrt2.7的电泳检测图;
11.图2为本发明提供的fanrt2.7蛋白结构分析图;
12.图3为本发明提供的红颜草莓与其它植物nrt2.7蛋白之间的系统发育关系图;
13.图4为本发明提供的fanrt2.7的启动子电泳检测图;
14.图5为本发明提供的fanrt2.7的时空表达模式图;
15.图6为本发明提供的fanrt2.7对不同氮形态处理30d的响应模式图;
16.图7为本发明提供的fanrt2.7对不同氮形态处理1h的响应模式图;
17.图8为本发明提供的fanrt2.7对硝态氮短期诱导的响应模式图;
18.图9为本发明提供的fanrt2.7对硝态氮长期处理的响应模式图;
19.图10为本发明提供的fanrt2.7对激素的响应模式图;
20.图11为本发明提供的fanrt2.7对其他非生物胁迫的响应模式图;
21.图12为本发明提供的fanrt2.7亚细胞定位图;
22.图13为本发明提供的fanrt2.7组织定位分析图。
具体实施方式
23.以下结合附图,对本发明上述的和另外的技术特征和优点作更详细的说明。
24.一、红颜草莓硝酸盐转运蛋白基因fanrtf2.7的基因和启动子的克隆与分析。
25.用准备的红颜草莓果实cdna为模板,以n27f和n27r为引物,扩增nrt2.7基因,并进行相关生物信息学分析和系统进化树分析。利用软件或网站进行生物信息学分析发现:fanrt2.7含1416bp的完整开放阅读框,编码471个氨基酸;所编码的氨基酸序列等电点为8.46,蛋白质分子量为50848.21;预测跨膜螺旋(tmhs)为12,跨膜螺旋的预测氨基酸数量(aas)为254.50042;氨基酸序列中没有信号肽;fanrt2.7蛋白结构域预测如图2所示。结果显示跨膜区域为47-69、82-99、119-141、153-175、180-199、204-226、264-286、293-315、335-352、359-381、391-413和425-447,没有复杂程度较低的区域;fanrt2.7的蛋白质二级结构分发现α-螺旋占45.65%,延伸链占16.14%,β-转角占7.01%,无规卷曲占31.21%。
26.克隆到的fanrt2.7与其它植物nrt2.7进行系统进化分析发现,与红颜草莓最相近的物种有森林草莓(fragaria vesca)和月季(rosa chinensis),其次有扁桃(prunus dulcis)、毛桃(prunus persica)、梅花(prunus mume)、甜樱桃(prunus avium)、苹果(malus domestica)、杜梨(pyrus betulifolia)和白梨(pyrus x bretschneideri),较远的有酸枣(ziziphus jujuba)、大麻(cannabis sativa)、拟南芥、芥菜型油菜(brassica juncear)、甘蓝型油菜(brassica napus)、巴西橡胶树(hevea brasiliens)、核桃(juglans regia)和欧洲栓皮栎(quercus suber)。虽然不同物种间有明显的变异性,但保守结构域也较多,可见克隆的fanrt2.7基因在进化过程中保持了一定的同源性。
27.以带酶切位点的特异性引物n27p f和n27p r为引物,克隆的fanrt2.7的启动子,
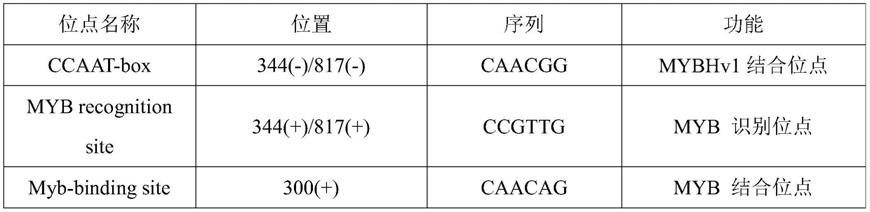
并进行plant care和place预测分析,plant care顺式元件分析如表1所示,place顺式元件分析如表2所示。采用plant care进行启动子顺式作用元件分析,发现fanrt2.7启动子上存在很多不同功能的顺式作用元件。有myb家族的相关结合或识别位点ccaat-box、myb recognition site、myb-binding site、myb-like sequence和mre,有mre、aca-motif、ae-box、atc-motif、ga-motif、g-box、sp1和tct-motif等11类与光响应相关的顺式作用元件,有cgtca-motif、tca-element、abre和p-box等激素应答元件(茉莉酸甲酯、水杨酸、脱落酸和赤霉素),也有auxrr-core、tga-element和cat-box与生长素、分生组织表达相关的元件,有ry-element的种子特异调控的元件,还有caat-box和tata-box等启动子核心元件。由此推测,fanrt2.7基因的表达也受光照和激素等多种因素调控。
28.表1 fanrt2.7基因启动子plant care顺式元件分析表
29.[0030][0031]
[0032]
表2 fanrt2.7启动子place顺式元件分析
[0033]
[0034][0035]
二、红颜草莓硝酸盐转运蛋白基因fanrtf2.7的表达模式
[0036]
1、红颜草莓硝酸盐转运蛋白基因fanrtf2.7的时空表达和响应模式。
[0037]
