dna聚合酶、核酸适配体、热启动dna聚合酶及方法和应用
技术领域
1.本发明涉及生物技术领域,尤其涉及一种修饰的dna聚合酶、核酸适配体、热启动dna聚合酶、试剂盒、生物合成dna聚合酶的方法、制备热启动dna聚合酶的方法及dna聚合酶和核酸适配体相配合用于检测核酸或者合成核酸的应用。
背景技术:
2.核酸扩增技术广泛用于研究人类疾病以及检测病原体。除pcr外,还有多种等温扩增技术可以用于核酸诊断技术,如环介导等温扩增(lamp)、重组酶聚合酶扩增(rpa)、链置换扩增(sda)和滚环扩增(rca)。据报道,这些技术还用于检测正在传播的严重急性呼吸综合征冠状病毒-2(sars-cov-2)。
3.核酸检测技术的一个关键挑战是由引物错配和非模板扩增引起的非特异性扩增。非特异性扩增强烈阻碍或降低目标模板的扩增,降低了检测的灵敏度。避免非特异性扩增的有效方法是阻断嗜热dna聚合酶在低温下的活性。在pcr反应中,用抗体或特异性适配体阻断dna聚合酶的活性是防止非特异性扩增的最常用方法。对dna聚合酶(如taq dna聚合酶)进行共价修饰是阻断其低温阶段活性的另一种方法。此外,在dna聚合酶的氨基酸中引入特定突变位点也会改变其性质并降低聚合酶的活性。
4.与pcr相比,等温扩增技术中也存在非特异性扩增,严重影响lamp等相关技术的应用。然而,目前解决等温扩增中非特异性扩增问题的方法很少。据报道,酰胺添加剂可以提高lamp中核酸扩增的特异性并减少了背景扩增。但有机溶剂的使用也有很多局限性,如无法用于冻干试剂的制备。已发现来自于腾冲嗜热厌氧菌的解旋酶可抑制非靶核酸的扩增,从而有效降低背景信号。然而,解旋酶水解datp并释放h
,导致溶液ph改变,限制了其在ph介导的比色lamp中的使用。
技术实现要素:
5.为了克服现有技术的不足,本发明的目的之一在于提供一种修饰的dna聚合酶,可提供广谱的制备热启动dna聚合酶的技术策略。
6.本发明的目的之一采用如下技术方案实现:一种修饰的dna聚合酶,包括dna聚合酶片段和融合于所述dna聚合酶片段的n端的g-四链体结合肽。
7.本发明的目的之二在于提供一种用于调控上述的dna聚合酶活性的核酸适配体。
8.本发明的目的之二采用如下技术方案实现:一种用于调控上述dna聚合酶活性的核酸适配体,所述核酸适配体的核苷酸序列包括g-四链体核心序列,所述g-四链体核心序列的二级结构为g-四链体;
9.所述g-四链体用于在第一预设温度时结合于所述dna聚合酶的g-四链体结合肽,以抑制所述dna聚合酶的活性,并用于在第二预设温度时脱离于所述dna聚合酶的g-四链体结合肽,以恢复所述dna聚合酶的活性;其中,所述第二预设温度高于所述第一预设温度。
10.本发明的目的之三在于提供一种热启动dna聚合酶。
11.本发明的目的之三采用如下技术方案实现:一种热启动dna聚合酶,包括上述的dna聚合酶和上述的核酸适配体,所述核酸适配体的g-四链体在第一预设温度结合于所述dna聚合酶的g-四链体结合肽,以抑制所述dna聚合酶的活性,并在第二预设温度脱离于所述dna聚合酶的g-四链体结合肽,以恢复所述dna聚合酶的活性;其中,所述第二预设温度高于所述第一预设温度。
12.本发明的目的之四在于提供一种试剂盒。
13.本发明的目的之四采用如下技术方案实现:一种试剂盒,包括上述dna聚合酶和上述核酸适配体。
14.本发明的目的之五在于提供一种生物合成上述dna聚合酶的方法。
15.本发明的目的之五采用如下技术方案实现:一种生物合成上述dna聚合酶的方法,包括如下步骤:
16.将dna聚合酶片段插入质粒载体,构建第一质粒;
17.在所述第一质粒的基础上,将g-四链体结合肽的编码序列插入所述dna聚合酶片段的n端,构建第二质粒;
18.将所述第二质粒转化到大肠杆菌菌株中进行培养并诱导蛋白质表达;
19.纯化蛋白质,获得所述的dna聚合酶。
20.本发明的目的之六在于提供一种制备热启动dna聚合酶的方法。
21.本发明的目的之六采用如下技术方案实现:一种制备热启动dna聚合酶的方法,所述dna聚合酶/适配体复合物包括上述的dna聚合酶和上述的核酸适配体,所述核酸适配体通过所述g-四链体结合于所述g-四链体结合肽而结合于所述dna聚合酶,所述方法包括如下步骤:
22.将所述核酸适配体溶解于第一缓冲液中,在90℃~100℃变性2分钟~8分钟后,冷却至20℃~30℃,获得处理后的核酸适配体;
23.将所述dna聚合酶和处理后的核酸适配体按预设的摩尔浓度比加入第二缓冲液中,在2℃~6℃下孵育30min~60min;
24.其中,所述第一缓冲液包括10mm ph 7.4tris-hcl、75mm kcl、0.5mm edta和0.2mg/ml牛血清白蛋白,所述第二缓冲液包括20mm ph 8.8tris-hcl、10mm(nh4)2so4、50mm kcl、2-8mm mgso4和0.1%吐温-20。
25.本发明的目的之七在于提供上述dna聚合酶和上述核酸适配体相配合用于检测核酸或者合成核酸的应用。
26.本发明的目的之七采用如下技术方案实现:上述dna聚合酶和上述核酸适配体相配合用于检测核酸或者合成核酸的应用。
