茄子e3泛素连接酶基因smdda1b及其在提高青枯病抗性的应用
技术领域
1.本发明属于分子生物学技术领域,更具体地,涉及茄子e3泛素连接酶基因smdda1b及其在提高青枯病抗性的应用。
背景技术:
2.茄子(solanum melongena l.)是茄科(solanaceae)茄属(solanum l.)一年生或多年生蔬菜作物,起源于印度及中国西南部地区。是亚洲、地中海、中欧及东南欧各地广泛栽培的夏季蔬菜。茄子营养丰富,具蛋白质、脂肪、碳水化合物及多种微量元素,除此以外,还富含维生素p和龙葵碱,具有保护心脑血管、抗衰老等作用,其果可供蔬食,其根、茎、叶均可入药,叶还可以作麻醉剂。
3.但茄子易受病害侵袭,尤其是青枯病,可造成茄子品质下降,大量减产。茄子受青枯病病害时,一般减产20%~30%,严重时可达50%~60%。青枯病造成的重大经济损失和难以治理的现状,已严重威胁到茄科植物的生产和发展。研究发现,茄科许多作物的栽培品种不抗青枯病,生产上也没有抗青枯病的茄子品种抗病品种多为半栽培品种或野生种。目前,对茄科青枯病抗病性遗传的报道多样,至今未有定论。
4.目前已验证的茄子青枯病抗性相关基因较少,茄子抗青枯病涉及多种因子,分子机制较为复杂,仍不明确。xiao等验证茄子re
‑
bw基因为抗青枯病重要基因,且能与青枯菌无毒效应子popp2发生互作。邵欣欣研究发现茄子erf转录因子smerf参与抗青枯病过程,并获得茄子抗青枯病的smerf相关基因smerf66及smerf88。李可等通过基因沉默(vigs)技术验证茄子eds1基因对抗青枯病具有正调控的作用。同时,chen等对茄子smnac转录因子进行功能鉴定分析得出smnac负调控茄子抗青枯病抗性。qiu等研究表明外施亚精胺(spd)可显著提高茄子对青枯病的抗性,而r2r3
‑
myb转录因子smmyb44能够直接结合亚精胺合成酶基因spds的启动子并增强其表达,促进亚精胺的合成。
5.泛素(ubiquitin,ub)最早发现于小牛胸腺当中,是普遍存在于真核生物中的保守低分子量(8.5kda)蛋白质,由76个氨基酸组成。泛素几乎参与真核生物的所有方面,包括生长发育和免疫调节。泛素/26s蛋白酶体系统(ubiquitin/26s proteasome system,ups)是主导泛素化的保守体系,由泛素、泛素激活酶(e1)、泛素结合酶(e2)、泛素连接酶(e3)和26s蛋白酶体5个基本部分组成,中心成分是泛素分子,泛素通过e1、e2和e3相互作用,与靶蛋白结合,由此改变蛋白质组成而调控真核生物的功能。在泛素系统中,e3连接酶的基因数量相对较高,且在识别底物特异性方面具有重要作用。
6.不同于e1激活酶和e2结合酶,e3连接酶大量且多样。据现有报道,e3连接酶根据其结构特征域及传递泛素至靶蛋白的作用机制,主要被分为三个家族,hect e3s,ring e3s,rbr e3s。其中,ring型家族最大,含有一个被称为ring的锌结合结构域或u
‑
box结合域。从结构上看,ring结构域和u
‑
box结构域类似,但u
‑
box结构域主要通过盐桥、离子螯合以及氢键从e2上转移泛素至靶蛋白。而ring结构域主要通过8个氨基酸与锌离子的螯合形成共价
键来转移泛素。crls(cullin
‑
ring
‑
ligases)是多亚基复合体,也是ring型家族中最大的家族,其主要由cullin蛋白、ring蛋白、底物受体以及adaptor蛋白组成。cullin蛋白为核心支架,c端锚定ring蛋白,n端与crl的adaptor互作,adaptor帮助crl连接底物受体。泛素化修饰通过构象激活crls,更改cullin
‑
ring接口,与nedd8的共价连接来进行翻译后修饰,以激活连接酶活性。
7.研究表明,e3连接酶不仅调控植物的生长发育,还在植物抗胁迫方面发挥着重要的作用。在拟南芥中,miel1为ring型e3连接酶,是防御反应的负调节因子。stpub17作为一种e3连接酶,是e3s stpob1的已知靶蛋白,且已被证实其正调控植物抗晚疫病等病害,近期研究表明stpub17的降解底物为rna结合蛋白stkh17,该基因负调控植物免疫,且stkh17功能需结合rna行使。ups也可作为一种正调控因子参与应激反应。烟草中,e3连接酶nbube3r1正调控免疫反应,通过vigs、y2h及co
