人源化抗人cd89抗体及其用途
技术领域
1.本发明涉及抗体和此类抗体的用途的领域。所述抗体特别适用于防止人iga与人cd89的结合。所述抗体特别适用于靶向人效应细胞。
背景技术:
2.免疫球蛋白的fc部分的受体(fcr)存在于各种类型的免疫细胞上,并在免疫系统的体液分支与细胞分支之间提供连接。抗体与fcr之间的相互作用为表达所述fcr的细胞提供抗原特异性识别性质。相互作用可发起多种应答,其中包括内吞作用、吞噬作用、转胞吞作用、胞吐作用、超氧化物产生、抗体依赖性细胞毒性以及细胞因子炎症介质的释放。免疫球蛋白的受体及其在免疫应答中的作用被广泛研究。
3.目前,识别了五类抗体恒定区,即igg、iga、igd、ige和igm。iga在先天免疫系统中发挥作用。它参与防止微生物和外来蛋白质穿透粘膜表面。iga还能够摧毁毒素和感染性生物体。此外,iga具有抗炎性质并且能够抑制诸如igg诱导的细胞因子释放和吞噬作用的功能。
4.不同类别的抗体恒定区可与不同的fc受体相互作用。能够结合iga的fc受体是cd89。人cd89能够与人iga1和人iga2的重链恒定区结合。cd89是糖基化跨膜受体并且也被称为fcαri。cd89具有两个细胞外结构域ec1和ec2、跨膜结构域和细胞内结构域。cd89与iga的相互作用由ec1细胞外结构域介导。对于参考序列,我们参考np_001991.1(免疫球蛋白αfc受体同种型前体)。该参考仅用于鉴定人cd89基因/蛋白。这并不旨在将如本文所述的人cd89限于数据库条目的特定序列。人cd89的天然变体在本发明的范围内。如果重组人cd89能够结合iga,并且能够结合如本文所述的抗体,则它也在本发明的范围内。cd89存在于髓系细胞的细胞表面上,所述髓系细胞包括嗜中性粒细胞、嗜酸性粒细胞以及大多数单核细胞和巨噬细胞。然而,受体不出现在肥大细胞和肠道巨噬细胞上。cd89的表达被发现是组成性的,并且与iga配体的存在无关。
5.cd89可与单体iga、聚合iga和iga复合物相互作用。据认为,单体iga与cd89瞬时结合,而聚合iga和iga复合物与cd89紧密结合。cd89可在促炎和抗炎应答两者中发挥作用。为了对iga结合作出反应,受体必须与另一种因子缔合,所述因子主要是二聚体形式的fcrγ-链。配体与cd89的结合可发起多种生物过程。通过配体与cd89的结合促进的细胞功能也依赖于缔合的fcrγ链。
6.细胞上cd89受体的交联可通过iga抗体、iga免疫复合物或抗cd89抗体的结合来实现。交联触发免疫应答,所述免疫应答取决于环境并且特别是取决于结合而可能具有积极和消极影响。cd89特异性抗体可用作调节免疫应答的工具/药物。例如,患有慢性炎症性疾病的个体可从抑制免疫应答的方法中获益。cd89受体在各种细胞类型上表达,所述细胞类型包括嗜中性粒细胞、嗜酸性粒细胞、单核细胞和巨噬细胞。特别是患有与表达cd89的细胞有关的疾病和/或iga相关疾病的个体可从使用cd89抗体的治疗中获益。目前,对于与嗜中性粒细胞有关的疾病,尚无已知的治疗。
技术实现要素:
7.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的人源化抗人cd89抗体,所述人源化抗人cd89抗体包含:
8.重链可变区,所述重链可变区包含氨基酸序列:
9.evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglex1vx2t ix3gx4gdityy pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
10.其中:
11.x1是l或w
12.x2是a或s
13.x3是n、s
14.x4是q、t或n,
15.其中所述重链可变区包含在除x1、x2、x3和x4以外的位置处相对于所指示氨基酸序列的0、1、2或3个氨基酸变异、插入、缺失、取代、添加或它们的组合;
16.和轻链可变区,所述轻链可变区包含氨基酸序列:
17.diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gkz1z2klliyy tsrlhsgvps rfsgsgsgtdz
3 tltisslqp edfatyz4cqq gktlpytfgq gtkleik
18.其中:
19.z1是a或t
20.z2是v或p
21.z3是y或f
22.z4是y或f,
23.其中所述轻链可变区包含在除z1、z2、z3和z4以外的位置处相对于所指示氨基酸序列的0、1、2或3个氨基酸变异、插入、缺失、取代、添加或它们的组合。
24.在一个优选的实施方案中,x1是l,x2是a,x3是s,x4是t,z1是a,z2是v,z3是y,并且z4是y。
25.在另一个优选的实施方案中,x1是l,x2是a,x3是s,x4是q,z1是a,z2是v,z3是y,并且z4是y。
26.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:138或139的氨基酸序列的重链,和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链。
27.如本文所述的抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。
28.在另一方面,如本文所述的抗体能够结合至表达人cd89的hek293f细胞上的人cd89的细胞外部分。表达人cd89的hek293f细胞的布达佩斯条约保藏由保藏人swiss pharma international ag,waldmannstr.8,8001zurich,switzerland以保藏人参考编号293f cg89克隆2保藏于德国布伦瑞克的莱布尼茨学院dsmz-德意志微生物保藏中心(leibniz-institut dsmz-deutsche sammlung von mikroorganismen und zellkulturen gmbh(dsmz)),指定登录号:dsm acc3341。
29.在一个方面,本公开提供了编码如本文公开的抗体或如本文公开的其抗原结合片段的一种或多种核酸分子。还提供了一种编码如本文公开的可变区,优选如本文所述的人源化抗人cd89抗体的重链可变区和/或轻链可变区的核酸。
30.在一个方面,本公开提供了一种载体,所述载体包含如本文所述的核酸分子。在一个方面,本公开提供了一种细胞(宿主细胞),所述细胞包含如本文公开的抗体、一种或多种核酸分子和/或载体。优选地,所述宿主细胞是哺乳动物、昆虫、植物、细菌或酵母细胞。更优选地,所述宿主细胞是人细胞。优选地,所述宿主细胞是杂交瘤细胞、中国仓鼠卵巢(cho)细胞、nso细胞或per-c6
tm
细胞(欧洲细胞培养物保藏中心(european collection of cell cultures,ecacc)96022940;wo1997000326;pau等人,2001.vaccine 19:2716-2721)。
31.在一个方面,本公开提供了一种产生如本文公开的抗体的方法。所述方法优选地包括收获所述抗体。优选地,所述抗体使用细胞产生,并且从所述细胞收获。优选地,所述细胞是杂交瘤细胞、中国仓鼠卵巢(cho)细胞、nso细胞或per-c6
tm
细胞。在另一个优选的实施方案中,所述抗体合成产生。
32.本公开的一个方面提供了一种药物组合物,所述药物组合物包含如所公开的抗体或其抗原结合片段、核酸和/或细胞。优选地,如本文公开的组合物或抗体或其抗原结合片段用于制造药物。优选地,所述药物用于治疗或预防慢性炎症性疾病。
33.在一个方面,本公开提供了一种用于治疗受试者中的慢性炎症性疾病的方法,所述方法包括向有需要的受试者施用治疗有效量的如本文公开的抗体或其抗原结合片段、核酸分子或载体或药物组合物。
34.在一个方面,本公开提供了一种用于治疗慢性炎症性疾病的抗体或其抗原结合片段。
35.在一个方面,本公开提供了一种抗体,所述抗体能够结合或结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止或阻止人iga与人cd89的结合,并且不会使所述细胞在37℃下过夜孵育后的细胞活力降低超过60%。如本文提及的在37℃下过夜孵育后的活力或表达的变化优选地是指相对于优选地是在相同条件下但没有所述抗体的情况下孵育时的对照值的变化。所述活力实验优选地在合适的培养基,如无血清培养基中进行。
36.在另一方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且当与在相同条件下但没有所述抗体的情况下孵育的相同细胞相比时,不会使所述细胞在37℃下过夜孵育后的磷脂酰丝氨酸表达增加超过20%。
37.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且当在37℃下、在nan3不存在下所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且当在4℃下、在nan3存在下与所述细胞结合时不能置换单体人iga或热聚iga。
38.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与重组人cd89分子结合,其中人cd89的氨基酸22-46被替换为食蟹猴cd89的氨基酸22-46。
39.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸47-71被替换为食蟹猴cd89的氨基酸47-71。
40.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸72-96被替换为食蟹猴cd89的氨基酸72-96。
41.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且其与嵌合cd89分子的结合未减少20%或更少,其中人cd89的氨基酸97-121被替换为食蟹猴cd89的氨基酸97-121。
42.在一个方面,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸58、59、73、74、76、106和107被替换为食蟹猴cd89的相应氨基酸58、59、73、74、76、106和107。
43.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:29-31的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:32-34的cdr1、cdr2和cdr3序列的轻链可变区。
44.在另一方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:27的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:28的氨基酸序列的轻链可变区。
45.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:45-47的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:48-50的cdr1、cdr2和cdr3序列的轻链可变区。
46.在另一方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:43的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:44的氨基酸序列的轻链可变区。
47.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:69-71的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:72-74的cdr1、cdr2和cdr3序列的轻链可变区。
48.在另一方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:67的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代
或添加的seq id no:68的氨基酸序列的轻链可变区。
49.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:77-79的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:80-82的cdr1、cdr2和cdr3序列的轻链可变区。
50.在另一方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:75的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:76的氨基酸序列的轻链可变区。
51.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:53-55的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:56-58的cdr1、cdr2和cdr3序列的轻链可变区。
52.在另一方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:51的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:52的氨基酸序列的轻链可变区。
53.在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的抗体,优选地所述抗体能够结合cd89的ec1细胞外结构域。
附图说明
54.图1.使用流式细胞术,小鼠抗人cd89抗体(上清液)与hek293f细胞上的膜结合全长人cd89的结合。虚线代表背景(即,无小鼠抗人cd89抗体的结合)。示出平均值
±
sd(n=2)。
55.图2.小鼠抗人cd89抗体(上清液)对血清人iga与rhucd89(a,elisa)或与hek293f细胞上的膜结合人cd89(b,facs)的结合的影响。示出平均值
±
sd(n=2)。
56.图3.纯化的cd89/iga阻断小鼠抗人cd89抗体与rhucd89(a,elisa)或与hek293f细胞上的膜结合人cd89(b,facs)的结合特性。示出平均值
±
sd(n=2)。
57.图4.纯化的cd89/iga阻断小鼠抗人cd89抗体对血清人iga与rhucd89(a,elisa)或与hek293f细胞上的膜结合人cd89(b,facs)的结合的影响。纯化的cd89/iga阻断小鼠抗人cd89抗体对分泌型人iga与hek293f细胞上的膜结合人cd89(c)的结合的影响。示出平均值
±
sd(n=2)。
58.图5.纯化的cd89/iga阻断小鼠抗人cd89抗体(10μg/ml)对(a,c)预防非聚集或热聚血清人iga与膜结合人cd89的结合、对(b,d)先前饱和的非聚集或热聚血清人iga置换为膜结合人cd89和对(e)诱导表达膜结合人cd89的hek293f细胞的细胞死亡(细胞活力和磷脂酰丝氨酸表达)的影响。(a,b)在代谢非活性条件下(即,在nan3存在下和低温环境温度(4℃)下)和(c,d,e)在代谢活性条件下(即,在nan3不存在下和生理环境温度(37℃)下)检测cd89/iga阻断小鼠抗人cd89抗体的影响。示出平均值
±
sd(n=2)。
59.图6.原型商业小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3对预防/抑制非聚
集(a)或热聚(b)血清人iga与hek293f细胞上的膜结合人cd89的结合的影响。示出平均值
±
sd(n=2)。
60.图7.纯化的cd89/iga阻断小鼠抗人cd89抗体(10μg/ml)与离体人嗜中性粒细胞上(a;来自5个不同健康供体的平均值
±
sd)、hek293f细胞上(b;平均值
±
sd(n=2))和单核细胞u937细胞上(c;平均值
±
sd(n=2))的膜结合人cd89的结合。
61.图8.纯化的cd89/iga阻断小鼠抗人cd89抗体对表达人cd89的离体原代人嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的影响。虚线表示仅iga包被的珠(即,不添加抗体)的吞噬作用。示出来自3个不同健康供体(1、2和3)的平均值
±
sd(n=2)。
62.图9.纯化的cd89/iga阻断小鼠抗人cd89抗体(20μg/ml)对(a;平均值
±
sd(n=3))血清人iga包被的琼脂糖珠诱导的表达人cd89的离体原代人嗜中性粒细胞的2维迁移和对(b;平均值
±
sd(n=3))来自表达人cd89的离体原代人嗜中性粒细胞的其相应上清液来源的趋化活性以及对(c;n=1)来自表达人cd89的离体原代人嗜中性粒细胞的其相应上清液来源的化学吸引ltb4水平的影响。虚线表示由仅iga包被的琼脂糖珠(即,不添加抗体)诱导的(a)2维粒细胞迁移,(b)粒细胞趋化和(c)粒细胞ltb4产生。在(a)、(b)和(c)中,示出来自3个健康供体(1、2和3)的数据。nd=未测定。
63.图10.纯化的cd89/iga阻断小鼠抗人cd89抗体对(a)血清人iga与表达人cd89的离体原代人嗜中性粒细胞的结合和对(b)来自所述细胞的其相应血清人iga介导的乳铁蛋白产生(脱粒标志物)的影响。虚线表示(a)粒细胞与仅iga包被的板的结合和(b)由仅iga包被的板(即,不添加抗体)诱导的其相应的乳铁蛋白产生。在(a)、(b)中示出来自3个健康供体(1、2和3)的平均值
±
sd(n=2)。
64.图11.非标记的纯化cd89/iga阻断小鼠抗人cd89抗体(10μg/ml)与pe缀合的商业小鼠抗cd89抗体克隆mip8a(a;cd89/iga阻断剂)、克隆a59(b;cd89/iga非阻断剂)和克隆a3(c;cd89/iga非阻断剂)对hek293f细胞上的膜结合人cd89的交叉竞争。示出平均值
±
sd(至少n=2)。
65.图12.对非标记的纯化cd89/iga阻断小鼠抗人cd89抗体与pe缀合的商业小鼠抗cd89抗体克隆mip8a(a;cd89/iga阻断剂)、克隆a59(b;cd89/iga非阻断剂)和克隆a3(c;cd89/iga非阻断剂)对hek293f细胞上的膜结合人cd89的交叉竞争的详细分析。示出平均值
±
sd(至少n=2)。
66.图13.(a)野生型人cd89、野生型牛fcγ2r及其来源的嵌合人/牛fcr构建体的示意性图示。(b)兔抗人cd89多克隆抗体和非聚集或热聚血清人iga与hek293f细胞上的膜结合人全长cd89(
‘
人ec1-ec2-cd89’;即)、膜结合嵌合人ec1-cd89/牛ec2-fcγ2r(
‘
人ec1-cd89’,即和)、膜结合嵌合牛ec1-fcγ2r/人ec2-cd89(
‘
人ec2-cd89’;即)以及膜结合牛全长fcγ2r(
‘
牛fcγ2r’,即)的结合(n=1)。(c)纯化的cd89/iga阻断小鼠抗人cd89抗体与hek293f细胞上的膜结合人全长cd89(
‘
人ec1-ec2-cd89’)、膜结合嵌合人ec1-cd89/牛ec2-fcγ2r(
‘
人ec1-cd89’)、膜结合嵌合牛ec1-fcγ2r/人ec2-cd89(
‘
人ec2-cd89’)以及膜结合牛全长fcγ2r(
‘
牛fcγ2r’)的结合(n=1)。灰色虚线代表背景(即,无小鼠抗人cd89抗体的结合)。
67.图14.(a)兔抗人cd89多克隆抗体和非聚集或热聚血清人iga与膜结合食蟹猴全长
cd89的结合。(b)纯化的cd89/iga阻断小鼠抗人cd89抗体与膜结合食蟹猴全长cd89的结合。灰色虚线代表背景(即,无小鼠抗人cd89抗体的结合)。
68.图15.(a)野生型人cd89、野生型食蟹猴cd89及其来源的嵌合人/食蟹猴cd89构建体的示意性图示。(b)兔抗人cd89多克隆抗体和非聚集或热聚血清人iga与hek293f细胞上的膜结合人全长cd89(
‘
人ec1-cd89’;即)、膜结合嵌合人/食蟹猴cd89-热点(
‘
δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89’,即)、膜结合嵌合人/食蟹猴cd89-i(
‘
δgln22-lys46人ec1-cd89’,即)、膜结合嵌合人/食蟹猴cd89-ii(
‘
δile47-ile71人ec1-cd89’,即)、膜结合嵌合人/食蟹猴cd89-iii(
‘
δgly72-gly96人ec1-cd89’,即)、膜结合嵌合人/食蟹猴cd89-iv(
‘
δarg97-gly121人ec1-cd89’,即)和膜结合嵌合食蟹猴全长cd89(
‘
食蟹猴ec1-cd89’,即)的结合(n=1)。(c)纯化的cd89/iga阻断小鼠抗人cd89抗体与hek293f细胞上的膜结合人全长cd89(
‘
人ec1-cd89’)、膜结合嵌合人/食蟹猴cd89-热点(
‘
δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89’)、膜结合嵌合人/食蟹猴cd89-i(
‘
δgln22-lys46人ec1-cd89’)、膜结合嵌合人/食蟹猴cd89-ii(
‘
δile47-ile71人ec1-cd89’)、膜结合嵌合人/食蟹猴cd89-iii(
‘
δgly72-gly96人ec1-cd89’)、膜结合嵌合人/食蟹猴cd89-iv(
‘
δarg97-gly121人ec1-cd89’)和膜结合嵌合食蟹猴全长cd89(
‘
食蟹猴ec1-cd89’)的结合(n=1)。灰色虚线代表背景(即,无小鼠抗人cd89抗体的结合)。
69.图16.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体与rhucd89(a,elisa)或hek293f细胞上的膜结合人cd89(b,facs)的结合特性。示出平均值
±
sd(n=2)。
70.图17.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体对非聚集(a)或热聚(b)血清人iga与hek293f细胞上的膜结合人cd89的结合的影响。纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体对分泌型人iga与hek293f细胞上的膜结合人cd89(c)的结合的影响。示出了平均值
±
sd(n=2)。
71.图18.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体(10μg/ml)与离体人嗜中性粒细胞上的膜结合人cd89的结合(来自3个不同健康供体的平均值
±
sd)。
72.图19.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体对表达人cd89的离体原代人嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的影响。虚线表示仅iga包被的珠(即,不添加抗体)的吞噬作用。示出来自3个不同健康供体(1、2和3)的平均值
±
sd(n=2)。
73.图20.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体(20μg/ml)对(a;平均值
±
sd(n=3))血清人iga包被的琼脂糖珠诱导的表达人cd89的离体原代人嗜中性粒细胞的2维迁移和对(b;平均值
±
sd(n=3))来自表达人cd89的离体原代人嗜中性粒细胞的其相应上清液来源的趋化活性以及对(c;平均值
±
sd(n=2))来自表达人cd89的离体原代人嗜中性粒细胞的其相应上清液来源的化学吸引ltb4水平的影响。虚线表示由仅iga包被的琼脂糖珠(即,不添加抗体)诱导的(a)2维粒细胞迁移,(b)粒细胞趋化和(c)粒细胞ltb4产生。示出来自2(b)至3((a)和(c))个健康供体(1、2和3)的数据。
74.图21.纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体对血清人iga介导的来自表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生(脱粒标志物)的影响。虚线表示由仅
iga包被的板(即,不添加抗体)诱导的乳铁蛋白产生。示出来自2个健康供体(1和2)的平均值
±
sd(n=2)。
75.图22:表达人cd89的hek293f细胞的布达佩斯条约保藏以登录号:dsm acc3341和鉴定参考编号:293f cd89克隆2保藏。保藏形式:(a)在原始保藏物情况下的接收(b)存活力声明。
76.图23:纯化的cd89/iga阻断小鼠抗人cd89抗体对血清人iga包被的乳胶珠诱导的表达人cd89的离体原代人嗜中性粒细胞的net释放的影响。虚线表示由仅iga包被的珠(即,不添加抗体)诱导的net释放。示出来自8个不同健康供体(1-8)的平均值
±
sd(n=3)。
77.图24:纯化的cd89/iga阻断小鼠抗人cd89抗体对诱导表达非引发和lps引发的膜结合人cd89的离体原代人嗜中性粒细胞的细胞死亡(碘化丙啶(pi)百分比)的影响。示出来自3个健康供体(1、2和3)的平均值
±
sd(n=1或2)。
78.图25:cd89/iga阻断人源化抗人cd89抗体10e7(即,源自与人恒定igg4/κ区组合的cd89/iga阻断小鼠抗人抗体10e7 vh和vl区的人源化vh1、2、3和vl1、2、3、4型式)(的上清液)与rhucd89(a,elisa)和hek293f细胞上的膜结合人cd89(b,facs)的特有的结合。cd89/iga阻断嵌合抗人cd89抗体10e7(即,嵌合物igg4/κ)用作参考。示出了平均值
±
sd(n=2)。
79.图26:纯化的cd89/iga阻断
‘
cdr2脱酰胺修复的’(即,vh3sqvl3 10e7和vh3stvl3 10e7)人源化抗人cd89抗体10e7型式vh3sqvl3和vh3stvl3(即,源自与人恒定igg4κ区组合的cd89/iga阻断小鼠抗人抗体10e7 vh和vl区的人源化vh3和vl3型式)与rhucd89(a,elisa)和hek293f细胞上的膜结合人cd89(b,facs)的特有的结合。cd89/iga阻断嵌合抗人cd89抗体10e7(即,嵌合物igg4/κ10e7)用作参考。示出平均值
±
sd(n=2)。
80.图27:纯化的cd89/iga阻断
‘
cdr2脱酰胺修复的’(即,vh3sqvl3 10e7和vh3stvl3 10e7)人源化抗人cd89抗体10e7型式vh3sqvl3和vh3stvl3(即,源自与人恒定igg4κ区组合的cd89/iga阻断小鼠抗人抗体10e7 vh和vl区的人源化vh3和vl3型式)对血清人iga(a)和分泌型人iga(b)与hek293f细胞上的膜结合人cd89的结合的影响。cd89/iga阻断嵌合抗人cd89抗体10e7(即,嵌合物igg4/κ10e7)用作参考。示出平均值
±
sd(n=2)。
81.图28:纯化的cd89/iga阻断
‘
cdr2脱酰胺修复的’人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3(10μg/ml)与离体人嗜中性粒细胞上的膜结合人cd89的结合(来自3个不同健康供体的平均值
±
sd)。
82.图29:纯化的cd89/iga阻断
‘
cdr2脱酰胺修复的’人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对表达人cd89的离体原代嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的影响。虚线表示仅iga包被的珠(即,不添加抗体)的吞噬作用。示出来自3个不同健康供体(1、2和3)的平均值
±
sd(n=2)。
83.图30:纯化的cd89/iga阻断
‘
cdr2脱酰胺修复’人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对人血清iga介导的来自表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生(脱粒标志物)的影响。虚线表示由仅iga包被的板(即,不添加抗体)诱导的乳铁蛋白产生。示出来自5个健康供体(1-5)的平均值
±
sd(n=2)。
84.图31:(a)体内labd小鼠模型的实验设置和使用cd89/iga阻断小鼠抗人cd89抗体10e7的治疗方案。(b)纯化的cd89/iga阻断小鼠抗人cd89抗体10e7对注射部位处抗小鼠胶原蛋白xvii人iga抗体诱导的表达人cd89的小鼠嗜中性粒细胞流入(influx)的体内作用。
示出平均值
±
sd。黑色圆圈和黑色方块分别代表每只单独小鼠的左耳中的pbs注射和右耳中的抗小鼠胶原蛋白xvii人iga注射。*p《0.05(双尾未配对学生t检验);ns=不显著。
具体实施方式
85.本公开描述了能够结合表达人cd89的细胞上的人cd89(人fcαri)的细胞外部分的抗体。如本文所述的抗体可用于当所述抗体与所述细胞结合时防止人iga与人cd89的结合。已经产生了可与cd89受体结合的几种抗体。认为结合至cd89的ec1结构域的单克隆抗体可阻断iga结合,而结合ec2结构域的抗体不阻止iga与受体的结合。特异性地干扰cd89上的iga结合位点的抗体在本领域中是已知的。例如,如morton等人,j.exp.med.1999年6月7日;189(11):1715-22和shen l.a.,j leukoc biol.1992年4月;51(4):373-8中描述的抗体mip8a、2d11或my43。mip8a是结合至人cd89并具有小鼠igg1恒定区的小鼠单克隆抗体。能够结合cd89的抗体(例如mip8a)可诱导嗜中性粒细胞死亡(wehrli等人,j immunol.2014年12月1日;193(11):5649-59)。
86.在一个方面,本发明提供了新的人源化抗体,所述人源化抗体能够结合表达人cd89的细胞上的人cd89(人fcαri)的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。
87.在一个方面,本发明提供了新的抗体,所述抗体能够结合表达人cd89的细胞上的人cd89(人fcαri)的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且与抗体mip8a相比,对表达人cd89的细胞诱导更少的细胞死亡。
88.术语“抗体”是指通常由两对相同的多肽链构成的免疫球蛋白分子,每对多肽链具有一条“重”(h)链和一条“轻”(l)链。人轻链被分类为κ(κ)和λ(λ)。重链被分类为μ、δ、γ、α或ε,并且将抗体的同种型分别定义为igm、igd、igg、iga和ige。每条重链由重链可变区(文中缩写为hcvr或vh)和重链恒定区构成。igd、igg和iga的重链恒定区由三个结构域ch1、ch2和ch3构成,并且igm和ige的重链恒定区由四个结构域ch1、ch2、ch3和ch4构成。每条轻链由轻链可变区(本文中缩写为lcvr或vl)和轻链恒定区构成。轻链恒定区由一个结构域cl构成。抗体的恒定区可介导免疫球蛋白与宿主组织或因子(包括免疫系统的各种细胞(例如,效应细胞))的结合。vh和vl区可进一步细分为称为互补决定区(cdr)的高变区,其间穿插有称为框架区(fr)的较保守区。每个vh和vl由三个cdr和四个fr构成,按照以下顺序从氨基末端到羧基末端排列:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4。轻链和重链的可变区一起形成抗体结合位点,并定义表位的特异性。本领域已知多种方法将氨基酸分配给抗体中的区或结构域。熟知的方法包括kabat方法和chothia方法(kabat sequences of proteins of immunological interest(national institutes of health,bethesda,md.(1987和1991);chothia等人conformations of immunoglobulin hypervariable regions in nature 1989;342(6252):877-83)。本公开的每个区或结构域的氨基酸分配与kabat的定义一致。
89.术语“抗体”包括鼠抗体、人源化抗体、去免疫化人抗体和嵌合抗体,以及为抗体的多聚体形式(如单体抗体的二聚体、三聚体或更高级多聚体)的抗体。抗体还包括单特异性、双特异性或多特异性抗体,以及包含具有所需特异性的抗原识别位点的免疫球蛋白分子的任何其他经修饰构型。它还包括与非抗体部分连接或附接的抗体。此外,术语“抗体”不受产