27.相比现有技术,本发明的有益效果在于:(1)本发明构建一种修饰的dna聚合酶以及与该dna聚合酶特异性结合的核酸适配体,dna聚合酶的活性在低温阶段被结合的核酸适配体严格抑制,但在加热到反应温度时完全恢复,在应用于等温扩增时,不会产生非特异性扩增,因此,它在分子诊断中具有很大的应用价值;(2)采用本发明构建的修饰的dna聚合酶以及与该dna聚合酶特异性结合的核酸适配体,成功地解决了lamp检测hpv dna和sars-cov-2rna的非特异性扩增问题;(3)g-四链体结合肽与g-四链体适配体的结合为可控dna聚合酶的开发提供了一条新途径。
附图说明
28.图1a为本发明实施例一提供的bst dna聚合酶、bst-lf和dna聚合酶的结构示意图,其中dna聚合酶融合有g-四链体结合肽,用g4p-bst和rhau23-bst表示;
29.图1b为本发明实施例一提供的纯化的bst-lf、rhau23-bst和g4p-bst的电泳图;
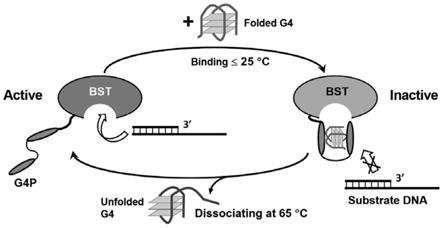
30.图1c为本发明实施例一提供的不同温度下,g-四链体核酸适配体调控dna聚合酶活性的原理示意图;
31.图1d为本发明实施例一提供的pcold-i-g4p-bst质粒图谱;
32.图1e为本发明实施例一提供的pcold-i-rhau23-bst质粒图谱;
33.图2a至图2c为本发明实施例二提供的dna聚合酶g4p-bst与核酸适配体t4b1 core g4、pdgfrb core g4、t4b1 fs-g4、pdgfrb fs-g4、t4b1 fd-g4和pdgfrb fd-g4的结合能力结果图;
34.图2d为本发明实施例二提供的dna聚合酶rhau23-bst与核酸适配体t4b1 fd-g4和pdgfrb fd-g4的结合能力结果图;
35.图2e为本发明实施例二提供的dna聚合酶rhau23-bst与核酸适配体hg53、gh5、hg3的结合能力结果图;
36.图3a为本发明实施例三提供的引物延伸实验的原理图;
37.图3b至图3d为本发明实施例三提供的使用dna聚合酶g4p-bst进行引物延伸实验的结果图;
38.图3e至图3h为本发明实施例三提供的使用dna聚合酶rhau23-bst进行引物延伸实验的结果图;
39.图4a为本发明实施例四提供的通过emsa方法检测dna聚合酶g4p-bst与非g4单链dna和非g4双链dna的结合能力的结果图;
40.图4b为本发明实施例四提供的通过emsa方法检测bst-lf与t4b1 fd-g4和t4b1 fs-g4的结合能力的结果图;
41.图4c为本发明实施例四提供的通过引物延伸实验检测非g4单链dna和非g4双链dna存在时g4p-bst的活性的结果图;
42.图4d为本发明实施例四提供的通过引物延伸实验检测t4b1 fd-g4和t4b1 fs-g4存在时g4p-bst的活性的结果图;
43.图5为本发明实施例五提供的g-四链体类型、“发夹”结构长度和环长度对核酸适配体(hg53)抑制dna聚合酶rhau23-bst活性能力的影响结果图;
44.图6a为本发明实施例六提供的在不存在核酸适配体的情况下,bst-lf在不同温度下进行引物延伸实验的结果图;
45.图6b为本发明实施例六提供的在不存在核酸适配体的情况下,bst 2.0热启动dna聚合酶在不同温度下进行引物延伸实验的结果图;
46.图6c为本发明实施例六提供的在不存在核酸适配体的情况下,rhau23-bst在不同温度下进行引物延伸实验的结果图;
47.图6d为本发明实施例六提供的在存在核酸适配体hg53-gvbq1的情况下,rhau23-bst在不同温度下进行引物延伸实验的结果图;
48.图6e为本发明实施例六提供的在存在核酸适配体hg53-h7的情况下,rhau23-bst
在不同温度下进行引物延伸实验的结果图;
49.图6f为本发明实施例六提供的rhau23-bst在核酸适配体hg53-h7存在的情况下的标准化活性结果图;
50.图7为本发明实施例七提供的用荧光lamp检测hpv 16基因dna的结果图;
51.图8a是荧光rt-lamp检测sars-cov-2rna的结果图;图8b是ph介导的比色rt-lamp检测sars-cov-2rna的结果图;
52.图9a为本发明实施例九提供的pcold-i-rhau23-taq质粒图谱;
53.图9b为本发明实施例九提供的在不存在核酸适配体的情况下,rhau23-taq在不同温度下进行引物延伸实验的结果图;
54.图9c为本发明实施例九提供的在存在核酸适配体hg53-h7的情况下,rhau23-taq在不同温度下进行引物延伸实验的结果图;
55.图9d为本发明实施例九提供的存在核酸适配体hg53-h7时rhau23-taq的活性相比不存在核酸适配体时的比值;
56.图10a为本发明实施例十提供的pcold-i-rhau23-rt质粒图谱;
57.图10b为本发明实施例十提供的在存在核酸适配体hg53-h7的情况下,rhau23-rt在不同温度下进行引物延伸实验的结果图;
58.图10c为本发明实施例十提供的在不存在核酸适配体的情况下,现有mmlv逆转录酶在不同温度下进行引物延伸实验的结果图;
59.图10d为本发明实施例十提供的存在核酸适配体hg53-h7时rhau23-rt逆转录酶活性相比不存在核酸适配体时现有mmlv逆转录酶活性的比值。
具体实施方式
60.下面,结合附图以及具体实施方式,对本发明做进一步描述,需要说明的是,在不相冲突的前提下,以下描述的各实施例之间或各技术特征之间可以任意组合形成新的实施例。