‑
ip实验表明bamv的复制酶为nbube3r1的可能底物。desaki等发现正调控植株免疫反应的e3泛素连接酶pub4可与lysm受体样激酶cerk1发生互作。
8.此外,e3连接酶在植物中介导多种植物信号通路,包括激素等。hana等研究表明,苹果u
‑
box e3s mdpub29通过调控sa途径提高植物对苹果轮纹病菌(botryosphaeria dothidea)的抗性。而苹果中btb
‑
back域e3s mdpob1通过降解mdpub29来调控苹果h2o2含量、sa相关基因及其信号基因的相对表达量。tir1是scftir1连接酶的底物识别亚基,可对aux/iaa蛋白的转录抑制因子进行蛋白降解。sa途径信号基因npr1可能为cul3的泛素化底物,两者通过adaptor蛋白间接相互作用。在拟南芥中,e3泛素连接酶atarre可负调控aba信号。
9.e3泛素连接酶存在于ups,在植物细胞降解蛋白的途径中起重要作用,且多有研究表明e3连接酶在植物抗病中发挥作用。茄子是重要的经济作物,但易受青枯病侵害,严重影响其经济价值,对于茄子抗青枯病的研究较多,但关于茄子e3泛素连接酶抗青枯病的报道几乎没有,因而研究e3泛素连接酶基因对茄子青枯病抗性,可为茄子抗青枯病提供理论参考及育种价值。
技术实现要素:
10.本发明所要解决的技术问题是克服现有技术中存在的上述问题,首先提供茄子e3泛素连接酶基因smdda1b。
11.本发明的第二个目的是提供茄子e3泛素连接酶基因smdda1b的应用。
12.本发明的第三个目的是提供一种提高茄子青枯病抗性的方法。
13.本发明的目的通过以下技术方案实现:
14.茄子e3泛素连接酶基因smdda1b,其核苷酸序列如seq id no:1所示,编码的氨基酸序列如seq id no:2所示。smdda1b的orf为504bp,167个氨基酸,18.27kda,具dda1结构域,属于crl的ddb型e3泛素连接酶的主要组成部分。
15.本发明还提供含有所述基因smdda1b的生物材料,所述生物材料包括但不限于载体、质粒、宿主细胞、植物。
16.在本研究中,本课题组以茄子抗青枯病的负调控转录因子smnac n端包含nam结构域的417bp碱基为诱饵,对茄子cdna文库进行筛选后获得e3泛素连接酶基因smdda1b,并进行功能验证。vigs等实验初步筛选出结果最明显,且之前未被进行功能验证的e3泛素连接
酶基因smdda1b,之后对其进行过表达实验,再次验证了其确实对植物抗青枯病具有正调控作用,推测为茄子抗青枯病的重要基因。
17.因此,本发明还提供上述茄子e3泛素连接酶基因smdda1b在开发、筛选茄子对青枯病抗性的功能产品的应用。
18.优选的,所述功能产品具有上调smdda1b基因的表达、转录或者其表达产物的功能。
19.更优选的,所述功能产品选自用于促进smdda1b基因表达的物质、smdda1b基因过表达基因重组构建体中的一种或两种。
20.从激素处理实验,vigs,过表达的激素信号途径基因表达以及过表达sa含量测定结果可得,smdda1b在抗茄子青枯病的防御反应中受到sa途径及ja途径的正调控,同时也可靶向sa及ja的信号途径基因,因此,优选的,smdda1b基因的表达受sa途径正调控。
21.本发明还提供一种提高茄子青枯病抗性的方法,包括以下步骤:
22.s1、构建以smdda1b基因作为靶基因的过表达载体;
23.s2、将构建的过表达载体转化到茄子中;
24.s3、筛选获得阳性转基因茄子。
25.与现有技术相比,本发明具有以下有益效果:
26.本发明利用茄子负调控转录因子smnac为诱饵,从cdna文库中筛选出e3泛素连接酶基因smdda1b并进行功能验证,结果表明茄子e3连接酶基因smdda1b正调控茄子抗青枯病。本研究以期为茄子抗青枯病的分子机制提供理论参考。
附图说明
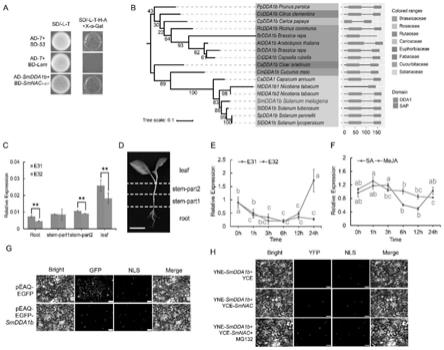
27.图1为茄子e3泛素连接酶基因smdda1b的获得,系统发育分析结果,组织特异性分析,亚细胞定位以及青枯菌接菌和激素处理结果;(a)smnac和smdda1b的y2h结果,bd
‑
53与ad
‑
t共转y2h
‑
gold是阳性对照,bd
‑
lam和ad
‑
