1.本发明涉及分子生物学技术领域,具体涉及一种高效分离带有野生稻血缘的抗白叶枯病基因及其家族成员的方法。
背景技术:
2.水稻(oryza sativa l.)是全世界近一半人口的主粮,然而水稻生长发育过程中时常遭遇各种病原物的危害,严重制约着水稻的高产稳产以及稻米品质。由稻黄单胞杆菌(xanthomonas oryzae pv.oryzae,简称xoo)引起的水稻白叶枯病(bacterialblight)是世界范围内最严重的细菌性病害,也是我国水稻三大病害之一,我国除新疆、西藏和东北的北部以外均有不同程度的发生,长江以南和江淮平原籼稻产区为常发区,遭受白叶枯病危害后易造成水稻减产20%~30%,严重时可达50%,甚至绝收。随着全球气候逐渐变暖,水稻白叶枯病发生面积在逐年增加,水稻产量损失严重,已严重威胁到国家粮食安全(陈功友,徐正银,杨阳阳,等.我国水稻白叶枯病菌致病型划分和水稻抗病育种中应注意的问题.上海交通大学学报(农业科学版),2019,37(1):71-77.)。从长远途径来说,要想在水稻白叶枯病害常发区获得理想的水稻产量,就必须不断培育抗病新品种,以抵御白叶枯病菌的不断变异。尤其是多渠道、多来源发掘出抗白叶枯病基因,并服务于育种实践,已成为目前水稻分子育种的重要目标。
3.回顾水稻新品种的培育历程,育种家们大都将来自栽培品种中的优良基因不断重组、聚合,导致栽培品种原有的遗传资源利用日趋饱和,许多改良品种具有相同或相似的遗传来源,造成了遗传上的单一性,尤其由于早期育种家们对栽培稻抗病特性的忽视,我们现有的栽培稻不具有抗病特性或携有的抗病基因单一。从远缘种群中引入优良基因创作新的水稻品种是扩大亲本间遗传差异的重要途径之一。栽培稻有20多个野生近缘种,包含有极其丰富的遗传变异。云南省是亚洲栽培稻的遗传多样性中心和起源中心之一,拥有中国的全部三种野生稻,即普通野生稻(oryza rufipogon griff.)、药用野生稻(oryza officinalis wall.)和疣粒野生稻(oryza granulatabaill.),而且每种野生稻具有许多亚种或生态类型,这些种质资源在表型和分子水平上都具有较高的遗传多样性,拥有显著的抗逆、抗病虫害、耐寒、耐旱和耐贫瘠等许多优良性状(陈玲,张敦宇,陈越,等.云南药用野生稻种质资源的白叶枯病抗性评价.南方农业学报,2019,50(7):1417-1425;程在全.云南野生稻遗传特性及其优良基因克隆研究.四川:四川大学,2006.)。因此,利用拥有的广谱高抗白叶枯病的野生资源培育水稻新品种,这不仅将大大降低生产成本,而且具有重要的生态意义。
4.前期我们利用广谱高抗白叶枯病的元江普通野生稻渗入系(元江普通野生稻与栽培稻合系35杂交后代bc2f
16
)精细定位到了一个位于水稻11号染色体的显性抗白叶枯病基因xa47(t)(陈玲,钟巧芳,王玲仙,等.与水稻广谱高抗白叶枯病基因xa45(t)紧密连锁的分子标记hxjy-14,2020a,中国,zl201910825566.0.;陈玲,王波,程在全,等.水稻广谱高抗白
叶枯病基因xa45(t)的共分离分子标记hxjy-1,2020b,中国,zl201910825603.0.;陈玲,程在全,王波,等.与水稻广谱高抗叶枯病基因xa45(t)紧密连锁分子标记r13i14,2020c,中国,zl201910825560.3.),之前暂命名为xa45(t),因与位于4号染色体的水稻抗白叶枯病基因xa1的等位基因xa45(t)重名(ji ch,ji zy,liu b,et al.xa1 allelic r genes activate rice blight resistance suppressed by interfering tal effectors.plant comm,2020,1(4),100087.),所以后将其改为xa47(t)。利用xa47(t)的共分离标记检测发现该基因带有元江普通野生稻血缘,目前从野生稻中克隆到的基因只有3个,分别为来自长雄野生稻(oryza longistaminataa.chev et roehr)的xa21(song wy,wang gl,chen ll,et al.a receptor kinase-like protein encoded by the rice disease resistance gene,xa21.science,1995,270(5243):1804-1806.)