tomato plants: similarity of functional domains to subtilisin
‑
like endoproteases. proceedings of the national academy of sciences of the united states of america 93: 6332
‑
6337.)。在拟南芥中,枯草杆菌类蛋白酶sbt3.3正调控其对细菌和真菌的抗性(ramirez v, lopez a, mauch
‑
mani b, gil mj, vera p (2013) an extracellular subtilase switch for immune priming in arabidopsis. plos pathogens 9.)。在马铃薯中,沉默半胱氨酸蛋白酶编码基因c14,导致其对致病疫霉更加感病(avrova ao, stewart he, de jong w, heilbronn j, lyon gd, et al. (1999) a cysteine protease gene is expressed early in resistant potato interactions with phytophthora infestans. molecular plant
‑
microbe interactions 12: 1114
‑
1119.)。番茄中的半胱氨酸蛋白酶pip1调控其对细菌、真菌和卵菌等病原菌的抗性(ilyas m, horger ac, bozkurt to, van den burg ha, kaschani f, et al. (2015) functional divergence of two secreted immune proteases of tomato. current biology 25: 2300
‑
2306.)。最近发现,番茄中半胱氨酸蛋白酶rcr3需要被质外体空间的p69b等蛋白酶激活,才能介导无毒蛋白效应子avr2和抗病蛋白cf2的识别,激发免疫反应。在拟南芥中还发现,天冬氨酸蛋白酶sap1和sap2可以切割丁香假单胞杆菌毒性效应子mucd,提高寄主抗性(wang y, garrido
‑
oter r, wu j, winkelmuller tm, agler m, et al. (2019) site
‑
specific cleavage of bacterial mucd by secreted proteases mediates antibacterial resistance in arabidopsis. nat commun 10: 2853.)。
5.上述研究结果表明,植物分泌的质外体蛋白酶在抑制病原菌侵染与提高寄主免疫过程中具有至关重要的作用。然而,水稻分泌的半胱氨酸蛋白酶在稻瘟病菌抗性中的作用机制仍不清楚。因此研究水稻半胱氨酸蛋白酶在稻瘟病菌抗性中的作用机制,不但可拓展人们对植物与病原菌互作机制的认识,同时为抗稻瘟病遗传育种与新型杀菌剂的研发提供参考。
技术实现要素:
6.基于本发明人的研究,首次发现osrd21蛋白具有半胱氨酸蛋白酶活性,能够对稻瘟病菌的外泌蛋白进行切割,使其失活,从而调控水稻对稻瘟病菌的抗性。osrd21基因在水稻中过表达的植株对稻瘟病菌具有显著抗性。由此完成本发明。
7.本发明首先提供一种水稻半胱氨酸蛋白酶,其特征在于,其氨基酸序列如seq id no.2 所示。
8.对应地,本发明还提供编码所述的水稻半胱氨酸蛋白酶的基因,其特征在于,其核苷酸序列如seq id no.1所示。
9.进一步地,本发明提供含有所述的基因的表达元件、重组载体、宿主细胞。
10.本发明还提供所述的水稻半胱氨酸蛋白酶、所述的基因在防治稻瘟病菌中的应用。