生所述抗体的任何特定方法的限制。例如,它包括单克隆抗体、重组抗体和多克隆抗体。本发明提供了如本文所述的抗体。此外,本发明提供了如本文公开的抗体的一部分、衍生物和/或类似物。所述一部分、衍生物和/或类似物在种类上而不一定在数量上保留所述抗体的抗原结合特性。一部分和/或衍生物的非限制性例子包括抗体的一部分是抗原结合部分,并且通常含有所述抗体的一个或多个可变结构域。非限制性实例是各种fab片段。一部分还可以是所谓的单结构域抗体片段。单结构域抗体片段(sdab,开发者ablynx称之为纳米抗体)是具有单个单体可变抗体结构域的抗体片段。像完整的抗体一样,它能够选择性地结合特定抗原。单结构域抗体片段的分子量仅为12-15kda,远小于由两条重蛋白链和两条轻链构成的普通抗体(150-160kda),并且甚至小于fab片段(约50kda,一条轻链和半条重链)和单链可变片段(约25kda,两个可变区,一个来自轻链且一个来自重链)。单结构域抗体本身并不比普通抗体小很多(通常为90-100kda)。单结构域抗体片段大多从骆驼科动物中发现的重链抗体工程化;这些被称为vhh片段一些鱼类也具有仅重链抗体(ignar,“免疫球蛋白新抗原受体”),从中可获得称为vnar片段的单结构域抗体片段。替代方法是将来自人或小鼠的普通免疫球蛋白g(igg)的二聚体可变区分裂成单体。尽管对单结构域抗体的大多数研究目前都是基于重链可变结构域,但源自轻链的纳米抗体也显示出与靶表位特异性结合。抗体部分的非限制性实例含有抗体或其等效物的重链和/或轻链的可变结构域。此类部分的非限制性实例是vhh、人结构域抗体(dab)和单抗体(unibodies)。优选的抗体部分或衍生物至少具有如本文所述抗体的重链和轻链的可变结构域。衍生物或一部分的非限制性实例是f(ab)片段和单链fv片段。双特异性抗体的功能部分包含双特异性抗体的抗原结合部分,或结合部分的衍生物和/或类似物。
[0090]“单链抗体”(scfv)具有包含与vh结构域连接的vl结构域的单个多肽链,其中vl结构域和vh结构域配对形成单价分子。单链抗体可根据本领域已知的方法制备(参见例如,bird等人,(1988)science 242:423-426和huston等人,(1988)proc.natl.acad.sci.usa 85:5879-5883)。“双抗体”具有两条链,每条链包含重链可变区,所述重链可变区与通过短肽接头连接的同一多肽链上的轻链可变区相连,其中同一链上的两个区域彼此不配对,而是与另一条链上的互补结构域配对,以形成双特异性分子。制备双抗体的方法是本领域已知的(参见例如,holliger p.等人,(1993)proc.natl.acad.sci.usa 90:6444-6448和poljak r.j.等人,(1994)structure 2:1121-1123)。结构域抗体(dab)是抗体的小功能结合单位,对应于抗体重链或轻链的可变区。结构域抗体在细菌、酵母和哺乳动物细胞系统中表达良好。结构域抗体及其生产方法的更多细节在本领域是已知的(参见例如,美国专利号6,291,158;6,582,915;6,593,081;wo04/003019和wo03/002609)。纳米抗体源自抗体的重链。纳米抗体通常包含单个可变结构域和两个恒定结构域(ch2和ch3),并保留原始抗体的抗原结合能力。纳米抗体可通过本领域已知的方法制备(参见例如,美国专利号6,765,087、美国专利号6,838,254、wo 06/079372)。单抗体具有igg4抗体的一条轻链和一条重链。单抗体可通过去除igg4抗体的铰链区来制备。单抗体及其制备方法的更多细节可在wo2007/059782中找到。
[0091]
抗体类似物的清单每年都在增加。利用可变结构域的序列和目前对许多不同抗体的3d结构的广泛知识,技术人员可将本发明的抗体转化为一种或另一种抗体类似物、部分或衍生物。
[0092]
除了结合分子之外,本发明的分子还可包含用于增加分子体内半衰期的部分,如但不限于聚乙二醇(peg)、人血清白蛋白、糖基化基团、脂肪酸和葡聚糖。此类另外的部分可使用本领域熟知的方法与结合部分缀合或以其他方式组合。
[0093]
还提供了包含如本文所述的抗体的可变结构域的嵌合抗原受体(car)。car是工程化受体,其将新的特异性(通常是抗体或其衍生物的抗原结合部分)与免疫细胞结合到靶细胞。所述受体被称为嵌合体,因为它们由不同来源的部分融合而成(t淋巴细胞经遗传修饰以表达一种或多种嵌合抗原受体(car;参见例如,eshhar,美国专利号7,741,465;eshhar,美国专利申请公布号2012/0093842)。在一些实施方案中,如本文公开的抗体可与活性化合物例如毒素偶联。此外,如所公开的抗体或抗原结合片段可与标记(例如,荧光蛋白、化学标记、有机染料、有色颗粒或酶)偶联。如本文公开的抗体可与药物偶联,以形成抗体-药物缀合物(adc)。本发明提供了抗体类似物、抗体部分和抗体衍生物,当这些分子与其他分子偶联或掺入时也是如此。
[0094]
在一些实施方案中,如本文公开的抗体是嵌合抗体。术语“嵌合抗体”是指包含源自两种不同物种(如人和小鼠)的氨基酸序列的抗体,通常是小鼠可变(来自重链和轻链)区和人恒定(重链和轻链)区的组合。在工作实施例(例如,实施例6)中描述了产生这种嵌合抗体的非限制性实例。在这种嵌合抗体中,小鼠igg1/κ恒定区被替换为人igg/κ恒定区。
[0095]
在一些实施方案中,如本文公开的抗体是人源化抗体。术语“人源化抗体”是指含有来自非人动物抗体的一些或全部cdr的抗体,而所述抗体的框架和恒定区含有源自人抗体序列的氨基酸残基。人源化抗体通常通过将来自小鼠抗体的cdr移植到人框架序列中,随后用某些人框架残基回代来自源抗体的对应小鼠残基来产生。人源化抗体也可在计算机中使用非人来源的抗体的氨基酸序列和蛋白质设计实验室(pdl)技术(美国专利569376、美国专利5693671、美国专利5585089)生成。与小鼠对应物相比,人源化抗体的优点是在体内耐受性增加。
[0096]
术语“去免疫化抗体”也指非人来源的抗体,其中通常在一个或多个可变区中,一个或多个表位已被去除,其具有构成人t细胞和/或b细胞表位的高倾向,目的是降低免疫原性。表位的氨基酸序列可全部或部分去除。然而,通常通过用一个或多个其他氨基酸取代一个或多个构成表位的氨基酸来改变氨基酸序列,从而将氨基酸序列改变成不构成人t细胞和/或b细胞表位的序列。视情况而定,氨基酸被存在于对应的人可变重链或可变轻链中的一个或多个对应位置处的氨基酸取代。
[0097]
在优选的实施方案中,如本文公开的抗cd89抗体是人源化的。
[0098]
在本文提及“脱酰胺修复的”氨基酸序列的情况下,旨在是指其中易于脱酰胺的氨基酸被不太易于或不易于脱酰胺的氨基酸替代的氨基酸序列。脱酰胺是化学反应,其中氨基酸天冬酰胺或谷氨酰胺的侧链中的酰胺官能团被除去或转化为另一个官能团。通常,天冬酰胺(asn)被转化为天冬氨酸或异天冬氨酸。谷氨酰胺(gln)可被转化为谷氨酸或焦谷氨酸。asn-gly位点最易于脱酰胺,并且天冬酰胺相较于谷氨酰胺更容易脱酰胺。天冬酰胺和/或谷氨酰胺的脱酰胺可改变抗体的结构以及其稳定性和/或功能。在抗体中,这种反应是不合乎需要的,因为它可改变抗体的结构、稳定性或功能(即,抗体-抗原结合),并且可导致抗体降解。可预测抗体可变区中易于脱酰胺的氨基酸(sydow等人plos one 2014;9(6):e100736)。因此,鉴定处于脱酰胺风险的残基,并且然后可用不太易于脱酰胺的氨基酸替代
易于脱酰胺的氨基酸来修复这些位点。
[0099]
在优选的实施方案中,如本文公开的人源化抗cd89抗体是脱酰胺修复的。在一个优选的实施方案中,易于脱酰胺的天冬酰胺被丝氨酸(s)、苏氨酸(t)或谷氨酰胺(q)氨基酸替代。
[0100]
在一些实施方案中,如本文公开的抗体是人抗体。术语“人抗体”是指仅由人免疫球蛋白序列的氨基酸序列组成的抗体。如果在小鼠、小鼠细胞或来源于小鼠细胞的杂交瘤中产生,则人抗体可含有鼠糖链。人抗体可以本领域已知的多种方式制备。嵌合抗体、人源化抗体、去免疫化抗体和人抗体都在本发明的范围内。
[0101]
可结合人cd89的抗体在通常用于抗体结合的条件下与受体结合。当抗体和人cd89受体在适合于抗体结合的条件下彼此接触时,所述抗体将与人cd89受体结合。所述抗体与如以编号dsm acc3341保藏的hek293f细胞上表达的膜结合人cd89结合,而所述抗体不与不在其细胞膜上表达人cd89的hek293f细胞显著结合。所述抗体与表达人cd89的细胞的结合可通过本领域技术人员已知的方法来检测。例如,通过使用携带荧光标记的第二抗体,并使用流式细胞术(facs)测量经标记的细胞。
[0102]
cd89是可结合iga的fc受体。所述受体也被称为fcαri。人cd89能够与人iga1和人iga2的重链恒定区结合。cd89是糖基化跨膜受体,并且具有两个细胞外结构域ec1和ec2、跨膜结构域和细胞内结构域。cd89与iga的相互作用由ec1细胞外结构域介导。对于参考序列,我们参考np_001991.1(免疫球蛋白αfc受体同种型前体)。所述参考仅用于鉴定人cd89基因/蛋白。这并不旨在将如本文所述的人cd89限于数据库条目的特定序列。可结合iga并且可被如本文所述的抗体结合的人cd89的天然变体在本发明的范围内。如果重组人cd89能够结合iga,并且能够结合如本文所述的抗体,则它也在本发明的范围内。
[0103]
cd89存在于髓系细胞的细胞表面上,所述髓系细胞包括嗜中性粒细胞、嗜酸性粒细胞以及大多数单核细胞和巨噬细胞。受体不出现在肥大细胞和肠道巨噬细胞上。cd89的表达被发现是组成性的,并且与iga配体的存在无关。术语“表达人cd89的细胞”是指表达人cd89的细胞。示例性细胞是嗜中性粒细胞、嗜酸性粒细胞、单核细胞和/或巨噬细胞。
[0104]
细胞上cd89受体的交联可通过iga抗体、iga免疫复合物或抗cd89抗体的结合来实现。iga的结合可触发免疫应答,所述免疫应答可具有积极和消极作用。
[0105]
术语“细胞外”的字面意思是细胞外。术语“细胞外部分”是指分子的在细胞膜外部的一部分。分子的这部分可与细胞外的其他分子相互作用。人cd89受体具有两个细胞外结构域,即ec1和ec2。这些结构域可与细胞外的分子(例如iga抗体)相互作用。已知iga与人cd89受体的ec1结构域结合。
[0106]
表达人cd89的细胞是表达人cd89分子的细胞。优选地,所述分子存在于细胞的细胞膜上。表达人cd89的细胞的实例包括但不限于嗜中性粒细胞、嗜酸性粒细胞、单核细胞和/或巨噬细胞。此外,在细胞膜上表达人cd89的经修饰的hek293f细胞系以编号:dsm acc3341保藏。
[0107]
术语“以防止结合”是指如本文公开的抗体或其抗原结合片段阻止iga与人cd89受体相互作用的能力。如果抗cd89抗体或其抗体片段与人cd89受体结合,则iga不能再与人cd89受体结合。优选地,如本文公开的抗cd89抗体的结合将iga与人cd89受体结合的能力阻断或降低至少50%,优选至少60%、至少70%、至少80%或至少90%。iga与cd89的结合的防
止或阻断或降低优选地用实施例中描述的方法来测量。优选地,使用表达cd89的细胞。优选地,所述细胞稳定表达人cd89。在表达cd89的细胞上滴定目标抗体。随后,将细胞与iga一起孵育。洗涤后,使用针对iga的经标记的抗体(优选荧光标记的抗体)检测与细胞结合的iga。可使用流式细胞仪(facs)测量iga在表达人cd89细胞的细胞膜上的结合。结合的iga的量指示经滴定抗体的阻断能力。在一定抗体浓度下iga的结合越少,表明所述抗体的阻断能力越强。在实施例中描述了优选的方法,其结果在图4a和图4b中描绘。测试抗体优选地在人cd89包被的孔上或在表达cd89的hek293f细胞上滴定。然后通过比较滴定曲线与使用阳性对照如mip8a所获得的曲线容易地确定阻断能力。阻断百分比通常以在与在其他方面相同的条件下mip8a的阻断相比时的百分比表示。比较测试抗体和对照抗体mip8a的结合百分比优选地在测试抗体刚刚达到其阻断能力的至少90%的抗体浓度下进行。在图4b中,这是约1ug/ml抗体浓度。在此实施例中,抗体8f3的阻断百分比被计算为约((800-200)/800)x100=约75%。抗体16d6的阻断百分比被计算为约((800-450)/800)x100=约45%。不结合cd89的对照igg1通常不会阻止iga与cd89的结合。
[0108]
免疫球蛋白a是尤其在血液和血清粘膜分泌物中发现的抗体同种型。人iga具有两个子类,即iga1和iga2,并且可以单体和二聚体形式产生。二聚体形式是最普遍的形式。二聚体iga的分泌形式也被称为分泌型iga。iga的分泌形式在其与fcαri结合时部分在空间上受到阻碍。这是因为一些siga的fcαri结合位点被裂解的聚合ig受体(裂解后称为分泌型组分)的区段所掩盖,所述受体帮助siga的分泌物进入肠腔。在与聚合受体二聚体iga(diga)结合之前,与fcαri结合,其亲和力与单聚体iga大致相同。术语“热聚iga”是指通过加热iga而形成的iga的复合物。这些聚集体可模拟免疫复合物,例如循环免疫复合物。热聚iga复合物可通过本领域技术人员已知的任何方法产生。
[0109]
术语“细胞死亡”是指生物细胞停止执行其功能的事件。细胞死亡可由多种原因引起,例如细胞凋亡、程序性细胞死亡、有丝分裂灾难、缺血性细胞死亡和/或免疫原性细胞死亡。术语“细胞活力”涉及细胞执行某些功能的能力,如代谢、生长、繁殖、某种形式的反应性和适应性。细胞死亡和细胞活力可通过熟练的技术人员已知的许多合适的测定进行评价,例如multitox-glo或multitox-fluor多重细胞毒性测定(promega,madison,wisconsin),或活细胞和死细胞测定(live and dead cell assay)(abcam;cambridge,massachusetts)。染料排除法常被用作测定死细胞的一种量度。诸如台盼蓝的染料不容易通过活细胞的细胞膜,但将进入死细胞,因为这些死细胞不能维持其细胞膜的完整性。在实施例部分中描述了用于测定细胞活力的合适的方法。
[0110]
特异性地干扰cd89上的iga结合位点的抗体是本领域中已知的。例如,如morton等人,j.exp.med.1999年6月7日;189(11):1715-22和shen l.a.,j leukoc biol.1992年4月;51(4):373-8中描述的抗体mip8a、2d11或my43。mip8a是结合至人cd89并具有小鼠igg1恒定区的小鼠单克隆抗体。如wehrli等人,j immunol.2014dec 1;193(11):5649-59中所述,mip8a显示诱导嗜中性粒细胞死亡。
[0111]
在一个方面,本公开提供了一种能够结合人cd89的细胞外部分的人源化抗人cd89抗体,所述人源化抗人cd89抗体包含重链可变区,所述重链可变区包含氨基酸序列:
[0112]
evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglex1vx2t ix3gx4gdityy pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0113]
其中:
[0114]
x1是l或w
[0115]
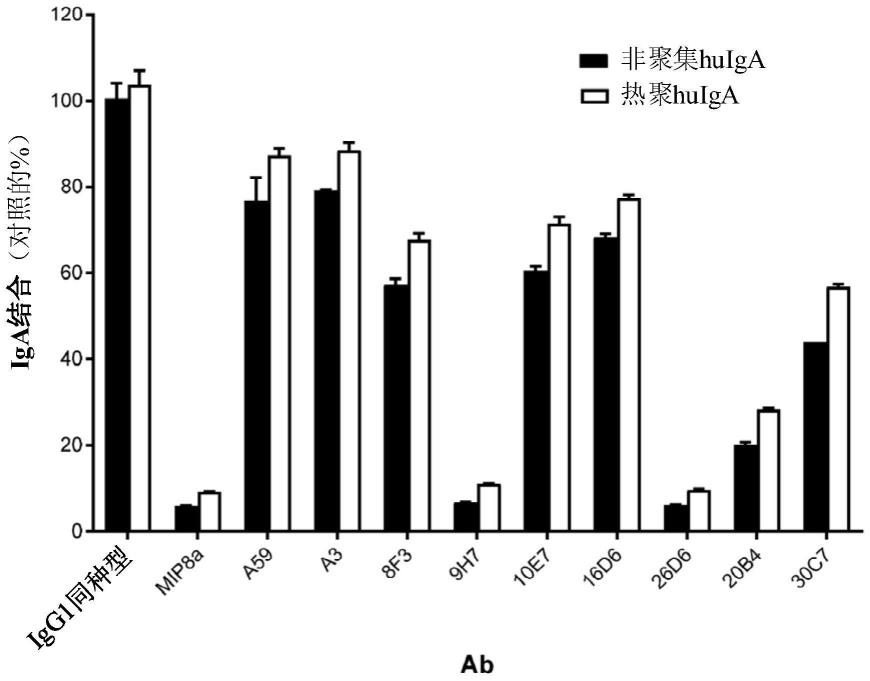
x2是a或s
[0116]
x3是n、s
[0117]
x4是q、t或n,
[0118]
其中所述重链可变区包含在除x1、x2、x3和x4以外的位置处相对于所指示氨基酸序列的优选0-3、优选0-2、优选0-1和优选0个氨基酸变异、插入、缺失、取代、添加或它们的组合;
[0119]
和轻链可变区,所述轻链可变区包含氨基酸序列:
[0120]
diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gkz1z2klliyy tsrlhsgvps rfsgsgsgtd z3tltisslqp edfatyz4cqq gktlpytfgq gtkleik
[0121]
其中:
[0122]
z1是a或t
[0123]
z2是v或p
[0124]
z3是y或f
[0125]
z4是y或f,
[0126]
其中所述轻链可变区包含在除z1、z2、z3和z4以外的位置处相对于所指示氨基酸序列的优选0-3、优选0-2、优选0-1和优选0个氨基酸变异、插入、缺失、取代、添加或它们的组合。
[0127]
在一些实施方案中,x1、x2、x3和x4是l、a、s和t,或l、a、s和q,或l、a、n和n,或w、a、n和n,或w、s、n和n,并且z1、z2、z3和z4是a、v、y和y。
[0128]
在其他实施方案中,x1、x2、x3和x4是l、a、n和n,或w、a、n和n,或w、s、n和n,并且z1、z2、z3和z4是a、p、f和y,
[0129]
在其他实施方案中,x1、x2、x3和x4是l、a、n和n,或w、a、n和n,或w、s、n和n,并且z1、z2、z3和z4是a、p、y和y,
[0130]
在其他实施方案中,x1、x2、x3和x4是l、a、n和n,或w、a、n和n,或w、s、n和n,并且z1、z2、z3和z4是t、v、y和f,
[0131]
在优选的实施方案中,x1、x2、x3和x4是l、a、s和t,并且z1、z2、z3和z4是a、v、y和y。在另一个优选的实施方案中,x1、x2、x3和x4是l、a、s和q,并且z1、z2、z3和z4是a、v、y和y。
[0132]
在优选的实施方案中,如本文公开的抗人cd89抗体包含氨基酸序列evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglelvat ix3gx4gdityy pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0133]
其中所述重链可变区包含在除x3和x4以外的位置处相对于所指示氨基酸序列的优选0-3、优选0-2、优选0-1和优选0个氨基酸变异、插入、缺失、取代、添加或它们的组合;
[0134]
和轻链可变区,所述轻链可变区包含含有0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列。
[0135]
在优选的实施方案中,x3是s,并且x4是t。
[0136]
在另一个优选的实施方案中,x3是s,并且x4是q。
[0137]
如本文公开的重链包含10个,优选9个、优选8个、优选7个、优选6个、优选5个、优选
4个、优选3个、优选2个、优选1个、优选0个氨基酸插入、缺失、取代或添加。在所述0-10个氨基酸插入、缺失、取代或添加中,不超过3个存在于重链可变区中。如本文公开的重链可变区包含3个、优选2个、优选1个、优选0个氨基酸插入、缺失、取代或添加。在优选的实施方案中,所述0-3个氨基酸插入、缺失、取代存在于除x1、x2、x3和x4以外的位置,并且重链可变区中的氨基酸插入、缺失、取代或添加(当存在时)是在框架区中。重链可变区中的氨基酸插入、缺失、取代或添加(当存在时)优选不是选自以下的氨基酸插入、缺失、取代或添加:在所述重链可变区的位置5的亮氨酸(l)、位置19的精氨酸(r)、位置40的丙氨酸(a)、位置42的甘氨酸(g)、位置44的甘氨酸(g)、位置75的丝氨酸(s)、位置84的天冬酰胺(n)、位置87的精氨酸(r)、位置88的丙氨酸(a)、位置93的缬氨酸(v)和位置114的亮氨酸(l)。
[0138]
如本文公开的轻链包含8个,优选7个、优选6个、优选5个、优选4个、优选3个、优选2个、优选1个、优选0个氨基酸插入、缺失、取代或添加。在所述0-8个氨基酸插入、缺失、取代或添加中,不超过5个存在于轻链恒定区中,并且不超过3个存在于轻链可变区中。如本文公开的轻链可变区包含3个、优选2个、优选1个、优选0个氨基酸插入、缺失、取代或添加。在优选的实施方案中,所述0-3个氨基酸插入、缺失、取代或添加存在于除z1、z2、z3和z4以外的位置,并且轻链可变区中的氨基酸插入、缺失、取代或添加(当存在时)是在框架区中。重链可变区中的氨基酸插入、缺失、取代或添加(当存在时)优选不是选自以下的氨基酸插入、缺失、取代或添加:在所述轻链可变区的位置8的脯氨酸(p)、位置15的缬氨酸(v)、位置18的精氨酸(r)、位置22的苏氨酸(t)、位置41的甘氨酸(g)、位置42的赖氨酸(k)、位置70的天冬氨酸(d)、位置72的苏氨酸(t)、位置75的异亮氨酸(i)、位置77的丝氨酸(s)、位置79的谷氨酰胺(q)、位置80的谷氨酰胺(q)、位置83的苏氨酸(t)和位置100的谷氨酰胺(q)。
[0139]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:138的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链可变区。
[0140]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:142的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:136的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:142的氨基酸序列的重链和具有seq id no:136的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3stvl3。
[0141]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:139的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链可变区。
[0142]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:143的氨基酸
序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:136的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:143的氨基酸序列的重链和具有seq id no:136的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3sqvl3。
[0143]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:119的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链可变区。
[0144]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:133的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:136的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:133的氨基酸序列的重链和具有seq id no:136的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3vl3。
[0145]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:119的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:123的氨基酸序列的轻链可变区。
[0146]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:133的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:137的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:133的氨基酸序列的重链和具有seq id no:137的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3vl4。
[0147]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:117的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:120的氨基酸序列的轻链可变区。
[0148]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:131的氨基酸
序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:134的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:131的氨基酸序列的重链和具有seq id no:134的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh1vl1。
[0149]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:117的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:121的氨基酸序列的轻链可变区。
[0150]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:131的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:135的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:131的氨基酸序列的重链和具有seq id no:135的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh1vl2。
[0151]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:117的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链可变区。
[0152]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:131的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:136的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:131的氨基酸序列的重链和具有seq id no:136的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh1vl3。
[0153]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:117的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:123的氨基酸序列的轻链可变区。
[0154]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:131的氨基酸
序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:137的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:131的氨基酸序列的重链和具有seq id no:137的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh1vl4。
[0155]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:118的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:120的氨基酸序列的轻链可变区。
[0156]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:132的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:134的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:132的氨基酸序列的重链和具有seq id no:134的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh2vl1。
[0157]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:118的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:121的氨基酸序列的轻链可变区。
[0158]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:132的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:135的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:132的氨基酸序列的重链和具有seq id no:135的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh2vl2。
[0159]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:118的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:122的氨基酸序列的轻链可变区。
[0160]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:132的氨基酸
序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:136的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:132的氨基酸序列的重链和具有seq id no:136的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh2vl3。
[0161]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:118的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:123的氨基酸序列的轻链可变区。
[0162]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:132的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:137的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:132的氨基酸序列的重链和具有seq id no:137的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh2vl4。
[0163]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:119的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:120的氨基酸序列的轻链可变区。
[0164]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:133的氨基酸序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:134的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:133的氨基酸序列的重链和具有seq id no:134的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3vl1。
[0165]
在一个方面,本公开提供了一种结合人cd89的人源化抗体,所述人源化抗体包含具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:119的氨基酸序列的重链可变区,和具有含0、1、2或3个氨基酸插入、缺失、取代或添加的seq id no:121的氨基酸序列的轻链可变区。
[0166]
在另一方面,本公开提供了一种结合cd89的人源化抗体,其中所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:133的氨基酸
序列的重链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述重链的可变区中;和具有含0、1、2、3、4、5、6、7或8个氨基酸插入、缺失、取代或添加的seq id no:135的氨基酸序列的轻链,其中不超过3个、优选不超过2个、优选不超过1个、优选0个氨基酸插入、缺失、取代或添加存在于所述轻链的可变区中。优选地,可结合人cd89的细胞外部分的所述人源化抗体包含具有seq id no:133的氨基酸序列的重链和具有seq id no:135的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是抗体10e7 vh3vl2。
[0167]
在一个方面,与包含具有seq id no:94的氨基酸序列的重链和具有seq id no:99的氨基酸序列的轻链的嵌合抗体相比,如本文公开的人源化抗体对人cd89的细胞外部分具有更高的亲和力。
[0168]