、来自普通野生稻的xa23(zhouyl,uzokwe vne,zhang ch.improvement of bacterial blight resistance of hybrid rice in china using the xa23 gene derived from wild rice(oryza rufipogon).crop prot,2011,30(6):637-644.)和来自小粒野生稻(oryza minuta presl.)的xa27基因(gu k,tian d,yang f,et al.high-resolution genetic mapping of xa27(t),a new bacterial blight resistance gene in rice,oryza sativa l.theor appl genet,2004,108(5):800-807.),这3个基因的克隆方式基本都沿用了常规的图位克隆法,但该方法费时费力,随着测序技术的到来,利用测序技术高效且快速地分离xa47(t)基因已成为可能。经接种9个白叶枯病菌发现,359份元江普通野生稻渗入系抗病谱多样化,存在67种抗病谱,抗病谱不同携带的抗白叶枯病基因可能有所不同,或者这些抗病基因很可能为同一个家族成员,由于这些渗入系遗传背景相似,参照来自渗入系g252中的xa47(t)基因序列,利用元江普通野生稻的其他渗入系可大量地分离到xa47(t)家族成员。那么,如何充分利用现代测序技术以及分子生物学技术从元江普通野生稻及其渗入系中高效地分离xa47(t)基因及其家族成员成为了急需要解决的科学问题。
技术实现要素:
5.有鉴于此,本发明的目的在于提供一种高效分离带有野生稻血缘的抗白叶枯病基因及其家族成员的方法,能够高效快速地克隆到xa47(t)基因及其家族成员。
6.为解决上述技术问题,本发明提供了以下技术方案:
7.本发明提供了一种高效分离带有野生稻血缘的抗白叶枯病基因的方法,所述方法包括:
8.通过对元江普通野生稻渗入系双亲进行建库和高通量测序获得连续基因组,将所述连续基因组与参考基因组进行共线性分析,根据目的基因定位情况,筛选出所述抗白叶枯病基因的候选基因,根据所述候选基因设计引物进行pcr扩增、pcr产物直接测序和抗病功能分析,从而分离出带有野生稻血缘的抗白叶枯病基因。
9.优选的,所述抗白叶枯病基因为xa47(t),所述基因的核苷酸序列如seq id no.13所示;所述基因xa47(t)的cds序列如seq id no.14所示。
10.优选的,所述元江普通野生稻渗入系双亲为元江普通野生稻和栽培稻合系35;所述参考基因组为日本晴基因组;所述目的基因定位和所述pcr扩增的模板为元江普通野生稻渗入系g252。
11.优选的,所述引物为214qc-9f/r;所述214qc-9f的核苷酸序列如seq id no.11所示,所述214qc-9r的核苷酸序列如seq id no.12所示。
12.优选的,所述抗病功能分析包括功能互补试验和基因编辑试验分析候选基因的抗病功能。
13.本发明还提供了一种高效分离带有野生稻血缘的抗白叶枯病基因家族成员的方法,所述方法包括:
14.筛选获得含有xa47(t)基因或其同源基因的元江普通野生稻渗入系材料;根据上述xa47(t)基因序列,获得来自元江普通野生稻和合系35中的xa47(t)基因,分别命名为xa47(t)
yp
和xa47(t)
hx
;
15.利用引物对所述元江普通野生稻渗入系不同材料进行扩增、测序,得到4种xa47(t)基因的基因型:xa47(t)
g252
、xa47(t)
yp
、xa47(t)
hx
以及xa47(t)
l234
;对由4种基因型编码得到的蛋白质进行相似性分析,从而确定其是否为带有野生稻血缘的抗白叶枯病基因家族成员。
16.优选的,所述筛选的步骤包括:
17.利用xa47(t)基因的共分离标记hxjy-1对元江普通野生稻渗入系进行pcr扩增,得到扩增片段大小为167bp的渗入系,结合渗入系的抗病谱筛选出含有xa47(t)基因或其同源基因的元江普通野生稻渗入系材料。
18.优选的,根据上述xa47(t)基因序列,利用blast程序从元江普通野生稻和合系35基因组r13i14和hxjy-14区段中进行检索,即获得来自元江普通野生稻和合系35中的xa47(t)基因。