61.本发明提供一种修饰的dna聚合酶,包括dna聚合酶片段和融合于dna聚合酶片段的n端的g-四链体结合肽。
62.作为一种实施方式,g-四链体结合肽为rhau23肽,rhau23肽的氨基酸序列如seq id no.1所示;或者,
63.g-四链体结合肽为g4p,g4p的氨基酸序列如seq id no.2所示。
64.作为一种实施方式,dna聚合酶片段耐受40℃以上的温度,且在温度高于40℃以上时具有活性。
65.作为一种实施方式,dna聚合酶片段来自于bst dna聚合酶、taq dna聚合酶和mmlv逆转录酶中的任意一种。
66.作为一种实施方式,dna聚合酶片段来自于bst dna聚合酶,dna聚合酶片段的氨基酸序列如seq id no.3所示。
67.作为一种实施方式,dna聚合酶片段来自于taq dna聚合酶,dna聚合酶片段的氨基酸序列如seq id no.4所示。
68.作为一种实施方式,dna聚合酶片段来自于mmlv逆转录酶,dna聚合酶片段的氨基
酸序列如seq id no.5所示。
69.本发明还提供一种用于调控上述dna聚合酶活性的核酸适配体,核酸适配体的核苷酸序列包括g-四链体核心序列,g-四链体核心序列的二级结构为g-四链体;
70.g-四链体用于在第一预设温度时结合于dna聚合酶的g-四链体结合肽,以抑制dna聚合酶的活性,并用于在第二预设温度时脱离于dna聚合酶的g-四链体结合肽,以恢复dna聚合酶的活性;其中,第二预设温度高于第一预设温度。
71.作为一种实施方式,g-四链体为规则的三层g-四链体、凸起的g-四链体、带有g-空位的g-四链体和规则的两层g-四链体中的任意一种。
72.作为一种实施方式,核酸适配体的核苷酸序列还包括g-四链体核心序列的5’端和/或3’端的旁侧dna序列。
73.作为一种实施方式,旁侧dna序列为侧翼单链dna序列或者侧翼双链dna序列或者用于形成“发夹”结构的dna序列。
74.作为一种实施方式,旁侧dna序列优选为用于形成“发夹”结构的dna序列,其中,形成的“发夹”结构的长度大于或等于7bp。
75.作为一种实施方式,第一预设温度为0℃~30℃,第二预设温度为45℃~70℃。
76.作为一种实施方式,第二预设温度为55℃~65℃。
77.作为一种实施方式,g-四链体核心序列为cstb核心序列,cstb核心序列的核苷酸序列如seq id no.6所示;或者,
78.g-四链体核心序列为kit-c核心序列,kit-c核心序列的核苷酸序列如seq id no.7所示;或者,
79.g-四链体核心序列为t4b1核心序列,t4b1核心序列的核苷酸序列如seq id no.8所示;或者,
80.g-四链体核心序列为pdgfrb核心序列,pdgfrb核心序列的核苷酸序列如seq id no.9所示;或者,
81.g-四链体核心序列为t1b1核心序列,t1b1核心序列的核苷酸序列如seq id no.10所示;或者,
82.g-四链体核心序列为gvbq1核心序列,gvbq1核心序列的核苷酸序列如seq id no.11所示;或者,
83.g-四链体核心序列为gvbq2核心序列,gvbq2核心序列的核苷酸序列如seq id no.12所示;或者,
84.g-四链体核心序列为g12核心序列,g12核心序列的核苷酸序列如seq id no.13所示。
85.本发明还提供一种热启动dna聚合酶,包括上述dna聚合酶和上述核酸适配体,核酸适配体的g-四链体在第一预设温度结合于dna聚合酶的g-四链体结合肽,以抑制dna聚合酶的活性,并在第二预设温度脱离于dna聚合酶的g-四链体结合肽,以恢复dna聚合酶的活性;其中,第二预设温度高于第一预设温度。
86.作为一种实施方式,第一预设温度为0℃~30℃,第二预设温度为45℃~70℃。
87.本发明还提供一种试剂盒,包括上述dna聚合酶和上述核酸适配体。
88.作为一种实施方式,试剂盒用于检测人乳头瘤病毒dna或者sars病毒rna。
89.本发明还提供一种生物合成上述dna聚合酶的方法,其特征在于,包括如下步骤:
90.将dna聚合酶片段插入质粒载体,构建第一质粒;
91.在第一质粒的基础上,将g-四链体结合肽的编码序列插入dna聚合酶片段的n端,构建第二质粒;
92.将第二质粒转化到大肠杆菌菌株中进行培养并诱导蛋白质表达;
93.纯化蛋白质,获得的dna聚合酶。
94.作为一种实施方式,dna聚合酶片段来自于嗜热脂肪芽孢杆菌的dna聚合酶的291-878残基,质粒载体为pcold-i,第一质粒为pcold-i-bst-lf质粒;
95.g-四链体结合肽为g4p,第二质粒为pcold-i-g4p-bst质粒;或者,g-四链体结合肽为rhau23,第二质粒为pcold-i-rhau23-bst质粒。
96.作为一种实施方式,dna聚合酶片段来自于taq dna聚合酶,质粒载体为pcold-i,第一质粒为pcold-i-taq-lf质粒;
97.g-四链体结合肽为g4p,第二质粒为pcold-i-g4p-taq质粒;或者,g-四链体结合肽为rhau23,第二质粒为pcold-i-rhau23-taq质粒。
98.作为一种实施方式,dna聚合酶片段来自于mmlv逆转录酶,质粒载体为pcold-i,第一质粒为pcold-i-rt-lf质粒;