在一个方面,本发明提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89(人fcαri)的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且与抗体mip8a相比诱导表达人cd89的细胞的更少细胞死亡。优选地,与抗体mip8a相比,所述抗体诱导少10%的细胞死亡。更优选地,与抗体mip8a相比,所述抗体诱导少20%的细胞死亡。更优选地,与抗体mip8a相比,所述抗体诱导少40%的细胞死亡。所述抗体的细胞死亡诱导性质优选地使用以编号:dsm acc3341保藏的表达人cd89的hek293f细胞来确定。
[0169]
靶向效应细胞可在结合抗人cd89抗体后裂解。如本文公开的抗体可用于靶向表达人cd89的细胞,而不会触发靶细胞的广泛细胞死亡或裂解。此类特征可用于维持靶细胞存活。存活细胞可对cd89抗体的结合作出反应,可能是通过改变的信号传导。存活表达cd89的细胞可对缺乏iga与cd89的结合的抗体作出反应,这是由于如本文公开的抗体的阻断性质。具有如本文公开的抗体的细胞可对独立于iga的其他刺激、病原体和/或免疫作用作出反应。
[0170]
在一个实施方案中,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且不会使所述细胞在37℃下过夜孵育后的细胞活力降低超过60%。优选地,所述抗体与所述细胞的结合不会使所述细胞在过夜孵育后的细胞活力降低超过50%、或超过40%、或超过30%、或超过20%、或超过10%或更少。所述抗体的细胞活力保护性质优选地使用以编号:dsm acc3341保藏的表达人cd89的hek293f细胞来确定。
[0171]
细胞通常在37℃下培养。刺激对细胞的影响可直接可见或在孵育期后可见。在影响可见之前,一些刺激需要细胞信号传导。术语“在37℃下过夜孵育”是指细胞与刺激物一起孵育过夜。过夜可意指例如12-16小时或8-24小时,之后测量细胞的特征,例如细胞活力或磷脂酰丝氨酸表达。
[0172]
在一些实施方案中,本公开提供了一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且不会使所述细胞在37℃下过夜孵育后的磷脂酰丝氨酸表达增加超过20%。优选地,所述抗体与所述细胞的结合不会使所述细胞在37℃下过夜孵育后的磷脂酰丝氨酸表达增加超过20%,更优选不会超过10%,更优选不会超过5%。抗体对磷脂酰丝氨酸表达的影响优选使用以编号dsm acc3341保藏的表达人cd89的hek293f细胞来确定。
[0173]
磷脂酰丝氨酸是磷脂并且是细胞膜的组分。它在细胞信号传导中发挥作用,并与细胞死亡和凋亡相关。磷脂酰丝氨酸在细胞的细胞膜上的表达可用作细胞死亡的标志物。磷脂酰丝氨酸在细胞的细胞膜中的表达可通过熟练的技术人员已知的方法来测量。磷脂酰丝氨酸面向细胞膜的细胞质(内)侧活跃。然而,当细胞经历凋亡时,磷脂酰丝氨酸不再局限于细胞溶质侧。相反,它在两侧之间迅速交换。磷脂酰丝氨酸表达通常通过检测细胞膜的向外指向表面上的磷脂酰丝氨酸来确定。磷脂酰丝氨酸的水平可与对照细胞(例如未处理的细胞)进行比较。示例性方法在实施例部分中详细描述。
[0174]
可通过将细胞与抗磷脂酰丝氨酸抗体一起孵育来测定磷脂酰丝氨酸的表达。并行地,细胞可与阴性对照(例如抗人cd19抗体)一起孵育。在洗涤并固定细胞后,可使用流式细胞仪(facs)测量膜磷脂酰丝氨酸表达。
[0175]
在一些实施方案中,提供了一种抗体,所述抗体:能够结合表达人cd89的细胞上的人cd89的细胞外部分;并且当在37℃下、在nan3不存在下所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合;并且当在4℃下、在nan3存在下与所述细胞结合时不能置换单体人iga或热聚iga超过90%。抗体对iga置换的影响优选使用以编号dsm acc3341保藏的表达人cd89的hek293f细胞来确定。
[0176]
在优选的实施方案中,所述抗体是人源化抗体。
[0177]
叠氮化钠与4℃低温的组合用于抑制细胞的代谢活性。叠氮化钠是可逆的线粒体呼吸抑制剂。对细胞代谢活性的抑制是指细胞的线粒体呼吸减少。在一些实施方案中,在代谢受抑制的细胞上,如本文公开的抗体不能置换人iga超过90%。
[0178]
在一个实施方案中,本公开提供了一种以20%或更少地与重组人cd89分子结合的抗体,其中人cd89的氨基酸22-46被替换为食蟹猴cd89的氨基酸22-46,同时所述抗体能够结合表达人cd89的细胞的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。优选地,所述抗体以10%或更少地与所述重组人cd89分子结合。具有这些特征的示例性抗体是抗体20b4。在此cd89分子中取代的氨基酸gln22-lys46是cd89的ec1结构域的一部分(seq id no:23)。在一些实施方案中,与mip8a相比,所述抗体与所述细胞的结合在表达人cd89的细胞中诱导较少细胞死亡。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的细胞活力降低超过60%。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的磷脂酰丝氨酸表达增加超过20%。
[0179]
iga系统在包括人、小鼠和兔子的各种物种之间不同。例如,没有鉴定人cd89基因的小鼠同源物。cd89同源物已在大鼠和牛中鉴定。本公开包括抗体与表达人cd89的人细胞和表达嵌合cd89分子的人细胞上的cd89结合的工作实施例。人/食蟹猴嵌合cd89分子具有人cd89分子的一部分和食蟹猴cd89的一部分。将所述部分组合,使得总体蛋白质结构保持完整。
[0180]
食蟹猴(cynomolgus/macaca fascicularis)cd89基因具有与人cd89相似的内含子/外显子结构,并且表现出与人基因的86%同源性(rogers等人2004,immunology)。用食蟹猴cd89的对应氨基酸取代人cd89的氨基酸可用于测试所述抗体的特异性和交叉反应性。用食蟹猴cd89取代可有助于鉴定抗人cd89抗体的表位。
[0181]
在一个实施方案中,本公开提供了一种以20%或更少地与嵌合cd89分子结合的抗
体,其中人cd89的氨基酸47-71被替换为食蟹猴cd89的氨基酸47-71,同时所述抗体能够结合表达人cd89的细胞的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。优选地,所述抗体以10%或更少地与所述重组人cd89分子结合。优选地,所述抗体以5%或更少地与所述重组人cd89分子结合。具有这些特征的示例性抗体是抗体20b4、8f3、30c7和16d6。氨基酸ile47-ile71是cd89的ec1结构域的一部分(seq id no:24)。iga可与cd89受体的此部分结合。因此,与cd89受体的此部分结合的抗体可能干扰iga与cd89的相互作用。
[0182]
在一个实施方案中,本公开提供了一种以20%或更少地与嵌合cd89分子结合的抗体,其中人cd89的氨基酸72-96被替换为食蟹猴cd89的氨基酸72-96,同时所述抗体能够结合表达人cd89的细胞的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。优选地,所述抗体以10%或更少地与所述重组人cd89分子结合。优选地,所述抗体以5%或更少地与所述重组人cd89分子结合。具有这些特征的示例性抗体是抗体8f3、10e7和16d6。氨基酸gly72-gly96是cd89的ec1结构域的一部分(seq id no:25)。ec1结构域的此部分被认为包含f-g环,其被预测位于ec1底部靠近细胞膜的位置中。
[0183]
在一个实施方案中,本公开提供了一种与嵌合cd89分子的结合不会降低20%或更少的抗体,其中人cd89的氨基酸97-121被替换为食蟹猴cd89的氨基酸97-121,并且所述抗体能够结合表达人cd89的细胞的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。优选地,所述抗体与所述重组人cd89分子的结合不会降低10%或更少。具有这些特征的示例性抗体是抗体20b4、8f3、30c7、10e7和16d6。氨基酸arg97-gly121是cd89的ec1结构域的一部分(seq id no:26)。例如,mip8a抗体可与ec1结构域的此部分结合。
[0184]
在一个实施方案中,本公开提供了一种以20%或更少地与嵌合cd89分子结合的抗体,其中人cd89的氨基酸58、59、73、74、76、106和107分别被替换为食蟹猴cd89的氨基酸58、59、73、74、76、106和107,同时所述抗体能够结合表达人cd89的细胞的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合。优选地,所述抗体以10%或更少地与所述重组人cd89分子结合。优选地,所述抗体以5%或更少地与所述重组人cd89分子结合。具有这些特征的示例性抗体是抗体8f3、10e7和16d6。氨基酸thr58和gln59被认为与人iga-cd89结合有关。
[0185]
在一些实施方案中,与mip8a相比,所述抗体与所述细胞的结合确实在表达人cd89的细胞中诱导较少细胞死亡。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的细胞活力降低超过60%。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的磷脂酰丝氨酸表达增加超过20%。
[0186]
如本文所述的抗体能够结合至表达人cd89的hek293f细胞上的人cd89的细胞外部分。表达膜结合的人cd89的示例性hek293f细胞以根据布达佩斯条约的编号:dsm acc3341保藏。这些hek293f细胞在其细胞表面上稳定表达人cd89。优选地,使用这些细胞允许比较靶向人cd89的不同抗体的作用。优选地,以研究cd89抗体结合后经历细胞死亡的细胞的百分比。使用稳定表达人cd89的细胞系通常减少细胞之间的表达差异,并与瞬时转染的细胞相比允许实验条件的更好比较。
[0187]
本公开的一个方面提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个
氨基酸插入、缺失、取代或添加的seq id no:29-31的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:32-34的cdr1、cdr2和cdr3序列的轻链可变区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:29-31的cdr1、cdr2和cdr3序列的重链可变区和具有seq id no:32-34的cdr1、cdr2和cdr3序列的轻链可变区。
[0188]
在另一方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:27的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:28的氨基酸序列的轻链可变区。在优选的实施方案中,0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:27的氨基酸序列的重链可变区和具有seq id no:28的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是8f3。具有这些cdr的抗体可与cd89的ec1结构域中的表位结合,特别是与seq id no:24和seq id no:25的序列的一部分结合。
[0189]
在一个方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:45-47的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:48-50的cdr1、cdr2和cdr3序列的轻链可变区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:45-47的cdr1、cdr2和cdr3序列的重链可变区和具有seq id no:48-50的cdr1、cdr2和cdr3序列的轻链可变区。
[0190]
在另一方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:43的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:44的氨基酸序列的轻链可变区。在优选的实施方案中,0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:43的氨基酸序列的重链可变区和具有seq id no:44的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是10e7。具有这些cdr的抗体可与cd89的ec1结构域中的表位结合,特别是与seq id no:25的序列的一部分结合。
[0191]
在一个方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:69-71的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:72-74的cdr1、cdr2和cdr3序列的轻链可变区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:69-71的cdr1、cdr2和cdr3序列的重链可变区和具有seq id no:72-74的cdr1、cdr2和cdr3序列的轻链可变区。
[0192]
在另一方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:67的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:68的氨基酸序列的轻链可变区。在优选的实施方案中,0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区。优选地,可结合人cd89的
细胞外部分的抗体包含具有seq id no:67的氨基酸序列的重链可变区和具有seq id no:68的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是20b4。具有这些cdr的抗体可与cd89的ec1结构域中的表位结合,特别是与seq id no:23和seq id no:24的序列的一部分结合。
[0193]
在一个方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:77-79的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:80-82的cdr1、cdr2和cdr3序列的轻链可变区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:77-79的cdr1、cdr2和cdr3序列的重链可变区和具有seq id no:80-82的cdr1、cdr2和cdr3序列的轻链可变区。
[0194]
在另一方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:75的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:76的氨基酸序列的轻链可变区。在优选的实施方案中,0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:75的氨基酸序列的重链可变区和具有seq id no:76的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是30c7。具有这些cdr的抗体可与cd89的ec1结构域中的表位结合,特别是与seq id no:24的序列的一部分结合。
[0195]
在一个方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:53-55的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:56-58的cdr1、cdr2和cdr3序列的轻链可变区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:53-55的cdr1、cdr2和cdr3序列的重链可变区和具有seq id no:56-58的cdr1、cdr2和cdr3序列的轻链可变区。
[0196]
在另一方面,本公开提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:51的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:52的氨基酸序列的轻链可变区。在优选的实施方案中,0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区。优选地,可结合人cd89的细胞外部分的抗体包含具有seq id no:51的氨基酸序列的重链可变区和具有seq id no:52的氨基酸序列的轻链可变区。具有这些特征的示例性抗体是16d6。具有这些cdr的抗体可与cd89的ec1结构域中的表位结合,特别是与seq id no:24和seq id no:25的序列的一部分结合。
[0197]
与mip8a相比,根据序列在本文提及的抗体与细胞的结合在表达人cd89的细胞中可诱导较少的细胞死亡。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的细胞活力降低超过60%。在一些实施方案中,在37℃下过夜孵育后,所述抗体与所述细胞的结合不会使所述细胞的磷脂酰丝氨酸表达增加超过20%。
[0198]
还提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:37-39的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或
2个氨基酸插入、缺失、取代或添加的seq id no:40-42的cdr1、cdr2和cdr3序列的轻链可变区。
[0199]
优选的实施方案提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:35的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:36的氨基酸序列的轻链可变区。在优选的实施方案中,所述氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区中。具有这些特征的示例性抗体是9h7。
[0200]
一个实施方案提供了一种结合人cd89的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:61-63的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:64-66的cdr1、cdr2和cdr3序列的轻链可变区。
[0201]
优选的实施方案提供了一种结合人cd89的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:59的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:60的氨基酸序列的轻链可变区。在优选的实施方案中,所述氨基酸插入、缺失、取代或添加位于轻链和/或重链可变区的框架区中。具有这些特征的示例性抗体是26d6。
[0202]
本公开的抗人cd89抗体或其抗原结合片段优选地包含如本文所述的重链可变区和轻链可变区。这种抗体具有良好特征。当然有可能通过修饰其中的一个或多个氨基酸来产生这种原始抗体的变体。当与所述原始的相比时,许多此类变体将表现出或多或少的相似。此类变体也包括在本公开的范围内。
[0203]
相对于原始抗体的序列,变体可具有氨基酸取代、插入、缺失或添加。氨基酸取代是用另一种氨基酸替代一种氨基酸。优选地,所述氨基酸被具有相似化学特性的氨基酸预先替代,这通常被称为保守取代。氨基酸缺失导致序列中一个或多个氨基酸的缺失。氨基酸插入导致在序列中出现一个或多个另外的氨基酸。氨基酸添加导致在氨基酸序列的起始或末端处出现一个或多个氨基酸。
[0204]
这种修饰的非限制性实例是包含焦谷氨酸盐而不是谷氨酸盐的抗体。此类修饰的其他非限制性例子是与所述原始抗体相比时,一个或多个氨基酸的插入、缺失、倒位和/或取代。优选地,氨基酸取代、插入、缺失或添加在可变结构域的cdr之外。优选地,氨基酸取代、插入、缺失或添加位于所述抗体的可变区的框架区内和/或恒定区内。可如本文所述地测试变体的cd89结合。
[0205]
在一些实施方案中,本发明抗体的恒定区是igg、iga、igd、ige或igm抗体(如igg1、igg2、igg3或igg4抗体)的恒定区。恒定区可包含修饰(如氨基酸取代),以赋予恒定区特定的特性。例如,igg4铰链区的突变使得所述抗体对半分子的交换更加稳定。其他修饰影响所述抗体的半衰期、添加或去除糖基化位点、提高产量、提高在大规模发酵罐中生产的抗体产物的均一性等。
[0206]
本发明的抗体优选地是鼠igg1、在恒定区中突变以减少或阻止补体激活或fc受体相互作用的人igg1、或人igg4、或突变以阻止半分子与其他igg4分子交换的人igg4。在优选的实施方案中,本文公开的抗体是人源化igg4或经人源化igg4突变以防止半分子与其他igg4分子交换,或在恒定区中经人源化igg1突变以减少或防止补体激活或fc受体相互作
用。
[0207]
如本文公开的抗体的恒定区中的一些变异是允许的。通常,恒定区中允许约0-10个氨基酸取代。通常允许多于10个的氨基酸变化。本发明的抗体可具有重链恒定区(ch1-ch2-ch3),其相对于天然存在的重链恒定区(h1-ch2-ch3)具有0-15个、优选0-10个、更优选0-5个、更优选5、4、3、2、1或0个氨基酸取代。这种抗体可具有轻链恒定区,其相对于天然存在的轻链恒定区具有0-5个,优选5、4、3、2、1或0个氨基酸取代。
[0208]
igg4中的一些变异在自然界中发生和/或在不改变所得抗体的免疫学特性的情况下被允许。具有igg4恒定区或突变的igg1恒定区的抗体至少具有抗体的大部分药理学特性,但不结合补体,并且因此将不在体内诱导与其结合的细胞的耗竭。优选地,所述恒定区是人抗体(嵌合)的恒定区。
[0209]
优选地,所述恒定区是在补体激活方面有缺陷的区域,优选人igg4恒定区或突变的人igg1恒定区。在优选的实施方案中,如本文公开的所述人源化抗体具有igg4或igg1同种型。
[0210]
本文公开的抗体及其抗原结合片段的cd89结合可在技术人员已知的许多合适的测定中得到证实。此类测定包括例如,亲和力测定,例如蛋白质印迹、放射免疫测定、facs和elisa(酶联免疫吸附测定)。实施例(例如,实施例2(a))详细描述了可用于测量cd89结合的许多测定中的一些,以及确定抗体对人cd89的相对结合亲和力的方法。
[0211]
术语“结合分子”涵盖(1)抗体,(2)抗体的抗原结合片段,和(3)抗体的衍生物,各自如本文所定义。术语“与cd89结合(binds to cd89)”或“可与cd89结合”或“与cd89结合(binding to cd89)”是指如本文定义的结合分子在体外测定如biacore
tm
(表面等离子体共振)或(生物层干涉法)中与人cd89受体的结合。结合分子具有约1x10-6
m或更低、约1x10-7
m或更低、约1x10-8
m或更低、约1x10-9
m或更低、约1x10-10
m或更低、约1x10-11
m或更低或1x10-12
m或更低的结合亲和力(kd)。所述结合优选是特异性的,这意味着cd89或其表位被结合分子特异性地结合。亲和力是与特定抗原或表位的结合强度的量度。特异性结合,或“特异性地识别”在本文定义为以至多1x10e-6
m、优选至多1x10e-7
m、1x10e-8
m或最优选至多1x10e-9
m的亲和力(kd)结合。
[0212]
如本文下文所述,如本文所述的人源化抗人cd89抗体的kd优选小于1.5nm,如小于1.4nm、小于1.3nm、小于1.2nm、小于1.1nm或小于1nm。与亲本小鼠抗体的kd相比(其是约1.7nm),这一kd出人意料地更低。
[0213]
术语“k
d”是指特定抗体-抗原相互作用的平衡解离常数,并且用来描述配体(如抗体)与蛋白质(如cd89)之间的结合亲和力。平衡解离常数越小,配体结合就越紧密,或配体与蛋白质之间的亲和力越高。kd可通过表面等离子体共振和基于生物层干涉法的测定测量。术语“抗cd89抗体”是指能够结合至cd89(优选人cd89)的如本文定义的抗体。
[0214]
术语“k
on”和“k
off”分别指配体(如抗体)与其靶蛋白(如cd89)缔合(缔合速率)和解离(解离速率)的速率常数。“k
on”表征配体(如抗体)与其靶蛋白(如cd89)结合的速率,并且“k
off”表征配体(如抗体)与其靶蛋白(如cd89)解离的速率。
[0215]
阻断iga与如本文公开的抗体及其抗原结合片段的cd89受体的相互作用的能力可在熟练的技术人员已知的许多合适的测定中得到证实。此类测定包括例如亲和力测定、elisa和facs。所呈现的实施例(例如实施例2(b))详细描述许多测定中的两种,facs和
elisa,它们可用于测试抗cd89抗体阻断iga与cd89受体的结合的能力。
[0216]
为了用elisa测定测试抗体的iga阻断特性,将重组cd89包被在板上。随后,使用封闭缓冲液封闭经包被的板,以防止非特异性结合。将具有重组cd89的板与目标抗体和/或杂交瘤上清一起孵育。随后,将iga与cd89一起添加至孔。洗涤后,使用elisa技术测量所结合的iga的量。所结合的iga的量表示所测试抗体的阻断能力,其中iga结合越少指示抗体的阻断能力越强。此外,可使用表达cd89的细胞来用facs测定测试抗体的iga阻断特性。优选地,所述细胞稳定表达人cd89。将表达cd89的细胞与目标抗体或目标杂交瘤上清一起孵育。随后,将细胞与iga一起孵育。洗涤后,使用针对iga的第二抗体,优选荧光二抗体标记结合至细胞的iga。可使用流式细胞仪(facs)测量iga在表达人cd89细胞的细胞膜上的结合。所结合的iga的量表示所测试抗体的阻断能力,其中iga结合越少指示抗体的阻断能力越强。
[0217]
为了分析如本文公开的纯化的抗人cd89抗体是否能够将先前饱和的iga置换为人cd89,熟练的技术人员可使用许多已知的合适测定。在实施例部分中公开了一种合适的测试方法。在此测定中,允许iga与表达cd89的细胞结合。然后,将抗cd89抗体添加至细胞。可用facs分析测量仍结合在细胞上的iga的量。所述测定在实施例2中详细描述。这种测定和其他测定可用于测定由抗人cd89抗体置换人iga。可使用代谢活跃的细胞(例如在37℃下孵育过夜)或使用代谢不活跃的细胞(例如在叠氮化钠存在下在4℃下孵育)来测量置换。
[0218]
在另一方面,本公开提供了编码如本文公开的抗体或如本文公开的其抗原结合片段的一种或多种核酸分子。还提供了一种编码如本文公开的可变区的核酸分子。如本公开所用的核酸通常但不排他地是核糖核酸(rna)或脱氧核糖核酸(dna)。基于遗传密码,技术人员可以确定编码如本文公开的抗体变体的核酸序列。基于遗传密码的简并性,64个密码子可以用于编码20个氨基酸和翻译末端信号。如技术人员已知的,不同生物体中的密码子使用偏好可能影响基因表达水平。技术人员可利用各种计算工具来优化密码子的使用,这取决于所需核酸将被表达的生物体。
[0219]
在另一方面,本公开提供了一种载体,所述载体包含如本文所述的核酸序列分子。如本文所用的术语“载体”是指能够将异源核酸序列引入宿主细胞中的核酸分子,如质粒、噬菌体或动物病毒。根据本发明的载体允许在宿主细胞中表达或产生由异源核酸序列编码的本发明的抗体。根据本发明使用的载体例如来源于动物病毒,其实例包括但不限于痘苗病毒(包括减毒衍生物,如改良的痘苗病毒安卡拉,mva)、新城疫病毒(ndv)、腺病毒或逆转录病毒。根据本发明的载体优选地包含表达盒,所述表达盒包含适用于在所选择宿主细胞中启动根据本发明的抗体的启动子的转录。用于在真核宿主细胞中表达根据本发明的多肽的合适启动子的实例包括但不限于:β-肌动蛋白启动子、免疫球蛋白启动子、5s rna启动子或病毒衍生启动子,如用于哺乳动物宿主的巨细胞病毒(cmv)、劳斯肉瘤病毒(rsv)和猿猴病毒40(sv40)启动子。
[0220]
当如本文公开的一种或多种核酸分子在细胞中表达时,所述细胞可产生根据本公开的抗体。因此,在一个实施方案中,提供了一种包含根据本公开的抗体、一种或多种核酸分子和/或载体的细胞。宿主细胞可以是哺乳动物、昆虫、植物、细菌或酵母细胞。所述细胞优选地是动物细胞,优选哺乳动物细胞,最优选人细胞。适合作为宿主细胞的哺乳动物细胞系的实例包括杂交瘤细胞、中国仓鼠卵巢(cho)细胞、nso细胞或per-c6
tm
细胞。出于本公开的目的,合适的细胞是能够包含并且优选产生所述抗体和/或所述核酸的任何细胞。本公开
还包括包含所述细胞的细胞培养物。
[0221]
术语“宿主细胞”是指已经引入有表达如本文所述的抗人cd89抗体的表达载体的细胞。所述术语不仅包括特定的受试者细胞,还包括这种细胞的子代。因为后代中可能由于环境影响或突变而发生某些修饰,所以这种子代可能与亲代细胞不同,但仍包括在术语“宿主细胞”的范围内。
[0222]
如本文公开的抗体可以通过技术人员已知的任何方法产生。在优选的实施方案中,使用细胞产生所述抗体,优选地其中所述细胞是杂交瘤细胞、cho细胞、nso细胞或per-c6
tm
细胞。在特别优选的实施方案中,所述细胞是cho细胞,优选地所述细胞在无血清培养基中培养。这包括从所述培养物中收获所述抗体。所述抗体优选地从培养基中纯化,优选地所述抗体被亲和力纯化。可替代地,所述抗体可以合成产生。
[0223]
各种机构和公司已经开发了用于大规模生产抗体的细胞系,例如用于临床使用。这些细胞也用于其他目的,如生产蛋白质。为蛋白质和抗体的工业规模生产而开发的细胞系在本文中进一步称为工业细胞系。因此,本公开的优选实施方案提供了为大规模生产所述抗体而开发的细胞系的用途。
[0224]
根据本发明的抗体表现出多种活性,所述活性可以有利地用于治疗和非治疗用途。特别地,根据本发明的抗体可用于个体的治疗。优选地,根据本发明的抗体可用于治疗免疫相关疾病或预防免疫相关疾病。在一些实施方案中,根据本发明的抗体优选地用于疗法,优选人疗法。在一些实施方案中,如本文公开的抗体可以用于研究目的。例如,用于体外实验、细胞培养、器官型培养和体内模型中。
[0225]
还描述了用于治疗或预防慢性炎症性疾病(cid)的方法。cid的实例包括炎症性肠病(ibd)如溃疡性结肠炎或克罗恩氏病、慢性阻塞性肺病(copd)、哮喘、过敏性和非过敏性鼻炎、食物过敏如乳糜泻以及皮肤病如线性iga大疱病或疱疹样皮炎。导致在cid中观察到的组织破坏的一个共同特征是多形核细胞,特别是嗜中性粒细胞和/或嗜酸性粒细胞的局部累积。多形核细胞是白细胞,其特征是细胞质中存在颗粒。
[0226]
iga与其受体cd89的结合可触发一系列事件,包括免疫细胞的激活,最终导致多形核细胞的迁移、累积和浸润。由于iga免疫复合物对cd89的交联潜在募集和激活嗜中性粒细胞,所以异常iga的存在可能导致促炎应答加剧,从而导致组织损伤。这可在特征在于血清(自身)iga水平增加的不同ccid中起作用,所述cid如iga肾病、亨诺克-斯奇赖恩紫癜(henoch-purpura)、强直性脊柱炎、薛格连氏综合征(syndrome)、酒精性肝硬化、乳糜泻、哮喘、ibd、类风湿性关节炎、线性iga大疱病和疱疹样皮炎(aleyd等人immunol rev 2015;268:123-138)。干扰iga与其受体cd89的结合可抑制信号传导级联和多形核细胞的累积。因此,可通过向需要这种治疗的患者施用有效剂量的本发明的抗体来治疗或预防cid。阻断iga与多形核细胞(如嗜中性粒细胞)上的受体之间的相互作用可停止炎症反应。因此,cid患者可从使用如本文公开的抗体治疗中受益。
[0227]
本发明提供了一种用于治疗患有炎症性疾病的受试者的方法,所述方法包括向所述受试者施用治疗有效量的如本文公开的抗体。还提供了一种制备用于治疗患有炎症性疾病的受试者的药物的方法。本公开描述了用于通过阻断iga与cd89之间的结合来防止免疫细胞激活的方法。
[0228]
本公开还包括一种药物组合物,所述药物组合物包含如本文公开的抗体或其抗原
结合片段,或编码所述抗体或其抗原结合片段的核酸,或包含如本文公开的抗体或其抗原结合片段或编码所述抗体或其抗原结合片段的核酸的细胞。提供了药物组合物,所述药物组合物包含根据本发明的多肽或其药学上可接受的盐,以及至少一种药学上可接受的载体、稀释剂和/或赋形剂。此类组合物特别适合用作药物。所述组合物可呈任何合适的形式,如液体、半固体和固体剂型。所选择的制剂的剂量和时间表可通过熟练技术人员熟知的标准程序来确定。此类程序包括从动物模型中推断和估计给药时间表,然后在人类临床剂量范围研究中确定最佳剂量。药物组合物中的剂量将取决于许多因素而变化,如所需的释放和药效学特征。
[0229]
如本文所用,“受试者”是人或动物。受试者包括但不限于哺乳动物,如人、猪、雪貂、海豹、兔、猫、狗、牛和马,以及鸟类,如鸡、鸭、鹅和火鸡。在本发明的优选实施方案中,受试者是哺乳动物。在特别优选的实施方案中,受试者是人。