19.优选的,所述引物包括214qc-2f/r、48qc-9f/r和48qc-11f/r,所述214qc-2f的核苷酸序列如seq id no.17所示,所述214qc-2r的核苷酸序列如seq id no.18所示;所述48qc-9f的核苷酸序列如seq id no.19所示,所述48qc-9r的核苷酸序列如seq id no.20所示;所述48qc-11f的核苷酸序列如seq id no.21所示,所述48qc-11r的核苷酸序列如seq id no.22所示。
20.优选的,所述xa47(t)
g252
、xa47(t)
yp
、xa47(t)
hx
以及xa47(t)
l234
基因即以其供体材料进行命名的xa47(t)基因;所述xa47(t)
g252
为上述的xa47(t)基因;所述xa47(t)
yp
基因的核苷酸序列如seq id no.15所示;所述xa47(t)
yp
基因的cds序列如seq id no.30所示;所述xa47(t)
hx
基因的核苷酸序列如seq id no.16所示;所述xa47(t)
hx
基因的cds序列如seq id no.32所示;所述xa47(t)
l234
基因的核苷酸序列如seq id no.27所示;所述xa47(t)
l234
基因的cds序列如seq id no.28所示。
21.本发明提供了一种高效分离带有野生稻血缘的抗白叶枯病基因及其家族成员的方法,利用pacbio smrt技术和dovetail hi-c技术成功地将元江普通野生稻和栽培稻合系35的基因组对应到水稻染色体上,并与日本晴基因组进行共线性分析,根据测序组装序列以及xa47(t)基因的定位情况,快速获得了元江普通野生稻和合系35中xa47(t)基因型。利用根据该基因型设计的引物扩增后的目的产物可直接进行测序获得目的基因的完整编码区,无需通过繁琐的载体连接、蓝白斑筛选以及酶切验证等步骤,该技术高效、快捷、省时省力,为分离新的基因提供了较好的借鉴方法。同时,本发明利用来自同一个亲本的多样化抗谱材料,高效快速地克隆到xa47(t)基因及其家族成员,避免了常规图位克隆的繁琐和耗时
费力,利用元江普通野生稻渗入系获得的家族成员,其遗传背景清晰,为后续研究家族成员的功能以及育种利用奠定了坚实的基础。
附图说明
22.图1为合系35与日本晴11号染色体共线性图。chr11表示11号染色体;29021106kb表示日本晴11号染色体总长度;29113644kb表示合系35中11号染色体总长度。
23.图2为元江普通野生稻5与日本晴11号染色体共线性图。chr11表示11号染色体;29021106kb表示日本晴11号染色体总长度;31935697kb表示元江普通野生稻11号染色体总长度。
24.图3为214qc-9f/r在元江普通野生稻渗入系g252中的扩增电泳图。m为dl5000的marker,由5000bp、3000bp、2000bp、1000bp、750bp、500bp、250bp和100bp共8条链组成。
25.图4为xa47(t)基因部分区域测序峰图。
26.图5为xa47(t)
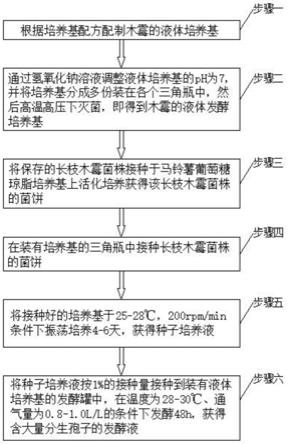
yp
、xa47(t)
g252
、xa47(t)
hx
以及xa47(t)
l234
4个成员的蛋白结构域。
27.图6为xa47(t)
l234
成员与xa47(t)
yp
和xa47(t)
hx
成员的比对图;其中,xa47_hx表示xa47(t)
hx
成员,xa47_yp表示xa47(t)
yp
成员,xa47_l234表示xa47(t)
l234
成员。
28.图7为xa47(t)
g252
成员与xa47(t)
yp
和xa47(t)
hx
成员的比对图;其中,xa47_hx表示xa47(t)
hx
成员,xa47_yp表示xa47(t)
yp
成员,xa47_g252表示xa47(t)
g252
成员。
具体实施方式
29.本发明提供了一种高效分离带有野生稻血缘的抗白叶枯病基因的方法,所述方法包括:
30.通过对元江普通野生稻渗入系双亲进行建库和高通量测序获得连续基因组,将所述连续基因组与参考基因组进行共线性分析,根据目的基因定位情况,筛选出所述抗白叶枯病基因的候选基因,根据所述候选基因设计引物进行pcr扩增、pcr产物直接测序和抗病功能分析,从而分离出带有野生稻血缘的抗白叶枯病基因。
31.本发明通过对元江普通野生稻渗入系双亲进行建库和高通量测序获得连续基因组。本发明中,所述建库和高通量测序之前优选的包括对元江普通野生稻渗入系双亲的dna提取;所述提取的方法优选为ctab法;所述元江普通野生稻渗入系双亲优选为元江普通野生稻和栽培稻合系35。本发明中,所述建库和高通量测序优选的包括pacbio sequel_smrt平台、dovetail hi-c建库技术和mecat组装系统。