11.优选地,通过转基因方法在植物中过量表达所述基因。
12.本发明还提供所述的基因在制备抗病转基因植物中的应用,其中通过转基因方法在转基因植物中过量表达。
13.优选地,所述植物为单子叶植物,优选地所述植物为水稻。更优选地,所述抗病性
指抗稻瘟病菌导致的植物病害。
14.本发明涉及水稻半胱氨酸蛋白酶编码基因osrd21在抗病过程中的应用,osrd21基因的敲除不影响稻瘟病菌的正常侵染;但是,osrd21基因的过表达转基因植物,对稻瘟病菌具有显著的抗性,能够抑制稻瘟病菌在寄主细胞内扩展。并且在与稻瘟病菌互作过程中,osrd21蛋白具有半胱氨酸蛋白酶活性,能对稻瘟病菌分泌的效应子进行切割,使其丧失活性,从而显著提高对稻瘟病菌的抗性。本发明涉及的转基因植物并不影响正常的生长和结实。
15.进一步地,本发明提供一种增强植物对稻瘟病抗性的方法,其特征在于,通过转基因方法在转基因植物中过量表达所述的基因。
16.本发明人研究发现,在水稻与稻瘟病菌互作过程中,水稻中的osrd21基因被诱导上调表达,该蛋白转录水平的提高能够显著提高水稻对稻瘟病菌的基础抗病性。过表达植株对稻瘟病菌表现为抗性,可推广实际应用。本发明有利于水稻抗病品种的培育,为后期筛选高抗水稻品种提供依据。例如,本发明能够提供osrd21基因过表达植株,利用ems诱变获得osrd21基因的沉默抑制子,并获得对应的转基因植株;另外,本发明能够进一步提供或应用利用上述转基因植株或者种子,进行杂交获得更高抗的转基因水稻植株。
附图说明
17.图1为osrd21基因响应稻瘟病菌侵染。水稻tp309接种稻瘟病菌后,不同时间段,osrd21基因转录水平的变化。
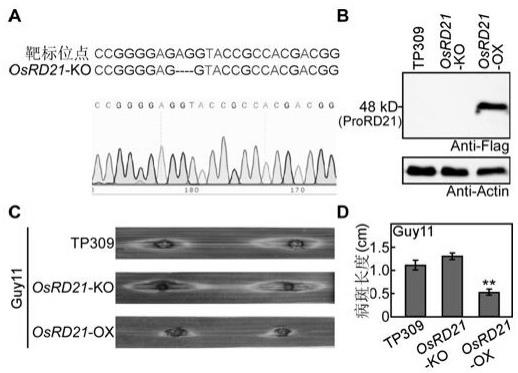
18.图2为osrd21基因敲除和过表达突变体的获得及致病性测定。其中,a为利用crispr/cas9技术获得osrd21基因敲除突变体靶标序列的构建,以及目的基因敲除的测序结果;b为osrd21基因敲除和过表达植株western验证结果;c为osrd21基因敲除和过表达植株稻瘟病菌抗性测定实验;d为病斑大小的统计结果。
19.图3为osrd21基因编码蛋白的半胱氨酸蛋白酶活性测定。
20.图4为osrd21基因编码蛋白对稻瘟病菌效应子的切割活性测定。其中,a为osrd21蛋白在烟草中表达纯化后western blot检测结果;b为稻瘟病菌的效应子mosef1与osrd21蛋白在烟草细胞中共表达后western blot检测结果。
具体实施方式
21.下面通过具体实施例对本发明进行阐述,以期更好的理解本发明,但不构成对本发明的限制。
22.实施例1:osrd21基因响应稻瘟病菌的侵染在稻瘟病菌与水稻互作早期,水稻感知病菌侵染后会通过抗病相关基因的上调表达,抵抗稻瘟病菌的侵染。水稻中的半胱氨酸蛋白酶包括rd21在植物免疫中具有重要作用,然而其是否响应稻瘟病菌侵染还不清楚。为了研究osrd21基因是否在抗稻瘟病过程中发挥作用,本发明利用荧光定量pcr技术,检测水稻中的osrd21基因的转录水平在稻瘟病菌侵染后是否上调表达。用稻瘟病菌野生型菌株guy11的孢子悬浮液,产孢方法详见实施例2。将上述孢子悬浮液调制浓度为1
×
105个/ml,喷晒到水稻感病品种tp309叶片上,先黑暗保湿24小时,随后光暗交替培养,分别提取侵染不同阶段(8、24、36、48和72小时)的水稻叶片rna。
rna的提取利用天根公司的rna提取试剂盒;提取后的rna反转录合成cdna,反转录利用takara的反转录试剂盒完成。设计特异的定量引物f1/r1(f1: gttgctattgaggctggtgg,r1:tccatgcgaacgtatccaga)用于检测osrd21基因的转录水平。荧光定量pcr反应体系为20
µ
l,包括cdna 20 ng,引物各0.2
µ