[0230]
术语抗体的“抗原结合片段”是指全长抗体的一个或多个部分,所述一个或多个部分保留与所述抗体所结合的相同抗原(即,人cd89)结合的能力。术语“抗原结合片段”还包括抗体的一部分,所述部分是通过非共价或共价缔合形成的较大分子的一部分,或者是具有一个或多个另外分子实体的抗体部分的一部分。另外的分子实体的实例包括氨基酸、肽或蛋白质,如链霉亲和素核心区,其可用于制备四聚体scfv分子(kipriyanov等人hum antibodies hybridomas 1995;6(3):93-101)。示例性抗原结合片段是抗体的vh和/或vl。抗原结合片段包括fab、f(ab')、f(ab')2、互补决定区(cdr)片段、单链抗体(scfv)、二价单链抗体和其他抗原识别免疫球蛋白片段。在一些情况下,如本文所用的术语“抗体”可理解为也包括其抗原结合片段。
[0231]
术语“人抗体”是指仅由人免疫球蛋白序列的氨基酸序列组成的抗体。如果在小鼠、小鼠细胞或来源于小鼠细胞的杂交瘤中产生,则人抗体可含有鼠糖链。人抗体可以本领域已知的多种方式制备。
[0232]
术语“表位”是指能够特异性结合抗体或t细胞受体或以其他方式与分子相互作用的抗原的一部分。“表位”在本领域中也称为“抗原决定簇”。表位通常由分子的化学活性表面分组组成,如氨基酸或碳水化合物或糖侧链。表位可以是“线性的”或“非线性/构象的”。一旦确定了所需的表位(例如,通过表位作图),则可产生针对所述表位的抗体。抗体的产生和表征也可提供关于所需表位的信息。从此信息,然后可以筛选与相同表位结合的抗体,例如通过进行交叉竞争研究来发现相互竞争结合的抗体,即抗体竞争与抗原的结合。
[0233]
如本文所用,“包括”及其变形以其非限制性的意义使用,意指包括所述词后面的项目,但不排除没有具体提到的项目。另外,动词“由
……
组成”可以由“基本上由
……
组成”替换,这意味着如本文定义的化合物或附属化合物可以包含除了具体鉴定的那些之外的一种或多种另外的组分,所述一种或多种另外的组分不改变本发明的独特特征。
[0234]
如本文所用的冠词“一个/种”(“a”和“an”)是指一个/种或多于一个/种(即,至少一个/种)所述冠词的语法宾语。举例来说,“一个(种)要素”意指一个(种)要素或多于一个(种)要素。
[0235]
当与数值(大约10,约10)结合使用时,词语“大约”或“约”优选地意指所述值可以是给定值10的所述值的
±
1%。
[0236]
如本文所用,术语“治疗(treatment)”、“治疗(treat)”和“治疗(treating)”是指
逆转、减轻、延迟疾病或障碍的发作或抑制其进展,或其一种或多种症状,如本文所述。在一些实施方案中,可以在一种或多种症状出现后施用治疗。在其他实施方案中,可以在不存在症状的情况下施用治疗。例如,可在症状发作之前向易感个体施用治疗(如,根据症状史和/或根据遗传因素或其他易感性因素)。治疗也可在症状已经消退之后继续进行,例如,以预防或延迟其复发。
[0237]
出于清楚和简明描述的目的,本文将特征描述为相同或单独实施方案的一部分。然而,将理解,本发明的范围可包括具有所描述特征中的全部或一些的组合的实施方案。
[0238]
在本技术中引用的全部专利和文献参考文件通过引用以其全部内容特此并入。
[0239]
在以下实施例中进一步解释本发明。这些实施例不限制本发明的范围,而仅用于阐明本发明。
[0240]
实施例
[0241]
实施例1.cd89/iga阻断小鼠抗人cd89单克隆抗体的产生
[0242]
(a).用于免疫的瞬时表达表面人cd89的hek293f细胞的产生
[0243]
将编码人全长cd89蛋白的cdna(swiss-prot号p24071.1;参见seq id no:1)优化用于哺乳动物表达,并由德国雷根斯堡的geneart合成(参见seq id no:2)。将此cdna亚克隆到pcdna3.1来源的表达质粒中。使用freestyle
tm 293表达系统(life technologies),将此全长人cd89质粒瞬时转染到freestyle
tm 293f细胞(life technologies)中。在2天后,收获这些hek293f细胞,用无菌磷酸盐缓冲盐水溶液(pbs)洗涤,并在pbs中以≈20.0x106个活细胞/ml等分,并且储存在-80℃下以获得细胞裂解物。在储存之前,使用1:20稀释的藻红蛋白(pe)缀合的小鼠抗人cd89抗体(克隆mip8a;biorad)以流式细胞计数法确认转染的hek293f细胞上的人cd89表面表达。
[0244]
(b).用于筛选的稳定表达表面人cd89的hek293f细胞的产生
[0245]
将编码人全长cd89蛋白的cdna(swiss-prot号p24071.1;参见seq id no:1)优化用于哺乳动物表达,并由德国雷根斯堡的geneart合成(参见seq id no:2)。将此cdna亚克隆到pcdna3.1来源的表达质粒中。使用freestyle
tm 293表达系统(life technologies),将此全长人cd89质粒转染到freestyle
tm 293f细胞(life technologies)中。使用125μg/ml g418/遗传霉素(gibco)选择稳定的人全长cd89转染的hek293f克隆2号。使用1:20稀释的pe缀合的小鼠抗人cd89抗体(克隆mip8a;biorad)以流式细胞计数法确认转染的hek293f细胞上的人cd89表面表达。
[0246]
(c).小鼠抗人cd89单克隆抗体的免疫和产生
[0247]
在第0天将四只balb/c小鼠(雌性,6-8周龄;charles river laboratories)皮下注射水包油乳化sigma adjuvant(sas;sigma)中的约500μl重组c末端多组氨酸标记的人细胞外cd89结构域(ncbi ref seq np_001991.1;sino biological inc)和人cd89瞬时转染的hek293f细胞裂解物(参见以上实施例1(a));将每只小鼠注射与250μl sas混合的250μl pbs中的25μg重组人cd89和人cd89转染的hek293f细胞裂解物(由5x106个活的表达膜结合cd89的细胞制备)。在第14天和第28天,通过皮下注射水包油乳化sas中的重组人cd89和人cd89瞬时转染的hek293f细胞裂解物增强这四只小鼠中的抗体应答;将每只小鼠注射与250μl sas混合的250μl pbs中的25μg重组人cd89和人cd89转染的hek293f细胞裂解物(由5x106个活的表达膜结合cd89的细胞制备)。最后,在第42天和第43天将两只小鼠
(小鼠3号和4号)腹膜内注射不含佐剂的重组人cd89和人cd89瞬时转染的hek293f细胞裂解物;将每只小鼠注射200μl pbs中的20μg重组人cd89和人cd89转染的hek293f细胞裂解物(由4x106个活的表达膜结合cd89的细胞制备)。另外,在第77天和第78天将两只小鼠(小鼠1号和2号)腹膜内注射不含佐剂的重组人cd89和人cd89瞬时转染的hek293f细胞裂解物;将每只小鼠注射200μl pbs中的20μg重组人cd89和人cd89转染的hek293f细胞裂解物(由4x106个活的表达膜结合cd89的细胞制备)。在第46天(对于融合i;小鼠3号和4号)或在第81天(对于融合ii;小鼠1号和2号),如下文所述使用标准杂交瘤技术(最初由和milstein在nature 1975,256:495中描述)将来自这些经免疫小鼠的脾细胞与sp2/0-ag14骨髓瘤细胞(dsmz)融合。简言之,将经免疫小鼠处死。从脾中挑取脾细胞,并在含glutamax培养基(sf培养基;invitrogen)的无血清opti-i中洗涤。将对数生长的sp2/0-ag14骨髓瘤细胞在sf培养基中洗涤,并添加到脾细胞中,从而产生5:1比率的脾细胞与骨髓瘤细胞。然后沉淀细胞,并去除上清液。然后在60秒时间段内逐滴添加1ml 37%(v/v)的聚乙二醇4000(merck)溶液,之后将细胞在37℃下再孵育60秒。然后在温和搅拌下缓慢添加8ml sf培养基,随后是5ml含glutamax/10%(v/v)胎牛血清(fcs;bodinco)的opti-i。在室温(rt)下30分钟后,将细胞沉淀,在含有glutamax/10% fcs的opti-i中洗涤以除去残留的聚乙二醇,并最终以在氨基蝶呤选择培养基中的0.1x106个细胞/200μl/孔的浓度铺板(即,opti-i,其含有补充有50x hybri-max
tm
氨基蝶呤(一种从头dna合成抑制剂;sigma)的glutamax/10% fcs)中。从第7天开始,每2-3天重新填满氨基蝶呤选择培养基,并且在第12-14天,用含有glutamax/10% fcs的opti-mem i替换氨基蝶呤选择培养基。
[0248]
(d).筛选小鼠抗人cd89单克隆抗体的存在
[0249]
从每次融合后第12-14天开始,使用以重组c-末端多组氨酸标记的人(细胞外)cd89(rhucd89;sino biological)作为靶蛋白的elisa,筛选来自生长中杂交瘤的上清液中是否存在igg类别(即,“高亲和力”igg,与“低亲和力”igm相反)的小鼠抗人cd89抗体。为此,在4℃-8℃下在16-24小时期间,使用半面积96孔eia板(corning),以0.5μg/ml将rhucd89包被在pbs中(25ng/50μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1%牛血清白蛋白(bsa;roche)封闭1小时。随后,将板与50μl未稀释的杂交瘤上清液/孔在室温下孵育1小时。并行地,将50μl培养基(含glutamax/10%fcs的opti-i)和50μl的10μg/ml的小鼠抗人cd89抗体克隆mip8a(biorad)(在培养基中稀释)分别作为阴性对照和阳性对照运行。在pbs/0.05%tween 20中充分洗涤后,在室温下用1:5000稀释的辣根过氧化物酶(hrp)缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)测定抗体在rhucd89上的结合,持续1小时,随后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m h2so4后,使用微板读数器(imark;biorad)在450nm波长(参考波长为655nm)下测量抗体在rhucd89上的结合(光密度)。
[0250]
从融合后第12-14天开始,还使用以膜结合人cd89作为靶蛋白的facs,筛选并证实来自生长中杂交瘤的上清液的igg类别(即,“高亲和力”igg,与“低亲和力”igm相反)的小鼠抗人cd89抗体产生。为此,将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa
(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与100μl未稀释的杂交瘤上清液/管在4℃下孵育30分钟。并行地,将100μl培养基(含glutamax/10% fcs的opti-i)、100μl的10μg/ml小鼠igg1同种型对照(bd biosciences)(在培养基中稀释)、100μl的10μg/ml小鼠igg2a同种型对照(bd biosciences)(在培养基中稀释)作为阴性对照运行,并将100μl的10μg/ml小鼠抗人cd89抗体克隆mip8a(biorad)(在培养基中稀释)作为阳性对照运行。未转染(即,对膜结合人cd89表达呈阴性)的野生型(wt)hek293f细胞也作为阴性对照细胞运行,以确定抗体特异性。在pbs/bsa/nan3中充分洗涤后,随后在4℃下将细胞与1:200稀释的pe缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量抗体在膜人cd89上的结合(几何平均荧光强度)。
[0251]
将双重cd89阳性(即在elisa中rhucd89 (数据未示出)和在facs中膜cd89 hek293f细胞(图1))杂交瘤细胞扩增并冷冻保存。来自这些双重cd89阳性杂交瘤的上清液显示与未转染的wt hek293f细胞(即,对膜结合人cd89表达呈阴性)无反应性。如图1所示,这种方法产生了21个产生小鼠抗人cd89特异性抗体的杂交瘤。随后,测试这些产生小鼠抗人cd89特异性抗体的杂交瘤的上清液阻断血清人iga在其受体cd89上的结合的能力(参见下文的实施例1(e))。
[0252]
(e).筛选cd89/iga阻断小鼠抗人cd89单克隆抗体的存在
[0253]
为了分析小鼠抗人cd89抗体对人血清iga与人cd89的结合的影响,通过使用elisa和facs分析来测定小鼠抗人cd89抗体在空间上阻碍人血清iga与人cd89的相互作用的能力。
[0254]
elisa:在4℃-8℃下在16-24小时期间,使用半面积平底96孔eia板(corning),将rhucd89(sino biological)以0.5μg/ml包被在pbs中(25ng/50μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1% bsa(roche)封闭1小时。随后,将板与25μl未稀释的杂交瘤上清液/孔在室温下孵育30小时。并行地,将25μl培养基(含glutamax/10% fcs的opti-i)和25μl的20μg/ml的小鼠抗人cd89抗体克隆mip8a(biorad)(在培养基中稀释)分别作为阴性对照和阳性对照运行。在此之后(即,不洗涤),将25μl的2μg/ml纯化的人(血清来源的)iga(bethyl laboratories)(在培养基中稀释)添加至这些孔中,并在室温下再孵育30分钟。在pbs/0.05% tween 20中充分洗涤后,在室温下用1μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定血清人iga在rhucd89上的结合,持续1小时。在pbs/0.05% tween 20中充分洗涤后,添加1:10,000稀释的hrp缀合的链霉亲和素(jackson immunoresearch),并在室温下孵育1小时,然后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m h2so4后,使用微板读数器(imark;biorad)在450nm波长(参考波长为655nm)下测量血清人iga在rhucd89上的结合(光密度)。
[0255]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))以10x106个细胞/ml置于冰冷冻了的pbs中,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与50μl未稀释的杂交瘤
上清液/管在4℃下孵育30分钟。并行地,将50μl培养基(含glutamax/10% fcs的opti-i)、50μl的20μg/ml小鼠igg1同种型对照(bd biosciences)(在培养基中稀释)、50μl的20μg/ml小鼠igg2a同种型对照(bd biosciences)(在培养基中稀释)作为阴性对照运行,并将50μl的20μg/ml小鼠抗人cd89抗体克隆mip8a(biorad)(在培养基中稀释)作为阳性对照运行。在此之后(即,不洗涤),将50μl的20μg/ml纯化的人(血清来源的)iga(bethyl laboratories)(在培养基中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0256]
如图2a所示,来自产生小鼠抗人cd89特异性抗体的杂交瘤的21个检查的上清液中的6个(即,8f3、9h7、10e7、26d6、20b4和30c7)显示血清人iga与rhucd89的结合的强烈/完全阻断,而来自产生小鼠抗人cd89特异性抗体的杂交瘤的21个检查的上清液中的1个(即,16d6)显示血清人iga与rhucd89的结合的中等/部分阻断。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且显示血清人iga与rhucd89的结合的强烈/完全阻断。
[0257]
如图2b所示,来自产生小鼠抗人cd89特异性抗体的杂交瘤的21个检查的上清液中的6个(即,8f3、9h7、10e7、26d6、20b4和30c7)显示血清人iga与膜结合人cd89的结合的强烈/完全阻断,而来自产生小鼠抗人cd89特异性抗体的杂交瘤的21个检查的上清液中的1个(即,16d6)显示血清人iga与膜结合人cd89的结合的中等/部分阻断。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(biorad)(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且显示血清人iga与膜结合人cd89的结合的强烈/完全阻断。
[0258]
使用蛋白g柱(ge healthcare)从上述产生cd89/iga阻断小鼠抗人cd89特异性抗体的杂交瘤上清液中纯化小鼠抗体。使用isostrip
tm
小鼠单克隆抗体同种型分型试剂盒(roche)对重链和轻链的同种型类别进行分型,并且所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,8f3、9h7、10e7、16d6、26d6、20b4和30c7)被发现是igg1/κ。此外,使用lal显色终点测定(hycult biotech)测定lps水平,并且所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,8f3、9h7、10e7、16d6、26d6、20b4和30c7)都含有《0.005eu lps/μg小鼠igg。随后,详细测试了这些纯化的cd89/iga阻断小鼠抗人cd89特异性抗体对人cd89的相对结合亲和力,对血清人iga与人cd89的结合的阻断作用,以及对血清人iga介导的表达人cd89的原代人嗜中性粒细胞的吞噬作用的阻断作用和对血清人iga介导的表达人cd89的原代人嗜中性粒细胞的迁移的阻断作用,如实施例2和3中所述。此外,通过与已知的商业cd89/iga阻断和非阻断小鼠抗人cd89特异性抗体的交叉竞争、通过人cd89/牛fcγ2r结构域作图、通过在食蟹猴cd89上的跨物种结合以及通过人/食蟹猴cd89表位作图来检查这些纯化的cd89/iga阻断小鼠抗人cd89特异性抗体的良好特异性,如实施例4中所述。
[0259]
实施例2.cd89/iga阻断小鼠抗人cd89单克隆抗体的结合表征
[0260]
(a).cd89/iga阻断小鼠抗人cd89抗体对人cd89的相对结合亲和力
[0261]
为了确定纯化的cd89/iga阻断小鼠抗人cd89抗体对人cd89的相对结合亲和力,使用了elisa和facs分析。
[0262]
elisa:在4℃-8℃下在16-24小时期间,使用半面积平底96孔eia板(corning),将rhucd89(sino biological)以0.5μg/ml包被在pbs中(25ng/50μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1% bsa(roche)封闭1小时。随后,将板与50μl滴定的(在封闭缓冲液中)纯化的小鼠抗人cd89抗体/孔在室温下孵育1小时。并行地,50μl滴定的(在封闭缓冲液中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)作为阳性对照运行。在pbs/0.05% tween 20中充分洗涤后,在室温下用1:5000稀释的辣根过氧化物酶(hrp)缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)测定抗体在rhucd89上的结合,持续1小时,随后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m h2so4后,使用微板读数器(imark;biorad)在450nm波长(参考波长为655nm)下测量抗体在rhucd89上的结合(光密度)。
[0263]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与100μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体/管在4℃下孵育30分钟。并行地,100μl滴定的(在pbs/bsa/nan3中)纯化的小鼠igg1同种型对照(bd biosciences)和100μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)分别作为阴性对照和阳性对照运行。在pbs/bsa/nan3中充分洗涤后,随后将细胞与1:200稀释的pe缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量抗体在膜人cd89上的结合(几何平均荧光强度)。
[0264]
如图3a所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都剂量依赖性地结合至rhucd89。基于它们的结合概况,发现了以下相对亲和力排序(从高至较低亲和力):9h7=26d6=20b4》8f3=10e7=30c7(=mip8a)》16d6。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示与rhucd89的剂量依赖性结合。
[0265]
如图3b所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都剂量依赖性地结合至膜人cd89。基于它们的结合概况,发现了以下相对亲和力排序(从高至较低亲和力):9h7=26d6=20b4(=mip8a)》8f3=10e7=30c7》16d6,其与在elisa中发现的相对亲和度排序一致。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示与膜人cd89的剂量依赖性结合。
[0266]
(b).cd89/iga阻断小鼠抗人cd89抗体的cd89/iga阻断能力的程度
[0267]
为了分析纯化的小鼠抗人cd89抗体的cd89/iga阻断的程度,通过使用elisa和facs分析来测定纯化的cd89/iga阻断小鼠抗人cd89抗体在空间上阻碍人iga与人cd89的相互作用的能力。
[0268]
elisa:在4℃-8℃下在16-24小时期间,使用半面积平底96孔eia板(corning),将rhucd89(sino biological)以0.5μg/ml包被在pbs中(25ng/50μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1% bsa(roche)封闭1小时。随后,将板与25μl滴定的(在封闭缓冲液中)纯化的小鼠抗人cd89抗体/孔在室温下孵育30分钟。并行地,25μl滴定的(在封闭缓冲液中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)作为阳性对照运行。在此之后(即,不洗涤),将25μl的2μg/ml纯化的人(血清来源的)iga(bethyl laboratories)(在封闭缓冲液中稀释)添加至这些孔中,并在室温下再孵育30分钟。在pbs/0.05% tween 20中充分洗涤后,在室温下用1μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定血清人iga在rhucd89上的结合,持续1小时。在pbs/0.05% tween 20中充分洗涤后,添加1:10,000稀释的hrp缀合的链霉亲和素(jackson immunoresearch),并在室温下孵育1小时,然后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m h2so4后,使用微板读数器(imark;biorad)在450nm波长(参考波长为655nm)下测量血清人iga在rhucd89上的结合(光密度)。
[0269]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体/管在4℃下孵育30分钟。并行地,50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠igg1同种型对照(bd biosciences)和50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)分别作为阴性对照和阳性对照运行。在此之后(即,不洗涤),将50μl的20μg/ml纯化的人(血清来源的)iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0270]
如图4a所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都剂量依赖性地阻止了血清人iga与rhucd89的结合。基于它们的cd89/iga阻断概况,发现了以下排序(从强至较弱的cd89/iga阻断程度):9h7=26d6=20b4》8f3=10e7=30c7(=mip8a)》16d6。令人感兴趣地,在这些经检查的纯化的小鼠抗人cd89抗体在空间上阻断血清人iga与rhucd89的结合的程度(此实施例)与它们对rhucd89的相应相对结合亲和力(参见以上实施例2(a))之间似乎存在强正相关关系。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示血清人iga与rhucd89的结合的剂量依赖性阻断。
[0271]
如图4b所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都剂量依赖性地阻止了血清人iga与膜人cd89的结合。基于它们的cd89/iga阻断概况,发现了以下排序(从强至较低的cd89/iga阻断程度):9h7=26d6=20b4(=mip8a)》8f3=10e7=30c7》16d6,其
与在elisa中发现的cd89/iga阻断程度一致。令人感兴趣地,在这些经检查的纯化的小鼠抗人cd89抗体在空间上阻断血清人iga与膜人cd89的结合的程度(此实施例)与它们对膜人cd89的相应相对结合亲和力(参见以上实施例2(a))之间似乎存在强正相关关系。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示血清人iga与膜人cd89的结合的剂量依赖性阻断。
[0272]
为了分析纯化的小鼠抗人cd89抗体的cd89/iga阻断的程度,通过使用facs分析来测定纯化的cd89/iga阻断小鼠抗人cd89抗体在空间上阻碍分泌型人iga与人cd89的相互作用的能力。
[0273]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))以10x106个细胞/ml置于冰冷冻了的pbs中,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体/管在4℃下孵育30分钟。并行地,50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)作为阳性对照运行。在此之后(即,不洗涤),将50μl的0.16μm纯化的人(初乳来源的)iga(biorad)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定分泌型人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的4%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量分泌型人iga在膜人cd89上的结合(几何平均荧光强度)。
[0274]
如图4c所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都剂量依赖性地阻止了分泌型人iga与膜人cd89的结合。基于它们的cd89/iga阻断概况,发现了以下排序(从强至较低的cd89/iga阻断程度):9h7=26d6=20b4(=mip8a)》8f3=10e7=30c7》16d6,其与在facs中发现的使用血清人iga的cd89/iga阻断程度(参见图4b)一致。令人感兴趣地,在这些经检查的纯化的小鼠抗人cd89抗体在空间上阻断分泌型人iga与膜人cd89的结合的程度(此实施例)与它们对膜人cd89的相应相对结合亲和力(参见以上实施例2(a))之间似乎存在强正相关关系。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示分泌型人iga与膜人cd89的结合的剂量依赖性阻断。
[0275]
人cd89被描述为对单体人iga具有低/中等亲和力的受体(ka≈106m-1
),而人iga免疫复合物与人cd89紧密结合(bakema等人immunol rev 2011;4:612-624)。为了模拟人iga免疫复合物,将人(血清来源的)iga(bethyl laboratories)在63℃下加热30分钟,并冷却至室温,然后在4℃ 12000
×
g下离心2分钟,以去除任何不溶性蛋白质沉淀。使用尺寸排除色谱分析将这种热聚(血清来源的)人iga与非聚集(血清来源的)人iga进行了比较,并且证明热聚人iga由≈40%的单体、≈15%的二聚体和≈45%的四聚体或更高级多聚体人iga组成,而非聚集人iga由≈70%的单体、≈20%的二聚体和≈10%的三聚体人iga组成。然后,通过使用facs分析(参见下文)测定我们的纯化的cd89/iga阻断小鼠抗人cd89抗体在空间
上阻碍这种热聚血清人iga相比非聚集血清人iga与膜结合人cd89之间的相互作用的能力。
[0276]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与50μl的20μl/ml纯化的小鼠抗人cd89抗体(在pbs/bsa/nan3中)在4℃下孵育30分钟。并行地,50μl的20μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在pbs/bsa/nan3中)和50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111);biorad)(在pbs/bsa/nan3中)分别作为阴性对照和阳性对照运行。此外,50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);bd biosciences)(在pbs/bsa/nan3中)和50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);santa cruz biotechnology)(在pbs/bsa/nan3中)作为额外的阴性对照运行。在此之后(即,不洗涤),将50μl的20μg/ml纯化的非聚集或热聚人(血清来源的)iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定非聚集或热聚血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0277]
如图5a所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都阻止了热聚血清人iga与膜人cd89的结合,达到与在非聚集血清人iga情况下发现的类似程度。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示热聚血清人iga与膜人cd89的结合的阻断,达到与在非聚集血清人iga情况下发现的类似程度。出人意料地,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示热聚血清人iga与膜人cd89的结合的部分(≈60%)阻断,达到与在非聚集血清人iga情况下发现的类似程度。
[0278]
总之,这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7阻止了单体、二聚体、三聚体、四聚体或更高级多聚体血清人iga(即,非聚集和热聚iga)和二聚体分泌型人iga与膜人cd89的结合。关于总结,参见表1a(即,血清人iga阻断)。
[0279]
(c).通过cd89/iga阻断小鼠抗人cd89抗体进行血清人iga置换
[0280]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体是否能够将先前饱和的血清人源iga置换为人cd89,通过使用facs分析来测定纯化的小鼠抗人cd89抗体对血清人iga置换为hek293f细胞上的膜结合人cd89的影响。
[0281]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃
下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与50μl的20μg/ml纯化的非聚集或热聚人(血清来源的)iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)在4℃下孵育30分钟。在此之后(即,不洗涤),将50μl的20μg/ml纯化的小鼠抗人cd89抗体(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。并行地,50μl的20μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在pbs/bsa/nan3中)和50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111);biorad)(在pbs/bsa/nan3中)分别作为阴性对照和阳性对照运行。此外,50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);bd biosciences)(在pbs/bsa/nan3中)和50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);santa cruz biotechnology)(在pbs/bsa/nan3中)作为额外的阴性对照运行。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab’)2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定非聚集或热聚血清人iga在膜人cd89上的结合30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0282]
如图5b所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都不同程度地置换了先前饱和的膜人cd89上的非聚集血清人iga。基于它们的血清人iga置换程度,发现以下排序(从强至较弱的人iga置换程度):9h7=26d6(=mip8a)》20b4》30c7》8f3=10e7》16d6。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也置换了先前饱和的膜人cd89上的非聚集血清人iga。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-没有置换先前饱和的膜人cd89上的非聚集血清人iga。
[0283]
如图5b所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(除16d6外)也在不同程度上置换了先前饱和的膜人cd89上的热聚血清人iga,并且其程度略低于在非聚集血清人iga的情况下所发现的程度。基于它们的血清人iga置换程度,发现以下排序(从强至较弱的人iga置换程度):9h7=26d6(=mip8a)》20b4》30c7》8f3=10e7。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也置换了先前饱和的膜人cd89上的非聚集血清人iga,尽管其程度略低于在非聚集血清人iga的情况下所发现的程度。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-没有置换先前饱和的膜人cd89上的热聚血清人iga。
[0284]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、(16d6)、26d6、20b4和30c7显示置换先前饱和的膜人cd89上的单体、二聚体和三聚体血清人iga(即非聚集iga),并且在稍微更低程度上置换先前饱和的膜人cd89上的四聚体或更高级多聚体
血清人iga(即热聚iga)。关于总结,参见表1a。
[0285]
表1a.在流式细胞计数和代谢非活性条件下(即,在nan3存在下和在低温环境温度(4℃)下),饱和浓度(即,10μg/ml)下的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体在膜人cd89上的iga阻断和iga置换概况。
[0286]
通过小鼠抗人cd89抗体进行的iga阻断或iga置换
[0287][0288]-=膜人cd89上没有通过小鼠抗cd89抗体的iga结合的阻断或没有通过小鼠抗cd89抗体的iga置换(图5a和5b中的76%-125% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的弱阻断或弱iga置换(图5a和5b中的51%-75% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的中等阻断或中等iga置换(图5a和5b中的26%-50% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的强阻断或强iga置换(图5a和5b中的0%-25% iga结合)。aggr=聚集的。
[0289]
由于所有上述实验(参见实施例2(b)和实施例2(c))都是在(流式细胞计数)代谢非活性条件(即,在低温环境温度(4℃)下,并在nan3(一种线粒体呼吸的可逆抑制剂)存在下,其防止抗体与受体结合后抗体-抗原复合物的加帽、脱落和内化)下进行的,所以我们还检查了我们的纯化的cd89/iga阻断小鼠抗人cd89抗体在代谢(活性)条件下,在空间上阻碍
非聚集和热聚血清人iga与膜结合人cd89的相互作用的能力,如下:
[0290]
人iga阻断设置:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以1.70x106个细胞/ml置于补充有125μg/ml g418/遗传霉素(gibco)的freestyle
tm 293培养基(life technologies)中。然后,将400μl/管(即,0.7x106个细胞)的这些细胞与或不与50μl的100μl/ml纯化的小鼠抗人cd89抗体(在freestyle
tm 293培养基中)在4℃下孵育30分钟。并行地,50μl的100μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在freestyle
tm 293培养基中)和50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111);biorad)(在freestyle
tm 293培养基中)分别作为阴性对照和阳性对照运行。此外,50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);bd biosciences)(在freestyle
tm 293培养基中)和50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);santa cruz biotechnology)(在pbs/bsa/nan3中)作为额外的阴性对照运行。在此之后(即,不洗涤),将50μl的100μg/ml纯化的非聚集或热聚人(血清来源的)iga(bethyl laboratories)(在freestyle
tm 293培养基中)添加至这些细胞中,并在37℃下在5% co2孵育箱中再孵育24小时。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定非聚集或热聚血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0291]
人iga置换设置:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以1.70x106个细胞/ml置于补充有125μg/ml g418/遗传霉素(gibco)的freestyle
tm 293培养基(life technologies)中。然后,将400μl/管(即,0.7x106个细胞)的这些细胞与50μl的100μg/ml纯化的非聚集或热聚人(血清来源的)iga(bethyl laboratories)(在freestyle
tm 293培养基中)在4℃下孵育30分钟。在此之后(即,不洗涤),将50μl的100μg/ml纯化的小鼠抗人cd89抗体(在freestyle
tm 293培养基中)添加至这些细胞中,并在37℃下在5% co2孵育箱中再孵育24小时。并行地,50μl的100μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在freestyle
tm 293培养基中)和50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111);biorad)(在freestyle
tm 293培养基中)分别作为阴性对照和阳性对照运行。此外,50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);bd biosciences)(在freestyle
tm 293培养基中)和50μl的100μg/ml纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);santa cruz biotechnology)(在pbs/bsa/nan3中)作为额外的阴性对照运行。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定非聚集或热聚血清人iga在膜人cd89上的结合,持续30分
钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0292]
如图5c所示,纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、26b6、20b4和30c7在代谢活性条件下强烈抑制了非聚集和热聚血清人iga与膜人cd89的结合。纯化的cd89/iga阻断小鼠抗人cd89特异性抗体16d6显示部分阻断非聚集血清人iga与膜人cd89的结合,而小鼠抗人cd89抗体16d6在代谢活性条件下不能阻断热聚血清人iga与膜人cd89的结合。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示在代谢活性条件下非聚集和热聚血清人iga与膜人cd89的结合的强阻断。出人意料地,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示部分阻断非聚集血清人iga与膜人cd89的结合,而小鼠抗人cd89抗体克隆a59和克隆a3在代谢活性条件下不能阻断热聚血清人iga与膜人cd89的结合。
[0293]
如图5d所示,纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、26b6、20b4和30c7在代谢活性条件下强烈置换了先前饱和的膜人cd89上的非聚集和热聚血清人iga。纯化的cd89/iga阻断小鼠抗人cd89特异性抗体16d6可部分置换先前饱和的膜人cd89上的非聚集血清人iga,而小鼠抗人cd89抗体16d6在代谢活性条件下不能置换先前饱和的膜人cd89上的热聚血清人iga。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也能够在代谢活性条件下强烈地置换先前饱和的膜人cd89上的非聚集和热聚血清人iga。出人意料地,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-能够部分置换先前饱和的膜人cd89上的非聚集血清人iga,而小鼠抗人cd89抗体克隆a59和克隆a3在代谢活性条件下不能置换先前饱和的膜人cd89上的热聚血清人iga。
[0294]
这些结果表明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、26d6、20b4和30c7在代谢活性条件下抑制膜人cd89上单体、二聚体和三聚体血清人iga(即,非聚集iga)以及四聚体或更高级多聚体血清人iga(即,热聚iga)的结合。此外,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、26d6、20b4和30c7在代谢活性条件下也置换了先前饱和的膜人cd89上的单体、二聚体和三聚体血清人iga(即,非聚集iga)以及四聚体或更高级多聚体血清人iga(即,热聚iga)。关于总结,参见表1b。
[0295]
表1b.在代谢(活性)条件下(即,在nan3不存在下和在生理环境温度(37℃)下),在24小时后饱和浓度(即,10μg/ml)下的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体在膜人cd89上的iga阻断和iga置换概况。
[0296]
通过小鼠抗人cd89抗体进行的iga阻断或iga置换
[0297][0298]-=膜人cd89上没有通过小鼠抗cd89抗体的iga结合的阻断或没有通过小鼠抗cd89抗体的iga置换(图5c和5d中的76%-125% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的弱阻断或弱iga置换(图5c和5d中的51%-75% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的中等阻断或中等iga置换(图5c和5d中的26%-50% iga结合), =膜人cd89上通过小鼠抗cd89抗体的iga结合的强阻断或强iga置换(图5c和5d中的0%-25% iga结合)。aggr=聚集的。
[0299]
cd89/iga阻断小鼠抗人cd89抗体克隆mip8a被描述为诱导表达cd89的人嗜中性粒细胞中的人细胞死亡(wehrli等人j immunol 2014,193:5649-5659)。取决于炎症微环境,通过暴露于二价小鼠抗人cd89抗体克隆mip8a,诱导这些人嗜中性粒细胞中的半胱天冬酶依赖性(通常在凋亡过程中观察到)或半胱天冬酶独立性(非凋亡)细胞死亡。因此,使用稳定的人全长cd89转染的hek293f细胞作为靶细胞,检查了我们的所产生的cd89/iga阻断小鼠抗人cd89抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7对人cd89介导的细胞死亡的影响。
[0300]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以1.25x106个细胞/ml置于补充有125μg/ml g418/遗传霉素(gibco)的freestyle
tm 293
immunol ther exp 2001;49:217-229;bakema等人immunol rev 2011;4:612-624)。更具体地说,原型cd89/iga阻断小鼠抗人cd89抗体克隆mip8a识别人cd89的ec1结构域内的表位(lu等人protein sci 2014;23:378-386),而原型cd89/iga非阻断小鼠抗人cd89抗体克隆a59和a3分别识别人cd89的ec2结构域内(morton j exp med 1999;189:1715-1722)和人cd89的ec1-ec2结构域的边界内(morton j exp med 1999;189:1715-1722)的表位。
[0305]
出人意料地,纯化的熟知cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)小鼠抗人cd89抗体克隆a59和克隆a3(在10μg/ml下检查时)显示非聚集和热聚血清人iga两者与膜人cd89的结合的部分但显著(≈60%)阻断(参见以上实施例2(b))。因此,通过使用facs分析,更详细地研究了小鼠抗人cd89抗体克隆a59和克隆a3在空间上阻碍血清人iga与膜结合人cd89的相互作用。
[0306]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa (sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)在4℃下孵育30分钟。并行地,50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠igg1同种型对照(bd biosciences)和50μl纯化的(在pbs/bsa/nan3中)小鼠抗人cd89抗体克隆mip8a(biorad)分别作为阴性对照和阳性对照运行。在此之后(即,不洗涤),将50μl的20μg/ml纯化的非聚集或热聚人(血清来源的)iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定非聚集或热聚血清人iga在膜人cd89上的结合30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0307]
如图6a和6b所示,小鼠抗人cd89特异性抗体克隆59和a3两者均剂量依赖性地且部分地抑制(即,≈60%最大抑制)非聚集和热聚血清人iga与膜人cd89的结合,而其相应的小鼠igg1同种型对照对非聚集和热聚血清人iga与膜人cd89的结合没有任何影响,从而证明小鼠抗人cd89特异性抗体59和a3对cd89/iga相互作用的抑制作用是特异性的。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且显示对非聚集和热聚血清人iga与膜人cd89的结合的剂量依赖性和完全阻止。
[0308]
这些结果证明小鼠抗人cd89特异性抗体克隆59和a3(分别识别人cd89的ec2结构域内(morton j exp med 1999;189:1715-1722)和人cd89的ec1-ec2结构域的边界内(morton j exp med 1999;189:1715-1722)的表位的原型抗体)能够特异性地且部分地抑制单体、二聚体、三聚体、四聚体或更高级多聚体血清人iga(即非聚集和热聚iga)与膜人cd89的结合。最可能地,小鼠抗人cd89特异性抗体克隆59和a3可在与ec2结构域或ec1-ec2边界结合之后以使血清人iga与人cd89的结合变得不那么“有利”或最佳(与识别人cd89上
的iga结合位点(即,ec1结构域)的抗人cd89抗体的空间位阻相反)的这样一种方式改变膜人cd89的折叠。
[0309]
实施例3.使用离体的表达人cd89的原代人嗜中性粒细胞进行cd89/iga阻断小鼠抗人cd89单克隆抗体的生物学表征
[0310]
(a).cd89/iga阻断小鼠抗人cd89抗体在表达人cd89的原代人嗜中性粒细胞上的结合
[0311]
为了确定纯化的cd89/iga阻断小鼠抗人cd89抗体在表达人cd89的原代人嗜中性粒细胞上的结合,使用了facs分析。
[0312]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞-出于比较目的,并行检查了稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))和表达人cd89的单核细胞u937细胞系(来自dr.rt urbanus,department of haematology,university medical centre utrecht,nl的慷慨馈赠)。在pbs中洗涤后,将粒细胞在4℃下以10x106个细胞/ml置于含有0.1% bsa的冰冷冻了的pbs(sigma-aldrich;pbs/bsa)中10分钟,所述pbs补充有50μg/ml人igg(阻断可能的fcγ受体;sigma-aldrich)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与100μl的10μl/ml纯化的小鼠抗人cd89抗体(在pbs/bsa中)在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的小鼠igg1同种型对照(biolegend)(在pbs/bsa中)作为阴性对照运行,并且100μl的10μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(biorad)(在pbs/bsa中)、10μg/ml的克隆a59(bd biosciences)(在pbs/bsa中)和10μg/ml的克隆a3(santa cruz biotechnology)(在pbs/bsa中)作为阳性对照运行。在pbs/bsa中充分洗涤后,随后将细胞与1:200稀释的pe缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)在4℃下孵育30分钟。在pbs/bsa中充分洗涤后,将细胞在4℃下在pbs/bsa中的2%甲醛中固定30分钟。使用流式细胞仪(cyan;beckmancoulter)测量抗体在来自离体人嗜中性粒细胞的膜人cd89上的结合(几何平均荧光强度)。
[0313]
如图7a所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(在10μg/ml下)结合至来自从多个供体(n=5)分离的离体人嗜中性粒细胞的膜人cd89。基于它们在中性粒细胞上的结合概况,发现了以下排序(从高至较低结合强度):9h7=26d6(=mip8a)》10e7=30c7=20b4(=a59=a3)》8f3=16d6,这与这些纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(在10μg/ml下)与来自hek293f细胞的膜全长人cd89的结合排序(参见图7b)即8f3=9h7=10e7=26d6=20b4=30c7(=mip8a=a59=a3)》16d6出人意料地显著不同。此外,纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(在10μg/ml下)与来自单核细胞u937细胞的膜人cd89的结合排序(参见图7c),即9h7=26d6(=mip8a)》10e7=30c7(=a59=a3)》8f3=16d6=20b4似乎与纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(在10μg/ml下)与来自离体人嗜中性粒细胞的膜人cd89的前述结合排序相似(除20b4外)。
[0314]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7识别来自离体人嗜中性粒细胞的膜人cd89上、来自hek293f细胞的膜人全长cd89上和来自单核细胞u937细胞的膜人cd89上的表位。然而,当离体表达人cd89的人嗜中性粒细胞(从高至较低结合强度;9h7=26d6(=mip8a)》10e7=30c7=20b4(=a59=a3)》8f3=16d6)对比表达人全长cd89的hek293f细胞(从高至较低结合强度;8f3=9h7=10e7=
26d6=20b4=30c7(=mip8a=a59=a3)》16d6)在此比较研究中用作靶细胞时,这些cd89/iga阻断小鼠抗人cd89特异性抗体的结合排序显著不同,这表明这些细胞上多种膜人cd89同种型的可能(缺乏)识别。事实上,除了全长cd89(也称为fcαria.1)外,已知原代人嗜中性粒细胞和单核细胞表达人cd89的两种选择性剪接变体转录物(patry等人j immunol 1996;156:4442-4448;pleass等人biochem j 1996;318:771-777;togo等人febs letters 2003;535:20-209),所述转录物缺少(1)人cd89的ec2结构域(gly195-thr216;swiss-prot号p24071.2)的一部分(称为fcαria.2或δ66ec2),或(2)人cd89的整个ec2结构域(gly121-thr216;swiss-prot号p24071.3)(称为fcαria.3或δec2)。由于在10μg/ml下小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7与hek293f细胞(仅表达全长人cd89型式)结合的程度相似(除低亲和力抗体16d6外;也参见以上实施例2(a)),而在10μg/ml下小鼠抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7(如分别识别人cd89的ec2结构域内(morton j exp med 1999;189:1715-1722)和人cd89的ec1-ec2结构域的边界内(morton j exp med 1999;189:1715-1722)的表位的人iga非阻断剂克隆a59和a3)的结合显著低于在10μg/ml下小鼠抗人cd89特异性抗体9h7和26d6(如识别人cd89的ec1结构域内的表位的人iga阻断剂mip8a(lu等人protein sci 2014;23:378-386))与原代人嗜中性粒细胞(表达全长人cd89、δec2和在较小程度上表达δ66ec2型式)的结合,所以表明(1)小鼠抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7识别人cd89的ec2结构或ec1-ec2结构域的边界内的表位(如人iga非阻断剂克隆a59和a3,它们不能与选择性剪接变体fcαria.3结合,因为它缺乏ec2结构域)或识别ec1结构域内的表位,所述表位在人cd89的选择性剪接变体(即,fcαria.2和/或fcαria.3)中不可接近,这是由于这些选择性剪接变体的“异常”蛋白质折叠(与全长人类cd89/fcαria.1的“正常”蛋白质折叠相反),以及(2)小鼠抗人cd89特异性抗体9h7和26d6识别人cd89的ec1结构域内的表位(如人iga阻断剂mip8a)。令人感兴趣地,选择性剪接变体fcαria.3(或δec2)缺乏与血清来源的人iga的结合,尽管在此剪接变体上存在整个ec1结构域(即,人cd89上的iga结合位点),这指示了这种选择性剪接变体fcαria.3的“异常”蛋白质折叠(与全长人cd89/fcαria.1的“正常”蛋白质折叠相反)。此外,人全长cd89可能形成两种不同的构象,即,由于由内向外信号传导,人cd89的非活性状态对比活性状态(brandsma等人immunol rev 2015,268:74-87)。因此,非活性人cd89显示对人iga的低亲和力结合,而活性人cd89显示对人iga的高亲和力结合(bracke等人blood 2001;97:3478-3483)。因此,全长人cd89的构象变化/状态(由内向外信号传导)也可导致我们的所产生的cd89/iga阻断小鼠抗人cd89特异性抗体与离体人嗜中性粒细胞上的全长人cd89的差异结合。
[0315]
与此一致,10μg/ml的小鼠抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7(如分别识别人cd89的ec2结构域内(morton j exp med 1999;189:1715-1722)和人cd89的ec1-ec2结构域的边界内(morton j exp med 1999;189:1715-1722)的表位的非阻断剂克隆a59和a3)的结合显著低于10μg/ml的小鼠抗人cd89特异性抗体9h7和26d6(如识别人cd89的ec1结构域内的表位的阻断剂mip8a(lu等人protein sci 2014;23:378-386))与单核细胞u937细胞的结合,所述单核细胞u937细胞也已知表达全长人cd89、δec2和δ66ec2型式(patry等人j immunol 1996;156:4442-4448;togo等人febs letters 2003;535:20-209)。
[0316]
这些结果还证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、
26d6、20b4和30c7识别来自离体人嗜中性粒细胞的膜人cd89上的非多态表位,因为这些抗体明确地与从所有5个经检查的供体中分离的人嗜中性粒细胞结合。
[0317]
(b).使用cd89/iga阻断小鼠抗人cd89单克隆抗体阻断血清人iga介导的表达人cd89的原代人嗜中性粒细胞的吞噬作用
[0318]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断小鼠抗人cd89抗体抑制表达人cd89的原代人嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的能力。
[0319]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。然后,将100μl/孔(即,在96孔平底板中0.2x106个细胞;greiner)的这些细胞与滴定的(在rpmi/10% fcs中)纯化的小鼠抗人cd89抗体在4℃下孵育20分钟。并行地,滴定的(在rpmi/10%fcs中)纯化的小鼠igg1同种型对照(biolegend)作为阴性对照运行,并且滴定的(在rpmi/1% fcs中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)、克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)作为对照运行。在此之后(即,不洗涤),将1.2μl细胞与珠粒比例为1:60的纯化的人(血清来源的)iga(mp biomedicals)包被的荧光乳胶珠(1μm大小和羧酸酯改性的聚苯乙烯;sigma-aldrich)添加至这些细胞中(制剂iga珠,参见aleyd等人j immunol 2014;192:2374-2383),并在37℃下再孵育30分钟。并行地,细胞与珠粒比例为1:60的bsa(sigma-aldrich)包被的荧光乳胶珠作为阴性对照运行(制剂bsa-珠,参见aleyd等人j immunol 2014;192:2374-2383)。在rpmi/10% fcs中洗涤并重悬于pbs/0.1% bsa(sigma-aldrich)中后,使用流式细胞仪(cyan;beckman coulter)测量血清人iga介导的离体人嗜中性粒细胞上的膜人cd89对荧光乳胶珠的吞噬作用(根据aleyd等人j immunol 2014;192:2374-2383的研究,使用几何平均荧光强度来计算吞噬指数)。
[0320]
如图8所示,所有纯化的cd89/iga阻断小鼠抗人cd89特异性抗体剂量依赖性地抑制了分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞中血清人iga介导的吞噬作用,尽管观察到一些供体间变化。基于其血清人iga介导的吞噬作用抑制的程度,发现以下排序(从强至较低的iga介导的吞噬作用抑制程度):8f3=9h7=10e7=26d6=30c7(=mip8a)》20b4》16d16。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞中血清人iga介导的吞噬作用的剂量依赖性抑制,尽管观察到一些供体间变化。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示对分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞中血清人iga介导的吞噬作用无抑制、几乎没有抑制或弱抑制。
[0321]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7抑制了血清人iga介导的表达膜人cd89的离体原代人嗜中性粒细胞的吞噬活性。关于总结,参见表2。
[0322]
表2.纯化的cd89/iga阻断小鼠抗人cd89特异性抗体对表达人cd89的离体原代人
嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的阻断作用。
[0323]
小鼠抗人cd89抗体对iga包被的乳胶珠的吞噬作用的阻断
[0324][0325]-=没有小鼠抗cd89抗体对经由膜人cd89的iga包被的乳胶珠的吞噬作用的阻断, =小鼠抗cd89抗体对经由膜人cd89的iga包被的乳胶珠吞噬作用的弱阻断, =小鼠抗cd89抗体对经由膜人cd89的iga包被的乳胶珠吞噬作用的中等阻断, =小鼠抗cd89抗体对经由膜人cd89的iga包被的乳胶珠吞噬作用的强阻断。
[0326]
(c).使用cd89/iga阻断小鼠抗人cd89单克隆抗体阻断血清人iga介导的表达人cd89的原代人嗜中性粒细胞的迁移、血清人iga介导的表达人cd89的原代人嗜中性粒细胞的趋化性以及血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的白三烯b4产生
[0327]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断小鼠抗人cd89抗体抑制(1)血清人iga介导的表达人cd89的原代人嗜中性粒细胞的迁移、(2)血清人iga介导的表达人cd89的原代人嗜中性粒细胞的趋化性和(3)血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的嗜中性粒细胞化学引诱物白三烯b4(ltb4)产生的能力。
[0328]