t共转y2h
‑
gold是阴性对照;(b)e3泛素连接酶基因smdda1b的系统发育分析结果;系统发育树分析smdda1b及smdda1b在其他不同科中的同源蛋白序列,用不同颜色标出,蛋白序列由ncbi获得,分支上的数字代表支持度,数值越大可信度越高,数值最高为100;(c)smdda1b在茄子“e31”、“e32”的根、茎部分1、茎部分2、叶中的相对表达情况;误差线表示3个生物学重复的标准差,“*”表示两数据的p值小于0.05,且大于0.01,具显著性差异,“**”表示两数据的p值小于0.01,具极显著性差异;(d)为茄子苗的组织部分示意图,分别为叶、茎的上部分、茎的下部分和根的示意图,图中白色标尺表示5cm长度;(e)e31、e32接种青枯菌后24h内的smdda1b相对表达情况分析,取样时间为接种青枯菌后0h、1h、3h、6h、12h、24h,误差线表示3个生物学重复的标准差,各数据间的多重比较结果采用字母标记法表示(p<0.05),最终结果表示为处理组基因相对于对照组基因的相对表达量;(f)e31被sa及meja处理后24h内的smdda1b的相对表达情况,取样时间分别为激素处理后的0h、1h、3h、6h、12h、24h,误差线表示3个生物学重复的标准差,各数据间的多重比较结果采用字母标记法表示(p<0.05),最终结果表示为处理组基因相对于对照组基因的相对表达量;(g)smdda1b的亚细胞定位结果。亚细胞定位的材料为本氏烟草,bright表示荧光显微镜的烟草明场,gfp表示细胞核或细胞膜等发出的绿色荧光,该基因将在此处进行亚细胞水平的表达,nls表示细胞核发出的红色荧光,作为参照,merge表示将前三者组合;
bright和merge的红色标尺及gfp和nls图中的白色标尺均表示1mm的长度;(h)smdda1b和smnac的bifc结果表明,材料为烟草;蛋白酶体抑制剂mg132在yne
‑
smdda1b和yce
‑
smnac农杆菌共同注射后注射;在yfp,nls,bright及merge图中的红色标尺和白色标尺都表示1mm的长度;
28.图2为smdda1b基因序列和氨基酸序列;(a)smdda1b的基因序列,smdda1b总长度为504bp,包含dda1域和sap域,分别用红色和蓝绿色标记;(b)smdda1b的氨基酸序列,全长76个氨基酸,结构域包括dda1和sap,分别用红色和蓝绿色标记;
29.图3为e31和e32对青枯病的抗病性检测;(a)e32接种青枯菌后第0天和第10天的表型;(b)e31接种青枯菌第0天和第10天e31的表型;利用青枯菌gmi1000分别接种81株e31和73株e32;结果表明,接种10天后,e32表现出明显的萎蔫表型,e31无明显变化;发病率和疾病指数统计表明,e32的发病率为89.04%,疾病指数为47.6,属于易感材料;e31的发病率为11.11%,病害指数为2.78,属于高抗材料;图中穴盘的大小为5
×
10,共50个孔;
30.图4为e31泛素连接酶基因smdda1b的vigs实验结果;(a)在对e31的smdda1b基因沉默后,分析了对照植株(仅注射ptrv2和ptrv1农杆菌液的植株)和处理植株(注射ptrv2
‑
smdda1b和ptrv1农杆菌液的植株)中smdda1b基因的相对表达;误差条代表三个以上生物复制品的标准差。低于0.01的p值被认为具有非常显著的差异,用**表示;(b)接种青枯菌4周后,对照株和处理株经e31处理后的病情指数进行统计;病害等级分为0级、1级、2级、3级、4级,0级为植株不感病,1级为植株1
‑
2叶萎蔫,2级为植株3
‑
4叶萎蔫,3级为除顶叶外所有叶片萎蔫,4级为植株死亡,纵坐标表示每种疾病级别的植物数量占总数的百分比,共有10株e31在本实验中被沉默。低于0.01的p值被认为具有非常显著的差异,用**表示,(c)接种青枯菌4周后,对照植株和处理植株的e31
‑
vigs表型发生变化,图中的白色标尺表示长度为5厘米;
31.图5为在番茄中过表达smdda1b实验结果;(a)在dna水平上,组织培养所得番茄单株株系的bar基因验证;(b)在rna水平上,组织培养所得番茄单株株系的smdda1b基因相对表达量分析。p<0.05,用*表示,p<0.01,用**表示;(c)野生型番茄苗(wt)及转基因番茄苗(oet1)接种青枯菌后第7天表型,包括正面图,近图及俯视图;(d)野生型番茄苗及转基因番茄苗接种青枯菌后第14天表型,ck表示相同条件下同时培养的未接种青枯菌的野生型及转基因番茄苗;(e)野生型番茄苗及过表达番茄苗接种14天的发病率统计图,其中oet1‑
