1.本发明涉及用于包含nc886的增强溶瘤(oncolytic)病毒活性的组合物及包含其的用于促进病毒生产的组合物。
背景技术:
2.溶瘤病毒(oncolytic virus)的特征为操纵活病毒(virus)的基因,不作用于正常细胞,针对癌细胞特异性增殖来破坏癌细胞。并且,除直接攻击癌细胞之外,具有促进人体适应性免疫反应的功能(immune stimulation),还具有特异性感染并破坏肿瘤血管内皮细胞来抑制新血管生成的功能。1904年,dock g研究组证实并报告了狂犬病病毒对于宫颈癌患者的抗癌作用,从此以后,正在进行并报告使用禽病毒、感冒病毒等的病毒的抗癌研究。2015年10月,美国安进(amgen)首次获得美国食品药品监督管理局(fda)的批准并销售黑色素瘤治疗剂lmlygic(talimogene laherparepvec),之后各种溶瘤病毒的研究和临床试验备受关注。但是,难以大量生产溶瘤病毒,当适用于临床时,具有活性甚微的问题。
3.并且,若溶瘤病毒真正进入细胞,则感染率低,具有无法治疗的问题。因此,临床效果受限,目前正在尝试与现有的免疫检查点抑制剂结合来治疗。
[0004][0005]
丙型肝炎病毒(hcv,hepatitis c virus)是一种据信全球感染人数超过1.8亿人的病原体,在未有效治疗的情况下,以引起如肝纤维化、肝硬化、肝癌的疾病而周知。丙型肝炎病毒具有6种基因型,均匀地分布在世界各地,根据地域和种族分布不同。在致病性丙型肝炎病毒感染中,1型感染人数最多,其次为2型,最后为3型,遗传1型大多分布在欧洲和北美洲,最难治疗,遗传2型合3型在远东地区多发。尤其,难以治疗的遗传1型丙型肝炎病毒患者占丙型肝炎病毒感染人数的大多数。全球人口的1%~2%被该病毒感染,但是,病毒以非常低的滴度(titer)存在于生物体内,因此,非常难以诊断,目前没有对于丙型肝炎病毒有效的治疗剂或疫苗。目前具有同时给药通过抑制丙型肝炎病毒的增殖来对于丙型肝炎病毒引起的疾病具有一定程度的疗效的干扰素
‑
α和利巴韦林(ribavirin)的方法,但据报告,该方法对于每种病毒株的效果不同,仅对40%的丙型肝炎病毒感染患者中具有持续效果,诱发许多副作用。为了解决如上所述的问题,正在进行许多研究。例如,在韩国公开专利号第10
‑
2012
‑
0131864号中公开了对于靶物质特异性地释放小干扰核糖核酸(sirna)的用于干扰核糖核酸(rna)的组合物及利用其来治疗丙型肝炎病毒相关疾病的组合物,在韩国公开号第10
‑
2011
‑
0046321号中公开了包含吲哚化合物的药剂学组合物。但是,现有的病毒治疗剂的治疗功效甚微,由此对于更有效的病毒治疗剂的要求不断增加。
技术实现要素:
技术问题
[0006]
本发明人在研究新近发现的作为非编码核糖核酸(non
‑
coding rna)的nc886时发
现,nc886促进病毒复制来帮助病毒的生产,提供原病毒(pro
‑
viral)的环境来增强病毒活性。确认了当nc886被基因敲除(knock
‑
down)时,病毒的信使核糖核酸(mrna)及蛋白质显著减少。如上所述,确认了在使用nc886拮抗剂的情况下,具有预防及治疗病毒的效果。如上所述,本发明提供包含nc886的用于增强溶瘤病毒活性的组合物、包含其的用于促进病毒生产的组合物和病毒生产方法以及包含nc886拮抗剂的病毒治疗组合物。
[0007]
在本发明中,在病毒感染的初始阶段,通过给药nc886拮抗剂来抑制病毒的增殖。并且,本发明提供如下的组合物:当利用病毒治疗癌症时,插入了nc886的病毒可增加癌症治疗效率。技术方案
[0008]
本发明可提供包含nc886或其活化剂的用于促进病毒生产的组合物。
[0009]
并且,本发明可提供包括添加nc886或其活化剂的步骤的病毒生产方法。
[0010]
并且,本发明可提供包含nc886或其活化剂的用于增强溶瘤病毒治疗剂的活性的组合物。
[0011]
并且,本发明可提供包含nc886或其活化剂、以及溶瘤病毒治疗剂的用于预防或治疗癌症的药学组合物。
[0012]
并且,本发明可提供溶瘤病毒治疗剂中的溶瘤病毒治疗辅助候选物质的筛选方法,其包括:步骤(a),将表达nc886的细胞与试验物质及抗癌病毒相接触;步骤(b),在上述细胞中分析上述nc886的表达程度;以及步骤(c),将相对于未处理上述试验物质的对照组使上述nc886的表达增加的试验物质判断为抗癌病毒辅助剂候选物质。