二维(2-d)迁移测定:使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃
下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以2.5x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中,且随后将100μl钙黄绿素-am标记的粒细胞(即,在96孔平底板中0.25x106个细胞/孔;greiner)与20μg/ml(在rpmi/10% fcs中)纯化的小鼠抗人cd89抗体在4℃下孵育20分钟。并行地,20μg/ml(在rpmi/10% fcs中)纯化的小鼠igg1同种型对照(biolegend)作为阴性对照运行,并且20μg/ml(在rpmi/10% fcs中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)、克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)作为对照运行。在此之后,每孔添加150μl rpmi/10% fcs,并且将细胞静置10分钟以再次变成单层。在此之后(即,不洗涤),将10μl纯化的人(血清来源的)iga(mp biomedicals)包被的琼脂糖4b珠(90μm大小且溴化氰激活的;ge healthcare)轻轻地添加至这些单层细胞(制剂3μg/ml iga珠,参见van der steen等人gastroentorol 2009;137:2018-2029),并在37℃下再孵育40分钟。并行地,bsa(sigma-aldrich)包被的琼脂糖4b珠作为阴性对照运行(制剂3μg/ml bsa珠,参见van der steen等人gastroentorol 2009;137:2018-2029)。然后,收集上清液并用于趋化性测定和lbt4 elisa(参见下文),并且洗涤琼脂糖珠以除去非结合/非迁移的钙黄绿素-am标记的粒细胞。随后,将粒细胞在0.2%(w/v)十六烷基三甲基溴化铵(sigma-aldrich)缓冲液中在室温下裂解30分钟,并且使用荧光计(fluostar/polarstar;bmg labtech)测量96孔平底板(greiner)中释放的钙黄绿素-am(反映iga结合/迁移的粒细胞的数量)。通过使用已知数量的裂解的钙黄绿素-am标记的粒细胞的标准曲线(即,0-0.3x106个细胞/孔)来定量iga结合/迁移的离体人嗜中性粒细胞的数量。
[0329]
趋化性测定:使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以1.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。为了测量趋化性,用29μl来自iga包被的琼脂糖珠刺激的原代人嗜中性粒细胞(即,来自另一个健康供体,参见上文)的上清液填充博伊登室(neuro probe)的下部隔室的孔。并行地,仅rpmi/10% fcs培养基和纯化的1或10nm ltb4(在rpmi中;sigma-aldrich)分别作为阴性对照和阳性对照运行。随后,用3μm孔径的聚乙烯吡罗烷酮包被的聚碳酸酯过滤器(neuro probe)覆盖下部隔室,然后将上部隔室组装到博伊登室的下部隔室上。在此之后,在上部隔室的孔中添加50μl钙黄绿素-am标记的粒细胞(即,0.05x106个细胞/孔)。在37℃下孵育40分钟后,测定离体人嗜中性粒细胞从上部隔室朝向下部隔室的孔的趋化性。为此,将下部隔室中的粒细胞在0.1%(w/v)十六烷基三甲基溴化铵(sigma-aldrich)缓冲液中在室温下裂解30分钟,并且使用荧光计(fluostar/polarstar;bmg labtech)测量96孔平底板(greiner)中释放的钙黄绿素am(反映趋化粒细胞的数量)。通过使用已知数量的裂解的钙黄绿素-am标记的粒细胞的标准曲线(即,0-0.05x106个细胞/孔)来定量iga包被的琼脂糖珠诱导的趋化离体人嗜中性粒细胞的数量。
[0330]
ltb4 elisa:测量来自iga包被的琼脂糖珠刺激的离体人嗜中性粒细胞的上清液中的ltb4水平(参见上文)。为此,根据制造商的说明书使用了市售ltb4竞争性elisa试剂盒(r&d systems)。
[0331]
如图9a所示,20μg/ml的所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的迁移,尽管观察到一些供体间变化。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的迁移的抑制,尽管观察到一些供体间变化。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示对血清人iga包被的琼脂糖珠介导的表达膜人cd89的离体人原代嗜中性粒细胞的迁移的高度可变(即,无、弱或中等)抑制。
[0332]
如图9b所示,20μg/ml的所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的趋化性,尽管观察到一些供体间变化。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的趋化性的抑制,尽管观察到一些供体间变化。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3
–
两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示对血清人iga包被的琼脂糖珠介导的表达膜人cd89的离体人原代嗜中性粒细胞的趋化性的高度可变(即,无、中等或强)抑制。
[0333]
如图9c所示,20μg/ml的所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4产生,尽管观察到一些供体间变化。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4产生的抑制,尽管观察到一些供体间变化。出人意料地,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-也显示对血清人iga包被的琼脂糖珠介导的表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4产生的抑制。
[0334]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7抑制了血清人iga介导的表达膜人cd89的离体原代人嗜中性粒细胞的迁移、趋化性和来自表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4释放。关于总结,参见表3和4。
[0335]
表3.纯化的cd89/iga阻断小鼠抗人cd89特异性抗体对血清人iga包被的琼脂糖珠诱导的表达人cd89的离体原代人嗜中性粒细胞的2-d迁移的阻断作用
[0336]
通过小鼠抗人cd89抗体阻断iga包被的琼脂糖珠诱导的2-d迁移
[0337][0338]-=,没有小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的2-d迁移的阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的2-d迁移的弱阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的2-d迁移的中等阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的2-d迁移的强阻断。nd=未测定。
[0339]
表4.纯化的cd89/iga阻断小鼠抗人cd89特异性抗体对血清人iga包被的琼脂糖珠诱导的(1)表达人cd89的离体原代人嗜中性粒细胞的趋化性和(2)通过表达人cd89的离体原代人嗜中性粒细胞的ltb4产生的阻断作用。
[0340]
通过小鼠抗人cd89抗体阻断iga包被的琼脂糖珠诱导的趋化性和ltb4产生
[0341][0342]-=没有小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的趋化性和ltb4产生的阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的趋化性和ltb4产生的弱阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的趋化性和ltb4产生的中等阻断, =小鼠抗cd89抗体对iga包被的琼脂糖珠诱导的经由膜人cd89的趋化性和ltb4产生的强阻断。nd=未测定。
[0343]
(d).使用cd89/iga阻断小鼠抗人cd89单克隆抗体阻断表达人cd89的原代人嗜中性粒细胞上的血清人iga结合和血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的乳铁蛋白产生
[0344]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断小鼠抗人cd89抗体(1)在空间上阻碍血清人iga与表达人cd89的原代人嗜中性粒细胞的相互作用和(2)抑制血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的乳铁蛋白产生的能力。
[0345]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代
人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中,且随后将100μl钙黄绿素-am标记的粒细胞(即,0.2x106个细胞/孔)与滴定的(在rpmi/10% fcs中)纯化的小鼠抗人cd89抗体在4℃下孵育20分钟。并行地,滴定的(在rpmi/10% fcs中)纯化的小鼠igg1同种型对照(biolegend)作为阴性对照运行,并且滴定的(在rpmi/10% fcs中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)、克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)作为对照运行。在此之后(即,不洗涤),将100μl的这些细胞(即,0.2x106个细胞/孔)添加至96孔平底elisa板(nunc-immuno maxisorp),所述板先前用100μl/孔10μg/ml的纯化的人(血清来源的)iga(mp biomedicals)或100μl/孔10μg/ml的bsa(用作阴性对照;sigma-aldrich)包被。在37℃下孵育30分钟后,收获上清液(180μl/孔)以除去非结合的粒细胞,并使用这些上清液(在若干离心清除步骤后)来测量乳铁蛋白生产水平(用作脱粒标志物;参见下文)。在洗涤板后,将粒细胞在0.2%(w/v)十六烷基三甲基溴化铵(sigma-aldrich)缓冲液中在室温下裂解30分钟,并且使用荧光计(fluostar/polarstar;bmg labtech)测量96孔平底板(greiner)中释放的钙黄绿素-am(反映iga结合的粒细胞的数量)。通过使用已知数量的裂解的钙黄绿素-am标记的粒细胞的标准曲线(即,0-0.3x106个细胞/孔)来定量iga结合的离体人嗜中性粒细胞的数量。
[0346]
测量用板结合的血清人iga刺激的原代人嗜中性粒细胞的上清液中的乳铁蛋白产生(代表脱粒程度)(参见上文)。为此,在4℃-8℃下将96孔平底elisa板(nunc-immuno maxisorp)用100μl/孔兔抗人乳铁蛋白抗体(1:5000;sigma-aldrich)包被16-24小时。在pbs/0.05% tween 20中充分洗涤后,在室温下将板用200μl/孔pbs/0.05% tween 20/0.5% bsa (sigma-aldrich)封闭1小时。然后将板与100μl/孔1:2稀释度的上清液(在封闭缓冲液中)在37℃孵育1小时。在pbs/0.05% tween 20中充分洗涤后,将板与碱性磷酸酶标记的兔抗人乳铁蛋白检测抗体(1:2500;mp biomedicals)在37℃下孵育1小时。在添加磷酸对硝基苯酯(sigma-aldrich)后,使用微板读数器(imark;biorad)在450nm波长下测量光密度。纯化的人乳铁蛋白(sigma-aldrich)用作标准物,以计算由血清人iga刺激的离体人嗜中性粒细胞释放的乳铁蛋白的量。
[0347]
如图10a所示,所有我们纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(除16d6外)都剂量依赖性地抑制了分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞与血清人iga的结合,尽管观察到一些供体间变化。基于它们的抑制程度,发现以下排序(从强至较低抑制程度):9h7=10e7=26d6=30c7(=mip8a)》8f3》20b4。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞与血清人iga的结合的剂量依赖性抑制,尽管观察到一些供体间变化。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示对分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞与血清人iga的结合无抑制。
[0348]
如图10b所示,所有我们纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(除16d6外)都剂量依赖性地抑制了血清人iga介导的分离自三个健康个体的表达人cd89的离体原
代人嗜中性粒细胞的乳铁蛋白产生,尽管观察到一些供体间变化。基于它们的抑制程度,发现以下排序(从强至较低抑制程度):9h7=10e7=26d6=30c7(=mip8a)》8f3》20b4。令人感兴趣地,这些经检查的纯化的小鼠抗人cd89抗体抑制血清人iga介导的人嗜中性粒细胞的乳铁蛋白产生的程度反映了其相应的对表达人cd89的人嗜中性粒细胞与血清人iga的结合的抑制程度(将图10b与图10a进行比较)。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))并行运行,并且也显示对iga介导的分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生的剂量依赖性抑制,尽管观察到一些供体间变化。相比之下,纯化的小鼠抗人cd89抗体克隆a59和克隆a3-两者均是熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770)-显示对iga介导的分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生无抑制。
[0349]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、26d6、20b4和30c7抑制了表达人cd89的离体原代人嗜中性粒细胞与血清人iga的结合,以及其相应的iga介导的乳铁蛋白产生(脱粒标志物)。关于总结,参见表5。
[0350]
表5.纯化的cd89/iga阻断小鼠抗人cd89特异性抗体对(1)表达人cd89的离体原代人嗜中性粒细胞与iga包被的板的结合和(2)其相应的iga介导的乳铁蛋白产生的阻断作用。
[0351]
通过小鼠抗人cd89抗体阻断iga结合和相应的iga介导的乳铁蛋白产生
[0352]
[0353][0354]-=没有小鼠抗cd89抗体对表达人cd89的粒细胞与iga包被的板的结合及其相应的iga介导的乳铁蛋白产生的阻断, =小鼠抗cd89抗体对表达人cd89的粒细胞与iga包被的板的结合及其相应的iga介导的乳铁蛋白产生的弱阻断, =小鼠抗cd89抗体对表达人cd89的粒细胞与iga包被的板的结合及其相应的iga介导的乳铁蛋白产生的中等阻断, =小鼠抗cd89抗体对表达人cd89的粒细胞与iga包被的板的结合及其相应的iga介导的乳铁蛋白产生的强阻断。
[0355]
(e).使用cd89/iga阻断小鼠抗人cd89单克隆抗体阻断血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的嗜中性粒细胞细胞外诱捕网(net)释放
[0356]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断小鼠抗人cd89抗体抑制血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的net释放的能力。
[0357]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从多个健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞以0.5x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。然后,将1.0x105个细胞/200μl/孔(在96孔u底板中(greiner))与20μg/ml纯化的小鼠抗人cd89抗体在4℃下孵育20分钟。并行地,纯化的小鼠igg1同种型对照(biolegend)作为阴性对照运行。在此之后(即,不洗涤),将3.0μl细胞与珠粒比例为1:300的纯化的人(血清来源的)iga (mp biomedicals)包被的非荧光乳胶珠(0.9μm大小和羧酸酯改性的聚苯乙烯;sigma-aldrich)添加至这些细胞中(制剂iga珠,参见aleyd等人j immunol 2014;192:2374-2383),并在37℃下再孵育30分钟。并行地,细胞与珠粒比例为1:300的bsa (sigma-aldrich)包被的非荧光乳胶珠作为阴性对照运行(制剂bsa-珠,参见aleyd等人j immunol 2014;192:2374-2383)。在此之后,将细胞在补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中洗涤两次,且随后将这些细胞以1.0x105个细胞/200μl/孔转移至96孔平底黑色板(fluotrac
tm 200;greiner)并在37℃下再孵育3小时。当指示时,添加100μg/ml dna酶i。然后,通过添加2.5μg/ml绿(invitrogen)检查细胞外dna的释放。使用荧光计(fluostar/polarstar;bmg labtech)测量荧光强度。
[0358]
如图23所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体抑制了血清人iga介导的来自分离自八个健康个体的表达人cd89的离体原代人嗜中性粒细胞的net释放,尽管观察到一些供体间变化。
[0359]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、
26d6、20b4和30c7抑制血清人iga介导的来自表达人cd89的离体原代人嗜中性粒细胞的net释放。
[0360]
(f).使用cd89/iga阻断小鼠抗人cd89单克隆抗体诱导表达人cd89的原代人嗜中性粒细胞的细胞死亡
[0361]
cd89/iga阻断小鼠抗人cd89抗体克隆mip8a已被描述为诱导表达cd89的人嗜中性粒细胞中的人细胞死亡(wehrli等人j immunol 2014,193:5649-5659)。取决于炎症微环境,通过暴露于二价小鼠抗人cd89抗体克隆mip8a,诱导这些人嗜中性粒细胞中的半胱天冬酶依赖性(通常在凋亡过程中观察到)或半胱天冬酶独立性(非凋亡)细胞死亡。因此,使用非引发(non-primed)(以模拟非炎症条件)和lps引发(lps-primed)(以模拟炎症条件)的表达人cd89的原代人嗜中性粒细胞作为靶细胞,检查了我们产生的cd89/iga阻断小鼠抗人cd89抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7对人cd89介导的细胞死亡的影响。
[0362]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从多个健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。然后,将这些细胞在37℃下非引发或lps引发(100ng/ml来自大肠杆菌0111:b4的超纯lps;invivogen)5分钟。洗涤后,将这些粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中,且随后将250μl粒细胞(即,在96孔平底板中的0.5x106个细胞/孔;falcon)与10μg/ml纯化的小鼠抗人cd89抗体与10μg/ml交联山羊抗小鼠igg特异性抗体(southern biotech)的组合在37℃下孵育5小时。并行地,纯化的小鼠igg1同种型对照(biolegend)和纯化的小鼠抗人cd89抗体克隆mip8a(biorad)分别作为阴性对照和阳性对照运行。在含有0.1% bsa的冰冷冻了的pbs(sigma-aldrich;pbs/bsa)中充分洗涤后,将粒细胞用3μm红色荧光dna复染剂碘化丙啶在4℃下染色30-60分钟。在pbs/bsa中充分洗涤后,将细胞在4℃下在pbs/bsa中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur或fortessa;bd biosciences)测定离体人嗜中性粒细胞的细胞死亡百分比(基于碘化丙啶染色)。
[0363]
如图24所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体和商业cd89/iga阻断小鼠抗人cd89抗体克隆mip8a都未触发非引发的分离自三个健康个体的表达人cd89的离体原代人嗜中性粒细胞的细胞死亡。相比之下,商业cd89/iga阻断小鼠抗人cd89抗体克隆mip8a诱导了lps引发的表达人cd89的离体原代人嗜中性粒细胞中的显著细胞死亡,尽管观察到一些供体间变化,而所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体未触发这些lps引发的表达人cd89的离体原代人嗜中性粒细胞的细胞死亡。
[0364]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7在非炎症性(即,非引发后)和炎症性(即,lps引发后)条件下未诱导表达人cd89的离体原代人嗜中性粒细胞的细胞死亡。
[0365]
(g).使用cd89/iga阻断小鼠抗人cd89单克隆抗体10e7阻断实验性诱导的人iga介导的自身免疫性皮肤病症线性iga大疱病
[0366]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体10e7的生物活性,测定了所产生的cd89/iga阻断小鼠抗人cd89抗体10e7抑制实验性诱导的人iga介导的自身免疫性皮肤病症线性iga大疱病(labd)的能力。
[0367]
labd是与iga自身抗体相关的慢性皮肤病,其特征在于亚表皮水疱伴以表达cd89的嗜中性粒细胞为主导的致密炎症浸润物(van der steen等人j immunol 2012;189:1594-1601)。在labd患者中,通常在真皮-表皮交界处发现线性iga沉积,并且这些iga自身抗体主要针对跨膜半桥粒抗原bp180/胶原蛋白xvii(otten等人curr mol med 2014;14:69-95),其通过人cd89的交联诱导持续的嗜中性粒细胞募集。因此,表达cd89的嗜中性粒细胞的持续激活(即,通过活性氧物质和促炎性细胞因子产生)和浸润(即,通过局部化学引诱物ltb4释放)导致labd患者中的严重组织损伤和症状加重。
[0368]
体内labd小鼠模型:在缺乏小鼠cd89同源物的双重转基因人cd89/人iga(tg hucd89/huiga)小鼠中,人cd89表达、调控、与人iga的相互作用和功能模拟人的情况(van egmond等人,blood 1999;93:4387-4394)。这些tg hucd89/huiga(在雌性与雄性之间相等分布)小鼠在其循环小鼠嗜中性粒细胞上表达膜人cd89(van egmond等人,blood 1999;93:4387-4394),在第0、2、4、6、8、10、12天在右耳中皮下注射10μl的7mg/ml抗小鼠胶原蛋白xvii人iga(自身)抗体(prof.dr.m.van egmond,dept molecular cell biology and immunology,vumc,amsterdam,nl)或在左耳中皮下注射10μl pbs。在有或没有纯化的cd89/iga阻断小鼠抗人cd89抗体10e7处理的情况下监测注射部位处表达人cd89的小鼠嗜中性粒细胞流入。为此,在第7天和第11天腹膜内注射100μl的1.5mg/ml纯化的cd89/iga阻断小鼠抗人cd89抗体10e7(关于处理方案,参见图31a)。并行地,100μl的1.5mg/ml小鼠igg1同种型对照(biolegend)作为阴性对照运行。在第14天,处死小鼠并切除耳朵,并且随后在液氮中速冻。将这些耳组织标本冷冻切片(6μm)并在室温下在丙酮中固定10分钟。然后,将这些空气干燥的冷冻切片与1:400稀释的alexa488缀合的大鼠抗小鼠ly-6g(gr-1染色,一种嗜中性粒细胞标记物;ebioscience)在室温下孵育1小时。在pbs中洗涤后,使用1μg/ml的dapi(invitrogen)将细胞核在室温下复染色5分钟。使用具有以下设置的vectra polaris显微镜进行铺瓦式扫描(tile scanning)以获得整个耳朵的图像:dapi msi 0.43ms,fitc 81.70ms和20倍放大倍率。用imagej/fiji软件分析冷冻切片的gr-1染色。测量小鼠耳朵的总面积(μm2)和特异性gr-1染色的面积(μm2)。定量被计算为gr-1面积(μm2)除以总面积(μm2)。使用以下公式来确定gr-1/总面积比率:
[0369][0370]
如图31b所示,纯化的cd89/iga阻断小鼠抗人cd89特异性抗体10e7显著(p《0.05)抑制了注射部位处的抗小鼠胶原蛋白xvii人iga抗体诱导的表达人cd89的小鼠嗜中性粒细胞流入。
[0371]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体10e7抑制了tg hucd89/huiga小鼠中实验性诱导的体内人iga介导的表达人cd89的小鼠嗜中性粒细胞流入。
[0372]
实施例4.通过cd89/iga阻断小鼠抗人cd89单克隆抗体识别的人cd89表位和cd89结构域的表征
[0373]
(a).非标记的cd89/iga阻断小鼠抗人cd89抗体与pe缀合的商业小鼠抗cd89抗体克隆mip8a(cd89/iga阻断剂)、克隆a59(cd89/iga非阻断剂)和克隆a3(cd89/iga非阻断剂)对人cd89的交叉竞争
[0374]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的优良特异性,通过与纯化的小
鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111))、纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770))和纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770))交叉竞争,测定由所产生的cd89/iga阻断小鼠抗人cd89抗体识别的表位的位置。
[0375]
人cd89由短细胞质尾、跨膜区和两个细胞外(ec)ig样结构域组成。通过短的结构域间铰链区域,这两个ig样ec结构域以彼此成约90
°
的角度折叠(ding等人j biol chem 2003;278:27966-27970)。人iga在人cd89上的结合位点位于膜远端ig样ec1结构域中,并且不位于膜近端ig样ec2结构域中(wines等人j immunol 1999;162:2146-2153;morton j exp med 1999;189:1715-1722;lu等人protein sci 2014;23:378-386)。因此,普遍认为,与人cd89的ec1结构域结合的小鼠抗人cd89抗体可阻断人cd89上的人iga结合,而与人cd89的ec2结构域结合的小鼠抗人cd89抗体不能阻断人cd89上的人iga结合(morton等人arch immunol ther exp 2001;49:217-229;bakema等人immunol rev 2011;4:612-624)。更具体地说,原型cd89/iga阻断小鼠抗人cd89抗体克隆mip8a识别人cd89的ec1结构域内的表位(lu等人protein sci 2014;23:378-386),而原型cd89/iga非阻断小鼠抗人cd89抗体克隆a59和a3分别识别人cd89的ec2结构域内(morton j exp med 1999;189:1715-1722)和人cd89的ec1-ec2结构域的边界内(morton j exp med 1999;189:1715-1722)的表位。通过facs分析测定所产生的纯化的cd89/iga阻断小鼠抗人cd89抗体与熟知的cd89/iga阻断和非阻断小鼠抗人cd89抗体(即,克隆mip8a、克隆a59和克隆a3)在膜人cd89上竞争的能力。
[0376]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与100μl的10μl/ml纯化的小鼠抗人cd89抗体(在pbs/bsa/nan3中)在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在pbs/bsa/nan3中)作为阴性对照运行,并且100μl的10μg/ml纯化的小鼠抗人cd89抗体克隆mip8a(biorad)(在pbs/bsa/nan3中)、10μg/ml的克隆a59(bd biosciences)(在pbs/bsa/nan3中)和10μg/ml的克隆a3(santa cruz biotechnology)(在pbs/bsa/nan3中)作为阳性对照运行。在此之后(即,不洗涤),将5μl未稀释的pe缀合的小鼠抗人cd89抗体克隆mip8a(biorad)、克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)添加至这些细胞,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量pe缀合的小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3在膜人cd89上的结合(几何平均荧光强度)。
[0377]
如图11a所示,与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体9h7、26d6和30c7预孵育完全(》90%)阻断了商业pe缀合的cd89/iga阻断小鼠抗人cd89抗体克隆mip8a在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7、16d6和20b4预孵育未阻断(《25%)商业pe缀合的cd89/iga阻断小鼠抗人cd89抗体克隆mip8a在膜人cd89上的随后结合。为供参考,与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89抗体克隆mip8a预孵育也完全(》90%)阻断了其pe
缀合的抗体对应物在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的熟知cd89/iga非阻断小鼠抗人cd89抗体克隆a59和a3浓度预孵育未阻断(《25%)商业pe缀合的cd89/iga阻断小鼠抗人cd89抗体克隆mip8a在膜人cd89上的结合。
[0378]
如图11b所示,与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体9h7、26d6和20b4预孵育部分(≈55%-65%)阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a59在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7、16d6和30c7预孵育未阻断(《25%)商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a59在膜人cd89上的随后结合。为供参考,与10μg/ml的非标记的纯化的cd89/iga非阻断小鼠抗人cd89抗体克隆a59预孵育完全(》90%)阻断了其pe缀合的抗体对应物在膜人cd89上的随后结合。此外,与10μg/ml的非标记的纯化的熟知cd89/iga非阻断小鼠抗人cd89抗体克隆a3预孵育也完全(》90%)阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a59在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的熟知cd89/iga阻断小鼠抗人cd89抗体克隆mip8a预孵育部分阻断了(≈50%)商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a59在膜人cd89上的随后结合。
