一种mlo基因突变体及其制备方法与应用
技术领域
1.本发明属于生物技术领域,尤其涉及一种mlo基因突变体及其制备方法与应用。
背景技术:
2.油菜是我国种植面积最大的油料作物,保障油菜产业的健康发展是稳定我国食用油供给 安全重要基石。油菜菌核病是油菜生产中的主要病害,年均发病率约为10%至30%,造成的 年均产量损失约为10%至20%,危害严重的年份减产可以达到80%以上。白粉病是油菜的另 外一种重要的病害,特别是在长江流域主要的油菜种植区,雨水充沛,极容易造成油菜白粉 病的大规模爆发,产生的病原菌孢子严重影响了菜籽的品质和收获指数。
3.由于油菜种质资源中缺乏菌核病的抗源种,严重阻碍了油菜抗病性的分子遗传改良进程。 基因编辑技术是一种新型的基因组操作工具,可以同时对多倍体作物中多个同源拷贝的基因 功能进行细致解析。同时产生的突变体材料可以剔除转基因元件,获得稳定遗传、不含转基 因元件的突变材料,具有较强的应用前景。
4.mlo(mildew resistance locus)是一类具有跨膜结构的基因家族。前期研究表明,不同 物种中mlo基因的家族成员数目存在较大差异,如模式植物拟南芥共有15个mlo基因家 族成员,小麦中有8个,大豆中有39个,葡萄中有17个。不同的家族成员之间的功能也分 化明显。atmlo2单个基因突变能够显著提高拟南芥对白粉病的抗性,是白粉病抗性的主效 负调控因子;atmlo6和atmlo12基因对于白粉病抗性也有部分辅助功能。对于其它的病原 菌mlo基因也表现出明显的功能分化。拟南芥mlo2mlo6mlo12三基因突变对炭疽病 (colletotrichum higginsianum)抗性略有增加,对灰霉病(botrytis cinerea)抗性明显增加, 对黑斑病(alternaria alternata)和叶斑病(alternaria brassicicola)的敏感性增加,而对印度 梨型孢(serendipita indica)和霜霉病(hyaloperonospora arabidopsidis)抗性却没有明显变化。 不同的物种之间,mlo基因的功能也不一样。大麦mlo基因突变体对小麦根腐病菌(bipolarissorokiniana)引起的根腐病敏感性增加,对真菌(ramularia collo-cygni)引起的红斑叶斑病敏 感性也增加;辣椒camlo2基因受到细菌性疮痂病(xanthomonas campetris pv.vesicatoria) 的诱导表达,沉默camlo2基因可以提高辣椒对细菌性疮痂病的抗性。因此植物mlo基因 家族在不同物种中的成员数,不同成员参与的抗病调节反应,单个基因在基因组中的拷贝数, 以及不同拷贝之间的功能可能都存在较大的差异。
5.油菜是一种异源四倍体作物,大多数基因一般存在2个和2个以上的拷贝,这些基因的 拷贝之间的功能往往存在分化和冗余的现象。我们利用生物信息学、分子生物学实验结合基 因编辑的方法,首次从油菜的基因组中鉴定到了bnmlo6参与了负向调控油菜菌核病的抗性。 bnmlo6基因的突变能够同时显著提高油菜对菌核病和白粉病的抗性,经检索,这在油菜中 为首次报道。
技术实现要素:
6.为了解决上述技术问题,本发明提供一种mlo基因突变体及其制备方法与应用。
7.本发明通过比较分析油菜接种菌核病前后的表达量,发现bnmlo6基因中有6个同源拷 贝受油病原菌诱导表达,推测它们可能参与了对菌核病的抗性反应,利用crispr/cas9基因 编辑的方法,我们获得了bnmlo6基因6个拷贝同时发生编辑的突变体株系。突变株系抗性 调查结果表明,bnaa03g60240d(mlo6-a03)、bnacnng04240d(mlo6-c03)、bnaa01g22680d (mlo6-a01)、bnac01g29530d(mlo6-c01)、bnaa09g13700d(mlo6-a09)和 bnacnng31840d(mlo6-c09)6个拷贝同时突变,可以显著提高油菜对菌核病和白粉病的抗 性。
