1.本发明属于基因工程技术领域,特别涉及一种半矮化香蕉基因编辑载体及其构建方法和应用。
背景技术:
2.株高是农作物的重要性状之一,合适的株高是保障产量稳定的一个重要因素。香蕉作为重要的经济作物,也是仅位于水稻(oryza sativa)、小麦(triticum aestivum)、玉米(zea mays)之后的第四大粮食作物,树体一般超过2米,极易受到高速风,低/高气旋台风的影响,这些台风吹倒了假茎,给商业种植园带来了不可挽回的经济损失。在集约化农业种植条件下,植株矮化可以通过增加抗倒伏能力和降低风雨造成的损害机会对作物农艺产生积极的影响,且矮化的香蕉品种适合密集的田间栽培,促进机械化维护,提高果实收集的效率,并通过减少喷雾漂移允许更精确的农药施用。
3.作为重要的粮食和经济作物,商业种植的香蕉多为三倍体(aaa)基因组类型,当今种植的cavendish香蕉实际上是其祖先的遗传复制品,在180多年前被首次繁殖,并且大面积种植的品种,如
‘
grand nain’(musa acuminata,aaa group,cavendish subgroup,大矮蕉),可能是因其理想的农艺性状而被选择的天然克隆变种。然而,三倍体香蕉基本上是不育的,为cavendish引入新的和有用的特性进行繁殖几乎是不可能的。
4.申请人曾将株高超过3米的香蕉优良品种
‘
gros michel’(m.acuminata,aaa group,又名大蜜舍、大蜜哈等),通过crispr/cas9基因编辑技术,对水稻ga20ox2基因在香蕉中的5个同源基因进行定点编辑,获得了具有半矮化表型的
‘
gros michel’香蕉突变株系(shao et al.,2019)。但该品种不抗香蕉枯萎病菌1号生理小种,即使获得了植株高度极显著下降的新种质,也只能在温室或资源圃中保存,无法进行商业化种植。因此,有必要提供一种植株矮化或半矮化的、且适合商业化种植的香蕉。
技术实现要素:
5.基于此,本发明的目的在于提供一种基于crispr/cas9的香蕉ga20ox2基因编辑载体及其构建方法和应用。
6.为实现上述目的,本发明具体技术方案如下:
7.一种半矮化香蕉基因编辑载体的构建方法,包括以下步骤:
8.s1.将核苷酸序列如seq id no.2、seq id no.3所示的接头引物与pylgrna-osu6a质粒混合,同时进行酶切和连接反应,得连接产物;
9.s2.由核苷酸序列如seq id no.4和seq id no.3所示的引物,以s1中所述的连接产物为模板进行pcr扩增反应,得产物a;
10.由核苷酸序列如seq id no.2和seq id no.5所示的引物,以s1中所述的连接产物为模板进行pcr扩增反应,得产物b;
11.s3.将所述产物a与所述产物b混合作为模板,以seq id no.6和seq id no.7为引
物,制备得到grna表达盒;
12.s4.将所述sgrna表达盒连接至pylcrispr/cas9pubi-h载体上。
13.在其中一些实施例中,所述grna表达盒中含有核苷酸序列如seq id no.1所示的靶位点。
14.在其中一些实施例中,步骤s1中所述的同时进行酶切和连接反应,反应体系中还包含有bsa i内切酶、atp和t4 dna连接酶;及/或
15.所述pylcrispr/cas9pubi-h载体中融入了gfp报告基因。
16.本发明的目的还在于提供一种半矮化香蕉基因编辑载体,具体技术方案如下:
17.一种半矮化香蕉基因编辑载体,由如上所述的半矮化香蕉基因编辑载体的构建方法制备得到。
18.本发明的另一个目的还在于提供一种工程菌,具体技术方案如下:
19.一种工程菌,所述工程菌为含有如上所述的半矮化香蕉基因编辑载体的细菌。
20.在其中一些实施例中,所述工程菌为含有上述的半矮化香蕉基因编辑载体的农杆菌eha105。
21.本发明还提供了一种半矮化香蕉基因定点突变试剂盒,具体技术方案如下:
22.一种基因定点突变试剂盒,包含如上所述的半矮化香蕉基因编辑载体或如上任一项所述的工程菌或核苷酸序列如seq id no.2-3所示的引物。
23.本发明还提供了一种核苷酸序列如seq id no.16所示的基因ma04g15900和核苷酸序列如seq id no.17所示的基因ma08g32850在半矮化香蕉育种中的应用。
24.在其中一些实施例中,所述应用为:使所述核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19、20、21、22、23和24中的至少一种片段。
25.在其中一些实施例中,所述应用为:使所述核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、27、28、29、30和31中的至少一种片段。
26.本发明还提供了一种半矮化香蕉的育种方法,具体技术方案如下:
27.一种半矮化香蕉的育种方法,包括以下步骤:使用如上所述的工程菌与三倍体香蕉品种
‘
grand nain’的胚性悬浮细胞系或二倍体aa基因型的贡蕉细胞系共培养,经液体培养基筛选、半固体培养基筛选诱导产生胚。
28.本发明还提供了一种半矮化香蕉,具体技术方案如下:
29.一种半矮化香蕉,由如上所述的半矮化香蕉的育种方法制备得到。
30.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19、20、21、22、23和24中的至少一种片段;及/或
31.所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、27、28、29、30和31中的至少一种片段。
32.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、21和23中的