[0016]
并且,本发明可提供包含nc886的表达抑制剂的用于预防或治疗病毒的药学组合物。
[0017]
并且,本发明可提供用于治疗病毒感染的候选物质的筛选方法,其包括:步骤(a),将表达nc886的细胞与试验物质相接触;步骤(b),在上述细胞中分析上述nc886的表达程度;以及步骤(c),当上述试验物质减少上述nc886的表达时,判断为用于治疗病毒感染的候选物质。发明的效果
[0021]
本发明提供nc886的药剂学用途,上述nc886提供原病毒的环境,本发明的包含nc886或其活化剂的组合物具有如下的效果:促进病毒复制来帮助病毒的生产,并可增强病毒的活性,尤其,溶瘤病毒治疗剂的功效。并且,本发明的包含nc886拮抗剂的组合物具有预防及治疗病毒的效果。
附图说明
[0022]
图1为示出nc886促进病毒复制的结果,图1a为指定的肝细胞中的作为内参对照组的利用溴乙烷(etbr)染色的指示非编码核糖核酸(ncrna)的诺瑟杂交结果,图1b及图1c为在感染丙型肝炎病毒后标记的时间点nc886被基因敲除或未被基因敲除的丙型肝炎病毒及人类宿主蛋白的蛋白质印迹结果,图1d为在指定的时间测定丙型肝炎病毒信使核糖核酸的结果(丙型肝炎病毒感染24小时之前转染对于nc886(以
‑
886记载)或穹窿体核糖核酸1
‑
1(vtrna1
‑
1)(以
‑
con记载)的抗寡剂,“none”表示未转染抗寡核苷酸(anti
‑
oligo)),图1e为利用抗
‑
仙台病毒(sev)抗体(anti
‑
sev antibody)执行的蛋白质印迹结果(仙台病毒被标
记的细胞株感染,在感染后的标记的时间获取细胞,进行裂解来用于蛋白质印迹,并将带定量化),图1f为以moi 1将过表达或未过表达nc886的293细胞感染呼吸道合胞病毒(rsv)后,在不同时间点准备总细胞裂解物,并执行用于检测病毒蛋白的蛋白质印迹的结果,图1g为将腺病毒(adv)感染至指示的细胞株并测定腺病毒粒子的结果。
[0023][0024]
图2为示出nc886抑制干扰素(ifn)反应的结果,图2a为示出感染病原体时的干扰素反应的示意图,图2b为当进行聚肌胞苷酸(poly(i:c))处理时测定干扰素b1的实时荧光定量聚合酶链式反应(qr t
‑
pcr)的结果(转染聚肌胞苷酸后标记的时间分离核糖核酸,通过实时荧光定量聚合酶链式反应测定干扰素b1和18s核糖体核糖核酸(18s rrna),将数值归一化为18s核糖体核糖核酸,并将293t:载体(vector)的0小时的值设置为1),图2c为利用包含干扰素
‑
β启动子元件的荧光素酶报告质粒及内部荧光素酶对照质粒将293细胞(w/wo 886过表达)共转染的结果(荧光素酶质粒转染0小时,仙台病毒及呼吸道合胞病毒转染9小时,试验(assay)在24小时后完成),图2d及图2e为利用包含干扰素的干扰素刺激反应元件位点的多聚剂及内部荧光素酶对照质粒的荧光素酶报告质粒将293细胞、w/wo 886过表达共转染,转染15小时后,在mock或moi 1的条件下使细胞感染呼吸道合胞病毒/仙台病毒,感染24小时后,裂解细胞以测定荧光素酶活性(图2d),或者获取上清液来测定rantes(也是干扰素调节因子
‑
3(irf
‑
3)调节介质,图2e)的诱导的结果。
[0025][0026]
图3为示出nc886通过干扰素启动子的各种因子诱导干扰素反应的结果,图3a为示出干扰素b1信使核糖核酸表达的结果,图3b为示出上述核因子κb(nf
‑
κb)靶信使核糖核酸被nc886表达弱化的结果,图3c为利用示出核因子κb的活性的荧光素酶确认核因子κb的活性的结果,图3d为示出使用具有4个干扰素调节因子3/干扰素调节因子7
‑
结合基序的报告质粒来确认荧光素酶的结果,图3e及图3f为测定作为干扰素调节因子3的活性形态的磷酸(phospho)
‑
干扰素调节因子3的结果,确认了当表达nc886时,其活性减少,当基因敲除nc886时,其活性增加的结果。
[0027][0028]
图4为示出如下的结果,即,在蛋白激酶(pkr)被基因敲除的情况下,当感染腺病毒时,病毒基因的表达增加,在此基因背景下,若nc886也被基因敲除,则表达减少。
[0029][0030]
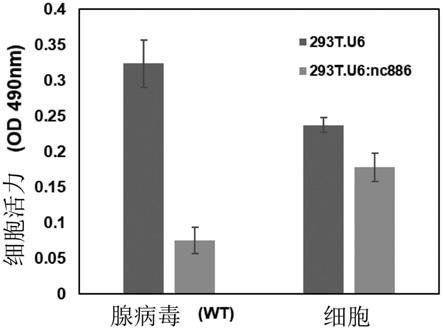