32.本发明将所述连续基因组与参考基因组进行共线性分析,筛选出所述抗白叶枯病基因的候选基因。本发明中,所述参考基因组为日本晴基因组;所述共线性分析优选的采用mcscanx软件进行。本发明中,所述筛选的方法优选包括:利用soft berry(http://linux1.softberry.com/)中的fgenesh程序对r13i14和hxjy-14两个标记之间的序列进行基因注释分析,在smart数据库(http://smart.embl-heidelberg.de/)预测注释基因的功能,根据注释的功能筛选出xa47(t)候选基因。本发明中,所述抗白叶枯病基因为xa47(t);所述基因的核苷酸序列如seq id no.13所示;所述基因xa47(t)的cds序列如seq id no.14所示。
33.本发明根据所述候选基因设计引物进行pcr扩增、pcr产物直接测序和抗病功能分
析,从而分离出带有野生稻血缘的抗白叶枯病基因。本发明中,所述目的基因定位和所述pcr扩增的模板优选为元江普通野生稻渗入系g252。所述抗病功能分析优选的包括功能互补试验和基因编辑试验分析候选基因的抗病功能。本发明中,所述引物优选为214qc-9f/r;所述214qc-9f的核苷酸序列如seq id no.11所示,所述214qc-9r的核苷酸序列如seq id no.12所示。本发明中,所述测序的平台选择本领域常规测序平台即可。在本发明的实施例中,所述测序优选由华大生物科技(武汉)有限公司完成,pcr扩增产物测序优选由北京擎科生物技术有限公司昆明分公司完成。
34.本发明还提供了一种高效分离带有野生稻血缘的抗白叶枯病基因家族成员的方法,所述方法包括:
35.筛选获得含有xa47(t)基因或其同源基因的元江普通野生稻渗入系材料;根据上述xa47(t)基因序列,获得来自元江普通野生稻和合系35中的xa47(t)基因,分别命名为xa47(t)
yp
和xa47(t)
hx
;
36.利用引物对所述元江普通野生稻渗入系材料进行扩增、测序,得到4种xa47(t)基因的基因型:xa47(t)
yp
、xa47(t)
g252
、xa47(t)
hx
以及xa47(t)
l234
;对由4种基因型编码得到的蛋白质进行相似性分析,从而确定其是否为带有野生稻血缘的抗白叶枯病基因家族成员。
37.本发明中,所述筛选的步骤包括:利用xa47(t)基因的共分离标记hxjy-1对元江普通野生稻渗入系进行pcr扩增,得到扩增片段大小为167bp的渗入系,结合渗入系的抗病谱筛选出含有xa47(t)基因或其同源基因的元江普通野生稻渗入系材料。本发明中,根据所述xa47(t)基因序列,利用blast程序从元江普通野生稻和合系35基因组r13i14和hxjy-14区段中进行检索,即获得来自元江普通野生稻和合系35中的xa47(t)基因。本发明中,所述引物包括214qc-2f/r、48qc-9f/r和48qc-11f/r,所述214qc-2f的核苷酸序列如seq id no.17所示,所述214qc-2r的核苷酸序列如seq id no.18所示;所述48qc-9f的核苷酸序列如seq id no.19所示,所述48qc-9r的核苷酸序列如seq id no.20所示;所述48qc-11f的核苷酸序列如seq id no.21所示,所述48qc-11r的核苷酸序列如seq id no.22所示。
38.本发明中,所述xa47(t)
g252
、xa47(t)
yp
、xa47(t)
hx
以及xa47(t)
l234
基因即以其供体材料进行命名的xa47(t)基因;所述xa47(t)
g252
为上述的xa47(t)基因;所述xa47(t)
yp
基因的核苷酸序列如seq id no.15所示;所述xa47(t)
yp
基因的cds序列如seq id no.30所示;所述xa47(t)
hx
基因的核苷酸序列如seq id no.16所示;所述xa47(t)
hx
基因的cds序列如seq id no.32所示;所述xa47(t)
l234
基因的核苷酸序列如seq id no.27所示;所述xa47(t)
l234
基因的cds序列如seq id no.28所示。
39.本发明中,所述的栽培稻合系35为我国选育的高产优质粳稻品种,公布于文献(殷富有,张敦宇,叶玉,等.普通野生稻栽培稻杂交后代白叶枯病抗性评价.江西农业学报,2010,22(8):81-84.)。