至少一种;和/或所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、28、29、30和31中的至少一种。
33.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19和24中的至少一种;和/或所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、28和30中的至少一种。
34.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19中的至少一种;和/或所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、28中的至少一种。
35.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19、20中的至少一种;和/或所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、28中的至少一种。
36.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18、19中的至少一种;和/或所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26、27、28中的至少一种。
37.在其中一些实施例中,所述半矮化香蕉的核苷酸序列如seq id no.16所示的基因ma04g15900中的核苷酸序列如seq id no.1所示的片段突变为seq id no.18和19所示的片段;所述半矮化香蕉的核苷酸序列如seq id no.17所示的基因ma08g32850中的核苷酸序列如seq id no.25所示的片段突变为seq id no.26和28所示的片段。
38.基于上述技术方案,本发明具有以下有益效果:
39.本发明针对香蕉基因的序列,设计得到基因编辑靶位点、接头引物和pcr扩增引物,并在构建半矮化基因编辑载体时结合特定的酶切连接反应和扩增反应,构建得到合理的grna表达盒,提供了一种针对特定香蕉的特定基因的有效且稳定的定向突变方法,从而使得香蕉获得半矮化性能,且突变体在植株变矮的同时,其叶片细胞形态受到的影响较小。提高了香蕉的抗倒伏能力,减轻台风等恶劣天气对香蕉种植园的影响。
40.进一步地,通过构建上述基因编辑载体及含有该载体的工程菌,应用于三倍体香蕉品种
‘
grand nain’的胚性悬浮细胞系的基因定点突变,有望得到具有稳定遗传的半矮化香蕉突变体,适合商业化种植。
附图说明
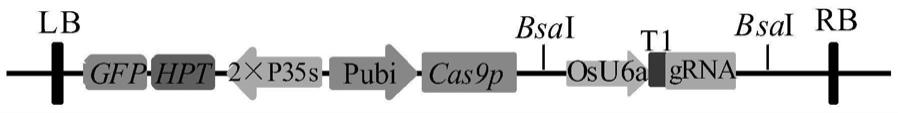
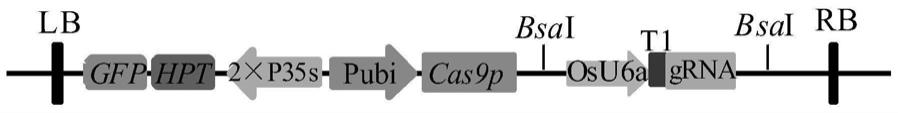
41.图1为pcrispr/cas9-osu6a-t1-grna基因编辑载体示意图;
42.图2为农杆菌介导的
‘
grand nain’香蕉遗传转化及植株再生;
43.图3为潮霉素基因hpt的pcr检测;
44.图4为潮霉素基因hpt的southern blot检测;
45.图5为
‘
grand nain’突变体的表型及植株不同生长期株高的变化;
46.图6为
‘
grand nain’叶片中内源gas含量的测定;
47.图7为
‘
grand nain’叶片细胞结构的分析。
具体实施方式
48.为了便于理解本发明,下面将参照实施例对本发明进行更全面的描述,以下给出了本发明的较佳实施例。但是,本发明可以以许多不同的形式来实现,并不限于本文所描述的实施例。提供这些实施例的目的是使对本发明的公开内容的理解更加透彻全面。应理解,下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如sambrook等人,分子克隆:实验室手册(new york:cold spring harbor laboratory press,1989)中所述的条件,或按照制造厂商所建议的条件。实施例中所用到的各种常用试剂,均为市售产品。
49.除非另有定义,本文所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。在本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不是旨在于限制本发明。本文所使用的术语“和/或”包括一个或多个相关的所列项目的任意的和所有的组合。
50.实施例1香蕉ga20ox2基因编辑载体的构建
51.1、根据香蕉的基因序列,分别以ma04g15900和ma08g32850基因的第二个外显子内的5'-ccg-gagctgacgctcggcaccgg-3'(seq id no.1)序列为靶标位点。
52.2、靶位点接头引物的设计及制备
53.根据靶位点的序列,采用水稻的u6a启动子来驱动sgrna,根据靶位点接头引物的设计原则,本试验中靶位点的接头引物序列为:
54.f:5'-gccgccggtgccgagcgtcagctc-3'(seq id no.2)
55.r:5'-aaacgagctgacgctcggcaccgg-3'(seq id no.3)
56.将接头引物用te缓冲液溶解成浓度为100μmol/l的母液,接着各取1μl加入到98μl的0.5
×
te缓冲液中,混合稀释到1μmol/l。采取约90℃30s的处理,然后转移至室温进行冷却,完成退火。
57.3、边切边连制备grna表达盒
58.采用边切边连的方式制备grna表达盒。配制10
×