12
,oet1‑
17
及oet1‑
31
‑2分别为过表达单株株系,oet1表示3个过表达株系的平均发病率;(f)野生型番茄苗及过表达番茄苗接种14天的病情指数统计图,其中oet1‑
12
,oet1‑
17
及oet1‑
31
‑2分别为过表达单株株系,oet1表示3个过表达株系的平均病情指数;(g)接种及未接种青枯菌的野生型番茄苗及过表达smdda1b番茄苗的sa含量测定结果;显著性差异用字母法标注;
32.图6为vigs及过表达中sa途径信号基因的表达情况分析;(a)vigs植株中sa途径信号基因表达情况分析结果,其中ptrv2代表对照植株,p
‑
smdda1b表示vigs处理植株ptrv2
‑
smdda1b;(b)过表达植株中sa途径信号基因表达情况分析结果,oet0代表t0代番茄过表达植株,wt代表野生型植株;p
‑
value的值高于0.01,低于0.05被认为具有显著性差异,用*表示,p
‑
value值低于0.01被认为具有极显著差异,用**表示。不同颜色的点代表不同的生物学重复,同一颜色的点代表技术重复。
具体实施方式
33.下面对本发明的具体实施方式作进一步说明。在此需要说明的是,对于这些实施方式的说明用于帮助理解本发明,但并不构成对本发明的限定。此外,下面所描述的本发明各个实施方式中所涉及的技术特征只要彼此之间未构成冲突就可以相互组合。
34.下述实施例以及实验例中所使用的试验方法如无特殊说明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂和材料;所用的设备,如无特殊说明,均为常规实验设备。
35.植物材料:茄子材料使用了华南农业大学(广东广州)曹必好教授的茄子高抗青枯病自交系“e31”(r)和茄子高感青枯病自交系“e32”(s)(图2,表4)。番茄材料使用华南农业大学(广东广州)邱正坤老师提供的money maker品种,该品种为不抗青枯病材料。烟草为本氏烟草,由本课题组提供。茄子cdna文库由本课题组提供。
36.实施例1smdda1b基因的克隆
37.rna提取,反转录cdna及rt
‑
qpcr数据分析参考qiu等(2019),相对表达量采用及计算方法(引物参考表1)。从系统发育树上均匀地选择了另外14个已测序全基因组的双子叶植物(表2),从ncbi下载每个物种的所有蛋白序列,对属于同一个基因不同可变剪接体的,只保留最长的。从pfam数据库下载dda1的隐马尔科夫模型文件,使用hmmsearch v3.3提取每个物种包含dda1的蛋白序列。把得到的蛋白序列与茄子的两个包含dda1的蛋白序列使用mafft v7.455进行多序列比对,使用iqtree v1.6.12构建系统发育树。hmmsearch使用了
“‑‑
cut_nc”对结果进行了限制,iqtree使用了
“‑
bb”参数来计算bootstrap值,除此之外,均使用了默认参数。使用itol绘制系统发育树。
38.对于所有包含dda1的蛋白序列,与pfam
‑
a数据库进行比对,扫描这些序列拥有的结构域,并使用ggplot2进行绘制。把系统发育树和结构域的图联合起来,可以发现早在双子叶植物出现以前,dda1就分成了两个不同的基因,一个只包含dda1,另外一个,也就是smdda1b所在的分支,包含dda1和sap两种结构域。本研究主要关注后者,于是把该分支提取出来,得到文章中的系统发育树。
39.表1 qrt
‑
pcr分析所用引物
40.[0041][0042]
表2 14种双子叶植物的smdda1b同源基因登录号
[0043][0044]