40.所述的hzhj19、pxo99、pb、scyc-b、t7147、y8、ym1、ym187、yn24、yjdp-2和yjws-2白叶枯病菌在非专利文献公开,申请人有保存,自本专利申请日起20年内可提供。
41.所述的hzhj19、ym1、ym187、yn24、yjdp-2和yjws-2菌株为本实验室在2013~2017年从自然发病的病叶上分离而来,公布于文献(陈玲,张敦宇,陈越,等.云南药用野生稻种质资源的白叶枯病抗性评价.南方农业学报,2019,50(7):1417-1425.)。
42.所述的pxo99菌株为菲律宾标准菌株6,采集自菲律宾,公布于文献(ji ch,ji zy,
liu b,et al.xa1 allelic r genes activate rice blight resistance suppressed by interfering tal effectors.plant comm,2020,1(4),100087.)。
43.所述的pb菌株,为pxo99菌株的突变株系,公布于文献(陈玲,张敦宇,陈越,等.云南药用野生稻种质资源的白叶枯病抗性评价.南方农业学报,2019,50(7):1417-1425.)。
44.所述的y8菌株为云南强致病型生理小种,采集自中国云南,公布于文献(殷富有,张敦宇,叶玉,等.普通野生稻栽培稻杂交后代白叶枯病抗性评价.江西农业学报,2010,22(8):81-84.)。
45.所述的scyc-b(中国标准菌株5号)和yn24(中国标准菌株9号)菌株,采集自中国东北稻区,公布于文献(吴宪,许晶,温嘉伟,等.东北水稻白叶枯病菌株遗传多样性分析及品种对白叶枯病抗性评价.吉林农业大学学报,2015,37(3):290-295.)。
46.所述的t7147(日本标准菌株2号),采集自日本,公布于文献(周永力,翟文学,章琦,等.xa21转基因水稻对白叶枯病的抗性及其遗传.植物病理学报,2001,31(2):123-129.)
47.所述的栽培稻jg30,是我国选育的矮秆中晚熟籼稻(栽培稻),公布于文献(金旭伟,王春连,杨清,等.水稻抗白叶枯病近等基因系cbb30的培育及xa30(t)的初步定位.中国农业科学,2007,40(6):1094-1100.)。
48.为使本发明的目的、技术方案和优点更加清楚明白,下面结合实施例对本发明进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
49.下述实施例中,为了避免因不必要的细节而模糊了本发明专利的技术方案,在实施例中仅出示了与本发明专利方案密切相关的技术方案和/或处理步骤,如无特殊说明,省略的其他细节方法均为常规方法。
50.下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
51.实施例1元江普通野生稻和栽培稻合系35基因组高通量测序
52.1.元江普通野生稻和栽培稻合系35的基因组dna提取
53.基于ctab方法提取基因组dna:采取样品的幼嫩叶片约200mg,用液氮研磨成粉状,将组织转移到预热65℃的2.0ml管中,加入900μl 2%ctab裂解缓冲液,涡旋混合。将离心管在65℃下孵育60分钟,然后在室温(rt)下以10,000rpm离心5分钟。用900μl体积的苯酚/氯仿/异戊醇(25:24:1)提取上清液,重复2次,然后在新管中以10,000rpm的速度在室温下离心10分钟。加入2/3体积的预冷(-20℃)异丙醇,-20℃放置2小时以上沉淀dna,室温12000rpm离心15分钟。加入75%无水乙醇洗涤沉淀并离心去除,然后将dna沉淀风干3-5分钟。用200μl ddh2o溶解沉淀后进行后续实验。
54.所述2%ctab裂解缓冲液的配制方法为:取4g十六烷基三甲基溴化铵(ctab),16.364gnacl,1m tris-hcl 20ml(ph8.0),0.5m edta8ml,先用70ml ddh2o溶解,再定容至200ml灭菌备用。
55.2.元江普通野生稻和栽培稻合系35染色体级别的连续基因组获取
56.将步骤1提取得到的dna送华大生物科技(武汉)有限公司,通过pacbio sequel_smrt平台进行建库和全基因组测序,结合dovetail hi-c建库技术以及mecat组装系统,获得元江普通野生稻和栽培稻合系35染色体级别的连续基因组。