图5为示出在nc886存在的情况下杀死癌细胞的活性(溶瘤作用,oncolytic effect)增加的结果。
具体实施方式
[0031]
以下,更详细地说明本发明。
[0032][0033]
本发明的nc886具有如下的效果:促进病毒复制,例如,当利用溶瘤病毒治疗癌症时,增强其的治疗效果。
[0034][0035]
本发明可利用nc886或其活化剂来促进病毒生产。尤其,当生产病毒时,可添加nc886或其活化剂来稳定地生产病毒。
[0036][0037]
丙型肝炎病毒的复制在一部分肝细胞株中进行。本发明人发现,在可复制丙型肝炎病毒的huh7类的肝细胞株(huh7、huh7.5、ft3
‑
7)的情况下,nc886的表达水平在原代肝细胞中高,相反,在未复制丙型肝炎病毒的肝细胞株的hepg2及hep3b中是沉默的(图1a)。由此,为了确认在丙型肝炎病毒复制中的nc886的作用,利用反义寡核苷酸在huh7.5细胞株中基因敲除nc886的结果,蛋白激酶在基因敲除nc886的细胞中活化。并观察到丙型肝炎病毒的信使核糖核酸及蛋白量减少,从上述结果确认了nc886对于复制丙型肝炎病毒是必需的(图1b及图1d)。为了确认nc886是否对于其他类型的病毒也有效,利用仙台病毒、呼吸道合胞病毒、腺病毒进行追加试验。在使表达nc886的293t细胞感染呼吸道合胞病毒及仙台病毒的结果,确认了病毒的增殖增加(图1e~图1f)。进而,确认了在利用crispr
‑
cas技术删除nc886的nthy
‑
ori 3
‑
1细胞中,腺病毒的增殖显著减少(图1g)。
[0038][0039]
丙型肝炎病毒为单正链核糖核酸(positive
‑
sense rna)病毒,仙台病毒和呼吸道合胞病毒为单负链核糖核酸(negative sense rna)病毒,腺病毒为双链脱氧核糖核酸(dna)病毒,因此,病毒基因表达及复制机制均不同。观察到各种病毒的增殖均被nc886促进,因此,nc886促进广泛范围的多种病毒的复制的可能性高,由此假设宿主的先天免疫反应抑制是促nc886的原病毒的作用的机制。为了确认其,研究了nc886对于主要宿主防御重要的干扰素
‑
β(ifn
‑
β,interferon
‑
β)信号的效果。干扰素
‑
β包含对于称为激活蛋白
‑
1(ap
‑
1)、干扰素调节因子及核因子κb的3种调节因素的结合位点(图2a)。聚肌胞苷酸(促进干扰素生产的合成核糖核酸)的转染诱导干扰素
‑
β的信使核糖核酸表达,相反,当nc886存在时,干扰素
‑
β的信使核糖核酸表达减少(图2b)。干扰素
‑
β启动子在作为不表达nc886的对照组细胞株的293t中通过聚肌胞苷酸、仙台病毒、呼吸道合胞病毒的感染活化,在制备成表达nc886的293t细胞中活化程度减少(图2c)。在干扰素
‑
β信号途径中,干扰素刺激基因(isgs)的转录活化是被干扰素刺激反应元件(isre)介导的。利用干扰素刺激反应元件测定荧光素酶表达的结果,irse荧光素酶示出与干扰素
‑
β荧光素酶相似的结果(图2d)。由此确认了nc886抑制干扰素
‑
β的信号。除干扰素
‑
β之外,当生产如rantes的趋化因子时,需要作为上述干扰素
‑
β启动子的3个调节因素的激活蛋白
‑
1、干扰素调节因子及核因子κb,经确认,当感染呼吸道合胞病毒时,nc886抑制rantes的诱导(图2e)。
[0040][0041]
评价了作为干扰素
‑
β启动子的结合的三种基因的核因子κb、激活蛋白
‑
1及干扰素调节因子(图2a)的贡献度。转染聚肌胞苷酸的结果,使干扰素
‑
β启动子活化,上述活化被nc886显著减弱。与之相比,在转染结构上活化状态的磷酸
‑
模拟干扰素调节因子3(phospho
‑
mimetic irf3)[=干扰素调节因子3(5d)]突变体的情况下,也使干扰素
‑
β启动子活化,上述活化也被nc886减弱。由于引入活化状态的干扰素调节因子3,在此条件下被nc886减弱的结果解释为nc886抑制核因子κb和激活蛋白
‑
1,因此证明了核因子κb和激活蛋白
‑
1具有贡献度(图3a)。作为支持这一观点的实验,利用聚肌胞苷酸诱导核因子κb靶信使核糖核酸的表达,并观察到上述核因子κb靶信使核糖核酸被nc886表达减弱(图3b)。利用示出核因子κb的活性的荧光素酶使293t:载体(对照组细胞株)及293t:nc886(表达nc886的细胞)感染呼吸道合胞病毒的结果,确认了293t:nc886(表达nc886的细胞)的核因子κb的活性