结果:本课题组先前研究获得茄子抗青枯病的负调控转录因子smnac,将其n端包含其nam结构域的139个氨基酸构建于pgbkt7载体,作为诱饵蛋白,对茄子cdna文库进行筛选,获得e3泛素连接酶基因smdda1b(图1a)。smdda1b的orf为504bp,167个氨基酸,18.27kda,具dda1结构域,属于crl的ddb型e3泛素连接酶的主要组成部分。拟南芥中,dda1基因(q9ffs4)已被研究,该基因与cul4、ddb1、cop10及det1形成蛋白复合体,将e2上的ub结合至脱落酸(aba)受体蛋白pyl8,完成泛素化过程。此外,dda1是crl4核心复合体进化过程中保守的基本成分,它可以直接与ddb1相互作用,促进底物补充或调节crl4底物复合体的整体拓扑结构。同时,smdda1b还具有sap结构域,也较为保守,sap被认为与连接酶的底物识别和活性有关,并常见于许多sumo e3。对smdda1b进行系统发育分析(图1 b),结果显示,15个双子叶植物中,smdda1b的同源蛋白均含有dda1及sap结构域,表明该结构域在双子叶植物中较为保守,可能在植物的生存进化中发挥较为重要的作用。值得一提的是,包含dda1结构域的基因只有两种,一种只包含dda1结构域,另一种包含dda1结构域及sap结构域(图2)。
[0045]
实施例2 smdda1b功能探索
[0046]
一、组织特异性分析
[0047]
分别取生长4
‑
5片真叶的“e31”及“e32”的根、茎、叶做组织特异性分析,各组织部位样品均含3个生物学重复,后进行rna提取,反转录cdna并进行rt
‑
qpcr,相对表达量采用计算方法。
[0048]
二、亚细胞定位
[0049]
利用一步克隆的方法将具有同源臂的目的基因片段(5
‘
端引物:ctgcccaaattcgcgaccggtatggaggatacctcatcatccatt;3’端引物:gcccttgctcaccataccggttgtgtccccccttaaccgtg)连接至age i单酶切的peaq
‑
egfp载体上,用大肠(dh5α)热激法进行筛选,并转入gv3101(psoup)农杆菌菌株中。将构好的农杆菌与核定位nls(sun et al.,2020)农杆菌划线,挑单菌落28℃,200rpm初摇,后阔摇,将农杆菌菌液6000rpm离心后,去上清,并用侵染液(10mm mgcl2,10mm mes,100μm as)重悬菌体,28℃静置活化2h后,将含目的基因农杆菌的侵染液与含核定位农杆菌的侵染液按照体积比1:1混合后,用去头的注射器将侵染液注射进烟草中,使烟草叶片呈水渍状,后放入22℃黑暗培养2
‑
3天。使用荧光显微镜可视化gfp荧光。该实验至少重复三次。
[0050]
三、双分子荧光互补实验(bifc)
[0051]
利用一步克隆法将目的基因与待验证基因分别构建于pspyne
‑
35s/puc
‑
spyne(yne)及pspyce
‑
35s/puc
‑
spyce(yce)载体(5’端限制性内切酶为bamh i,3
‘
端为sal i)上形成重组载体,并转入gv3101(psoup)农杆菌菌株中。将构好的农杆菌与核定位nls农杆菌划线,挑单菌落28℃,200rpm初摇,后阔摇,将农杆菌菌液6000rpm离心后,去上清,并用侵染液(10mm mgcl2,10mm mes,100μm as)重悬菌体,od600调至0.6,28℃静置活化2h后,将含yne
‑
目的基因农杆菌的侵染液、yce
‑
目标基因农杆菌侵染液与含核定位农杆菌的侵染液按照体积:1:1:1混合后,用去头的注射器将侵染液注射进烟草中,使烟草叶片呈水渍状,后放入22℃黑暗培养3
‑
4天。使用荧光显微镜可视化gfp荧光。该实验至少重复三次。
[0052]
表3 bifc实验引物汇总表
[0053][0054][0055]
结果:从“e31”中克隆得到smdda1b基因全长,并验证其是否具有茄子抗青枯病的功能。以茄子抗病材料“e31”及感病材料“e32”(图3)的根茎叶为材料,进行组织特异性分析。结果表明,smdda1b基因在抗病材料和感病材料的根、茎、叶中均有表达,且在叶中表达量最大,抗病材料中smdda1b基因的表达量普遍高于感病材料(图1c
‑
d)。之后,对其进行亚
细胞定位以确定其在亚细胞水平的表达区域,结果表明,peaq
‑
egfp
‑
smdda1b只在核中发荧光,smdda1b在核中表达(图1g),且bifc结果表明,smnac可与smdda1b在植物细胞核内进行互作。
[0056]
四、青枯菌接种
[0057]
青枯菌菌株为gmi1000,由华南农业大学(广东广州)提供。接种方法采用常规的“断根灌根法”,病情指数分5个等级(0级为植株不发病、1级为1