99.g-四链体结合肽为g4p,第二质粒为pcold-i-g4p-rt质粒;或者,g-四链体结合肽为rhau23,第二质粒为pcold-i-rhau23-rt质粒。
100.本发明还提供一种制备热启动dna聚合酶的方法,热启动dna聚合酶上述dna聚合酶和上述核酸适配体,核酸适配体通过g-四链体结合于g-四链体结合肽而结合于dna聚合酶,方法包括如下步骤:
101.将核酸适配体溶解于第一缓冲液中,在90℃~100℃变性2分钟~8分钟后,冷却至20℃~30℃,获得处理后的核酸适配体;
102.将dna聚合酶和处理后的核酸适配体按预设的摩尔浓度比加入第二缓冲液中,在2℃~6℃下孵育30min~60min;
103.其中,第一缓冲液包括10mm ph 7.4tris-hcl、75mm kcl、0.5mm edta和0.2mg/ml牛血清白蛋白,第二缓冲液包括20mm ph 8.8tris-hcl、10mm(nh4)2so4、50mm kcl、2-8mm mgso4和0.1%吐温-20。
104.作为一种实施方式,dna聚合酶和核酸适配体的预设的摩尔浓度比的范围为1:8~1:1。
105.本发明还提供一种上述dna聚合酶和上述核酸适配体相配合用于检测核酸或者合成核酸的应用。
106.作为一种实施方式,在应用于检测核酸时,核酸为人乳头瘤病毒dna或者sars病毒rna。
107.本发明构建了g-四链体结合肽融合的dna聚合酶以及与该聚合酶特异性结合的核酸适配体,dna聚合酶的活性在低温阶段被结合的核酸适配体严格抑制,但在加热到反应温度时完全恢复,在应用于等温扩增时,不会产生非特异性扩增,因此,它在分子诊断中具有很大的应用价值;采用本发明构建的修饰的dna聚合酶以及与该dna聚合酶特异性结合的核酸适配体,成功地解决了lamp检测hpv dna和sars-cov-2rna的非特异性扩增问题;g-四链
体结合肽与g-四链体适配体的结合为可控dna聚合酶的开发提供了一条新途径。
108.实施例一:
109.修饰的dna聚合酶的构建。
110.通过内切酶nde i和xba i将bst dna聚合酶大片段(bst-lf)插入pcold-i载体以构建pcold-i-bst-lf质粒。在pcold-i-bst-lf的基础上,将g4p的编码序列插入bst-lf的n端,构建了pcold-i-g4p-bst质粒,将rhau23的编码序列插入bst-lf的n端,构建了pcold-i-rhau23-bst质粒。
111.将质粒pcold-i-bst-lf、pcold-i-g4p-bst和pcold-i-rhau23-bst转化到大肠杆菌菌株bl21中。细胞在含有50μg/ml氨苄青霉素的lb培养基中,37℃培养至od 0.8-1.0。然后,添加0.4mm iptg并在16℃下再培养16h来诱导蛋白质表达。根据说明书使用his-tag钴树脂(thermo-scientific)纯化蛋白质,蛋白质储存在含有20mm tris-hcl、150mm nacl、1mm dtt、0.5mm edta和50%甘油的缓冲液中。图1a示出了bst dna聚合酶、bst-lf和修饰的dna聚合酶的结构示意图,其中修饰的dna聚合酶融合有g-四链体结合肽,用g4p-bst和rhau23-bst表示。图1b为纯化的bst-lf、rhau23-bst和g4p-bst的电泳图。
112.其中,bst-lf来自于嗜热脂肪芽孢杆菌(ara98840.1)的dna聚合酶i的291-878残基,也就是嗜热脂肪芽孢杆菌dna聚合酶中缺少5'-3'核酸外切酶结构域的部分,bst-lf的氨基酸序列如seq id no.3所示。bst-lf在较宽的温度范围内具有活性,并且具有较强的链置换活性,被广泛应用于环介导等温扩增(lamp)。rhau23是一种来自人类dhx36蛋白的23个氨基酸的肽,它与g-四链体特异性相互作用,rhau23的氨基酸序列如seq id no.1所示。g4p是一种由两个串联的rhau23组成的小蛋白质(64个氨基酸),g4p比rhau23具有更强的g-四链体结合亲和力,g4p的氨基酸序列如seq id no.2所示。pcold-i-g4p-bst质粒图谱见图1d,pcold-i-rhau23-bst质粒图谱见图1e。
113.本实施例提供的修饰的dna聚合酶,通过在n端融合有g-四链体结合肽,能够与具有g-四链体的核酸适配体相结合。如图1c所示,在低温阶段,融合g-四链体结合肽的dna聚合酶与g-四链体结合,阻止dna聚合酶与底物dna相互作用,在高温阶段,g-四链体dna去折叠并从dna聚合酶中分离,从而dna聚合酶可以重新作用于底物。
114.实施例二:
115.通过凝胶迁移实验(emsa)检测修饰的dna聚合酶与核酸适配体结合亲和力。
116.寡核苷酸购自sangon生物技术有限公司(中国)。互补单链dna(ssdna)溶解在含有10mm tris-hcl(ph7.4)、75mm kcl和0.5mm edta的缓冲液中,在95℃变性5分钟后,以0.1℃/s的速度冷却至25℃,从而配对成双链dna(dsdna)。