[0379]
如图11c所示,与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体9h7、26d6和20b4预孵育部分(≈55%-65%)阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a3在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7、16d6和30c7预孵育未阻断(《25%)商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a3在膜人cd89上的随后结合。为供参考,与10μg/ml的非标记的纯化的cd89/iga非阻断小鼠抗人cd89抗体克隆a3预孵育完全(》90%)阻断了其pe缀合的抗体对应物在膜人cd89上的随后结合。此外,与10μg/ml的非标记的纯化的熟知cd89/iga非阻断小鼠抗人cd89抗体克隆a59预孵育也完全(》90%)阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a3在膜人cd89上的随后结合,而与10μg/ml的非标记的纯化的熟知cd89/iga阻断小鼠抗人cd89抗体克隆mip8a预孵育部分阻断了(≈55%)商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a3在膜人cd89上的随后结合。
[0380]
为了分析上述交叉竞争的程度,还使用稳定的人全长cd89转染的hek293f细胞(克隆号2;参见以上实施例1(b)),进行了与滴定的(与仅使用10μg/ml相反)交叉竞争性非标记的cd89/iga阻断小鼠抗人cd89特异性抗体的预孵育,然后与pe缀合的小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3的孵育。
[0381]
如图12a所示,所有“mip8a”交叉竞争性的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,9h7、26d6和30c7)都剂量依赖性地阻断了商业pe缀合的cd89/iga阻断小鼠抗人cd89抗体克隆mip8a在膜人cd89上的结合。基于它们的“mip8a”交叉竞争概况,发现了以下排序(从强至较低的“mip8a-pe”阻断程度):9h7=26d6(=mip8a)》30c7。关于总结,参见表6。
[0382]
如图12b所示,所有“a59”交叉竞争性的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,9h7、26d6和20b4)都剂量依赖性地阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a59在膜人cd89上的结合。基于它们的“a59”交叉竞争概况,发现了以下排序(从强至较弱的“a59-pe”阻断程度):(a59=a3)》9h7=26d6=20b4(=mip8a)。关于总结,参见表6。
[0383]
如图12c所示,所有“a3”交叉竞争性的非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,9h7、26d6和20b4)都剂量依赖性地阻断了商业pe缀合的cd89/iga非阻断小鼠抗人cd89抗体克隆a3在膜人cd89上的结合。基于它们的“a3”交叉竞争概况,发现了以下排序(从强至较弱“a3-pe”阻断程度):(a59=a3)》9h7=26d6=20b4(=mip8a)。关于总结,参见表6。
[0384]
表6.非标记的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体与pe缀合的商业小鼠抗cd89抗体克隆mip8a(cd89/iga阻断剂)、克隆a59(cd89/iga非阻断剂)和克隆a3(cd89/iga非阻断剂)在膜人cd89上的交叉竞争概况交叉竞争
[0385][0386]-=没有阻断pe缀合的商业小鼠抗cd89抗体克隆mip8a(cd89/iga阻断剂)、克隆a59(cd89/iga非阻断剂)或克隆a3(cd89/iga非阻断剂)在膜人cd89上的结合, =对pe缀合的商业小鼠抗cd89抗体克隆mip8a(cd89/iga阻断剂)、克隆a59(cd89/iga非阻断剂)或克隆a3(cd89/iga非阻断剂)在膜人cd89上的结合的弱阻断, =对pe缀合的商业小鼠抗cd89抗体克隆mip8a(cd89/iga阻断剂)、克隆a59(cd89/iga非阻断剂)或克隆a3(cd89/iga非阻断剂)在膜人cd89上的结合的强阻断。
[0387]
这些结果证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7与人cd89表位结合,所述表位与由商业小鼠抗人cd89抗体克隆mip8a、克隆a59或克隆a3识别的人cd89表位不同,因为它们的交叉竞争概况与使用这些商业小鼠抗人cd89抗体获
得的交叉竞争概况显著不同。此外,这些结果证明,作为一个组的cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7和16d6(具有相同交叉竞争概况的三种抗体)对比作为一个组的cd89/iga阻断小鼠抗人cd89特异性抗体9h7和26b6(具有相同交叉竞争概况的两种抗体)对比cd89/iga阻断小鼠抗人cd89特异性抗体20b4对比cd89/iga阻断小鼠抗人cd89特异性抗体30c7与不相同的cd89表位结合,因为它们的交叉竞争概况显著不同。此外,这些结果证明,作为一个组的cd89/iga阻断小鼠抗人cd89特异性抗体9h7和26b6(具有相同交叉竞争概况的两种抗体)对比商业小鼠抗人cd89抗体克隆mip8a似乎结合至相似的cd89表位,因为它们的交叉竞争概况相同。这些结果还证明,cd89/iga阻断小鼠抗人cd89特异性抗体8f3、10e7和16d6似乎结合至相似的cd89表位,因为它们的交叉竞争概况是相同的。最后,这些结果表明,cd89/iga阻断小鼠抗人cd89特异性抗体9h7和26b6似乎结合至相似的cd89表位,因为它们的交叉竞争概况是相同的。
[0388]
(b).cd89/iga阻断小鼠抗人cd89抗体与膜全长人cd89和各种膜嵌合人cd89/牛fcγ2r构建体的结合(人cd89的ec1对比ec2结构域作图)
[0389]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的良好特异性,通过结构域作图确定由产生的cd89/iga阻断小鼠抗人cd89抗体识别的一个或多个表位的位置。通过facs分析确定cd89/iga阻断小鼠抗人cd89抗体与在hek293f细胞的表面上表达的人cd89的ec1结构域或ec2结构域结合的能力。
[0390]
在结构上,人cd89和牛fcγ2r高度同源,并且彼此密切相关(ravetch等人annu rev immunol 1991;9:457-492;zhang等人j immunol 1995;155:1534-1541)。因此,通过在这两种受体蛋白之间交换ig样ec1和ec2结构域,设计了嵌合人/牛受体(还参见图13a)。在功能上,人cd89和牛fcγ2r是完全不同的,在于人cd89结合人iga而不结合牛igg2,而牛fcγ2r结合牛igg2而不结合人iga。产生并瞬时表达了以下人cd89构建体:(1)膜全长人cd89构建体,所述构建体含有人cd89的ig样ec1和ig样ec2结构域二者(参见seq id no:1)并且因此称为“人ec1-ec2-cd89”,(2)人cd89的膜嵌合ig样ec1结构域与牛fcγ2r构建体的ig样ec2结构域组合(参见seq id no:3和seq id no:4,即分别与牛跨膜细胞内区或与人跨膜细胞内区组合)并且因此称为“人ec1-cd89”;和(3)牛fcγ2r的膜嵌合ig样ec1结构域与人cd89构建体的ig样ec2结构域组合(参见seq id no:7)并且因此称为“人ec2-cd89”。此外,还产生了膜全长牛fcγ2r构建体,所述构建体含有牛fcγ2r的ig样ec1和ig样ec2结构域两者(参见seq id no:9),并且因此称为“牛fcγ2r”。将编码上述“人ec1-ec2-d89”、“人ec1-cd89”、“人ec2-cd89”和“牛fcγ2r”构建体的cdna优化用于哺乳动物表达并由geneart,regensburg,germany合成(分别参见seq id no:2,seq id no:5、seq id no:6、seq id no:8和seq id no:10)。将这些cdna亚克隆到pcdna3.1来源的表达质粒中。
[0391]
使用freestyletm 293表达系统(invitrogen),将freestyl
etm 293f细胞(invitrogen)用“人cd89”、“人ec1-cd89”、“人ec2-cd89”和“牛fcγ2r”构建体瞬时转染。在48小时和/或72小时后,通过facs分析对cd89/iga阻断小鼠抗人cd89抗体与经转染的细胞上的上述嵌合人/牛受体的结合进行分析。为此,将瞬时转染的hek293f细胞在4℃下以10x106个细胞/ml置于冰冷冻了的磷酸盐缓冲盐水中10分钟,所述盐水含有0.1% bsa (sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与100μl的10μl/ml纯化的小鼠抗人cd89
抗体(在pbs/bsa/nan3中)在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的小鼠igg1同种型对照(bd biosciences)(在pbs/bsa/nan3中)作为阴性对照运行,并且100μl的10μg/ml(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)、克隆a59(bd biosciences)和克隆a3(santa cruz biotechnology)作为阳性对照运行。在pbs/bsa/nan3中充分洗涤后,随后将细胞在4℃下与1:200稀释的pe缀合的山羊抗小鼠igg fcγ特异性抗体(jackson immunoresearch)孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(型号facscalibur;bd biosciences)测量抗体的结合。
[0392]
除了上述cd89/iga阻断小鼠抗人cd89抗体与嵌合人/牛受体的结合外,还对这些瞬时转染的细胞检查了这些嵌合人/牛受体的(1)膜表面表达水平和(2)适当折叠。为此,将瞬时转染的hek293f细胞在4℃下以10x106个细胞/ml置于冰冷冻了的磷酸盐缓冲盐水中10分钟,所述盐水含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞(1)与或不与100μl的2.5μg/ml纯化的兔抗人cd89多克隆抗体(sino biological)(在pbs/bsa/nan3中),和(2)与或不与100μl的10μg/ml纯化的非聚集或热聚人(血清来源的;参见以上实施例2(b))iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)在4℃下孵育30分钟。在pbs/bsa/nan3中重复洗涤后,在4℃下分别用5μg/ml的1:200稀释的fitc缀合的f(ab')2片段山羊抗兔igg重/轻链特异性抗体(jackson immunoresearch)和生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定(1)兔抗人cd89多克隆抗体和(2)非聚集或热聚血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(型号facscalibur;bd biosciences)测量(1)兔抗人cd89多克隆抗体和(2)非聚集或热聚血清人iga在膜嵌合人/牛受体上的结合。
[0393]
如图13b所示,如通过兔抗人cd89多克隆抗体在这些细胞上的结合所证明,所有嵌合人cd89/牛fcγ2r受体(即,“人ec1-cd89”和“人ec2-cd89”的两种型式)和全长人cd89都在瞬时转染的细胞的膜表面上表达。此外,如通过非聚集和热聚血清人iga两者的结合所证明,这些膜表达的嵌合人cd89/牛fcγ2r受体(即,“人ec1-cd89”的两种型式)和全长人cd89似乎显示适当的蛋白质折叠。如所预期,缺少ec1结构域(即,人cd89上的iga结合位点)的嵌合人cd89/牛fcγ2r受体“人ec2-cd89”不与非聚集和热聚血清人iga结合。如所预期,兔抗人cd89多克隆抗体以及非聚集和热聚血清人iga不与模拟转染的(mock-transfected)细胞结合,也不与全长牛fcγ2r转染的细胞结合。
[0394]
如图13c所示,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,8f3、9h7、10e7、16d6、26d6、20b4和30c7)都显示与经转染的293f细胞上的“人ec1-cd89”的两种型式结合,而不与“人ec2-cd89”结合。此外,所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,8f3、9h7、10e7、16d6、26d6、20b4和30c7)都显示与全长人cd89“人ec1-ec2-cd89)转染的细胞结合,但显示与模拟转染的细胞和全长牛fcγ2r转染的细胞均无结合。如所预期,识别人cd89的ec1结构域内的表位(lu等人protein sci 2014;23:378-386)的纯化的熟知cd89/iga阻断小鼠抗人cd89抗体克隆mip8a显示与“人ec1-cd89”的两种型式
中重复洗涤后,在4℃下分别用5μg/ml的1:200稀释的fitc缀合的f(ab')2片段山羊抗兔igg重/轻链特异性抗体(jackson immunoresearch)和生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定(1)兔抗人cd89多克隆抗体和(2)非聚集或热聚血清人iga在膜食蟹猴cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(型号facscalibur;bd biosciences)测量(1)兔抗人cd89多克隆抗体和(2)非聚集或热聚血清人iga在膜食蟹猴cd89上的结合。
[0401]
如图14a所示,如通过交叉反应性兔抗人cd89多克隆抗体在这些细胞上的结合所证明,全长食蟹猴cd89在瞬时转染的细胞的膜表面上表达。此外,如通过交叉反应性非聚集和热聚血清人iga两者的结合所证明,这种膜表达的全长食蟹猴cd89似乎显示适当的蛋白质折叠。
[0402]
如图14b所示,纯化的cd89/iga阻断小鼠抗人cd89特异性抗体16d6和30c7显示针对经转染的293f细胞上的食蟹猴全长cd89的中等/弱交叉反应性。纯化的cd89/iga阻断小鼠抗人cd89抗体8f3、9h7、10e7、26d6和20b4未识别经转染的293f细胞上的食蟹猴全长cd89。商业纯化的小鼠抗人cd89特异性抗体克隆a59和克隆a3显示针对经转染的293f细胞上的食蟹猴全长cd89的强交叉反应性,而商业纯化的小鼠抗人cd89特异性抗体克隆mip8a未显示在食蟹猴全长cd89上的任何结合。
[0403]
这些结果证明,小鼠抗人cd89抗体16d6和30c7似乎识别最可能是食蟹猴全长cd89的ec1结构域中的线性和/或非线性/构象表位。
[0404]
全长食蟹猴cd89蛋白(即,met1-lys287;ncbi参考序列:xp_005590398.1)的预测氨基酸序列显示与全长人cd89蛋白(即,met1-lys287;swiss-prot号p24071.1)的氨基酸序列的86%同源性,并且食蟹猴cd89的细胞外区(即,gln22-asn227;ncbi参考序列:xp_005590398.1)的预测氨基酸序列显示与人cd89蛋白的细胞外区(即,gln22-asn227;swiss-prot号p24071.1)的氨基酸序列的83%同源性。更详细地,食蟹猴cd89蛋白的ec1结构域(即,gln22-gly121;ncbi参考序列:xp_005590398.1)、短铰链区(即,leu122-lys125)、ec2结构域(即,pro126-asn220)和膜近端“接头”区(即,arg221-asn227)的预测氨基酸序列分别显示与人cd89蛋白的其相应对应物的氨基酸序列的72%、100%、93%和85%同源性。
[0405]
考虑到我们产生的cd89/iga阻断小鼠抗人cd89抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7似乎识别人全长cd89的ec1结构域内的线性和/或非线性/构象表位(即,gln22-gly121;swiss-prot号p24071.1)(参见以上实施例4(b)),并不出人意料地,大多数的我们的cd89/iga阻断小鼠抗人cd89抗体(即,8f3、9h7、10e7、26d6和20b4)不与经转染的293f细胞上的全长食蟹猴cd89的低同源(即,72%氨基酸序列)ec1结构域(即,gln22-gly121;ncbi参考序列:xp_005590398.1)交叉反应。
[0406]
(d).cd89/iga阻断小鼠抗人cd89抗体与各种膜嵌合人cd89/食蟹猴cd89构建体的结合(人cd89的ec1结构域内的表位作图)
[0407]
为了分析纯化的cd89/iga阻断小鼠抗人cd89抗体的良好特异性,通过表位作图确定由产生的cd89/iga阻断小鼠抗人cd89抗体识别的一个或多个表位。通过facs分析确定cd89/iga阻断小鼠抗人cd89抗体与在hek293f细胞的表面上表达的人cd89的ec1结构域内
的表位结合的能力。
[0408]
在结构上,全长人cd89和全长食蟹猴cd89高度同源(即,氨基酸序列;也参见以上实施例4(c))并且彼此密切相关(rogers等人immunol 2004;113:178-186)。然而,全部识别人cd89的ec1结构域内的表位的我们所产生的cd89/iga阻断小鼠抗人cd89抗体(参见以上实施例4(b))显示与食蟹猴cd89的ec1结构域无(即,8f3、9h7、10e7、26d6和20b4)、弱(即,30c7)或中等(即,16d6)交叉物种反应性(参见以上实施例4(c))。因此,通过将来自人cd89的ec1结构域的部分(即,长度为25个氨基酸的多肽)与来自食蟹猴cd89的ec1结构域的相互对应物(还参见图15a)进行交换而设计了嵌合人cd89/食蟹猴cd89受体,以确定人cd89的ec1结构域内的关键区域,所述区域由我们所产生的cd89/iga阻断小鼠抗人cd89抗体识别。在功能上,人cd89和食蟹猴cd89是相似的,在于人cd89和食蟹猴cd89两者均结合血清人iga(参见以上实施例4(c))。产生并瞬时表达了以下人cd89构建体:(1)膜全长人cd89构建体,其含有人cd89的全长ec1结构域(参见seq id no:1)并且因此称为“人ec1-cd89”;(2)膜嵌合人cd89/食蟹猴cd89构建体(i),其由来自人cd89的ec1结构域的gln22-lys46替换为来自食蟹猴cd89的ec1结构域的相互gln22-arg46组成(参见seq id no:13)并且因此称为
‘
δgln22-lys46人ec1-cd89’;(3)膜嵌合人cd89/食蟹猴cd89构建体(ii),其由来自人cd89的ec1结构域的ile47-ile71替换为来自食蟹猴cd89的ec1结构域的相互ile47-arg71组成(参见seq id no:15)并且因此称为
‘
δile47-ile71人ec1-cd89’;(4)膜嵌合人cd89/食蟹猴cd89构建体(iii),其由来自人cd89的ec1结构域的gly72-gly96替换为来自食蟹猴cd89的ec1结构域的相互asp72-gly96组成(参见seq id no:17)并且因此称为
‘
δgly72-gly96人ec1-cd89’;和(5)膜嵌合人cd89/食蟹猴cd89构建体(iv),其由来自人cd89的ec1结构域的arg97-gly121替换为来自食蟹猴cd89的ec1结构域的相互arg97-gly121组成(参见seq id no:19)并且因此称为
‘
δarg97-gly121人ec1-cd89’。此外,膜全长食蟹猴cd89构建体,其含有人食蟹猴cd89的全长ec1结构域(参见seq id no:11)并且因此称为“食蟹猴ec1-cd89”。
[0409]
除了替换长度为25个氨基酸的连续多肽的ec1结构域部分(即,构建体i-iv,参见上文),接触人cd89上的氨基酸残基(热点)的人iga(根据herr等人,nature 2003;423:614-620;bakema等人immunol rev 2011;4:612-624;lu等人protein sci 2014;23:378-386)也被替换为来自食蟹猴cd89的ec1结构域的相互(reciprocal)氨基酸(还参见图15a)。为此,产生并瞬时表达了以下嵌合人cd89/食蟹猴cd89构建体:膜嵌合人cd89/食蟹猴cd89构建体,其由来自人cd89的ec1结构域的thr58、gln59、arg73、arg74、lys76、his106和tyr107替换为来自食蟹猴cd89的ec1结构域的相互ile58、trp59、glu73、lys74、gly76、leu106和ser107组成(参见seq id no:21)并且因此称为“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)。
[0410]
将编码上述“人ec1-cd89”、“δgln22-lys46人ec1-cd89”、“δile47-ile71人ec1-cd89”、“δgly72-gly96人ec1-cd89”、“δarg97-gly121人ec1-cd89”、“δthr58,gln59;δarg73;arg74;lys76;δhis106,tyr107人ec1-cd89”(热点)和“食蟹猴ec1-cd89”构建体的cdna优化用于哺乳动物表达并由geneart,regensburg,germany合成(分别参见seq id no:2、seq id no:14、seq id no:16、seq id no:18、seq id no:20、seq id no:22和seq id no:12)。将这些cdna亚克隆到pcdna3.1来源的表达质粒中。
[0411]
使用freestyletm 293表达系统(invitrogen),将freestyl
etm 293f细胞
cd89”以及全长人cd89和全长食蟹猴cd89结合至非聚集和热聚血清人iga两者。相比之下,膜表达的嵌合人cd89/食蟹猴cd89构建体“δile47-ile71人ec1-cd89”和“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)未显示与非聚集和热聚血清人iga的任何结合。
[0414]
这些结果证明,血清人iga结合至人cd89的ec1结构域内的氨基酸序列ile47-ile71(即,iqcqaireayltqlmiiknstyrei;参见seq id no:24),并且此ile47-ile71氨基酸序列内的氨基酸残基thr58和gln59似乎对这种血清人iga/人cd89相互作用至关重要,因为血清人iga显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体无结合。关于总结,参见表7。
[0415]
如图15c所示并且如所预期(参见以上实施例4(c)),所有我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体(即,8f3、9h7、10e7、16d6、26d6、20b4和30c7)显示与全长人cd89的强结合,但显示与全长食蟹猴cd89无(即,8f3、9h7、10e7、26d6和20b4)、弱(即,30c7)或中等(即,16d6)结合。如所预期(参见以上实施例4(c)),商业小鼠抗人cd89抗体克隆mip8a也显示与全长人cd89的强结合,但显示与全长食蟹猴cd89无结合。如所预期(参见以上实施例4(c)),商业小鼠抗人cd89抗体克隆a59和克隆a3显示与全长人cd89和全长食蟹猴cd89两者的强结合。关于总结,参见表7。
[0416]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体8f3显示与“δgln22-lys46人ec1-cd89”和“δarg97-gly121人ec1-cd89”构建体的强结合,但分别显示与“δile47-ile71人ec1-cd89”和“δgly72-gly96人ec1-cd89”构建体的中等或无结合,这表明小鼠抗人cd89抗体8f3识别人cd89的ec1结构域内的氨基酸序列ile47-ile71和gly72-gly96(即,分别iqcqaireayltqlmiiknstyrei和grrlkfwnetdpefvidhmdankag;参见seq id no:24和25)中的线性或非线性/构象表位。这些氨基酸序列ile47-ile71和gly72-gly96中的氨基酸残基thr58、gln59、arg73、arg74和lys76似乎对小鼠抗人cd89抗体8f3与人cd89的结合至关重要,因为小鼠抗人cd89抗体8f3显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体无结合。关于总结,参见表7。
[0417]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体9h7显示与“δgln22-lys46人ec1-cd89”、“δile47-ile71人ec1-cd89”和“δgly72-gly96人ec1-cd89”构建体的强结合,但显示与“δarg97-gly121人ec1-cd89”构建体的弱结合,这表明小鼠抗人cd89抗体9h7识别人cd89的ec1结构域内的氨基酸序列arg97-gly121(即,ryqcqyrighyrfrysdtlelvvtg;参见seq id no:26)中的线性或非线性/构象表位。此氨基酸序列arg97-gly121中的氨基酸残基his106和tyr107似乎对小鼠抗人cd89抗体9h7与人cd89的结合至关重要,因为小鼠抗人cd89抗体9h7显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的弱结合。关于总结,参见表7。
[0418]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体10e7显示与“δgln22-lys46人ec1-cd89”、“δile47-ile71人ec1-cd89”和“δarg97-gly121人ec1-cd89”构建体的强结合,但显示与“δgly72-gly96人ec1-cd89”构建体的弱结合,这表明小鼠抗人cd89抗体10e7识别人cd89的ec1结构域内的氨基酸序列gly72-gly96(即,grrlkfwnetdpefvidhmdankag;参见seq id no:25)中的线性或非线性/构象表位。此氨基酸序列gly72-gly96中的氨基酸残基arg73、arg74和lys76似乎对小鼠抗人cd89抗体10e7与人
cd89的结合至关重要,因为小鼠抗人cd89抗体10e7显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的无结合。关于总结,参见表7。
[0419]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体16d6显示与“δgln22-lys46人ec1-cd89”和“δarg97-gly121人ec1-cd89”构建体的强结合,但分别显示与“δile47-ile71人ec1-cd89”和“δgly72-gly96人ec1-cd89”构建体的弱或中等结合,这表明小鼠抗人cd89抗体16d6识别人cd89的ec1结构域内的氨基酸序列ile47-ile71和gly72-gly96(即,分别iqcqaireayltqlmiiknstyrei和grrlkfwnetdpefvidhmdankag;参见seq id no:24和25)中的线性或非线性/构象表位。这些氨基酸序列ile47-ile71和gly72-gly96中的氨基酸残基thr58、gln59、arg73、arg74和lys76似乎对小鼠抗人cd89抗体16d6与人cd89的结合至关重要,因为小鼠抗人cd89抗体16d6显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体无结合。关于总结,参见表7。
[0420]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体26d6显示与“δgln22-lys46人ec1-cd89”、“δile47-ile71人ec1-cd89”和“δgly72-gly96人ec1-cd89”构建体的强结合,但显示与“δarg97-gly121人ec1-cd89”构建体的弱结合,这表明小鼠抗人cd89抗体26d6识别人cd89的ec1结构域内的氨基酸序列arg97-gly121(即,ryqcqyrighyrfrysdtlelvvtg;参见seq id no:26)中的线性或非线性/构象表位。此氨基酸序列arg97-gly121中的氨基酸残基his106和tyr107似乎对小鼠抗人cd89抗体26d6与人cd89的结合至关重要,因为小鼠抗人cd89抗体26d6显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的中等结合。关于总结,参见表7。
[0421]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体20b4显示与“δgly72-gly96人ec1-cd89”和“δarg97-gly121人ec1-cd89”构建体的强结合,但分别显示与“δgln22-lys46人ec1-cd89”和“δile47-ile71人ec1-cd89”构建体的中等或无结合,这表明小鼠抗人cd89抗体20b4识别人cd89的ec1结构域内的氨基酸序列gln22-lys46和ile47-ile71(即,分别qegdfpmpfisaksspvipldgsvk和iqcqaireayltqlmiiknstyrei;参见seq id no:23和24)中的线性或非线性/构象表位。氨基酸序列ile47-ile71中的氨基酸残基thr58和gln59似乎对小鼠抗人cd89抗体20b4与人cd89的结合不重要,因为小鼠抗人cd89抗体20b4显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的强结合。关于总结,参见表7。
[0422]
如图15c所示,我们的纯化的cd89/iga阻断小鼠抗人cd89特异性抗体30c7显示与“δgln22-lys46人ec1-cd89”、“δgly72-gly96人ec1-cd89”和“δarg97-gly121人ec1-cd89”构建体的强结合,但显示与“δile47-ile71人ec1-cd89”构建体无结合,这表明小鼠抗人cd89抗体30c7识别人cd89的ec1结构域内的氨基酸序列ile47-ile71(即,对应地iqcqaireayltqlmiiknstyrei;参见seq id no:24)中的线性或非线性/构象表位。此氨基酸序列ile47-ile71中的氨基酸残基thr58和gln59似乎对小鼠抗人cd89抗体30c7与人cd89的结合不重要,因为小鼠抗人cd89抗体30c7显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的强结合。关于总结,参见表7。
[0423]
如图15c所示,纯化的熟知cd89/iga阻断小鼠抗人cd89抗体克隆mip8a显示与“δgln22-lys46人ec1-cd89”、“δile47-ile71人ec1-cd89”和“δgly72-gly96人ec1-cd89”构
建体的强结合,但显示与“δarg97-gly121人ec1-cd89”构建体无结合,这表明小鼠抗人cd89抗体克隆mip8a识别人cd89的ec1结构域内的氨基酸序列arg97-gly121(即,ryqcqyrighyrfrysdtlelvvtg;参见seq id no:26)中的线性或非线性/构象表位。此氨基酸序列arg97-gly121中的氨基酸残基his106和tyr107似乎对小鼠抗人cd89抗体克隆mip8a与人cd89的结合至关重要,因为小鼠抗人cd89抗体克隆mip8a显示与“δthr58,gln59;δarg73,arg74,lys76;δhis106,tyr107人ec1-cd89”(热点)构建体的中等结合。关于总结,参见表7。