bsa i酶切连接反应液,加入atp、pylgrna-osu6a质粒(提前配制成20ng/μl进行保存)、上述序列为seq id no.2和seq id no.3的接头引物(最终浓度0.05μmol/l)、bsa i内切酶,neb t4 dna ligase等,反应体系为10
×
bsa i内切酶buffer 1μl,atp(20mmol/l)0.5μl,质粒pylgrna-osu6a(20ng/μl)1μl,接头引物(1μmol/l)0.5μl,bsa i 5u,neb t4 dna ligase 35u,ddh2o补充至10μl。以37℃5min,20℃5min,反应5个循环。
59.其中,pylgrna-osu6a质粒由华南农业大学刘耀光院士课题组惠赠。
60.4、扩增grna表达盒
61.采用2轮巢式pcr的方法扩增grna表达盒,第一轮扩增为2个反应,引物搭配如下:
62.反应1:u-f t1r,具体引物序列如下:
63.u-f:5'-ctccgttttacctgtggaatcg-3'(seq id no.4)
64.t1r:5'-aaacgagctgacgctcggcaccgg-3'(seq id no.3)
65.反应2:t1f grna-r,具体引物序列如下:
66.t1f:5'-gccgccggtgccgagcgtcagctc-3'(seq id no.2)
67.grna-r:5'-cggaggaaaattccatccac-3'(seq id no.5)
68.反应体系如下:10
×
kod buffer 1.5μl,dntp 0.6μl,primer f 1μl,primer r 1μl,mgso
4 0.6μl,kod 0.3u,步骤3的连接产物1μl,ddh2o 9μl,total volume为15μl。
69.pcr反应程序为:95℃10s,69℃15s,68℃20s,反应25-28个循环。
70.第二轮是overlapping pcr,用位置特异引物扩增出grna表达盒产物。
71.各取第一轮扩增中反应1 反应2的产物1μl稀释10倍,合为一管,取其中1μl作为二轮pcr的模板,反应体系如下:10
×
kod buffer 3μl,dntp 1.2μl,primer(pt1l)3μl,mgso
4 1.2μl,kod 0.6u,稀释10倍的一轮产物1μl,ddh2o 20μl,total volume为30μl。pcr反应程序为:95℃10s,58℃15s,68℃20s,反应15-20个循环。
72.所述的位置特异引物pt1l为引物对b1'/bl,具体序列如下:
73.uctcg-b1':ttcagaggtctctctcgactagtggaatcggcagcaaagg(seq id no.6)
74.grcggt-bl:agcgtgggtctcgaccgacgcgtccatccactccaagctc(seq id no.7)
75.5、纯化grna表达盒产物
76.将步骤4制备得到的t1-u6a的产物过柱纯化,用超微量紫外可见分光光度计检测浓度。
77.再用bsa i内切酶,以边切边连法连接到改造后的pylcrispr/cas9pubi-h载体(即在该载体中将gfp报告基因与hpt基因融合)。具体步骤如下:
78.本试验选择1个靶点,则表达盒用量约10ng,载体用量为60-80ng,采用边切边连的方式,反应体系如下:10
×
bsa i内切酶buffer 1.5μl,cas9质粒(pylcrispr/cas9pubi-h)60-80ng,t1-u6a的产物0.5μl,bsa i 10u,ddh2o up to 10μl。37℃条件下反应10min。加入0.75μl atp(20mmol/l)和35u neb t4 dna连接酶,在pcr仪上按照以下程序:37℃2min,10℃3min,20℃5min,最后37℃2min,反应10-15个循环。
79.其中,为了提高遗传转化效率,载体中融入了gfp报告基因,将增强型绿色荧光蛋白egfp基因的cds序列进行合成,通过酶切连接与载体中的hpt基因进行融合,新载体命名为pcrispr/cas9-osu6a-t1-grna,新载体示意图如图1。
80.6、转化大肠杆菌
81.取1-1.5μl连接产物通过电激法转化e.coli dh10b感受态细胞,电激电压为1500-1600v/20μl感受态细胞/1μl电激杯,电激后加入1ml soc,37℃条件下培养1-1.5h。涂板培养基为lb 25μg/ml kan,0.3~0.5mmol/l iptg,适量x-gal,阳性克隆产生蓝色菌斑,然后pcr检测阳性克隆。
82.7、重组质粒转化农杆菌
83.将鉴定无误的重组质粒用热激法转化根癌农杆菌菌株eha105感受态,步骤如下:a.吸取10μl质粒加至100μl eha105感受态细胞中,冰上放置30min;b.将离心管转入液氮中冷冻2min;c.将离心管转入37℃水浴5min;d.将离心管转入冰上放置2-3min;e.离心管中加入800μl yeb液体培养基,在28℃,220rpm条件下振荡培养3h;f.以4000rpm离心5min,去上清,加入800μl yeb液体培养基重悬;g.吸取200μl菌液进行涂板;h.将涂布的平板在28℃条件下倒置培养2-3d;i.菌落pcr阳性鉴定,待平板上长出合适大小的单菌落,随机挑取10个,进行hpt基因的pcr扩增,同时将检测为阳性的菌落,进行扩繁保种,并保存在-80℃冰箱备
用。
84.其中,pcr反应体系如下:单菌落1个,hpt-f 1μl,hpt-r 1μl,pcr mix 10μl,ddh2o8μl,total volume为20μl。pcr反应程序为:98℃预变性3min,98℃变性30s、56℃退火30s、72℃延伸1min,反应30个循环,最后72℃延伸5min。
85.其中引物序列如下:
86.hpt-f:5'-atgaaaaagcctgaactcaccgc-3'(seq id no.8)
87.hpt-r:5'-tccatcacagtttgccagtgata-3'(seq id no.9)
88.实施例2:
‘
grand nain’转基因株系的获得
89.本试验以三倍体香蕉品种
‘
grand nain’的胚性悬浮细胞系为材料,进行遗传转化,将含有pcrispr/cas9-osu6a-t1-grna载体的eha105农杆菌从-80℃冰箱取出,先进行活化,待达到合适od
600
值后,接着进行侵染悬浮细胞系、共培养、液体培养基筛选、半固体培养基筛选诱导产生胚及胚成熟、成熟胚萌发、萌发芽生根等过程(邵秀红等,“gros michel”香蕉胚性细胞悬浮系及遗传转化体系的建立.分子植物育种,2019,17(3):846
–
854)。待植株长至6cm左右高时,经过炼苗,移栽至营养杯中,在玻璃温室中培养。
90.其中,所述液体培养基筛选为将胚性细胞团在含有潮霉素b的液体培养基中筛选。筛选过程中,取适量细胞团在荧光显微镜下观察,成功转化的细胞团含有gfp绿色荧光(图2b)。三次液体筛选后,将细胞团转移至半固体筛选培养基中(图2a),1-2个月内发育成白色抗性体细胞胚(图2c),抗性体胚在半固体筛选培养基中继代培养1个月左右成熟,然后转移至萌发培养基得到抗性芽(图2d),将抗性芽转移至光下培养,接着在没有筛选压的生根培养基上培养,其中未转基因的对照1周左右生根,而转基因的抗性芽生根时间稍晚,随着时间的延长,未转基因的对照植株根部正常生长且植株较高(图2e),而转基因的株系根部稍短但植株高度与对照差异不明显(图2f)。植株在生根培养基上培养1个月后,经过炼苗,转移至温室中培养。
91.实施例3:
‘
grand nain’转基因株系的阳性鉴定
92.1、香蕉基因组dna的提取
93.选取其中9个转基因抗性株系和对照株系的嫩叶,用来提取基因组dna,根据tiangen植物基因组dna提取试剂盒的操作方法进行。提取的dna,用1%的琼脂糖凝胶电泳和nanodrop 2000c超微量紫外分光光度计检测dna的质量和浓度,并做好记录,计算总dna量。
94.2、hpt基因pcr鉴定
95.依据hpt基因的序列设计特异引物一对,并由生工生物工程(上海)股份有限公司合成,引物的序列为:seq id no.8和seq id no.9。
96.pcr扩增体系如下:dna模板1μl,hpt-f(2.5μmol/l)2μl,hpt-r(2.5μmol/l)2μl,10
×
buffer 2μl,dntp(1μmol/l)3μl,1u taq酶0.5μl,ddh2o 9.5μl,total volume 20μl。pcr反应程序为94℃预变性3min,接着94℃变性30s、56℃退火30s、72℃延伸30s,30个循环,最后72℃延伸10min。
97.结果显示,9个抗性株系均有约500bp的目标片段,而对照没有(图3),初步显示hpt基因已整合到
‘
grand nain基因组中。
98.2、southern blot检测
99.为了进一步验证转基因抗性株系是否阳性,本试验检测了hpt基因在抗性株系基因组中的拷贝数。待这9个抗性株系的叶片长至合适大小时,取其新鲜叶片速冻,然后委托钟鼎公司进行southern印迹杂交检测。结果显示,阳性质粒和9个抗性株系均有杂交条带,而对照没有条带(图4),从而进一步验证了这9个抗性株系均是阳性的。
100.实施例4:
‘
grand nain’转基因阳性株系靶位点的突变分析
101.‘
grand nain’转基因阳性株系在温室生长早期,与对照相比,株高的差异不明显。为了检验2个靶标基因是否发生了突变,分别选取在温室中生长180d后7个阳性株系的新叶和中部叶片并将其进行混合,将每个株系的混合叶分别研磨均匀后各取适量进行dna的提取,对2个靶基因分别设计引物,采用二轮pcr方法对靶位点进行扩增,其中第一轮pcr反应体系如下:kod 1μl,10
×
buffer 5μl,dntps 5μl,mgso
4 3μl,引物f1.5μl,引物r1.5μl,dna 1μl,ddh2o 32μl,total volume 50μl。
102.第一轮的引物具体名称及序列如下:
103.ma04g15900-f:cattgccgctgaagctttcttgtacacc(seq id no.10)
104.ma04g15900-r:tttccatctggtcctaccgtgatcaagc(seq id no.11)
105.ma08g32850-f:acgtcgcattctaacctcttcggtgctt(seq id no.12)
106.ma08g32850-r:aagatgacaaaccagacaaccatgctcc(seq id no.13)
107.pcr反应程序为:95℃预变性3min,95℃变性30s、58℃-68℃退火30s、68℃延伸1min 10s,反应11个循环;接着95℃变性30s、58℃退火30s、68℃延伸1min 10s,反应34个循环,然后68℃5min,12℃∞。
108.将第一轮的pcr产物稀释10倍后用作第二轮的dna(pcr模板),pcr反应体系如下:kod 1μl,10
×
buffer 5μl,dntps 5μl,mgso4 3μl,引物f1.5μl,引物r1.5μl,dna 5μl,ddh2o 28μl,total volume 50μl。
109.第二轮的引物具体名称及序列如下:
110.ma04g15900-13-f:acttgacctggcaataagcttaggagt(seq id no.14)
111.ma04g15900-13-r:tgaccacctccacctggtcctgttgaag(seq id no.15)