117.将20nm 5'-fam标记的核酸适配体dna(表1)溶解在第一缓冲液中,在95℃变性5分钟后,以0.1℃/s的速率冷却至25℃;其中,第一缓冲液包括10mm tris-hcl(ph值7.4)、75mm kcl、0.5mm edta和0.2mg/ml牛血清白蛋白。然后将各核酸适配体dna与指定浓度的融合g-四链体结合肽的dna聚合酶加入第二缓冲液中,在4℃下结合1小时,形成dna和蛋白结合的复合物;其中,第二缓冲液包括20mm tris-hcl(ph 8.8),10mm(nh4)2so4、50mm kcl、8mm mgso4和0.1%吐温-20。
118.核酸适配体和dna聚合酶的结合通过凝胶迁移实验来检测。具体地,在含有75mm kcl的1
×
tbe缓冲液中,dna和蛋白结合的复合物在4℃下在含有75mm kcl的8%或12%非变
性聚丙烯酰胺凝胶上电泳2小时。通过chemidoc mp成像系统(bio-rad)对dna和蛋白结合的复合物进行拍照,并通过image quant 5.2软件进行定量分析。
119.结果如图2所示,其中图2a至图2c为修饰的dna聚合酶g4p-bst与核酸适配体t4b1 core g4、pdgfrb core g4、t4b1 fs-g4、pdgfrb fs-g4、t4b1 fd-g4和pdgfrb fd-g4的结合能力结果图,其中core g4表示核心g-四链体,fs-g4表示侧翼含单链dna的g-四链体,fd-g4表示侧翼含双链dna的g-四链体;图2d为修饰的dna聚合酶rhau23-bst与核酸适配体t4b1 fd-g4和pdgfrb fd-g4的结合能力结果图,图2e为修饰的dna聚合酶rhau23-bst与核酸适配体hg53、gh5、hg3的结合能力结果图,其中核酸适配体hg53、gh5、hg3均为可折叠成g-四链体和“发夹”结构的单链dna,而上述的fd-g4适配体是由两个互补的单链dna配对而成的。
120.从图中可以看出,无论t4b1的凸起的g-四链体(凸起的g-四链体也称为bulged g-四链体)和pdgfrb的凸起的g-四链体是否具有旁侧dna序列,g4p-bst显示出与t4b1的凸起的g-四链体和pdgfrb的凸起的g-四链体均具有强的结合亲和力。其中,kd值范围为1-5nm。rhau23-bst与含有g-四链体和侧翼双链dna的核酸适配体也具有强的结合亲和力,kd处于nm水平。并且rhau23-bst与核酸适配体hg53、gh5、hg3的结合亲和力与rhau23-bst与含有g-四链体和侧翼双链dna的核酸适配体的结合亲和力相似,kd值范围为3.4-7.4nm。
121.表1
122.[0123][0124]
实施例三:
[0125]
通过引物延伸实验检测核酸适配体对dna聚合酶活性的调节。
[0126]
将表2中列出的各核酸适配体dna溶解在第一缓冲液中至10μm,在95℃下加热5min,然后以0.1℃/s的速率冷却至25℃;其中,第一缓冲液包括10mm tris-hcl(ph值7.4)、75mm kcl、0.5mm edta和0.2mg/ml牛血清白蛋白。将指定比例的融合g-四链体结合肽的dna聚合酶(100nm)与核酸适配体加入第二缓冲液中,在4℃下放置30min形成dna和蛋白结合的复合物;其中,融合g-四链体结合肽的dna聚合酶与核酸适配体的摩尔浓度比分别为1:0、1:1、1:2、1:4和1:8,第二缓冲液包括20mm tris-hcl(ph 8.8),10mm(nh4)2so4、50mm kcl、8mm mgso4和0.1%吐温-20。添加2.5mm dntp,100nm引物和100nm模板dna到dna和蛋白结合的复合物中,并在25℃或65℃下反应5分钟或者40分钟,其中,引物和模板dna的序列请参阅表3。反应完成后添加四倍体积的终止缓冲液(99%甲酰胺、0.1%sds和20mm edta)终止反应。样品在95℃下煮沸5分钟,然后在1
×
tbe缓冲液中的12%尿素变性聚丙烯酰胺凝胶上电泳。通过chemidoc mp成像系统(bio-rad)上对引物和全长产物进行拍照,并使用image quant 5.2软件进行数字化定量。
[0127]
表2
[0128]
[0129][0130]
表3
[0131][0132]
图3a为引物延伸实验的原理图,图3b至图3d为使用修饰的dna聚合酶g4p-bst进行引物延伸实验的结果图,图3e至图3h为使用修饰的dna聚合酶rhau23-bst进行引物延伸实验的结果图。
[0133]
从图中可以看出,当仅以核心g-四链体(core g4)作为核酸适配体时,也就是核酸适配体不具有任何旁侧dna序列,无论是在25℃还是65℃,g4p-bst的活性几乎不受影响,详见图3b;当使用含侧翼单链dna的g-四链体作为核酸适配体时,也就是使用fs-g4适配体,g4p-bst的活性在25℃时表现出不同程度的抑制,该抑制与修饰的dna聚合酶/核酸适配体的比率相关,在65℃时修饰的dna聚合酶的活性基本恢复,详见图3c。当使用含侧翼双链dna的g-四链体作为核酸适配体时,也就是使用fd-g4适配体,只要dna聚合酶/核酸适配体的比例小于1,g4p-bst在25℃时的活性几乎完全被抑制,fd-g4适配体对g4p-bst活性的抑制强烈,在65℃下仍表现出不同程度的抑制,详见图3d。由于g4p-bst与上述三种核酸适配体的结合亲和力相似,但含侧翼dna的适配体明显效果更好因此,g-四链体的侧翼区域的dna也是适配体发挥作用所必需的。
[0134]