8.为达上述目的,本发明采用如下的技术方案:
9.本发明的目的之一在于提供一种突变mlo6基因,在油菜darmor基因组的命名分别为 bnaa03g60240d(mlo6-a03)、bnacnng04240d(mlo6-c03)、bnaa01g22680d(mlo6-a01)、 bnac01g29530d(mlo6-c01)、bnaa09g13700d(mlo6-a09)和bnacnng31840d(mlo6-c09), 所述突变mlo6基因与野生型mlo6基因相比,mlo6-a03拷贝为纯合突变,缺失了15个 碱基,使得编码的氨基酸序列356号位点后缺失-l-v-q-p-g;mlo6-c03拷贝为缺失13个碱 基和缺失2个碱基的杂合突变,分别在氨基酸357号和356号位点后造成了移码突变; mlo6-a01拷贝为缺失2个碱基和单碱基g插入的杂合突变,都在365号氨基酸位点后造成 了移码突变;mlo6-c01拷贝的一条链缺失12个碱基,在氨基酸357号位点将l替换成了r, 随后丢失-v-q-p-g四个氨基酸;mlo6-a09拷贝的一条链缺失3个碱基,在氨基酸356号缺 失了v;mlo6-c09拷贝一条链缺失2个碱基,在氨基酸355号位点后面造成了移码突变。
10.本发明的目的之二在于提供一种上述突变mlo6基因编码的多肽。
11.本发明的目的之三在于提供一种构建体,其包含上述的突变mlo6基因。
12.本发明的目的之四在于提供一种重组细胞,所述重组细胞是通过上述的构建体转化受体 细胞获得的。
13.本发明的目的之五在于提供一种获得上述突变mlo6基因的方法,其包括如下步骤:
14.(1)设计bnmlo6-sgrna2,序列如seq id no.30所示;
15.(2)将步骤(1)中的sgrna2构建至载体质粒骨架中;
16.(3)将构建的基因编辑载体导入油菜受体材料中,并获得再生苗;
17.(4)转基因植株的鉴定和突变检测。
18.进一步的,所述步骤(2)中的载体质粒骨架为pkse401。
19.进一步的,所述步骤(3)中将构建的基因编辑载体导入油菜受体材料中是采用农杆菌介 导的油菜胚轴遗传转化方法。
20.本发明的目的之六在于提供上述突变mlo6基因在植物抗菌核病中的应用。
21.与现有技术相比,本发明具有如下有益效果:
22.通过本研究,我们证明了crispr/cas9系统是甘蓝型油菜复杂基因组编辑的有效工具, 可以成功敲除了油菜目标基因的多个同源拷贝。对油菜中感病基因bnmlo6的敲除,能够激 发叶片细胞中胼胝质的自发堆积,调节乙烯和茉莉酸抗病信号路径。因此,bnmlo6基因在 油菜中可能参与了多条抗病信号路径,负向调控对白粉病和菌核病的抗性,为利用
bnmlo6 基因协同调控油菜多种病原菌抗性的研究提供了参考。研究获得的基因突变产生的抗病材料, 对油菜的抗病性相关基础研究和抗病育种都具有重要意义。
附图说明
23.图1为实施例1中bnmlo6基因的6个拷贝的表达量检测结果图。
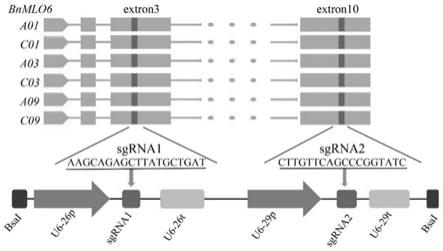
24.图2为实施例1中两个靶点sgrna1和sgrna2的示意图。
25.图3为实施例1中bnmlo6-c01拷贝基因page电泳分型部分结果图。
26.图4为实施例1中mlo6-212单株t0代bnmlo6基因靶点突变结果图。
27.图5a为实施例1中田间mlo6-212的t1代植株白粉病感染结果图。
28.图5b为实施例1中室内mlo6-212的t1代植株白粉病感染结果图。
29.图6为实施例1中mlo6-212t1代胼胝质观察及分析结果图。
30.图7a为实施例1中核盘菌侵染24小时后的结果图。
31.图7b为实施例1中核盘菌侵染36小时后的结果图。
32.图7c为实施例1中叶片菌斑面积分析图。图8为实施例1中多个基因在野生型和mlo6-212突变体t2代接菌后的表达变化图。