‑
2片叶萎蔫、2级为3
‑
4片叶萎蔫、3级为除顶端叶片,其余叶片均萎蔫、4级为植株死亡)。e31、e32断根后,对对照组进行清水处理,处理组接种青枯菌处理。后于早上8:00开始取样,分别取0h、1h、3h、6h、12h、24h的样,每个样含3个生物学重复,后提取rna,反转成cdna后进行rt
‑
qpcr,最终结果表示为处理组基因相对于对照组基因的相对表达量。各时间点数据的多重比较结果用字母标记法表示。
[0058]
五、激素处理
[0059]
对4
‑
5片真叶的茄子苗喷施1mm sa(jia et al.,2013;mahesh et al.,2018)及0.25mm meja(deng et al.,2021),对照组喷施清水,26℃,16h光照,22℃,8小时黑暗培养。从早上8:00开始取样,分别取0h、1h、3h、6h、12h、24h的样,每个时间点至少3个生物学重复,将样进行rna提取,并反转录cdna,做qpcr。引物参考表1,各时间点数据的多重比较结果用字母标记法表示。
[0060]
结果:我们利用sa及meja对e31进行处理,qpcr测定smdda1b基因在激素处理后一天内的相对表达量变化情况,如图1f所示,e31被sa处理后,smdda1b表现出先下降,再上升的趋势;表明短期内喷施sa,基因表达受其影响,且最终呈上升趋势,sa对基因表达具有一定促进作用;而对e31喷施meja后,smdda1b表达量持续下降,表明短期内喷施meja可抑制smdda1b表达。
[0061]
再将青枯菌分别接种至茄子抗病材料e31与感病材料e32中,qpcr测定基因相对表达量变化,分析青枯菌侵染不同材料是否会调动smdda1b基因的表达,结果显示,smdda1b在抗病植株中表达量呈先下降后上升的趋势,而在感病植株中呈先下降后趋于平缓的趋势,表明青枯菌侵染抗病植株可提高e31中smdda1b基因的表达(图1e)。
[0062]
六、vigs实验
[0063]
将目的基因截取300bp的特异片段,运用双酶切(5
‘
端酶切位点为ecor i,引物:ggaattccctccgaacaatgccaca;3’端酶切位点为sma i,引物:tcccccggggaaatccccttgccgtct)的方法将其构建在ptrv2载体上,后转入农杆菌gv3101菌株。以清水处理4
‑
5片真叶的e31植株及按体积比1:1注射ptrv2,ptrv1空载农杆菌菌液植株为对照,以ptrv2
‑
smdda1b与ptrv1空载农杆菌菌液以体积比1:1注射e31植株为处理组,注射后黑暗条件下16℃处理一天,后正常培养1
‑
2周,(26℃,16h光照,22℃,8h黑暗),每个处理至少10个生物学重复,后取样,进行rna及qpcr。之后采用“断根灌根法”进行青枯菌(gmi1000)接菌(od
600
=0.6),接种一个月左右观察表型。
[0064]
结果:为进一步验证e3泛素连接酶基因smdda1b是否抗茄子青枯病,对其进行基因沉默(vigs)实验并统计接种青枯菌后的发病指数,liu等(2005)制定了茄子对青枯病的抗性评价标准(表5)。结果显示,将高抗材料e31中smdda1b的基因表达量降低后接种青枯菌(图4a),e31表现出明显的感病症状,且发病率达到100%,发病指数为70,属于茄子高感青
枯病水平(图4b
‑
c)。初步表明smdda1b与茄子抗青枯病相关,且可能为茄子抗青枯病网络的关键位点(图4b)。
[0065]
七、过表达实验
[0066]
利用5
‘
端上游引物gagaacacgggggactctagaatggaggatacctcatcatccattc及3
‘
端下游引物gtggctagcgttaacactagttcatgtgtccccccttaaccg进行过表达目的基因smdda1b片段扩增,后利用限制性核酸内切酶xba i及spe i 37℃,1h双酶切过表达载体pcambia
‑
1380,并利用翊圣一步克隆试剂盒进行载体重组,利用pcambia
‑
1380的通用引物(5
‘