112.ma04g15900-15-f:acagtgcctggcaataagcttaggagt(seq id no.32)
113.ma04g15900-15-r:cagatccctccacctggtcctgttgaag(seq id no.33)
114.ma04g15900-16-f:atcacgcctggcaataagcttaggagt(seq id no.34)
115.ma04g15900-16-r:gatcagcctccacctggtcctgttgaag(seq id no.35)
116.ma04g15900-18-f:atgtcacctggcaataagcttaggagt(seq id no.36)
117.ma04g15900-18-r:gtagagcctccacctggtcctgttgaag(seq id no.37)
118.ma04g15900-21-f:actgatcctggcaataagcttaggagt(seq id no.38)
119.ma04g15900-21-r:atgagccctccacctggtcctgttgaag(seq id no.39)
120.ma04g15900-25-f:cacgatcctggcaataagcttaggagt(seq id no.40)
121.ma04g15900-25-r:cactcacctccacctggtcctgttgaag(seq id no.41)
122.ma04g15900-26-f:attcctcctggcaataagcttaggagt(seq id no.42)
123.ma04g15900-26-r:ctataccctccacctggtcctgttgaag(seq id no.43)
124.ma04g15900-wt-f:ctcagacctggcaataagcttaggagt(seq id no.44)
125.ma04g15900-wt-r:tcgaagcctccacctggtcctgttgaag(seq id no.45)
126.ma08g32850-13-f:acttgaaacccactacagagacttcttcgag(seq id no.46)
127.ma08g32850-13-r:tgaccacatctgttgccgtcgaacacc(seq id no.47)
128.ma08g32850-15-f:acagtgaacccactacagagacttcttcgag(seq id no.48)
129.ma08g32850-15-r:cagatccatctgttgccgtcgaacacc(seq id no.49)
130.ma08g32850-16-f:atcacgaacccactacagagacttcttcgag(seq id no.50)
131.ma08g32850-16-r:gatcagcatctgttgccgtcgaacacc(seq id no.51)
132.ma08g32850-18-f:atgtcaaacccactacagagacttcttcgag(seq id no.52)
133.ma08g32850-18-r:gtagagcatctgttgccgtcgaacacc(seq id no.53)
134.ma08g32850-21-f:actgataacccactacagagacttcttcgag(seq id no.54)
135.ma08g32850-21-r:atgagccatctgttgccgtcgaacacc(seq id no.55)
136.ma08g32850-25-f:cacgataacccactacagagacttcttcgag(seq id no.56)
137.ma08g32850-25-r:cactcacatctgttgccgtcgaacacc(seq id no.57)
138.ma08g32850-26-f:attcctaacccactacagagacttcttcgag(seq id no.58)
139.ma08g32850-26-r:ctataccatctgttgccgtcgaacacc(seq id no.59)
140.ma08g32850-wt-f:ctcagaaacccactacagagacttcttcgag(seq id no.60)
141.ma08g32850-wt-r:tcgaagcatctgttgccgtcgaacacc(seq id no.61)
142.以上引物名称中的13,15,16,18,21,25,26分别为7个阳性株系的编号。
143.pcr反应程序,除了延伸时间为20s外,其他的与第一轮的相同。
144.然后对pcr产物进行胶回收,每个样品取400ng送北京迈基诺公司进行高通量测序,测序结果分析软件为xshell、xftp和notepad,脱靶预测网站为http://www.rgenome.net/cas-offinder/。
145.结果表明,7个阳性株系都检测到突变,与对照相比,突变体中ma04g15900(seq id no.16)和ma08g32850基因(seq id no.17)的突变率均较高。
146.seq id no.16:
147.atggactgcaatcccacctccatcctcctgcgcactcccttggacctcaacaaggagcgagacgctggcgccggcggcgtcgtcttcaactcttcggtcctcgggaaccaagccagcatacccaaggcattcatctggccgcagtgccacagacccgccaccatcgacgatctcgacgccccggtggtggacctcggcggctttctccgcggcgacgaagcgtcgacggcgcgcgcggtggagtacgtcagggccgcctgctccacccacggcttcttccaggtcgccaaccatggcgtcgacgcgtcgctggcccgagaggcgatggactgcgttgacgggttctttaagctccctctctgtcacaagctacgcgcccggcggaagcccggtagcgtgtggggctacaccggggcgcacgccgaccgcttctcctccgagctgccatggaaggagaccctctcctttggctaccacgaggccggcggcggcgaccgcgtcgtcgtcgactacttcgcttccatcttaggccctgatttcgataggacggggtaagccatcgcaccatcttacgataagcacgaatcggcgaaatccactggcattgccgctgaagctttcttgtacaccaggctggttcttcagaggtactgcgaggcgatgaagaagctatcgctggtgatcatggagctcctggcaataagcttaggagtggaaagaagccactacagagacttcttcgaggacagttgctcaataatgagatgcaactactatcctccatgtcaggagccggagctgacgctcggcaccggcccgcactgcgaccccacctcgctgaccatccttcaacaggaccaggtggagggcctcgaggtcttctccgccaacaagtggcggtcggtccggccgattcgcgacgctttggtgatcaacattggtgacaccttcatggtacgtaacctgctgcctctctttctttctatagacaagactaatagtagtggagacgccaaagggcaaacaccatggagtaacagtgcagagttgctggtcgctgtctcctcttctttactgtttgcttgatcacggtaggaccagatggaaacattagcatcaaataattaagccaggattaagctcaaaa
aaaagaaaaaagaaaaaaaaagacacatacatacatacatacatacatacatcatcaactacaccaatgcttgtcttaggttcacaggattgagtatgtgctagacgacagggttcgatgtgcagcgctcaaaatgatgtgggtgcgtccccgagagggacagctgcttatgtattttacgtacccgttttatatgtgctatgctataaaacgtgtttggcccaattttactacgagtgcactcagatgcatatatccttttctaagtgattgaagcgtccgactccaaaacactatcgaacaggacgtgtgttttgtatttaatatctaatgtctcagatattgtacactaaaattcgatctttgcaagttggtgtacctattcaaagtttgcttttagattttataggaagcttccaaagaaattaaagagaaaagagagagagagagagagagagagagagagagaaccaaggaagaacctgaatgttctattgcgatgatgaagaatggctcccatgtctaattattgtatgcccccctgtctttgttctgcaacataggcgttgtccaatggaaggtacaaaagctgcctgcaccgggctgtggtgaacaggcatcgggaacggaagtcgctggctttcttcgtgtgccccagggaggaccgcaccatccggccaccgccgggggacctcggcggctcgcggctgtacccggacttcacgtgggcggagttcatgcagttcacgcagcgccactaccggtctgacacgaggacactccacagcttcaccaattggctctcctcctctaactccctccgccagtccacctag
148.seq id no.17:
149.atccctggtgtgtcctccgcaatggactgcaaccccacctccgttgtcctgcgcccttccttggccctcgataaagagagagaagccggctccggcggcgtcgtcttcagctctgcgctcctcccgaaccaagccaccatccccaaggccttcatctggcctcagtgcgacaggccgaccgccatcgaggagctggactcccctgtcgtagacctcgatggctttcttcgtggcgacgaagcgtccacggcgcgggcggtcgactgcgtcagggccgcctgctccacccacggcttcttccaagtcgccaaccacggcgtggacgcgtcgctcggccgggacgcgcttggctgcatggacgagttcttcgggctcccgctctgccacaagttacgcgcccggaggaagcccggaaacatgtggggctacgtcggcgctcacgccgaccgcttctcctcggagctgccctggaaggagaccctctccttcggctatcatgaggccggcgacgaccgcgtcgttatcgactacttcacgtccaccttgggcaatgatttccagaggatggggtgagccattagattaccctacaatcgacatgcatcgagacccaaagggaatggcattgccattcacgtcgcattctaacctcttcggtgcttgattaggttggtcttccagagatactgcgaggccatgaagaagctatctctggtgatcatggagctcctggcgatgagcctgggagtggacagaacccactacagagacttcttcgaggacagtcggtcgataatgagatgcaactactaccctccatgtccggagccggagctgacgctcggcaccggcccacactgcgatcccacctcgctcaccattcttcgacaggaccaagtggacgggctcgaggtgttcgacggcaacagatggcggtccgtccggcctattcgcgacgcccttgtgatcaacatcggtgatacgttcatggtacgtgagctgtctctgcgcctactgattactcaaaagttctctttctattaggtgtacatgcaacataaaatgttcggtaatttagacactgcaaaactgagattacgtacttagaagacaattccactgctgggaaccttaatttccttgatgaagtgaatccatgtatggcatgtcggtgaagtgcatcaaatccaagatttagcagcaatatttaggatgatttgctgcatgccatatcatgtcagccacaagctcatcatcttcaacattctgactaactcttccgcaaagtaatccactaaaagagagatgagaggacacacaagatgtaaagattcaggagcatggttgtctggtttgtcatcttgcagaaagaaacagatctgcattgtcttattgcaacagtgaaatcacctgcaatttctgatagcttctgtgcctttgattttggttttgtttgggaaccaaggcactgtccaacgggaggtacaaaagctgcctgcaccgggccgtggtgaacaggcgccggcaacgaaagtcgctggccttcttcctctgccccagggaggaccgcgtggtccggccgccgccgggggagctcggcggcgcgaggctgtacccggacttcacgtgggcggagctgttggagttcacgcagcgccactaccgggccgacatgaagacgctccacagcttcaccgactggcttctctcctcctcttccccatcccctctccggtccacctgatcatcccccttctccctcatccacccactgtaactcacaacagaccgaataatctatagacgccatgcgttctagttcatcgatcgtctccgttttaggactggttcgtggatctcgagttaggagttgaatttgcaaggtgggaagaggagaggtggacgtgatggcgccacaaatgacaagcgaccaacacatgaagttacaggagcagatccggtatctgata
150.在ma04g15900(seq id no.16)和ma08g32850基因(seq id no.17)中,针对序列为
seq id no.1的靶点,主要检测到1、2、3、8、13和19碱基缺失以及1和3碱基插入等8种类型的突变,其中单碱基插入( t)和单碱基缺失(-g或-c)是主要的突变类型,分别在7、6和5个株系中检测到,其次是13碱基缺失和单碱基插入( g)类型的突变,分别在2个株系中检测到,然后是2、3、8和19碱基缺失以及3碱基插入类型的突变,分别在1个株系中检测到(表1-2)。-g类型的突变在ma04g15900和ma08g32850基因中均检测到,且碱基缺失的位置一致,而-c类型的突变只在ma04g15900基因中检测到(表1)。
151.表1 ma04g15900的靶位点突变后的序列及对应突变类型、植株
152.5'-ccg-gagctgacgctcggcaccgg-3'wtseq id no.1seq id no.5'-ccg-ga*ctgacgctcggcaccgg-3'-1plant13,16,18,21,25185'-ccg-gag*tgacgctcggcaccgg-3'-1plant15,16,18,21,25195'-ccg-gag***acgctcggcaccgg-3'-3plant 21205'-ccg-g***tgacgctcggcaccgg-3'-3plant 13215'-ccg-gagc********ggcaccgg-3'-8plant 15225'-ccg-g*******************-3'-19plant 13235'-ccg-gagggcctgacgctcggcaccgg-3' 3plant 1624
153.表2 ma08g32850的靶位点突变后的序列及对应突变类型、植株
154.5'-agccg-gagctgacgctcggcaccgg-3'wtseq id no.25seq id no.5'-agccg-ga*ctgacgctcggcaccgg-3'-1plant13,15,16,18,21,25265'-agccg-g**ctgacgctcggcaccgg-3'-2plant 25275'-agccg-gagtctgacgctcggcaccgg-3' 1plant13,15,16,18,21,25,26285'-agccg-gaggctgacgctcggcaccgg-3' 1plant 13,15295'-agccg-g*************caccgg-3'-13plant 13,16305'-*******************caccgg-3'-19plant 1331