减弱(图3c)。使用具有4个干扰素调节因子3/干扰素调节因子7
‑
结合基序的报告质粒测定荧光素酶,由此确认nc886是否抑制干扰素调节因子3元件。上述质粒被聚肌胞苷酸活化,上述活化被nc886减弱(图3d)。通过干扰素调节因子3(5d)使上述启动子活化的结果,确认了作为活化状态的突变体的干扰素调节因子3(5d)不受nc886的影响,在293t:载体与293t:886之间等同地诱导。测定作为干扰素调节因子3的活性形态的磷酸
‑
干扰素调节因子3的结果确认了,在表达nc886的情况下,其活性减弱,在基因敲除nc886的情况下,其活性增加(图3e及3f)。因此,证实了干扰素调节因子途径也被nc886抑制。综上所述,证实了核因子κb、激活蛋白
‑
1、干扰素调节因子三种因子均被nc886调节,它们以除蛋白激酶之外还被各种途径调节而周知。
[0042][0043]
在本发明中,上述病毒为属于腺病毒科(adenoviridae)、黄病毒科(flaviviridae)、肺泡病毒科(pneumoviridae)或副粘病毒科(paramyxoviridae)的病毒,优选为腺病毒(adeno virus)、仙台病毒(sendai virus)、呼吸道合胞病毒(respiratory syncytial virus)或丙型肝炎病毒(hcv,hepatitis c virus)。
[0044][0045]
本发明可提供包含nc886或其活化剂的用于增强溶瘤病毒治疗剂的活性的组合物。
[0046][0047]
研究了腺病毒增殖中的蛋白激酶及nc886的作用。感染野生型腺病毒并8小时后获取细胞,分离核糖核酸后,从腺病毒转录的核糖核酸中,测定作为早期基因(early gene)的e1b和作为中期基因的va i,将在细胞中表达的18s核糖体核糖核酸作为内参对照(loading control)一同测定。与野生型(泳道(lane)5
‑
6)细胞相比,观察到通过crispr
‑
cas方法使蛋白激酶缺失的(泳道1
‑
2)细胞的e1b和va i的表达增加,确认了蛋白激酶为抗病毒蛋白。为了检验nc886是否通过抑制蛋白激酶来促进腺病毒增殖,在蛋白激酶缺失的状态下,删除nc886并观察,若上述假设为真,则在蛋白激酶缺失的细胞中,nc886无需抑制蛋白激酶,因此,经预测即使nc886缺失也没有影响。但是,在nc886也缺失的情况下,确认了e1b和va i的表达显著减少(图4)。这为证实nc886是腺病毒增殖所必需的结果,为示出其基质与蛋白激酶抑制活性分开的结果。
[0048][0049]
腺病毒为在细胞中增殖后向外释放时杀死细胞的溶瘤病毒之一。为了确认nc886和作为溶瘤病毒的腺病毒的关系,在使作为不表达nc886的对照组细胞株的293t.u6和作为表达nc886的细胞株的293t.u6:nc886感染野生型腺病毒的结果,确认了表达nc886的细胞中的细胞存活率更低(图5)。由此,确认在nc886存在下发生更多的溶瘤(oncolysis)作用,可利用其来提供用于增强溶瘤病毒的活性的组合物。
[0050][0051]
在本发明中,上述溶瘤病毒可以为腺病毒、呼肠孤病毒(reovirus)或新城鸡瘟病毒(newcastle disease virus)。溶瘤病毒在癌细胞中良好地增殖并破坏细胞。但是,在一部分细胞中,nc886是沉默的,根据在本发明中获取的信息,在这种癌细胞中病毒增殖减少,因此,溶瘤病毒效果不佳。在此情况下确认了,若与病毒一同给药nc886,则可提高溶瘤效
率。
[0052][0053]
并且,本发明可提供用于预防或治疗癌症的药学组合物,其包含:nc886或其活化剂;以及病毒治疗剂。在本发明中,上述病毒治疗剂可以为溶瘤病毒,但并不限定于此。明确的是,这种所有病毒及其用途属于本发明的发明要求保护范围。上述溶瘤病毒可以为腺病毒、呼肠孤病毒或新城鸡瘟病毒。上述癌症可以为乳腺癌、皮肤癌、骨癌、前列腺癌、肝癌、肺癌、非小细胞肺癌、脑癌、喉癌、胆囊癌、胰腺癌、直肠癌、甲状旁腺癌、甲状腺癌、肾上腺癌、神经组织癌、头颈癌、结肠癌、胃癌或支气管癌。
[0054][0055]