上游引物ggctcctacaaatgccatcattgcg;3’下游引物ataatttatcctagtttgcgcgc)检测,获得重组过表达载体,后转入gv3101农杆菌菌株。番茄过表达植株的获取采用农杆菌介导转化法。先在含有ms固体培养基(ms粉4.43g/l,蔗糖30g/l,0.5%植物凝胶)的组培瓶中播种mm番茄种子,待长出两片子叶后,切取子叶中间段及下胚轴作为外植体放在固体预培养基(ms固体培养基,0.2μg/l反式玉米素tzt)上,26℃黑暗培养1天。用6000rpm,5min离心含有重组载体的农杆菌,去上清,用ms培养基重悬菌体,调至od600为0.6,并加入100μm的as,28℃黑暗静置1h。将预培养1天的外植体放入侵染液中,黑暗晃动侵染10分钟,后滤去侵染液,并用滤纸吸干外植体水分,将外植体放在固体预培养基上,28℃黑暗培养1天后放入固体筛选培养基(ms固体培养基,2μg/l tzt,0.2mg/l tim,0.5μg/l iaa,相应抗生素)进行正常培养(26℃,光照16h,22℃,黑暗8h),并每两周换一次,选择生长健壮的外植体继续进行实验,待外植体长出愈伤组织,分化出芽时,转入芽伸长(ms固体培养基,0.2μg/l tzt,0.2mg/l tim,相应抗生素)培养基中继续使芽伸长,待茎长约3
‑
4cm时,切下芽并将其插入生根培养基(ms固体培养基,0.2mg/l tim,0.5μg/l iaa,相应抗生素)中进行生根,待植株生根并生长至瓶盖时,开盖进行练苗,一周后转移至灭菌的土中,套透明塑料袋处理一周,取样,进行植株bar基因(5
‘
端上游引物atgagcccagaacgacgcccg,3’端下游引物ttagatctcggtgacgggcaggacc)dna检测,smdda1b基因相对表达量检测,完成过表达实验。
[0067]
八、酵母双杂交(y2h)
[0068]
利用一步克隆的方法将具有同源臂的smnac n端417bp片段连接至ecor i(5
‘
端)和bamh i(3’端)双酶切的pgbkt7载体上,具体引物为5’端上游引物atggccatggaggccgaattcatgggtgttcaagaaaaagatcct,3’端下游引物ccgctgcaggtcgacggatcctaatctatattcatgcatgatccaattag。制作酵母y2h gold感受态,将y2h gold菌株在ypda平板上划线,30℃培养2
‑
3天,挑取直径2
‑
3mm的酵母单菌落,接种于3
‑
4ml ypda培养基,30℃,220rpm,培养12
‑
18h,至摇浓为止,后加入50mlypda,继续阔摇3
‑
5h,摇浓后,3000rpm,离心5min,去上清,并用30ml ddh2o重悬清洗两次,离心去上清后,用现配的1ml 1
×
te/liac重悬,即得感受态。
[0069]
取100μl y2h gold感受态于1.5ml离心管中,加入0.5μg pgbkt7
‑
bait及cdna文库质粒(0.5μg),5μl carry dna,600μl无菌的peg/liac溶液,混匀,30℃,200rpm摇动培养20min,后加入70μl dmso混匀,42℃水浴15min,冰浴2min。室温14000rpm,离心15s,弃上清,用100μl 1
×
te缓冲液重悬细胞,取100μl涂布于相应缺陷培养基上,30℃倒置培养3
‑
4天。
[0070]
结果:以番茄money marker为材料,对smdda1b进行超表达实验,以获得能够稳定遗传基因的植株,从正面验证smdda1b的功能。通过对组织培养获得的番茄苗进行bar基因检测,从60株组培苗中获得的12棵t0代转基因植株(图5a),并利用rt
‑
qpcr技术进行smdda1b基因的相对表达量检测,选取过表达效果较好的3个单株进行t1代繁殖,分别为
t0‑
12
,t0‑
17
及t0‑
31
‑2(图5b)。之后对所得t1代株系及野生型番茄苗进行青枯菌接种,结果表明野生型番茄苗与过表达番茄苗的发病时间相同,都为接种青枯菌后第五天发病,但接种青枯菌的14天内,野生型植株的发病率及病情指数都明显高于过表达植株(图5e
‑
f,表6)。观察接种青枯菌后第7天及第14天的wt及过表达植株,野生型番茄苗基本萎蔫,过表达番茄苗呈现部分萎蔫,比野生型植株表型出更抗青枯病表型(图5c
‑