采用fanrt2.7定量引物,进行基因的表达模式分析。fanrt2.7的时空表达模式所示。在不同组织中fanrt2.7基因的相对表达量亦不相同。叶片、瘦果和花中的表达量最高;茎、根和果实中的表达量居中;芽中的表达量最低。随着果实的成熟,果实(包含种子)中的fanrt2.7基因的表达量。在成熟果中草莓瘦果种子也是成熟的时期的fanrt2.7基因表达量最高;在大绿果期,草莓果实迅速变大,瘦果种子开始发育,其中的fanrt2.7基因表达量也较高;其他时期的fanrt2.7基因表达量均较低。由此可知,fanrt2.7基因在叶片、瘦果和花中表达量较高,在果实成基因的表达量上升。
[0038]
2、红颜草莓硝酸盐转运蛋白基因fanrtf2.7对不同氮形态和硝态氮的响应模式
[0039]
不同氮形态处理30d的红颜草莓苗后,取fanrt2.7基因表达量较高的叶、花、大绿果和成熟果研究对不同氮形态的响应模式。在成熟叶和花中硝氮处理和硝氮-铵氮处理后fanrt2.7基因的表达量极显著上升。在大绿果中只有硝氮-铵氮处理后fanrt2.7基因的表达量极显著上升;在成熟果中,硝氮处理和铵氮处理后fanrt2.7基因的表达量反而会极显著下降。另外,在不同氮形态水培条件下处理1h后如图7,硝态氮和硝氮-铵氮处理后水培根中fanrt2.7基因表达量反而均极显著下降。由此可知,fanrt2.7基因响应硝酸盐和含硝酸盐的氮形态处理。同浓度硝态氮处理草莓组培苗后,fanrt2.7基因的表达情况如图8所示。饥饿培养7d后,fanrt2.7基因表达均会下降。之后随着不同hn和ln的硝酸盐恢复供应,基因的表达在48h内会有一个先上升再下降的过程,分别在1h和12h时达到fanrt2.7基因的表达最高值。在不同氮形态响应发现fanrt2.7响应硝酸盐的基础上,取硝态氮的响应做进一步的研究。如图9所示,硝氮处理后成熟叶和花中fanrt2.7基因的相对表达量均显著或极显著高于对照处理。尤其在叶片中的上升最显著。相对于对照处理,硝氮处理后成熟叶中fanrt2.7基因的相对表达量极显著上升了5.02倍。其次,在花中fanrt2.7基因的相对表达量也极显著上升,分别上升了0.54倍和0.42倍。但是在根和果实(成熟果)中fanrt2.7基因的相对表达量在硝氮处理后表现出显著下降。由此可知,响应硝态氮fanrt2.7基因的相对表达量在根和果中下降,在叶和花中则显著上升。
[0040]
3、红颜草莓硝酸盐转运蛋白基因fanrtf2.7对激素的响应模式
[0041]
喷洒meja、sa和naa可以极显著提高叶片中fanrt2.7的基因表达;而在果实中,喷洒这5种激素均可以极显著提高fanrt2.7的基因表达,其中喷洒meja、sa和naa可以使基因的表达提高10倍以上;果实中fanrt2.7响应激素的程度更显著一些。这些结果可以表明,fanrt2.7的基因表达可能受meja、sa和naa的诱导。
[0042]
4、红颜草莓硝酸盐转运蛋白基因fanrtf2.7对其他非生物胁迫的响应模式
[0043]
其他非生物胁迫处理也影响草莓fanrtf2.7的表达如图11所示。fanrt2.7基因的表达会受低温处理极显著提高到4.16倍,而高温处理后则极显著降低。干旱胁迫时fanrt2.7基因的表达会极显著降低,而复水后基因表达会有所上升,到复水24h时基因表达仍极显著低于正常水平。当黑暗胁迫时,fanrt2.7基因的表达会极显著降低到0.04倍,而恢复光照后基因的表达水平会再逐渐上升,光照1h和6h基因分别极显著下降到0.12倍和0.44倍。盐胁迫处理后基因的表达也会极显著下降。由此可知,fanrt2.7基因的表达会响应低温、复水和光照而提高,响应高温、干旱、黑暗和盐等胁迫条件而下降。
[0044]
5、红颜草莓硝酸盐转运蛋白基因fanrtf2.7亚细胞定位分析和组织定位分析
[0045]
本采用拟南芥原生质体对fanrt2.7进行亚细胞定位分析,可以发现fanrt2.7定位在质膜上。用含有pcambia1391z-profanrt2.7载体的农杆菌花序法侵染拟南芥,筛选鉴定出t2代阳性植株,对拟南芥植株进行gus染色。如图12、13所示,其中,种皮不能被染色,子叶和上胚轴可以被染色;根部前端不能被染色;叶片和茎可以被染色,幼小的花絮染色很浅;茎、叶和花被片可以被染色;花被片和柱头可以被染色;果荚可以被染色较深,但是其中的种子种皮不能被染色。由此可知,fanrt2.7基因在果荚、叶片和茎中表达较高,在花、种子中也有表达,但在果皮和根中表达量很低。
[0046]
综上所述,本技术提供的红颜草莓硝酸盐转运蛋白基因fanrtf2.7的基因和启动子的克隆与分析及对不同氮形态、硝态氮、激素和其他非生物胁迫的响应模式分析应用。通过克隆红颜草莓硝酸盐转运蛋白基因fanrtf2.7的基因和启动子的克隆,从而进行不同氮形态、硝态氮、激素和其他非生物胁迫的响应模式分析,应用对筛选种质材料、培育优质品种和应对非生物胁迫等方面均具有重要的生产意义。
[0047]
以上所述仅为本发明的较佳实施例,对本发明而言仅仅是说明性的,而非限制性的。本专业技术人员理解,在本发明权利要求所限定的精神和范围内可对其进行许多改变,修改,甚至等效,但都将落入本发明的保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。