本发明可提供溶瘤病毒治疗剂中的溶瘤病毒治疗辅助剂候选物质的筛选方法,其包括:步骤(a),将表达nc886的细胞与试验物质及抗癌病毒相接触;步骤(b),在上述细胞中分析上述nc886的表达程度;以及步骤(c),将相对于未处理上述试验物质的对照组使上述nc886的表达增加的试验物质判断为抗癌病毒辅助剂候选物质。
[0059][0060]
上述溶瘤病毒可以为腺病毒、呼肠孤病毒或新城鸡瘟病毒。
[0061][0062]
本发明可提供包含nc886的表达抑制剂的用于预防或治疗病毒的药学组合物。例如,上述病毒可以为属于腺病毒科、黄病毒科、肺泡病毒科或副粘病毒科的病毒,上述病毒可以为仙台病毒、呼吸道合胞病毒或丙型肝炎病毒。nc886在双链脱氧核糖核酸病毒、单正链核糖核酸病毒及单负链核糖核酸病毒均促进病毒增殖。上述nc886的表达抑制剂可以为小干扰核糖核酸、短发夹核糖核酸(shrna)、微小核糖核酸(mirna)、核酶(ribozyme)、脱氧核酶(dnazyme)、肽核酸(peptide nucleic acids)或反义寡核苷酸。上述nc886的表达抑制剂可包含对于由seq id no:1的碱基序列组成的nc886的反义序列。上述seq id no:1的碱基序列可以为5
‘
ucgaaccccagca cagagau
‑3’
。在上述5
‘
ucgaaccccagcacagagau
‑3’
中,各个末端5nt可以为被2
′‑
o
‑
甲氧基取代的核苷酸。即,5
‘
ucgaacccca gcacagagau
‑3’
可以为核糖核苷酸(ribonucleotide),5
‘
ucgaacc ccagcacagagau
‑3’
可以为脱氧
‑
核糖核苷酸(deoxy
‑
ribonucleotid e)。碱基与碱基之间被修饰为硫代磷酸骨架(phosphorothioate backb one)。
[0063][0064]
本发明可提供用于治疗病毒感染的候选物质的筛选方法,其包括:步骤(a),将表达nc886的细胞与试验物质相接触;步骤(b),在上述细胞中分析上述nc886的表达程度;以及步骤(c),当上述试验物质减少上述微小核糖核酸的表达时,判断为用于治疗病毒感染的候选物质。
[0068][0069]
在本发明中,上述病毒可以为属于黄病毒科、肺泡病毒科或副粘病毒科的病毒,上述病毒可以为仙台病毒、呼吸道合胞病毒或丙型肝炎病毒。
[0070][0071]
以下,为了帮助理解本发明,例举实施例来详细说明。但是,下述实施例仅例示本发明的内容,本发明的范围并不限定于下述实施例。本发明的实施例为了向本技术领域的普通技术人员更完整地说明本发明而提供。
[0072][0073]
实施例1.细胞株、病毒及抗体
[0074][0075]
hep
‑
2细胞购自美国模式培养物集存库(atcc),以如文献[bao,x.et al.,virology.,408,224
‑
231(2010)]所说明的方式保持。仙台病毒购自harles river laboratories,inc。在37℃的温度下,在hep
‑
2细胞中使rst长株(long strain)增殖,如文献[ren,j.et al.,j.gen.virol.,92,2153
‑
2159(2011)]中所记载的方法,通过蔗糖梯度(sucros e gradient)纯化。通过crispr
‑
cas技术利用nthy
‑
ori
‑3‑
1制备了删除蛋白激酶和nc886的表达的细胞株(oncotarget.2016nov 15;7(46):75000
‑
75012.doi:10.18632/oncotarget.11852.lee ek et al.),在包含10%的胎牛血清(fbs)和1%的青霉素/链霉素(penicillin/strept omycin)抗生素的无血清细胞冻存(rpmi)培养基保持。如上所述,依次使用多克隆生物素结合山羊抗
‑
呼吸道合胞病毒抗体(cat#:7950
‑
0104,bio
‑
rad)和链霉亲和素过氧化物酶聚合物(streptavidin peroxid ase polymer;cat#:s2438,sigma
‑
aldrich),在hep
‑
2细胞通过免疫染色确定了病毒滴度(viral titer)。对于β
‑
肌动蛋白(β
‑
actin)的单克隆抗体(cat#a1978)购自sigma。利用包含10%的胎牛血清及1%的抗霉素(antibiotic
‑