具体实施方式
33.以下实施例中遗传转化受体材料为甘蓝型油菜中双6号,载体质粒骨架pkse401由中国 农业大学陈其军老师实验室提供,所用大肠杆菌dh5a和农杆菌gv3101感受态购买于唯地 生物技术有限公司,核盘菌由中国农科院油料所刘胜毅老师课题组提供,引物由金开瑞生物 工程有限公司合成,测序由奥科科技有限公司完成。
34.实施例1
35.1.试验方法
36.(1)半定量rt-pcr
37.取未接种和接种菌核菌后不同时间的中双6号叶片,采用天根公司rna提取试剂盒提取 rna,操作方法参照说明书,用分光光度计估算rna浓度。按照全式金反转录试剂盒说明 书操作合成第一链cdna。以第一链cdna为模板,用bnmlo6基因各同源拷贝的特异引物 进行pcr扩增,bnactin基因作为内参,用1%的琼脂糖凝胶电泳分离鉴定,其中引物序列如 表1。
38.表1 引物序列
[0039][0040]
(2)sgrna设计与载体构建
[0041]
利用拟南芥mlo6基因蛋白序列,在油菜基因组数据库(https://www.genoscope.cns.fr/brassicanapus/)中进行blast比较分析,共鉴定出六个油菜mlo6 基因的同源拷贝,分别为bnaa03g60240d(mlo6-a03)、bnacnng04240d(mlo6-c03)、 bnaa01g22680d(mlo6-a01)、bnac01g29530d(mlo6-c01)、bnaa09g13700d(mlo6-a09) 和bnacnng31840d(mlo6-c09),其对应的核苷酸序列分别为seq id no.53-58所示。采 用sgrna靶点设计网站(http://crispr.hzau.edu.cn/crispr2/)分别在六个基因拷贝的第三个外 显子和第十个外显子的保守序列位置,设计了两个sgrna,分别为bnmlo6-sgrna1(seq idno.29:aagcagagcttatgctgat)和bnmlo6-sgrna2(seq id no.30:cttgttcagc ccggtatc)。两个靶点采用的方式构建到基因编辑终载体中(参见ma x,zhang
factor 1)、 erf2(ethylene response factor 2)、pr4(pathogenesis-related 4)和ora059(octadecanoid
‑ꢀ
responsive arabidopsis ap2/erf 59)接种核盘菌后基因的表达情况,引物见表2。
[0054]
表2 防御相关基因引物
[0055][0056][0057]
2.试验结果
[0058]
(1)接种核盘菌后bnmlo6基因的表达分析和crispr/cas9基因编辑载体构建
[0059]
通过rt-pcr对接种后基因拷贝的表达量检测表明,bnmlo6基因的6个拷贝的表达量在接种后均呈 现不同升高的趋势,结果参见图1,其中a01/c01/a03/c03拷贝表达量上升更为明显。初步推定bnmlo6 基因可能与油菜菌核病抗性相关。
[0060]
为了进一步研究基因的功能,在bnmlo6基因的第3个和第10个外显子的保守区间分别设计了两个 靶点sgrna1和sgrna2,参见图2,并完成了crispr/cas9基因编辑载体的构建。
[0061]
(2)t0代基因突变分析
[0062]
对t0代植株进行nptii标记基因的检测,筛选转基因阳性植株。检测的494株再生苗中,阳性植株 137株,阳性率为27.7%。用page电泳法在137株阳性植株中筛选差异条带后,得到编辑植株41株,载 体的突变效率为29.9%,其中bnmlo6-c01拷贝基因page电泳分型部分结果参见图3。
[0063]
page分型得到的41株编辑植株中,有10株显示六个拷贝发生了带型的改变(图3),有可能发生 了突变。对这10个单株的突变位点进行pcr扩增后测序分析,发现在sgrna2靶点处,两个和四个基因 拷贝发生了编辑的各有2株,五个和六个拷贝发生了编辑的各有3株。