m, sybr premix extaq (takara bio inc. shiga, japan) 10
µ
l,及无菌超纯水6.8
µ
l。所用仪器为abi prism 7500,数据分析采用7500系统测序检测软件(v.1.4)。
23.结果显示,相比于不接种稻瘟病菌的水稻(0小时),侵染24、36、48和72小时后的水稻中osrd21基因的转录水平均具有显著的上调表达,尤其是72小时其上调倍数超过3倍(图1)。上述结果表明osrd21基因响应稻瘟病菌的侵染上调表达,预示其在抵抗稻瘟病菌侵染的过程中发挥重要作用。
24.osrd21基因核苷酸序列如seq id no:1所示,其氨基酸序列如seq id no:2所示。
25.实施例2:osrd21基因敲除突变体的获得及抗性测定cm培养基配置方法:量取20
×
硝酸盐(120g硝酸钠,10.4g氯化钾,10.4g七水硫酸镁,30.4g磷酸二氢钾,加蒸馏水溶解至1l) 50ml,1000
×
微量元素(2.2g七水硫酸锌,1.1g硼酸,0.5g四水氯化锰,0.5g七水硫酸铁,0.17g六水氯化钴,0.16g五水硫酸铜,0.15g二水锰酸钠,5g edta四钠,加蒸馏水溶解至100ml) 1ml,维生素溶液(0.01g生物素,0.01g维生素b6,0.01g维生素b1,0.01核黄素,0.01对氨基苯甲酸,0.01烟酸,加蒸馏水溶解至100ml) 1ml,称取葡萄糖10g,蛋白胨2g,酵母提取物1g,酪蛋白氨基酸1g,琼脂粉15g,加蒸馏水定量至1l,分装于三角瓶中,121℃灭菌20分钟,冷却后备用。
26.稻瘟病菌产孢培养基配置方法:用玉米粉和水稻秆配制,称取100g水稻秆加入1l水煮沸30分子,再加入40g玉米粉和15g琼脂粉煮沸20分钟,最后用蒸馏水定容到1l,分装于三角瓶中,121℃灭菌20分钟,冷却后备用。
27.水稻叶片穿刺接种稻瘟病菌方法:1.首先诱导稻瘟病菌产生分生孢子,将cm培养基上的稻瘟病菌guy11菌丝块接种到sdc培养基上,28℃黑暗培养4天,然后将表面菌丝刮掉,黑光灯下诱导3天,即可获得分生孢子;2.温室培养1月左右的水稻苗用于接种实验,保湿平铺于培养皿中,叶片用针孔穿刺扎伤;3. 收集上述sdc平板上的分生孢子,浓度为1
×
105个/ml,点滴到上述水稻叶片穿刺部位,黑暗保湿培养24小时,随后光暗交替培养5
‑
7天。
28.4. 统计病斑长度。
29.5. 每个处理重复三次。
30.通过crispr/cas9的方法,根据http://cbi.hzau.edu.cn/cgi
‑
bin/crispr网站,设计osrd21基因的敲除靶标序列(图2中a)。将该序列构建到psgrna载体上,再与pcas9进行gateway反应,转化大肠杆菌获得质粒后测序,再通过农杆菌介导的转化在水稻感病品种tp309背景下获得敲除osrd21基因的敲除突变体。获得突变体水稻苗后,提取叶片基因组,设计靶点附近的测序引物,克隆osrd21基因靶点附近的序列约400 bp,送公司测序,结果表明该基因已被敲除(图2中a);本发明通过水稻叶片穿刺接种稻瘟病菌野生型菌株guy11孢子液,浓度为1x105个/ml,发现osrd21基因的沉默突变体的病斑长度(1.3cm)较对照
(1.1cm)更长,说明osrd21基因的沉默突变体更为感病(图2中c)。
31.实施例3:osrd21基因过表达植株的获得及抗性测定利用酶切连接的方法将目的基因osrd21的编码区(seq id no:1所示序列)构建到pcam2300载体中,引物设计时osrd21基因上下游引物分别添加xbai和psti酶切位点(f2:gctctagaatggccgcccgcgccgcc, r2:aactgcagtgcggtgttcagctta),扩增osrd21基因;再将目的基因和载体用限制性内切酶xbai和psti进行酶切线性化,通过t4连接酶将载体和片段进行连接,连接体系为10ul(包括1