antimycotic)的dmem培养基,在5%的co2及37℃的条件下培养hek
‑
293t细胞(美国模式培养物集存库(amer ican type culture collection))。通过向上述hek
‑
293t细胞插入plpcx载体中的u6启动子来制备了质粒“plpcx
‑
u6”及nc886表达质粒“plpcx
‑
u6
‑
pre
‑
886”。293t细胞为nc886表达沉默的细胞株,在该细胞株中将nc886人工表达的细胞株为293t.u6:nc886,当制备该细胞株时,利用空白载体(empty vector)制备的对照组细胞株为293t.u6(rna.2011jun;17(6):1076
‑
89.doi:10.1261/rna.2701111.epub 2011apr 25.pmid:21518807.lee k.et al.)。腺病毒为5型野生型病毒,由西班牙的dr.alemany实验室提供,在hek
‑
293细胞中增殖后,通过标准氯化铯方法(https://bio
‑
protocol.org/bio101/e201)纯化。在hek
‑
293及hek
‑
293ft细胞感染后,通过结晶紫(crystal violet)染色方法确定病毒滴度。
[0076][0077]
实施例2.报告基因分析法(reporter gene assay)
[0078][0079]
如文献[jeon sh et al.,febs lett.,21;586(19):3477
‑
842012sep]中所述,将包含干扰素
‑
β启动子的质粒、包含与荧光素酶报告基因相连的干扰素
‑
β启动子的干扰素刺激反应元件位点的多个拷贝的质粒转染至细胞内。如文献[lee k et al.rna.,17(6):1076
‑
89.,2011jun]所记述,使用fugene 6(roche,indianapolis,in)来将对数生长的293细胞3倍转染为报告质粒。转导15小时后,利用聚肌胞苷酸处理细胞,或者利用仙台病毒或呼吸道合胞病毒分别感染2小时及15小时。荧光素酶对于内部对照组rr活性(internal control rr activity)正常化。
[0080][0081]
实施例3.丙型肝炎病毒实验
[0082][0083]
使huh7.5细胞在补充有10%的胎牛血清、1%的mem非必须氨基酸溶液、100u/ml的青霉素及100mg/ml的链霉素(gibco)的dmem培养基中生长。将huh7.5细胞以3
×
105/孔接种
在6孔板并培养一夜。利用丙型肝炎病毒jfh1(104ffu/ml)上清液感染huh7.5细胞,在4小时内利用磷酸盐缓冲液(pbs)清洗2次后,为了去除丙型肝炎病毒jfh1而替换新的培养基。以15ul的rnai
‑
max向细胞转染100nm的抗
‑
nc886或抗
‑
穹窿体核糖核酸。转染12小时、24小时及48小时后,利用包含蛋白酶/磷酸酶抑制剂(thermofisher)的ripa裂解液(thermofisher)裂解细胞,在13000rpm的条件下,离心分离15分钟来收集了包含蛋白质的上清液。使用bca蛋白质分析试剂盒(23225,thermofisher)来测定细胞提取物的蛋白浓度。对于各个样品,在10%的十二烷基硫酸钠
‑
聚丙烯酰胺凝胶电泳凝胶对25μg的总蛋白进行电穿孔(electroporated)后,转移到硝酸纤维素膜。使用5%的脱脂乳(skin milk)将膜(membrane)封闭(blocking)1小时,在4℃的温度下封闭抗
‑
丙型肝炎病毒核心(ma1
‑
080,thermo scientific)、抗
‑
ns3(ab65407,abcam)、查尔斯
·
赖斯的抗
‑
ns5a、抗
‑
蛋白激酶(ab32052,abcam)、抗
‑
磷酸
‑
蛋白激酶(ab32036)及β
‑
肌动蛋白(a2228,sigma)的一抗。除使用作为其他病毒克隆的jc1(107ffu/ml)之外,通过如上所述的方法执行。利用抗
‑
886寡核苷酸转染,在12小时、24小时、48小时及72小时后,根据制造商的说明,通过使用qi amp病毒核糖核酸迷你试剂盒(qiamp viral rna mini kit,52904,qiagen)来分离丙型肝炎病毒核糖核酸。实时荧光定量聚合酶链式反应(qrt
‑