d,表7)。结果表明,过表达smdda1b可提高植株对青枯菌抗性,smdda1b具有抗青枯病的功能。后对接种及未接种青枯菌的野生型番茄苗及过表达smdda1b番茄苗进行sa含量测定,结果显示,未接种青枯菌的野生型番茄苗sa含量显著低于过表达番茄苗,接种青枯菌后,wt及过表达番茄苗的sa均显著上升,但野生型番茄苗sa含量仍显著低于过表达番茄苗,表明,植株受到生物胁迫时可诱导sa含量上升,同时,smdda1b与sa途径具相关性,过表达smdda1b可诱导sa含量升高,植株表现出更强的抗病性。
[0071]
同时,对smdda1b过表达植株及vigs植株进行激素信号途径基因表达量分析。sa途径信号基因中筛选eds1、glua、npr1、tga、sgt1、pad4、pr
‑
1a及ics1,8个正调控茄子青枯菌抗性的基因。发现与对照相比,vigs中sa途径信号基因除tga外,其余基因均表现出显著性差异,且沉默植株中sa途径信号基因表达量下降(图6a)。在过表达植株中,与野生型相比,sa途径信号基因中除pr
‑
1a外,其余基因表达量具显著性差异,且呈上升趋势。该结果初步表明sa途径正调控smdda1b的表达(图6b)。
[0072]
讨论:在本研究中,本课题组以茄子抗青枯病的负调控转录因子smnac n端包含nam结构域的417bp碱基为诱饵,对茄子cdna文库进行筛选后获得e3泛素连接酶基因smdda1b,并进行功能验证。从结果看出,smdda1b在e31、e32接种青枯菌后的表达趋势不同,抗病植株中先下降后上升,在感病植株中先下降后趋于平缓的趋势。该结果表明,植株在受到病原胁迫时,其先天免疫系统会做出反应,即病原相关分子模式激发的免疫反应(pti)和效应蛋白激发的免疫反应(eti),pti是基础防卫反应,具非特异性,而eti则是由植物的抗病蛋白(r蛋白)识别病原的效应蛋白引起的特异性反应。在植株受到病原胁迫时,植株首先做出非特异性防御反应后,病原再释放效应蛋白抑制pti,这就导致e3泛素连接酶smdda1b表达在抗病植株及感病植株中都呈下降趋势,后植株进行eti反应,此时,smdda1b表达在抗病植株中上升。该结果说明,e3泛素连接酶smdda1b很有可能是正调控茄子抗青枯病的基因。同时,也侧面表明,e3泛素连接酶的基因具特异性。
[0073]
青枯菌含多种分泌系统,但主要通过iii型分泌系统发挥作用。t3ss可以将自身的效应因子注射到寄主体内引起寄主感病或超敏反应(hr)。同时,有研究表明,植物体内的ups可特异性识别病原效应子,在植物
‑
病原体互作中发挥作用。gabri
è
le等研究表明,在体外,ups可靶向turnip yellow mosaic virus的运动蛋白69k,调节其活性。在烟草中,ring型e3泛素连接酶ntrfp1可介导双子病毒编码的βc1降解。在此过程中,依据smdda1b对青枯菌防御反应的特异性,其可能与青枯病的毒性基因,即效应蛋白发生互作反应,将其泛素化后降解,以此进行植株防御反应。
[0074]
此外,从激素处理实验,vigs,过表达的激素信号途径基因表达以及过表达sa含量测定结果可得,smdda1b在抗茄子青枯病的防御反应中受到sa途径及ja途径的正调控,同时也可靶向sa及ja的信号途径基因。例如,先前有研究表明,e3连接酶cul3
bpm
可靶向myc2、myc3及myc4,降低myc蛋白丰度,调控ja通路等。
[0075]
vigs等实验初步筛选出结果最明显,且之前未被进行功能验证的e3泛素连接酶基因smdda1b,之后对其进行过表达实验,再次验证了其确实对植物抗青枯病具有正调控作用,且推测为茄子抗青枯病的重要基因。
[0076]
表4 e31及e32对茄子抗青枯病的病情指数统计表
[0077][0078]
表5茄子抗青枯病的病情指数评价标准表
[0079][0080]
表6野生型番茄苗与过表达番茄苗接种青枯菌14天内的发病率及病情指数统计表
[0081][0082]
表7野生型与过表达番茄接种青枯菌后14天发病率及病情指数统计表
[0083]
[0084][0085]
以上对本发明的实施方式作了详细说明,但本发明不限于所描述的实施方式。对于本领域的技术人员而言,在不脱离本发明原理和精神的情况下,对这些实施方式进行多种变化、修改、替换和变型,仍落入本发明的保护范围内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。