µ
l 线性化载体骨架,4
µ
l osrd21基因片段和5
µ
l连接酶),16℃连接6小时,转化大肠杆菌后验证菌落,获得阳性质粒,即构成pcam2300
‑
osrd21的载体。pcam2300载体由actin启动子启动基因表达,并自带flag标签,可用于后期的验证。
32.将制备的载体转化水稻,获得osrd21基因过表达的植株;获得的转基因植株, 通过western杂交实验检测,发现osrd21基因翻译形成的蛋白正常表达,在sds
‑
page胶上能够检测到目的条带,而对照则没有(图2中b),证明获得的转基因植株能用于后续实验。
33.利用水稻叶片穿刺接种稻瘟病菌方法测定稻瘟病菌抗性,具体方法详见实施例2。收集稻瘟病菌野生型菌株guy11孢子液,浓度为1x105个/ml,发现osrd21基因的过表达水稻的病斑长度(0.5cm)显著小于对照(1.1cm),说明osrd21基因过表达水稻更为抗病(图2中d)。上述结果表明osrd21基因的过表达植株能够显著提高水稻对稻瘟病菌的病性。
34.实施例4:osrd21蛋白半胱氨酸蛋白酶活性测定生物素标记的dcg
‑
04测定半胱氨酸蛋白酶的活性方法:将0.5mg目的蛋白溶解在50mm的乙酸钠溶液中,ph调制6.0,再加入10mm l
‑
半胱氨酸和2
µ
m的dcg
‑
04。对照中加入0.2mm的e
‑
64(e3132,sigma),室温下孵育5小时。孵育后的蛋白用加入两倍体积的预冷丙酮沉淀蛋白,10000g离心1分钟,去掉上请,加入70%的丙酮洗两次,晾干。沉淀用tbs溶解,加入蛋白上样缓冲液,煮沸5分钟。利用链霉亲和素偶联的hrp,通过western杂交的方法测定活性,western杂交后在sds
‑
page胶上能显示出条带的证明具有半胱氨酸蛋白酶活性(图3)。
35.利用pbin
‑
gfp载体,含35s启动子,克隆osrd21基因的cdna全长,并通过酶切连接的方法构建获pbin
‑
osrd21
‑
gfp的载体;通过农杆菌介导的转化,在烟草细胞中表达osrd21基因,并利用商品化的gfp珠子(chromotek),将osrd21蛋白进行纯化。进一步利用生物素标记的dcg
‑
04测定半胱氨酸蛋白酶的活性。研究结果显示,在只加入dcg04而不加入其他蛋白或者加入对照蛋白的情况下,能够在sds
‑
page胶上检测到osrd21蛋白条带,且大小为22 kd;而加入半胱氨酸蛋白酶抑制子e64和moers1的情况下,不能检测到osrd21蛋白的条带;上述结果表明osrd21蛋白具有半胱氨酸蛋白酶活性(图3)。
36.实施例5:osrd21蛋白对稻瘟病菌效应子的切割活性测定利用实施例4中的pbin载体,在烟草细胞中共表达水稻中osrd21基因及其缺失突变基因(融合表达gfp标签)和稻瘟病菌效应子编码基因mosef1(融合表达his标签),其中缺失突变分别指缺失seq id no.2前端的140个氨基酸或末端的85个氨基酸;共表达48小时后,提取烟草叶片总蛋白,利用gfp
‑
trap珠子进行蛋白的纯化。同时利用实施例4中的方法对osrd21基因及其缺失突变基因编码蛋白的半胱氨酸蛋白酶的活性进行测定,western结果显示这些蛋白在sds
‑
page胶上能够检测到目的条带,且大小正确,表明他们能够正常表达且具有半胱氨酸蛋白酶活性(图4中a);同时,western杂交结果显示,稻瘟病菌的效应子mosef1与osrd21蛋白在烟草细胞中共表达后,mosef1蛋白在sds
‑
page胶上显示出多条非目
的条带(图4中b中星号标注的条带),而单独表达mosef1蛋白则只有目的条带,表明osrd21蛋白的半胱氨酸蛋白酶活性能够对稻瘟病菌的效应子mosef1进行切割(图4中b)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。