pcr)实验使用iq5多色实时聚合酶链式反应检测系统(iq5multicolor real
‑
time pcr detection system;bio
‑
rad laboratories)来执行。正向引物:5'
‑
tgcacggtctacgagac
‑
3',探针:fam
‑
5'
‑
ccggggcactcgcaagcaccc
‑
3'
‑
gmbh,反向引物:5'
‑
gaccccccctcccgggagag
‑
3'(靶尺寸(target siz):222bp)。
[0084][0085]
实施例4.rantes酶联免疫吸附试验(elisa)
[0086]
根据制造商的说明(r&d system),通过商业酶联免疫吸附法(elisa)确定antes浓度。
[0087][0088]
实施例5.聚合酶链式反应
[0089][0090]
为了确认nc886促进腺病毒的增殖,通过逆转录聚合酶链式反应(rt
‑
pcr)确认细胞的转录阶段的代表基因的表达。以moi(感染复数(multiplicity of infection))=5感染腺病毒,8小时后,通过trizol(15596018,invitrogen)方法提取总核糖核酸,通过使用amfiri vert cdna synthesis platinum master mix(r5600,gendepot)来制备互补脱氧核糖核酸(cdna)。以下述表1中公开的引物碱基序列且利用amfisure pcr master mix(2x)(p0311,gendepot),对所制备的互补脱氧核糖核酸和作为病毒基因的e1b 55k、vai rna和用作对照组的人的基因18s核糖体核糖核酸进行聚合酶链式反应(polymeras e chain reaction),并通过琼脂糖胶(agarose gel)进行电泳来确认。
[0091][0091]
表1
[0093][0094][0095][0096][0097]
实施例6.测定细胞的存活力
[0098][0099]
将腺病毒以moi=5感染图中所示的细胞株,42小时后测定细胞的存活。利用celltiterone solution cell proliferation assay(mts)(g3581,promega)分析了存活力。
[0100][0101]
实施例7.nc886促进病毒复制
[0102][0103]
丙型肝炎病毒复制在一部分肝细胞株中进行。本发明人发现,在可复制丙型肝炎病毒的huh7类的肝细胞株(huh7、huh7.5、ft3
‑
7)的情况下,nc886的表达水平在原代肝细胞中高,相反,在未复制丙型肝炎病毒的肝细胞株的hepg2及hep3b中是沉默的(图1a)。由此,为了确认在丙型肝炎病毒复制中的nc886的作用,在利用反义寡核苷酸在huh7.5细胞株中基因敲除nc886的结果,蛋白激酶在基因敲除nc886的细胞中活化。并观察到丙型肝炎病毒的信使核糖核酸及蛋白量也减少,从上述结果确认了nc886对于复制丙型肝炎病毒是必需的(图1b及图1d)。为了确认nc886是否对于其他类型的病毒也有效,利用仙台病毒、呼吸道合胞病毒、腺病毒进行追加试验。在使表达nc886的293t细胞感染呼吸道合胞病毒及仙台病毒的结果,确认了病毒的增殖增加(图1e~图1f)。进而,确认了在利用crispr
‑
cas技术删除nc886的nthy
‑
ori 3
‑
1细胞中,腺病毒的增殖显著减少(图1g)。
[0104][0105]
实施例8.nc886拮抗干扰素反应
[0106][0107]
丙型肝炎病毒为单正链核糖核酸病毒,仙台病毒和呼吸道合胞病毒为单负链核糖核酸病毒,腺病毒为双链脱氧核糖核酸病毒,因此,病毒基因表达及复制机制均不同。观察到各种病毒的增殖均被nc886促进,因此,nc886促进多种病毒的复制的可能性高,由此假设宿主的先天免疫反应抑制是促nc886的原病毒的作用的机制。为了确认其,研究了nc886对于主要宿主防御重要的干扰素
‑
β信号的效果。干扰素
‑
β包含对于3种激活蛋白
‑
1、干扰素调节因子及核因子κb的结合位点(图2a)。聚肌胞苷酸(促进干扰素生产的合成核糖核酸)的转染诱导干扰素
‑
β的信使核糖核酸表达,相反,当nc886存在时,干扰素
‑
β的信使核糖核酸表
达减少(图2b)。干扰素
‑