155.其中*是为显示突变体植株的靶位点序列与野生型(wt)相比而作的标记,不代表任何碱基,即*标记表示基因片段中碱基被敲除。
156.实施例5:
‘
grand nain’转基因阳性株系的半矮化表型
157.将萌发的芽转入生根培养基后,定期观察转基因和对照株系的生长情况,包括株高、叶片及根的生长。当移栽至营养杯,并转移到温室培养后,定期观察植株的长势,在温室培养45d后,分别测量各株系的株高,之后将各株系转移至大盆(直径48cm,高38cm)中继续生长,并且在培养100d和180d后,分别测量各株系的株高,在温室培养100d后测株高的同时,测量每个株系展开叶(从上向下数)的第一至第四个叶片的叶长和叶宽。
158.转移至温室培养的突变体,早期随着培养时间的延长,表型变化不明显,突变株系与对照的高度相近,其叶片的大小和形态也与对照相似(图5a)。选取其中7个株系在培养45、100和180d后分别进行株高的测定,结果表明,在后两个发育时期,突变体的株高均显著低于对照植株(图5b)。突变体在温室中培养120d后的表型如图5c。突变体在生长过程中,其叶片与对照相比有一定差异,在温室培养100d后测量了各株系的叶长和叶宽,结果如图5d,可以看出,突变体的叶长和叶宽均显著短于对照。这些结果表明,突变体的叶长和叶宽与其植株高度的变化是一致的,即植株变矮,其叶长和叶宽也相应变短和变窄。
159.实施例6:
‘
grand nain’转基因阳性株系gas含量的分析
160.gas是影响植物生长发育的重要因素之一。为了检验靶位点的突变对gas生物合成
的影响,选取在温室培养180d后的
‘
grand nain’具有典型半矮化表型的2个株系和对照,分别取其上部新鲜叶片,采用高效液相色谱-质谱联用(hplc-ms/ms)技术,检测样品中多种内源gas的含量。具体步骤如下:
161.1、激素提取
162.a.将选取的各个株系叶片的全部样品在液氮中充分研磨至粉碎,准确称量约1g样品;
163.b.向粉末中加入异丙醇/盐酸提取缓冲液10ml,在4℃条件下震荡30min;
164.c.接着加入20ml二氯甲烷,在4℃条件下震荡30min;
165.d.在4℃条件下,以13000rpm离心5min,取下层有机相;
166.e.在避光条件下,以氮气吹干有机相,然后用400μl甲醇(含0.1%甲酸)溶解;
167.f.用0.22μm滤膜过滤后,进hplc-ms/ms检测。
168.2、液质检测
169.a.标准溶液配制。以甲醇(含0.1%甲酸)为溶剂配制梯度为0.1、0.2、0.5、2、5、20、50和200ng/ml的ga1/3/4/5/6/8/
19
/
20
/
29
/
44
/
53
标准溶液,并在实际绘制标准曲线方程时去掉线性不好的点。
170.b.液相条件。色谱柱:poroshell 120sb-c18反相色谱柱(2.1
×
150,2.7μm);柱温:30℃;流动相:a:b=(甲醇/0.1%甲酸):(水/0.1%甲酸);洗脱梯度:0-1min,a=20%;1-9min,a递增至80%;9-10min,a=80%;10-10.1min,a递减至20%;10.1-15min,a=20%;进样体积:2μl。
171.c.质谱条件。气帘气:15psi;喷雾电压:4500v;雾化气压力:65psi;辅助气压力:70psi;雾化温度:400℃。
172.本试验测定了
‘
grand nain’代表性株系15和26号叶片中内源gas的含量,结果如图6。可以看出,与对照相比,2个株系中叶片ga
53
和ga
19
的含量均极显著降低,ga
20
的含量极显著升高;活性gas只测出ga1和ga3,其中ga1的含量均极显著降低,对照和突变体中ga3的含量均较低;非活性ga8的含量显著降低,对照和突变体中ga5的含量均较低,而对照ga6的含量稍高,2个株系ga6的含量均有降低,但不显著。以上的结果表明,
‘
grand nain’叶片中gas的生物合成和分解代谢均有减弱。
173.实施例7:
‘
grand nain’转基因阳性株系细胞结构的变化
174.‘
grand nain’突变体除了变矮一些,其叶片的变化不明显。为了检验靶位点的突变对植株叶片结构的影响,选取在温室培养100d后的
‘
grand nain’3个株系和对照,分别取其植株中部的叶片,且每个株系选取相同的部位,将叶片沿叶脉的方向剪成0.5cm
×
0.5cm大小,剪下后均立即放入70%faa固定液中,然后参考li等(2015)的方法,将材料经固定、脱水、渗蜡、包埋、切片、展片与粘片、烘干、脱蜡、染色、封片等一系列步骤,做成石蜡切片。将石蜡切片于leica dm4b正置荧光显微镜下进行观察并拍照,同时添加合适的比例尺。
175.通过叶片细胞结构的石蜡切片分析发现,选取的15,17和26号等3个突变体的细胞结构是类似的,因此,以代表性突变体15号的为例,分析这种细胞学方面的差异,结果如图7(图7a和b分别表示wt和15号株系植株中部叶片的横切面,bar=100μm,其中,uc为上角质层,ue为上表皮,pl为栅栏组织,sl为海绵组织,le为下表皮,lc为下角质层。)。可以看出,对照株系叶片的角质层细胞规整、平滑,上表皮和下表皮细胞呈规则的圆球状(图7a),而突变
体的角质层细胞、上表皮和下表皮细胞也呈规则的状态(图7b)。这些结果表明,编辑后的突变体在植株高度变矮的同时,其叶片的细胞形态没有显著变化。
176.以上所述实施例的各技术特征可以进行任意的组合,为使描述简洁,未对以上实施例中的各个技术特征所有可能的组合都进行描述,然而,只要这些技术特征的组合不存在矛盾,都应当认为是本说明书记载的范围。
177.以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。