[0064]
在全部6个基因拷贝发生编辑的3株中,mlo6-212的突变情况最好,参见图4,其中a03拷贝为纯合 突变,缺失了15个碱基,使得编码的氨基酸序列356号位点后缺失-l-v-q-p-g。c03拷贝为缺失13个碱 基和缺失2个碱基的杂合突变,分别在氨基酸357号和356号位点后造成了移码突变。a01拷贝为缺失2 个碱基和单碱基g插入的杂合突变,都在365号氨基酸位点后造成了移码突变。c01拷贝的一条链缺失12 个碱基,在氨基酸357号位点将l替换成了r,随后丢失-v-q-p-g这四个氨基酸。a09拷贝的一条链缺失 3个碱基,在氨基酸356号缺失了v。c09拷贝一条链缺失2个碱基,在氨基酸355号位点后面造成了移 码突变。以上序列分析结果表明,dna序列上的突变造成了mlo6-212单株每个拷贝的氨基酸序列发生了 变化,可能会引起bnmlo6基因发生功能上的改变。
[0065]
(3)t1代突变分析及白粉病抗性观察
[0066]
任意选取mlo6-212单株的10个t1代单株提取dna,对其中的部分单株测序表明,这些t1代植株中 的六个bnmlo6基因拷贝的突变类型均与t0代一致,遗传了上一代的突变,没有发现新的突变类型,说 明基因编辑产生的突变能够有效地遗传给下一代。
[0067]
对转基因基地和地下室生长间种植的基因编缉突变后代及野生型的白粉病发病情况观察表明, bnmlo6基因的突变能够显著改善油菜对白粉病的抗性。基地中所有野生型植株茎秆上均披有一层白粉, 感染了很重的白粉病。但是mlo6-212的t1代植株上均看不见白粉,没有感染白粉病的痕迹(图5a)。 种植在室内植物生长间里的植株也具有同样的表型(图5b)。我们种植株系mlo6-212于汉川转基因基地, 对大田的白粉病发病情况进行了统计调查。由于2021年春季油菜开花和角果发育期阴雨天气多,田间油 菜的白粉病发病情况较为严重,几乎所有的野生型植株茎杆上都感染了白粉病病菌,发病率接近100%, 而bnmlo6基因的突变材料发病率为50-61%,极显著低于野生型材料。
[0068]
(4)bnmlo6基因突变材料的胼胝质分析
[0069]
对mlo6-212突变材料的t1代胼胝质显微镜观察结果表明,在没有白粉病感染的情况下,bnmlo6突 变材料自发地出现了胼胝质堆积(参见图6)。相比于野生型,突变材料中单个视野内观察到的胼胝质光 点数量,由7.33个增加到了35.29个,增加了486.3%。胼胝质的增加可能与bnmlo6基因突变引起的白 粉病抗性存在关联。
[0070]
(5)bnmlo6基因突变材料的菌核病抗性鉴定
[0071]
选取六个bnmlo6同源拷贝均有突变的t1单株mlo6-212的t2代种子,在生长间室内种植,五叶期后 取叶片进行核盘菌接种,以野生型为对照。接种鉴定结果表明(参见图7a-7c),bnmlo6突变材料的叶片 菌斑面积与野生型之间有显著性差异。在接种24小时后(图7a),相对于野生型,bnmlo6突变材料平均 菌斑面积从1.37cm2减少到了1.10cm2,减少了19.5%(p=0.012),达到显著水平;接菌36小时后(图7b), 相对于野生型,bnmlo6突变材料平均菌斑面积从3.95cm2减少到了3.45cm2,减少了12.5%(p=0.045), 也达到显著水平。
[0072]
(6)突变材料抗病相关基因表达分析
[0073]
利用rt-pcr鉴定erf1、erf2、pr4、ora059在野生型和mlo6-212突变体t2代接菌后的基因表达 变化情况。结果表明,突变材料中erf1基因的两个拷贝在未接菌和接菌24h后表达量高于野生型,接菌 36h后两者表达量趋于一致。erf2基因的三个拷贝在未接菌时,突变体和野生型表达量相差不大;但在接 菌24h和36h后,突变体中3个拷贝的表达量都明显高于野生型。pr4基因未接菌时,在突变体和野生型 中的表达量都很低;接菌24h和36h后,在野生型中pr4基因的4个拷贝几乎没有受到诱导,检测不到表 达量,但是在突变体中,pr4基因的4个拷贝表达量都明显提高。ora059基因未接菌时,表达量在突变 体和野生型中都很低,接菌后突变体和野生型中ora059基因的两个拷贝表达量都提高了,但是在突变体 中提高得更为显著(参见图8)。