β启动子在作为不表达nc886的对照组细胞株的293t中通过聚肌胞苷酸、仙台病毒、呼吸道合胞病毒的感染活化,在制备成表达nc886的293t细胞中活化程度减少(图2c)。在干扰素
‑
β信号途径中,干扰素刺激基因的转录活化是被干扰素刺激反应元件介导的。利用干扰素刺激反应元件测定荧光素酶表达的结果,干扰素刺激反应元件荧光素酶示出与干扰素
‑
β荧光素酶相似的结果(图2d)。由此确认了nc886抑制干扰素
‑
β的信号。除干扰素
‑
β之外,当生产如rantes的趋化因子时,需要作为上述干扰素
‑
β启动子的3个调节因素的激活蛋白
‑
1、干扰素调节因子及核因子κb,经确认,当感染呼吸道合胞病毒时,nc886抑制rantes的诱导(图2e)。
[0108][0109]
实施例9.nc886通过抑制蛋白激酶来抑制干扰素
‑
β启动子
[0110][0111]
评价了作为干扰素
‑
β启动子的结合的三种基因的核因子κb、激活蛋白
‑
1及干扰素调节因子(图2a)的贡献度。转染聚肌胞苷酸的结果,使干扰素
‑
β启动子活化,上述活化被nc886显著减弱。与之相比,在转染结构上活化状态的磷酸
‑
模拟干扰素调节因子3[=干扰素调节因子3(5d)]突变体的情况下,也使干扰素
‑
β启动子活化,上述活化也被nc886减弱。由于引入活化状态的干扰素调节因子3,在此条件下被nc886减弱的结果解释为nc886抑制核因子κb和激活蛋白
‑
1,因此证明了核因子κb和激活蛋白
‑
1具有贡献度(图3a)。作为支持这一观点的实验,利用聚肌胞苷酸诱导核因子κb靶信使核糖核酸的表达,并观察到上述核因子κb靶信使核糖核酸被nc886表达减弱(图3b)。利用示出核因子κb的活性的荧光素酶使293t:载体(对照组细胞株)及293t:nc886(表达nc886的细胞)感染呼吸道合胞病毒的结果,确认了293t:nc886(表达nc886的细胞)的核因子κb的活性减弱(图3c)。使用具有4个干扰素调节因子3/干扰素调节因子7
‑
结合基序的报告质粒测定荧光素酶,由此确认nc886是否抑制干扰素调节因子3元件。上述质粒被聚肌胞苷酸活化,上述活化被nc886减弱(图3d)。通过干扰素调节因子3(5d)使上述启动子活化的结果,确认了作为活化状态的突变体的干扰素调节因子3(5d)不受nc886的影响,在293t:载体与293t:886之间等同地诱导。测定作为干扰素调节因子3的活性形态的磷酸
‑
干扰素调节因子3的结果确认了,在表达nc886的情况下,其活性减弱,在基因敲除nc886的情况下,其活性增加(图3e及3f)。因此,证实了干扰素调节因子途径也被nc886抑制。综上所述,证实了核因子κb、激活蛋白
‑
1、干扰素调节因子三种因子均被nc886调节,它们以除蛋白激酶之外还被各种途径调节而周知。
[0112][0113]
实施例10.nc886促进腺病毒增殖
[0114][0115]
腺病毒的复制可在细胞表面表达柯萨奇病毒和腺病毒受体(coxsackievirus and adenovirus receptor(car))的几乎所有细胞感染,并可发生复制。通常,腺病毒在感染后表达病毒基因,相反,宿主细胞利用称为蛋白激酶的防御系统调节增殖而周知。在删除蛋白激酶的nthy
‑
ori
‑3‑
1细胞中感染病毒后,获取了病毒基因e1b 55k和vai的核糖核酸表达与亲本细胞相比增加的结果(比较图4的泳道6和2)。nc886/蛋白激酶均被基因敲除(ko)的细胞中,e1b和vai的表达显著减少(比较图4的泳道6和4)。上述结果证实了nc886在腺病毒的增殖中是必需的。同时,在该细胞株中,不仅基因敲除了nc886,还基因敲除了蛋白激酶,因
此,nc886在腺病毒的增殖中必需的原因在于抑制蛋白激酶之外的其他机制。
[0116][0117]
实施例11.nc886通过腺病毒促进细胞凋亡
[0118][0119]
在正常的细胞培养条件下,293t.u6:nc886的生长速度略慢于293t.u6。在感染腺病毒后,确认了nc886表达293t的存活率显著减少(图5)。腺病毒以增殖后从感染的细胞中出来时示出细胞溶解(oncolysis)而周知,因此,除了上述逆转录聚合酶链式反应结果之外,nc886表达细胞中的腺病毒基因的表达也增加,由此促进病毒增殖,从而可以得出更好地使细胞凋亡的结果。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。