57.实施例2xa47(t)的克隆
58.1.xa47(t)候选基因的预测
59.选用mcscanx软件将实施例1中元江普通野生稻和栽培稻合系35的11号染色体的连续基因组与高质量参考基因组即日本晴基因组进行共线性分析。随后利用soft berry(http://linux1.softberry.com/)中的fgenesh程序对r13i14和hxjy-14两个标记之间的序列进行基因注释分析,在smart数据库(http://smart.embl-heidelberg.de/)预测注释基因的功能,根据注释的功能筛选出xa47(t)候选基因。
60.根据xa47(t)基因位于分子标记r13i14和hxjy-14之间的事实,通过共线性分析得到图1和图2。从图1和图2可看出,xa47(t)基因的此定位区段在元江普通野生稻、合系35和日本晴之间排列顺序比较一致,表明可参照目前研究较为清楚的日本晴基因组从该区段进行xa47(t)基因的预测。进而根据上述步骤从该区域注释到3个候选基因,其中一个是含有nbs-lrr结构域的loc_os11g46200基因,其余两个基因与转座相关,与抗病不相关,因此将loc_os11g46200作为xa47(t)基因的候选基因。
61.2.xa47(t)候选基因的克隆
62.根据步骤1得到的候选基因的编码区上下游区域,设计10对特异引物,采用ctab法提取xa47(t)的供体亲本g252的基因组dna,利用pcr进行扩增,取7μlpcr扩增产物在1%琼脂糖凝胶上5v/cm恒压电泳,凝胶成像系统成像后,根据pcr产物,筛选出一个特异引物214qc-9f/r,其中,214qc-9f的核苷酸序列如seq id no.11所示,214qc-9r的核苷酸序列如seq id no.12所示,该引物的扩增片段大小正确且带型清晰,结果如图3所示。将该pcr产物直接送北京擎科生物技术有限公司昆明分公司进行测序,测序结果如图4所示。由图4可见,测序峰图信号强,峰型完整且为单峰,表明测序结果较好。
63.所述的pcr扩增采用45μlpcr反应体系进行;45μlpcr反应体系为:max master mix 24μl,10μmol/l上、下游引物各2.4μl,20ng/μl模板dna 6μl,ddh2o 10.2μl;pcr反应条件为:94℃预变性3min;94℃变性15sec,55℃复性15sec,68℃延伸5min,共35个循环;68℃延伸10min。
64.测序结果显示,xa47(t)候选基因的核苷酸(dna)长度为4240bp,编码区(cds)序列为2409bp,编码802个氨基酸,具体核苷酸序列见序列表seq id no.13,具体cds序列见序列表seq id no.13编码获得的氨基酸序列如seq id no.34所示。
65.实施例3候选基因功能分析
66.1.功能互补试验验证候选基因功能
67.提取xa47(t)的供体亲本g252的总rna,反转录为cdna后,参照xa47(t)候选基因编码区序列,利用软件bioxm2.6从xa47(t)的dna序列中预测出其编码区(cds区),在预测出的编码区两端设计特异引物g252-cds-f/r,利用g252的cdna进行pcr扩增,将预期大小正确的片段与载体pce2连接,将连接产物送北京擎科生物技术有限公司昆明分公司进行测序,获得xa47(t)编码区序列,同时获得pce2-xa47cds质粒。
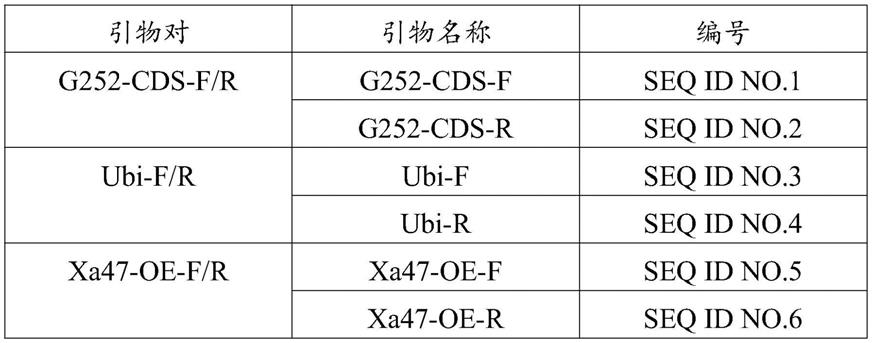
68.选择pcambia1305质粒作为植物表达载体骨架,用xbai和bsteii对其进行双酶切,利用引物ubi-f/r扩增质粒pjet-ubi中的ubiqutin启动子,利用xa47-oe-f/r引物扩增质粒pce2-xa47cds中的xa47(t)cds序列,切胶回收目的片段后,通过同源重组的方法构建ubi-xa47(t)过表达载体,通过农杆菌介导法转化感白叶枯病的栽培稻jg30材料。本实施例所采用的引物序列如下表1所示。
69.表1引物序列
[0070][0071]
对上述转化栽培稻jg30材料后的t1代阳性转基因苗进行孕穗期接种hzhj19、pxo99、pb、scyc-b、t7147、y8、ym1、ym187、yn24、yjdp-2和yjws-2共11个白叶枯病强致病菌,每个菌株接种3株转化苗,同时以感病材料金刚30作为对照,接种21天后调查病斑长度,结果如表2所示。其中,病斑长度的单位为cm。
[0072]