[0074]
图8中每一行代表相关基因一个同源拷贝的表达量变化,每个从上到下,erf2的三个拷贝为: bnaa09g0212800zs、bnac02g0416900zs、bnac09g0247600zs;erf1两个拷贝为:bnaa01g0293300zs、 bnac01g0362300zs;pr4四个拷贝为:bnaa03g0296000zs、bnaa03g0296200zs、bnac03g0354600zs、 bnac03g0354700zs;oar059两个拷贝为:bnaa10g0042700zs、bnac05g0044000zs;从左到右样品依次 为:野生型未接菌,突变体未接菌,野生型接菌24h,突变体接菌24h,野生型接菌36h,突变体接菌36h。
[0075]
在多倍体植物中由于存在基因冗余,进行基因敲除往往需要多个拷贝同时编辑。甘蓝型油菜作为一种 异源四倍体作物,在大多数情况下携带一个基因的两个以上同源拷贝。crispr/cas9系统是甘蓝型油菜多 重基因组编辑的有效工具。我们利用darmor的基因组中bnmlo6同源基因的保守序列选择了两个靶点, 分别靶向油菜bnmlo6基因的6个同源拷贝的保守的外显子序列。突变检测结果发现,根据第一个靶点设 计的sgrna1没有在目标位点检测到编辑,而根据第二个靶点设计的bnmlo6-sgrna2实现了目标位点的 编辑突变。bnmlo6-sgrna2靶点的编辑效率较高,达到了29.9%。其中,六个拷贝同时突变的编辑植株 占编辑植株的7.32%(3/41),阳性植株的2.19%。从3株6个基因拷贝同时被编辑的单株中选出了突变 较为理想的单株mlo6-212进行后续的功能鉴定。
[0076]
油菜bnmlo6基因的多个基因拷贝同时突变后,t1和t2代株系在室内植物生长间和大田条件下都表 现出很强的白粉病抗性。在油菜bnmlo6编辑材料中出现了自发的胼胝质堆积,胼胝质的数量比野生型显 著增加。胼胝质除了在早期作为防止真菌病原体进入的物理屏障外,还可能形成扩散屏障,阻止病原菌的 进一步扩散,在植物抗病早期的机械防御中起到重要作用。可见,油菜bnmlo6基因突变后产生的白粉病 抗性,与细胞中胼胝质的积累量提高有关。
[0077]
关于mlo基因与菌核病抗性关系的研究尚未见报道。接种核盘菌后,油菜bnmlo6基因的6个同源 拷贝的表达量均检测到不同程度的增加,有可能参与了对菌核病菌的防御反
应。菌核病接种结果表明, mlo6-212突变体的平均菌斑面积在接种24h和36h后均显著小于野生型。该结果说明bnmol6基因可能 负向调控了早期的油菜对菌核病的防御反应。
[0078]
植物激素介导的信号通路sa和ja/et参与植物对不同营养型病原菌的防御,拟南芥mlo基因的突 变影响ja/et信号通路基因的表达。ja/et通路的标志基因erf1,erf2,pr4和ora59在接种核盘菌后 的bnmlo6突变体中的表达量提高,说明mlo基因敲除后促进了ja/et信号通路基因的表达,从而可能介 导了该信号通路的防御,在感病早期提高了对菌核病菌的抗性。
[0079]
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发 明设计精神的前提下,本领域技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利 要求书确定的保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。