g4p-bst与fd-g4适配体结合后,其活性被严重抑制,即使升温到65℃也没有完全恢复;相比之下rhau23-bst与fd-g4适配体结合后,其在65℃下的活性几乎完全恢复,详见图3e。核酸适配体对这两个聚合酶的抑制效果不同与g4p和rhau23对g4结合能力相关。这说明我们可以通过改变g-四链体结合肽来调节适配体对酶活性的抑制能力。
[0135]
rhau23-bst与核酸适配体hg53、gh5、hg3在25℃下被强烈的抑制,且抑制作用至少持续40分钟。而在65℃时,rhau23-bst完全恢复活性。这表明以可折叠成g-四链体和“发夹”结构的单链dna作为核酸适配体足以调节融合g-四链体结合肽的dna聚合酶的活性。
[0136]
由此可见,核酸适配体对修饰的dna聚合酶的活性的调节受到核酸适配体中的旁侧dna序列和基于g-四链体调控的热启动dna聚合酶中的g-四链体结合肽的影响。并且,修饰的dna聚合酶g4p-bst和rhau23-bst的活性在25℃时被抑制,25℃是接近室温的温度,在该温度下聚合酶活性的抑制可以有效阻止非特异性扩增。
[0137]
实施例四:
[0138]
通过emsa和引物延伸实验检测g-四链体和g-四链体结合肽对热启动dna聚合酶的必要性。
[0139]
参照实施例二的实验方法,通过emsa方法检测dna聚合酶g4p-bst与非g4单链dna和非g4双链dna的结合能力,以及通过emsa方法检测bst-lf与t4b1 fd-g4和t4b1 fs-g4的结合能力。
[0140]
参照实施例三的实验方法,通过引物延伸实验检测非g4单链dna和非g4双链dna存在时g4p-bst的活性,以及通过引物延伸实验检测t4b1 fd-g4和t4b1 fs-g4存在时g4p-bst的活性。
[0141]
图4示出了实验结果,其中,图4a为通过emsa方法检测dna聚合酶g4p-bst与非g4单链dna和非g4双链dna的结合能力的结果图,图4b为通过emsa方法检测bst-lf与t4b1 fd-g4和t4b1 fs-g4的结合能力的结果图,图4c为通过引物延伸实验检测非g4单链dna和非g4双链dna存在时g4p-bst的活性的结果图,图4d为通过引物延伸实验检测t4b1 fd-g4和t4b1 fs-g4存在时bst-lf的活性结果图。
[0142]
从图中可以看出,从核酸适配体中去除g-四链体或者从dna聚合酶中去除g-四链体结合肽都显著降低了dna聚合酶与核酸适配体的结合亲和力,均导致无法调节dna聚合酶的活性。因此,核酸适配体中的g-四链体和dna聚合酶中的g-四链体结合肽是控制热启动dna聚合酶的两个不可或缺的部分。
[0143]
实施例五:
[0144]
通过引物延伸实验验证核酸适配体抑制修饰的dna聚合酶的能力可通过g-四链体类型和“发夹”结构的长度进行调节。
[0145]
5.1在温度为25℃下,通过引物延伸实验检测含有不同g-四链体类型的hg53核酸适配体存在下修饰的dna聚合酶rhau23-bst的活性。
[0146]
本实验中,选用四种不同g-四链体类型的八种核酸适配体进行实验,八种核酸适配体分别为cstb、kit-c、pdgfrb、t1b1、t4b1、gvbq1、gvbq2和g12。其中,核酸适配体cstb和kit-c的g-四链体类型为规则的三层g-四链体,核酸适配体pdgfrb、t1b1和t4b1的g-四链体类型为凸起的g-四链体,核酸适配体gvbq1和gvbq2的g-四链体类型为带有g-空位的g-四链体,核酸适配体g12的g-四链体类型为规则的两层g-四链体。rhau23-bst与核酸适配体的摩尔浓度比为1:2。
[0147]
结果如图5a所示,从图中可以看出,所有八种核酸适配体均能有效抑制修饰的dna聚合酶的活性。其中,含有非规则g-四链体的核酸适配体(t4b1、gvbq1和gvbq2)和含有非稳定的双层g-四链体的核酸适配体(g12)都可以抑制修饰的dna聚合酶的活性至少40分钟。
[0148]
5.2在温度为25℃下,通过引物延伸实验检测含有不同“发夹”结构长度的hg53核酸适配体存在下修饰的dna聚合酶rhau23-bst的活性。
[0149]
本实验中,分别选用“发夹”结构长度为8bp、7bp、6bp、5bp和4bp的hg53核酸适配体,rhau23-bst与核酸适配体的摩尔浓度比为1:2。
[0150]
结果如图5b所示,其中h8表示“发夹”结构长度为8bp的核酸适配体,h7表示“发夹”结构长度为7bp的核酸适配体,h6表示“发夹”结构长度为6bp的核酸适配体,h5表示“发夹”结构长度为5bp的核酸适配体。从图中可以看出,“发夹”区域的长度至少需要7bp,核酸适配体才能有效发挥作用。
[0151]
5.3在温度为25℃下,通过引物延伸实验检测含有不同环长的hg53核酸适配体存在下修饰的dna聚合酶rhau23-bst的活性。
[0152]
与gh5和hg3相比,hg53有一个额外的环连接g-四链体和“发夹”结构。该环主要影响dna末端“发夹”结构的形成,本实施例进行的实验中,分别取环长度为2nt、4nt、6nt、8nt、10nt和12nt的hg53核酸适配体进行实验,rhau23-bst与核酸适配体的摩尔浓度比为1:2。
[0153]
结果如图5c所示,从图中可以看出,该环的长度需要为6nt或更多。另外,以删除g-四链体或者突变其序列的核酸适配体进行实验,发现删除g-四链体或突变其序列使核酸适配体不再起到抑制修饰的dna聚合酶的作用,可见g-四链体是适配体的关键成分。