表2 t1代遗传转化苗ubi-xa47(t)表型鉴定
[0073]
[0074]
[0075][0076]
由表2可以看出,感病材料金刚30病斑在12cm以上,表现较强的感病特性,转化苗的病斑在5cm以下,对11个菌株达到抗到高抗的水平,说明遗传转化株系的白叶枯病抗性是由xa47(t)基因提供,该基因具有很强的白叶枯病抗性,对于抗白叶枯病水稻育种具有重要应用价值。
[0077]
2.基因编辑实验评估候选基因功能
[0078]
根据crispr-p2.0在线软件(http://crispr.hzau.edu.cn/crispr2)在xa47(t)整个dna序列(正反链)里找出含有gn20gg或n20gg的pam序列,选择中靶效率评分大于0.5的特异性较高的作为sgrna候选序列,通过查找共找到3个sgrna序列,将3个sgrna序列进行体外验证。将体外验证较好的sgrna-2的pam前面20bp的序列作为spacer序列,分别设计上、下游sgrna的oligo序列sgf/r,并在sgf的5'端加入同源臂ggca,sgr的5'端加入同源臂aaac,送北京擎科生物技术有限公司昆明分公司合成。
[0079]
利用合成的oligo序列制备两端具有粘性末端的双链dna片段(oligo-xa47(t))。利用cip酶切10ug pos-sgrna载体,制作出线性化pos-sgrna载体。将线性化pos-sgrna载体与oligo-xa47(t)相连,转化大肠杆菌,获得阳性克隆,构建出pos-sgrna-oligo-xa47(t)载体。利用lr clonase
tm ii enzyme mix将pos-sgrna-oligo-xa47(t)质粒与ph-ubi-cas9-7质粒进行lr反应,最终获得crispr/cas9-xa47(t)载体,并用引物seq-u3和seq-cas对crispr/cas9-xa47(t)载体进行测序来验证其正确性,测序公司为北京擎科生物技术有限公司昆明分公司。将验证正确的载体通过农杆菌介导法转化xa47(t)的供体亲本g252。本实施例所用引物序列如表3所示。
[0080]
表3引物序列
[0081][0082][0083]
对转化后的t1代阳性基因敲除苗进行孕穗期接种上述11个白叶枯病菌株,供体亲本g252作为对照,结果显示g252对11个菌株均抗病,敲除苗对11个菌株都感病,该结果与过表达植株的表型相对应,表明本发明所述基因xa47(t)抗不同的白叶枯病菌,具有广谱高抗的特性。
[0084]
实施例4 xa47(t)家族成员的分离
[0085]
本实施例所述的pcr扩增程序和体系与实施例1相同。
[0086]
1.xa47(t)基因家族成员的供体亲本筛选
[0087]
利用xa47(t)的共分离标记hxjy-1在359份元江普通野生稻渗入系中进行pcr扩增,筛选出扩增片段大小为167bp的渗入系有262份,且这262份渗入系抗病谱依然为67种类型,为了获得对白叶枯病菌具有广谱高抗的抗性基因,本实施例从262份渗入系中筛选出抗6-9个白叶枯病菌的渗入系,这些渗入系呈现出16个抗病谱,从16个抗病谱中选出1~3份代表材料用于xa47(t)基因家族成员的分离,具体的材料抗病谱如表4。
[0088]
表4部分渗入系抗病谱
[0089]
[0090]
[0091][0092]
2.元江普通野生稻和合系35基因的xa47(t)基因获取
[0093]
根据xa47(t)基因序列,利用blast程序从元江普通野生稻和合系35基因组r13i14和hxjy-14区段中进行检索,获得来自元江普通野生稻和合系35基因的xa47(t)基因。得到的xa47(t)
yp
基因序列如seq id no.15所示,xa47(t)
hx
基因如seq id no.16所示。利用dnaman软件比对发现,xa47(t)
hx
基因与loc_os11g46200的编码区完全一致,相似性为100%。
[0094]
3.xa47(t)基因家族成员的基因分离
[0095]
根据xa47(t)
yp
和xa47(t)
g252
基因的编码区上下游序列,设计特异引物,以及利用之前参照loc_os11g46200编码区上下游设计的10对特异引物,通过pcr技术在元江普通野生稻渗入系dna中进行扩增,取7μlpcr扩增产物在1%琼脂糖凝胶上5v/cm恒压电泳,凝胶成像系统成像后,根据跑胶结果,共筛选出4对可从元江普通野生稻渗入系dna中扩增的引物,引物序列如下表5,选取片段大小正确的pcr产物直接送北京擎科生物技术有限公司昆明分公司进行测序。
[0096]
表5引物序列
[0097][0098]
根据测序结果发现渗入系中出现了4种基因型,包括xa47(t)
yp
、xa47(t)
g252
、xa47(t)
hx
以及xa47(t)
l234
基因型,通过比对,xa47(t)
hx
与日本晴中的loc_os11g46200基因完全一致,而loc_os11g46200基因的cds区域已经明确。xa47(t)
l234
基因的核苷酸序列如seq id no.27所示。利用软件bioxm2.6从xa47(t)
yp
和xa47(t)
l234
基因型的dna序列中预测出其cds区,在预测出的编码区两端设计特异引物对yp-cds-f/r(其中yp-cds-f的核苷酸序列如seq id no.23所示,yp-cds-r的核苷酸序列如seq id no.24所示)和l234-cds-f/r(其中l234-cds-f的核苷酸序列如seq id no.25所示,l234-cds-r的核苷酸序列如seq id no.26所示),这两对引物分别在元江普通野生稻和渗入系l234的cdna中进行扩增,将预期大小正确的片段与载体pce2连接,将连接产物进行测序,获得xa47(t)
yp
和xa47(t)
l234
编码区序列。根据4个成员的cds序列通过bioxm2.6软件将其分别翻译成蛋白质序列,具体的序列如表6所
示。继续利用dnaman软件分析4个蛋白之间的相似性,结果如表7所示。进一步利用在线软件smart预测4个蛋白结构,结果如图5所示。
[0099]
表6不同成员的序列
[0100][0101]
表7不同基因型蛋白相似性分析
[0102] xa47(t)
hx
xa47(t)
yp
xa47(t)
g252
xa47(t)
l234
xa47(t)
hx
100%100%//xa47(t)
yp
95.2%100%//xa47(t)
g252
89.0%91.9%100%/xa47(t)
l234
94.8%99.0%92.1%100%
[0103]
从表7可见,4个蛋白之间相似性较高,说明它们极有可能互为同一个家族成员。从图5可见,4个蛋白结构域基本一致,综合说明4个基因型同为一个基因家族。那4个家族成员中,因xa47(t)
yp
和xa47(t)
hx
分别来自渗入系的双亲,说明xa47(t)
yp
是完全带有野生稻血缘的基因型,xa47(t)
hx
是来自栽培稻合系35。
[0104]
随后利用dnaman软件将xa47(t)
g252
和xa47(t)
l234
蛋白分别与xa47(t)
yp
和xa47(t)
hx
进行多重比对,确定xa47(t)
g252
和xa47(t)
l234
是否带有野生稻血缘,结果如图6和图7所示。
[0105]
可以看出,相对于xa47(t)
yp
和xa47(t)
hx
蛋白,xa47(t)
g252
和xa47(t)
l234
蛋白已发生变异,其中xa47(t)l
234
蛋白变异较xa47(t)
g252
小,但2个成员的大部分蛋白区域还是与xa47(t)
yp
一致,说明xa47(t)
g252
和xa47(t)
l234
基因型携带了野生稻血缘。
[0106]
本实施例从元江普通野生稻渗入系中获得了4个xa47(t)家族成员,其中的3个成员都带有野生稻血缘,是具有利用价值的基因资源。所述的基因家族指来源于同一个祖先,由一个基因通过基因重复而产生两个或更多的拷贝而构成的一组基因,它们在结构和功能上具有明显的相似性,编码相似的蛋白质产物。
[0107]
4.获得的家族成员在元江普通野生稻渗入系中的分布情况
[0108]
根据上述结果分析4个成员在元江普通野生稻渗入系中分布情况,结果如表8所示。
[0109]
表8 xa47(t)基因不同家族成员在元江普通野生稻中的分布情况
[0110][0111][0112]
可以看出,这些家族成员在渗入系中以纯合或杂合状态存在,表明元江普通野生稻渗入系中的xa47(t)具有多样化,即存在一定变异也存在一定的保守型,它们互为基因家族,表明利用本发明所述方法还可从其余的渗入系材料中分离出新的家族成员。
[0113]
以上所述仅为本发明的实施例,并非因此限制本发明的专利范围,凡是利用本发明说明书及附图内容所作的等效结构或等效流程变换,或直接或间接运用在其他相关的技术领域,均同理包括在本发明的专利保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。