[0154]
实施例六:
[0155]
核酸适配体在不同温度下对bst dna聚合酶活性抑制能力的检测。
[0156]
参照实施例三的实验方法,通过引物延伸实验测试在不同温度下核酸适配体hg53-gvbq1和hg53-h7对三种bst dna聚合酶的抑制作用。其中,三种bst dna聚合酶分别为bst-lf、bst 2.0热启动dna聚合酶(bst 2.0热启动dna聚合酶是neb公司的商品化的热启动dna聚合酶)和修饰的dna聚合酶rhau23-bst;检测温度为25℃-65℃;bst-lf、rhau23-bst反应终浓度为100nm,bst 2.0热启动dna聚合酶为0.32u/μl;核酸适配体使用浓度为200nm;反应缓冲液为第二缓冲液,包括20mm tris-hcl(ph 8.8)、10mm(nh4)2so4、50mm kcl、8mm mgso4和0.1%吐温-20。
[0157]
图6为本实施例的实验结果图,请参阅图6a至图6c,在没有核酸适配体结合的情况下,三种bst dna聚合酶(bst-lf、bst 2.0热启动dna聚合酶和rhau23-bst)在25℃-65℃合成dna时表现出相似的活性。请参阅图6d和图6e,rhau23-bst与核酸适配体hg53-h7和hg53-gvbq1结合后的热启动酶的活性在低于30℃以下受到极大抑制。图6f示出了通过比较扩增后的全长产物与rhau23-bst的分数来确rhau23-bst在核酸适配体hg53-h7存在的情况下的标准化活性结果图,从图中可以看出,在低于30℃时,核酸适配体hg53-h7使rhau23-bst的活性抑制了80%以上,在高于35℃时,rhau23-bst的活性逐渐恢复,并在高于50℃时,rhau23-bst的活性恢复到90%以上。从实验结果可以看出,结合核酸适配体的rhau23-bst的活性可以通过升高温度(热启动)来精确启动,在低温下有效抑制非特异性扩增;而商品化的bst 2.0热启动dna聚合酶的活性在低温下几乎没有被抑制。因此,本技术提供的修饰的dna聚合酶有助于其在检测核酸方面的广泛应用。
[0158]
实施例七:
[0159]
热启动dna聚合酶用于检测人乳头瘤病毒(hpv)dna。
[0160]
通过环介导等温扩增(lamp)进行检测,其中,lamp为现有技术,在此不再详述。本实验中,将hpv16的e6-e7基因的质粒十倍梯度稀释直到1拷贝每μl,根据e6-e7基因的dna序列设计如表4所示的引物,lamp反应中引物使用终浓度为fip/bip 1.6μμ,loop-f/loop-b 0.4μμ,f3/b3 0.2μμ。rhau23-bst反应终浓度为100nm。bst 2.0热启动dna聚合酶(neb)为0.32u/μl。适配体使用浓度为200nm。bst-lf和商品化的bst 2.0热启动dna聚合酶为对照。如图7a所示,在bst-lf存在的情况下,lamp反应的扩增曲线在50分钟内荧光没有增加,表明bst-lf未能检测到hpv16的dna序列。相比之下,请参阅图7b和图7c,当dna聚合酶rhau23-bst与核酸适配体hg53-h7的dna和蛋白结合的复合物用于lamp反应时,或者商品化的bst 2.0热启动dna聚合酶(图中用warm-start表示)用于lamp反应时,典型的扩增曲线根据模板
dna的浓度依次出现。由于rhau23-bst的活性受温度控制的限制,修饰的dna聚合酶rhau23-bst与核酸适配体hg53-h7的dna和蛋白结合的复合物可以在室温下放置8小时而不会影响后续lamp反应的准确性,详见图7b。相反,随着在室温下放置时间的增加,在bst 2.0热启动dna聚合酶存在下的样品的扩增信号逐渐降低,并且需要更长的时间才能达到平台期,详见图7c。因此,bst 2.0热启动dna聚合酶不能有效防止低温下非特异性扩增引起的引物耗尽,这与实施例六中图6b中的结果一致。
[0161]
表4
[0162][0163][0164]
实施例八:
[0165]
热启动dna聚合酶用于检测sars-cov-2。
[0166]
rt-lamp:在rt-lamp反应体系中加入5u warmstart rtx逆转录酶(neb),100nm核酸适配体组装的热启动dna聚合酶以及0.5
×
gelgreen,热启动dna聚合酶的修饰的dna聚合酶为rhau23-bst,核酸适配体为hg53-h7,修饰的dna聚合酶与核酸适配体摩尔比为1:2。其中,rt-lamp为现有技术,在此不再详述。本实验中,所用引物如表5所示,引物浓度:fip/bip为1.6μm,f3/b3为0.2μm,loop-f/b为0.4μm。扩增反应在65℃下在quantstudio 7flex(thermo scientific)的实时荧光定量pcr系统上进行。
[0167]
ph介导的比色rt-lamp:在ph介导的比色rt-lamp反应体系中加入5u warmstart rtx逆转录酶(neb),100nm核酸适配体组装的热启动dna聚合酶以及0.5
×
gelgreen,热启动dna聚合酶的修饰的dna聚合酶为rhau23-bst,核酸适配体为hg53-h7,修饰的dna聚合酶与核酸适配体的摩尔比为1:2。反应在65℃下进行指定时间,并通过甲酚红的颜色变化进行检测。
[0168]
图8a为使用荧光rt-lamp检测具有指定拷贝数的sars-cov-2假病毒rna的结果图;图8b为用比色rt-lamp检测sars-cov-2假病毒rna的结果图,其中,sars-cov-2假病毒rna的拷贝数与荧光rt-lamp检测中的相同。
[0169]
如图8a所示,核酸适配体hg53-h7结合rhau23-bst或现有的热启动bst dna聚合酶(warmstart 2.0bst)可有效防止非特异性扩增产生的假阳性结果。从图8b中可以看出,比色rt-lamp也显示了热启动bst dna聚合酶在防止非特异性扩增产生的假阳性结果方面的优势。
[0170]
表5
[0171][0172]
实施例九:
[0173]
g-四链体结合肽和含有g-四链体的核酸适配体的组合策略适用于taq dna聚合酶。
[0174]
参照实施例一的构建方法构建dna聚合酶rhau23-taq。其中,将质粒pcold-i-rhau23-bst中编码bst dna聚合酶大片段的序列替换成编码taq dna聚合酶的基因序列,构建质粒pcold-i-rhau23-taq,其中,pcold-i-rhau23-taq质粒图谱见图9a所示。
[0175]
参照实施例三的方法对rhau23-taq的活性进行检测。其中,rhau23-taq反应终浓度为100nm,hg53-h7 200nm。以不加核酸适配体的样品为对照,在不同温度下进行引物延伸30分钟。反应缓冲液为第二缓冲液,包括20mm tris-hcl(ph 8.8)、10mm(nh4)2so4、50mm kcl、2mm mgso4和0.1%吐温-20。
[0176]
在不同温度下,rhau23-taq结合含g-四链体的核酸适配体后,rhau23-taq的相对活性通过比较其全长(图中用fl表示)延伸产物与rhau23-taq的全长产物比例来计算。如图9b所示,rhau23-taq在25℃~65℃下都显示有活性。请参阅图9c和9d,当200nm hg53-h7存在时,rhau23-taq的活性在35℃下几乎全部抑制;当温度升高35℃以上时,rhau23-taq的活性逐渐恢复,特别是当温度升高50℃以上时,rhau23-taq的活性几乎完全恢复。因此,g-四链体结合肽和含有g-四链体的核酸适配体的结合策略适用于taq dna聚合酶,可以将taq dna聚合酶改造为修饰的dna聚合酶,使其具备热启动能力。
[0177]
可以理解,在其他实施例中,通过构建质粒pcold-i-g4p-taq来构建修饰的dna聚合酶g4p-taq也是可以的。
[0178]
实施例十:
[0179]
g-四链体结合肽和含有g-四链体的核酸适配体的结合策略适用于逆转录酶。
[0180]
参照实施例一的构建方法构建逆转录酶rhau23-rt。其中,将质粒pcold-i-rhau23-bst中编码bst大片段的序列替换成编码mmlv逆转录酶(rt)的基因序列,构建质粒pcold-i-rhau23-rt,其中pcold-i-rhau23-rt质粒图谱见图10a所示。
[0181]
参照实施例三的方法进行逆转录引物延伸实验。其中,使用rna为模板(rna模板的核苷酸序列如seq id no.14),与rna配对的dna为引物(dna引物的核苷酸序列如seq id no.15)。rhau23-rt反应终浓度为100nm,hg53-h7 200nm。反应缓冲液为第二缓冲液,包括20mm tris-hcl(ph 8.8)、10mm(nh4)2so4、50mm kcl、8mm mgso4和0.1%吐温-20。以未融合g-四链体结合肽以及未添加核酸适配体的样品为对照,在不同温度下进行引物延伸15分钟。
[0182]
请参阅图10c,现有的mmlv逆转录酶在25℃-65℃之间具有逆转录酶活性。请参阅图10b和图10d,当使用g-四链体结合肽和含有g-四链体的核酸适配体的结合策略对现有的mmlv逆转录酶进行改造之后,改造后的逆转录酶在30℃以下的逆转录活性被强烈抑制,当温度升高到35℃以上,改造后的逆转录活性能够大部分恢复,特别是在温度为50℃~60℃之间,改造后的逆转录活性恢复的更佳。因此,g-四链体结合肽和含有g-四链体的核酸适配体的结合策略适用于逆转录酶,可以将逆转录酶改造为修饰的逆转录酶,使其具备热启动的性质。
[0183]
可以理解,在其他实施例中,通过构建质粒pcold-i-g4p-rt来构建逆转录酶g4p-rt也是可以的。
[0184]
综上所述,g-四链体结合肽和含有g-四链体的核酸适配体的结合策略适用于多种dna聚合酶,可以将dna聚合酶改造为修饰的dna聚合酶,使其具备热启动的性质。
[0185]
上述实施方式仅为本发明的优选实施方式,不能以此来限定本发明保护的范围,本领域的技术人员在本发明的基础上所做的任何非实质性的变化及替换均属于本发明所要求保护的范围。
[0186]
[0187]
[0188]
[0189]
[0190]
[0191]
[0192]
[0193]
[0194]
[0195]
[0196]
[0197]
[0198]
[0199]
[0200]
[0201]
[0202]
[0203]
[0204]
[0205]
[0206]
[0207]
[0208]
[0209]
[0210]
[0211]
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