[0424]
如图15c所示并且如所预期,纯化的熟知cd89/iga非阻断小鼠抗人cd89抗体克隆a59和克隆a3,两者均识别人类cd89的ec2结构域内的表位(参见以上实施例4(b)),显示与所有检查的嵌合人cd89/食虫猴cd89构建体的强结合,因为在这些构建体中的任一者中人ec2结构域没有改变。关于总结,参见表7。
[0425]
表7.纯化的cd89/iga阻断小鼠抗人cd89特异性抗体和血清人iga与hek细胞上的膜结合的嵌合人cd89/食蟹猴cd89构建体的结合。
[0426]
小鼠抗人cd89抗体和人iga与嵌合人cd89/食蟹猴cd89构建体的结合
[0427][0428]-=没有小鼠抗cd89抗体或人iga与膜嵌合人cd89/食蟹猴cd89构建体的结合,-/ =小鼠抗cd89抗体或人iga与膜嵌合人cd89/食蟹猴cd89构建体的弱结合, =小鼠抗cd89
抗体或人iga与膜嵌合人cd89/食蟹猴cd89构建体的中等结合, =小鼠抗cd89抗体或人iga与膜嵌合人cd89/食蟹猴cd89构建体的强结合。
[0429]
实施例5.cd89/iga阻断小鼠抗人cd89单克隆抗体的分子遗传学表征
[0430]
将产生cd89/iga阻断小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7的杂交瘤细胞用pbs洗涤,并等分在含有5x106个细胞的微量小瓶中,并且作为沉淀储存在-80℃下。通过使用rneasy mini分离试剂盒(qiagen),将这些细胞沉淀用于分离rna。测定rna浓度(a260 nm),并将rna储存在-80℃下。通过逆转录酶,使用revertaid
tm h minus第一链cdna合成试剂盒(fermentas)从2μg rna合成cdna,并储存在-80℃下。基于同种型小鼠igg1/κ,设计了如表8所示的引物来扩增小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7的可变(v)区。
[0431]
表8.用于扩增小鼠抗人cd89特异性抗体8f3、9h7、10e7、16d6、26d6、20b4和30c7的cdna的pcr引物。
[0432]
[0433][0434]
s=有义;as=反义;vl=可变轻链区;vh=可变重链区;ck=恒定κ(kappa)轻链区;ch=恒定igg1重链区;*根据bioceros bv内部编码系统进行编号;简并引物:k=g或t,s=g或c,r=a或g,m=a或c,w=a或t,y=c或t,h=a或c或t,并且n=任何碱基。
[0435]
引物385、386、387、389和391是设计用于与小鼠抗体轻链的信号肽退火的有义引物;引物394和605是与小鼠κ轻链的恒定区退火的反义引物。引物609是与小鼠vl区的小鼠框架1(fr1)退火的简并引物。引物405、407、409和412是与小鼠抗体重链的信号肽退火的有义引物;引物416是设计用于与重链igg1的恒定区退火的反义引物。使用表8中所示的引物组合进行各种pcr。将产生的pcr产物亚克隆到pcr
tm-blunt ii-载体中。随后,对克隆的插入物进行测序。
[0436]
从cd89/iga阻断小鼠抗人cd89特异性抗体8f3获得分别来自重链和轻链序列反应的总计4个和5个信息序列。基于此信息,确定了小鼠抗人cd89抗体8f3的vh和vl区的共有氨基酸序列,并分别在seq id no:27和28中列出。小鼠抗人cd89抗体8f3的vh和vl区的cdr的氨基酸序列分别在seq id no:29-31和32-34中列出。
[0437]
从cd89/iga阻断小鼠抗人cd89特异性抗体9h7获得来自重链和轻链序列反应两者的总计4个信息序列。基于此信息,确定了小鼠抗人cd89抗体9h7的vh和vl区的共有氨基酸序列,并分别在seq id no:35和36中列出。小鼠抗人cd89抗体9h7的vh和vl区的cdr的氨基酸序列分别在seq id no:37-39和40-42中列出。
[0438]
从cd89/iga阻断小鼠抗人cd89特异性抗体10e7获得来自重链和轻链序列反应两者的总计4个信息序列。基于此信息,确定了小鼠抗人cd89抗体10e7的vh和vl区的共有氨基酸序列,并分别在seq id no:43和44中列出。小鼠抗人cd89抗体10e7的vh和vl区的cdr的氨基酸序列分别在seq id no:45-47和48-50中列出。
[0439]
从cd89/iga阻断小鼠抗人cd89特异性抗体16d6获得来自重链和轻链序列反应两者的总计4个信息序列。基于此信息,确定了小鼠抗人cd89抗体16d6的vh和vl区的共有氨基酸序列,并分别在seq id no:51和52中列出。小鼠抗人cd89抗体16d6的vh和vl区的cdr的氨基酸序列分别在seq id no:53-55和56-58中列出。
[0440]
从cd89/iga阻断小鼠抗人cd89特异性抗体26d6获得重链和轻链序列反应两者的总计4个信息序列。基于此信息,确定了小鼠抗人cd89抗体26d6的vh和vl区的共有氨基酸序列,并分别在seq id no:59和60中列出。小鼠抗人cd89抗体26d6的vh和vl区的cdr的氨基酸序列分别在seq id no:61-63和64-66中列出。
[0441]
从cd89/iga阻断小鼠抗人cd89特异性抗体20b4获得分别来自重链和轻链序列反应的总计4个和6个信息序列。基于此信息,确定了小鼠抗人cd89抗体20b4的vh和vl区的共有氨基酸序列,并分别在seq id no:67和68中列出。小鼠抗人cd89抗体20b4的vh和vl区的cdr的氨基酸序列分别在seq id no:69-71和72-74中列出。
[0442]
从cd89/iga阻断小鼠抗人cd89特异性抗体30c7获得来自重链和轻链序列反应两者的总计4个信息序列。基于此信息,确定了小鼠抗人cd89抗体30c7的vh和vl区的共有氨基酸序列,并分别在seq id no:75和76中列出。小鼠抗人cd89抗体30c7的vh和vl区的cdr的氨基酸序列分别在seq id no:77-79和80-82中列出。
[0443]
实施例6.cd89/iga阻断嵌合小鼠/人igg4/κ(即,将小鼠恒定igg1/κ结构域替换成人恒定igg4/κ结构域)抗人cd89单克隆抗体的产生
[0444]
基于测定的cd89/iga阻断小鼠抗人cd89抗体的小鼠v区(参见以上实施例5),进行设计以产生cd89/iga阻断嵌合小鼠/人抗人cd89抗体型式。为此,在geneart(regensburg,germany)订购灰仓鼠优化的cdna序列,seq id no:83(编码嵌合小鼠/人重igg4链8f3)、seq id no:84(编码嵌合小鼠/人重igg4链10e7)、seq id no:85(编码嵌合小鼠/人重igg4链16d6)、seq id no:86(编码嵌合小鼠/人重igg4链20b4)和seq id no:87(编码嵌合小鼠/人重igg4链30c7)以及seq id seq id no:88(编码嵌合小鼠/人轻κ链8f3)、seq id no:89(编码嵌合小鼠/人轻κ链10e7)、seq id no:90(编码嵌合小鼠/人轻κ链16d6)、seq id no:91(编码嵌合小鼠/人轻κ链20b4)和seq id no:92(编码嵌合小鼠/人轻κ链30c7),所述cdna序列编码人信号肽,随后是与人稳定的(即,s239p;根据angal等人在mol.immunol.,第30卷,第1期,第105-108页,1993中)igg4恒定区连接的小鼠vh链,或随后是与人κ恒定区连接的小鼠vl链。使用合适的限制性酶,将产生的cdna亚克隆到pcdna3.1来源的表达质粒中。随后使用freestyle
tm 293表达系统(invitrogen),在293-f细胞(invitrogen)中瞬时表达嵌合抗体。使用常规亲和色谱法蛋白a柱,从上清液中纯化表达的cd89/iga阻断嵌合抗人cd89抗体。此后,使用lal显色终点测定法(hhycult biotech)测定lps水平,并且所有我们纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体(即,8f3、10e7、16d6、20b4和30c7)都含有《0.001eu lps/μg嵌合igg。
[0445]
对于嵌合氨基酸序列,参见seq id no:93(嵌合小鼠/人重igg4链8f3)、seq id no:94(嵌合小鼠/人重igg4链10e7)、seq id no:95(嵌合小鼠/人重igg4链16d6)、seq id no:96(嵌合小鼠/人重igg4链20b4)、seq id no:97(嵌合小鼠/人重igg4链30c7)、seq id no:98(嵌合小鼠/人轻κ链8f3)、seq id no:99(嵌合小鼠/人轻κ链10e7)、seq id no:100(嵌合小鼠/人轻κ链16d6)、seq id no:101(嵌合小鼠/人轻κ链20b4)和seq id no:102(嵌
合小鼠/人轻κ链30c7)。
[0446]
实施例7.cd89/iga阻断嵌合小鼠/人抗人cd89抗体的结合表征
[0447]
(a).cd89/iga阻断嵌合小鼠/人抗人cd89抗体对人cd89的相对结合亲和力
[0448]
为了确定纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体对人cd89的相对结合亲和力,使用了elisa和facs分析。
[0449]
elisa:在4℃-8℃下在16-24小时期间,使用半面积96孔eia板(corning),将rhucd89(sino biological)以0.5μg/ml包被在pbs中(25ng/50μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1% bsa(roche)封闭1小时。随后,将板与和不与50μl滴定的(在封闭缓冲液中)纯化的嵌合小鼠/人抗人cd89抗体/孔在室温下孵育1小时。在pbs/0.05% tween 20中充分洗涤后,在室温下用1:5000稀释的辣根过氧化物酶(hrp)缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)测定抗体在rhucd89上的结合,持续1小时,随后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m h2so4后,使用微板读数器(imark;biorad)在450nm波长(参考波长为655nm)下测量抗体在rhucd89上的结合(光密度)。为了进行比较,小鼠抗人cd89抗体对应物并行运行,并如实施例2(a)中所描述监测它们的结合。
[0450]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与100μl滴定的(在pbs/bsa/nan3中)纯化的嵌合小鼠/人抗人cd89抗体/管在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的人igg4同种型对照抗体(bristol-myers squibb)(在pbs/bsa/nan3中)作为阴性对照运行。在pbs/bsa/nan3中充分洗涤后,随后在4℃下将细胞与1:200稀释的pe缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量抗体在膜人cd89上的结合(几何平均荧光强度)。为了进行比较,小鼠抗人cd89抗体对应物并行运行,并如实施例2(a)中所描述监测它们的结合。
[0451]
如图16a所示,所有纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体都剂量依赖性地结合至rhucd89。基于它们的结合概况,发现了以下相对亲和力排序(从高至较低亲和力):20b4》8f3=10e7=30c7》16d6。用于比较和一致,它们的小鼠抗人cd89抗体对应物显示了相似的相对亲和力排名,即20b4》8f3=10e7=30c7》16d6。更具体地,嵌合小鼠/人抗人cd89抗体20b4、8f3、10e7、30c7和16d6分别导致9、31、16、22和108ng/ml的以下相对亲和力(即,半最大结合ec50),而相应的小鼠抗人cd89抗体20b4、8f3、10e7、30c7和16d6分别导致13、52、20、29和119ng/ml的相对亲和力,这表明嵌合小鼠/人抗人cd89抗体20b4、8f3、10e7、30c7和16d6针对rhucd89的结合亲和力在嵌合过程中似乎保持不变。
[0452]
如图16b所示,所有纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体都剂量依赖性地结合至膜人cd89。基于它们的结合概况,发现了以下相对亲和力排序(从高至较低亲和力):20b4》30c7》8f3=10e7》16d6。用于比较,它们的小鼠抗人cd89抗体对应物显示了略微不同的亲和力排序,即20b4》8f3=10e7=30c7》16d6。更具体地,嵌合小鼠/人抗人cd89抗体637b4、8f3、10e7、30c7和16d6分别导致296、1321、1251、615和1789ng/ml的以下相对亲
和力(即,半最大结合ec50),而相应的小鼠抗人cd89抗体20b4、8f3、10e7、30c7和16d6分别导致260、714、637、484和1622ng/ml的相对亲和力,这表明(1)嵌合小鼠/人抗人cd89抗体20b4、30c7和16d6针对膜结合cd89的结合亲和力在嵌合过程中似乎保持不变,和(2)嵌合小鼠/人小鼠抗人cd89抗体8f3和10e7针对膜结合cd89的结合亲和力在嵌合过程中似乎略微降低。
[0453]
(b).cd89/iga阻断嵌合小鼠/人抗人cd89抗体的cd89/iga阻断能力的程度
[0454]
为了分析纯化的嵌合小鼠/人抗人cd89抗体的cd89/iga阻断的程度,通过使用facs分析来测定纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体在空间上阻碍血清人iga与人cd89的相互作用的能力。
[0455]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与50μl滴定的(在pbs/bsa/nan3中)纯化的嵌合小鼠/人抗人cd89抗体/管在4℃下孵育30分钟。并行地,50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂(zhang等人clin exp immunol 2000;121:106-111),biorad)作为阳性对照运行。此外,50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a59(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);bd biosciences)(在pbs/bsa/nan3中)和50μl的20μg/ml纯化的小鼠抗人cd89抗体克隆a3(一种熟知的cd89/iga非阻断剂(monteiro等人j immunol 1992;148:1764-1770);santa cruz biotechnology)(在pbs/bsa/nan3中)作为阴性对照运行。在此之后(即,不洗涤),将50μl的20μg/ml纯化的非聚集或热聚((参见以上实施例2(b))人(血清来源的)iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清iga α链特异性抗体(jackson immunoresearch)测定血清人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的2%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量非聚集或热聚血清人iga在膜人cd89上的结合(几何平均荧光强度)。
[0456]
如图17a和17b所示,所有纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体都剂量依赖性地阻止了非聚集和热聚血清人iga与膜人cd89的结合。基于它们的cd89/iga阻断概况,发现了以下排序(从强至较低cd89/iga阻断程度):20b4》30c7》8f3=10e7》16d6。令人感兴趣地,在这些经检查的嵌合小鼠/人抗人cd89抗体在空间上阻断血清人iga与膜人cd89的结合的程度(此实施例)与它们对膜人cd89的相应相对结合亲和力(参见以上实施例7(a))之间似乎存在强正相关关系。为供参考,小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示血清人iga与膜人cd89的结合的剂量依赖性阻断。此外,小鼠抗人cd89特异性抗体克隆59和a3剂量依赖性地且部分地抑制了(即,≈60%最大抑制)非聚集和热聚血清人iga与膜人cd89的结合,这证实了我们之前使用这两种熟知的cd89/iga非阻断剂的结果(参见以上实
施例2(d))。
[0457]
为了分析纯化的嵌合小鼠/人抗人cd89抗体的cd89/iga阻断的程度,通过使用facs分析来测定纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体在空间上阻碍分泌型人iga与人cd89的相互作用的能力。
[0458]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))以10x106个细胞/ml置于冰冷冻了的pbs中,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与50μl滴定的(在pbs/bsa/nan3中)纯化的嵌合小鼠/人抗人cd89抗体/管在4℃下孵育30分钟。并行地,50μl滴定的(在pbs/bsa/nan3中)纯化的小鼠抗人cd89抗体克隆mip8a(biorad)作为阳性对照运行。在此之后(即,不洗涤),将50μl的0.20μm纯化的人(初乳来源的)iga(biorad)(在pbs/bsa/nan3中稀释)添加至这些孔中,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定分泌型人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的4%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量分泌型人iga在膜人cd89上的结合(几何平均荧光强度)。
[0459]
如图17c所示,所有纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体都剂量依赖性地阻止了分泌型人iga与膜人cd89的结合。基于它们的cd89/iga阻断概况,发现了以下排序(从强至较低cd89/iga阻断程度):20b4》30c7》8f3=10e7》16d6,其与在facs中发现的使用血清人iga的cd89/iga阻断程度(参见图17a和17b)一致。令人感兴趣地,在这些经检查的纯化的小鼠抗人cd89抗体在空间上阻断分泌型人iga与膜人cd89的结合的程度(此实施例)与它们对膜人cd89的相应相对结合亲和力(参见以上实施例7(a))之间似乎存在强正相关关系。为供参考,纯化的小鼠抗人cd89抗体克隆mip8a(一种熟知的cd89/iga阻断剂)(zhang等人clin exp immunol 2000;121:106-111)并行运行,并且也显示分泌型人iga与膜人cd89的结合的剂量依赖性阻断。
[0460]
总之,这些结果证明,cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7阻止了单体、二聚体、三聚体、四聚体或更高级多聚体血清人iga(即,非聚集和热聚iga)和二聚体分泌型人iga与膜人cd89的结合。
[0461]
实施例8.使用离体表达人cd89的原代人嗜中性粒细胞进行cd89/iga阻断嵌合小鼠/人抗人cd89单克隆抗体的生物学表征
[0462]
(a).cd89/iga阻断嵌合小鼠/人抗人cd89抗体在表达人cd89的原代人嗜中性粒细胞上的结合
[0463]
为了确定纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体在表达人cd89的原代人嗜中性粒细胞上的结合,使用了facs分析。
[0464]
使用lymphoprep
tm
(axis-shield)梯度离心、然后在nh4cl裂解缓冲溶液中裂解红细胞,从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞在4℃下以10x106个细胞/ml置于含有0.1% bsa的冰冷冻了的pbs(sigma-aldrich;pbs/bsa)中10分钟,所述pbs补充有50μg/ml人igg(阻断可能的fcγ受体;sigma-aldrich)。
然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与100μl的10μl/ml嵌合小鼠/人抗人cd89特异性抗体(即,8f3、10e7、16d6、20b4和30c7)(在pbs/bsa中)在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的人igg4同种型对照抗体(bristol-myers squibb)(在pbs/bsa中)作为阴性对照运行。在pbs/bsa中充分洗涤后,随后将细胞与1:100稀释的异硫氰酸荧光素缀合的小鼠抗人igg4特异性抗体(sigma-aldrich)在4℃下孵育30分钟。在pbs/bsa中充分洗涤后,将细胞在4℃下在pbs/bsa中的2%甲醛中固定30分钟。使用流式细胞仪(cyan;beckmancoulter)测量抗体在来自离体人嗜中性粒细胞的膜人cd89上的结合(几何平均荧光强度)。
[0465]
如图18所示,所有我们的纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体(在10μg/ml下)结合至来自从多个供体(n=3)分离的离体人嗜中性粒细胞的膜人cd89。基于它们在嗜中性粒细胞上的结合概况,发现了以下排序(从高至较低结合强度):8f3=10e7=30c7=20b4》16d6。
[0466]
这些结果还证明,我们的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7识别来自离体人嗜中性粒细胞的膜人cd89上的非多态表位,因为这些抗体明确地与从所有3个经检查的供体中分离的人嗜中性粒细胞结合。
[0467]
(b).使用cd89/iga阻断嵌合小鼠/人抗人cd89单克隆抗体阻断血清人iga介导的表达人cd89的原代人嗜中性粒细胞的吞噬作用
[0468]
为了分析纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断嵌合小鼠/人抗人cd89抗体抑制表达人cd89的原代人嗜中性粒细胞对血清人iga包被的乳胶珠的吞噬作用的能力。
[0469]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。然后,将100μl/孔(即,在96孔平底板中0.2x106个细胞;greiner)的这些细胞与滴定的(在rpmi/10% fcs中)纯化的嵌合小鼠/人抗人cd89抗体(即,8f3、10e7、16d6、20b4和30c7)在4℃下孵育20分钟。在此之后(即,不洗涤),将1.2μl细胞与珠比例为1:60的纯化的人(血清来源的)iga(mp biomedicals)包被的荧光乳胶珠(1μm大小且羧酸酯改性的聚苯乙烯;sigma-aldrich)添加至这些细胞中(制剂iga珠,参见aleyd等人j immunol 2014;192:2374-2383),并在37℃下再孵育30分钟。并行地,细胞与珠粒比例为1:60的bsa (sigma-aldrich)包被的荧光乳胶珠作为阴性对照运行(制剂bsa-珠,参见aleyd等人j immunol 2014;192:2374-2383)。在rpmi/10% fcs中洗涤并重悬于pbs/0.1% bsa(sigma-aldrich)中后,使用流式细胞仪(cyan;beckman coulter)测量血清人iga介导的离体人嗜中性粒细胞上的膜人cd89对荧光乳胶珠的吞噬作用(根据aleyd等人j immunol 2014;192:2374-2383的研究,使用几何平均荧光强度来计算吞噬指数)。为了比较,商业小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3并行运行,如实施例3(b)中所述。
[0470]
如图19所示,所有纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体剂量依赖性地抑制了分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞中血清人iga介导的吞噬作用,尽管观察到一些供体间变化。基于其血清人iga介导的吞噬作用抑制的程度,发现以下排序(从强至较低的iga介导的吞噬作用抑制程度):8f3=10e7=30c7》
20b4》16d16。
[0471]
这些结果证明,cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体8f3、10e7、16d6、20b4和30c7抑制了血清人iga介导的表达膜人cd89的离体原代人嗜中性粒细胞的吞噬活性。
[0472]
(c).使用cd89/iga阻断嵌合小鼠/人抗人cd89单克隆抗体阻断血清人iga介导的表达人cd89的原代人嗜中性粒细胞的迁移、血清人iga介导的表达人cd89的原代人嗜中性粒细胞的趋化性以及血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的白三烯b4产生
[0473]
为了分析纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断嵌合小鼠/人抗人cd89抗体抑制(1)血清人iga介导的表达人cd89的原代人嗜中性粒细胞的迁移、(2)血清人iga介导的表达人cd89的原代人嗜中性粒细胞的趋化性和(3)血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的嗜中性粒细胞化学引诱物白三烯b4(ltb4)产生的能力。
[0474]
二维(2-d)迁移测定:使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以2.5x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中,且随后将100μl钙黄绿素-am标记的粒细胞(即,在96孔平底板中0.25x106个细胞/孔;greiner)与20μg/ml(在rpmi/10% fcs中)纯化的嵌合小鼠/人抗人cd89抗体(即,8f3、10e7、16d6、20b4和30c7)在4℃下孵育20分钟。并行地,20μg/ml(在rpmi/10% fcs中)纯化的人igg4同种型对照(sigma-aldrich)作为阴性对照运行。在此之后,每孔添加150μl rpmi/10% fcs,并且将细胞静置10分钟以再次变成单层。在此之后(即,不洗涤),将10μl纯化的人(血清来源的)iga(mp biomedicals)包被的琼脂糖4b珠(90μm大小且溴化氰激活的;ge healthcare)轻轻地添加至这些单层细胞(制剂3μg/ml iga珠,参见van der steen等人gastroentorol 2009;137:2018-2029),并在37℃下再孵育40分钟。并行地,bsa(sigma-aldrich)包被的琼脂糖4b珠作为阴性对照运行(制剂3μg/ml bsa珠,参见van der steen等人gastroentorol 2009;137:2018-2029)。然后,收集上清液并用于趋化性测定和lbt4 elisa(参见下文),并且洗涤琼脂糖珠以除去非结合/非迁移的钙黄绿素-am标记的粒细胞。随后,将粒细胞在0.2%(w/v)十六烷基三甲基溴化铵(sigma-aldrich)缓冲液中在室温下裂解30分钟,并且使用荧光计(fluostar/polarstar;bmg labtech)测量96孔平底板(greiner)中释放的钙黄绿素-am(反映iga结合/迁移的粒细胞的数量)。通过使用已知数量的裂解的钙黄绿素-am标记的粒细胞的标准曲线(即,0-0.3x106个细胞/孔)来定量iga结合/迁移的离体人嗜中性粒细胞的数量。为了比较,商业小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3并行运行,如实施例3(c)中所述。
[0475]
趋化性测定:使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以1.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中。为了测量趋化性,用29μl来自iga包被的
琼脂糖珠刺激的原代人嗜中性粒细胞(即,来自另一个健康供体,参见上文)的上清液填充博伊登室(neuro probe)的下部隔室的孔。并行地,仅rpmi/10% fcs培养基和纯化的1或10nm ltb4(在rpmi中;sigma-aldrich)分别作为阴性对照和阳性对照运行。随后,用3μm孔径的聚乙烯吡罗烷酮包被的聚碳酸酯过滤器(neuro probe)覆盖下部隔室,然后将上部隔室组装到博伊登室的下部隔室上。在此之后,在上部隔室的孔中添加50μl钙黄绿素-am标记的粒细胞(即,0.05x106个细胞/孔)。在37℃下孵育40分钟后,测定离体人嗜中性粒细胞从上部隔室朝向下部隔室的孔的趋化性。为此,将下部隔室中的粒细胞在0.1%(w/v)十六烷基三甲基溴化铵(sigma-aldrich)缓冲液中在室温下裂解30分钟,并且使用荧光计(fluostar/polarstar;bmg labtech)测量96孔平底板(greiner)中释放的钙黄绿素am(反映趋化粒细胞的数量)。通过使用已知数量的裂解的钙黄绿素-am标记的粒细胞的标准曲线(即,0-0.05x106个细胞/孔)来定量iga包被的琼脂糖珠诱导的趋化离体人嗜中性粒细胞的数量。
[0476]
ltb4 elisa:测量来自iga包被的琼脂糖珠刺激的离体人嗜中性粒细胞的上清液中的ltb4水平(参见上文)。为此,根据制造商的说明书使用了市售ltb4竞争性elisa试剂盒(r&d systems)。
[0477]
如图20a所示,20μg/ml的所有我们的纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体(除16d6外)都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的迁移,尽管观察到一些供体间变化。
[0478]
如图20b所示,20μg/ml的所有我们的纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体(除16d6外)都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的趋化性,尽管观察到一些供体间变化。
[0479]
如图20c所示,20μg/ml的所有我们的纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体(除16d6外)都抑制了血清人iga包被的琼脂糖珠介导的分离自三个健康个体的表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4产生,尽管观察到一些供体间变化。
[0480]
这些结果证明,cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体8f3、10e7、20b4和30c7抑制了血清人iga介导的表达膜人cd89的离体原代人嗜中性粒细胞的迁移、趋化性和来自表达膜人cd89的离体原代人嗜中性粒细胞的化学吸引ltb4释放。
[0481]
(d).使用cd89/iga阻断嵌合小鼠/人抗人cd89单克隆抗体阻断血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的乳铁蛋白产生
[0482]
为了分析纯化的cd89/iga阻断嵌合小鼠/人抗人cd89抗体的生物活性,测定了所产生的cd89/iga阻断嵌合小鼠/人抗人cd89抗体抑制血清人iga介导的来自表达人cd89的原代人嗜中性粒细胞的乳铁蛋白产生的能力。
[0483]
使用lymphoprep
tm
(axis-shield)梯度离心、随后在nh4cl裂解缓冲溶液中裂解红细胞来从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。然后,将这些原代人嗜中性粒细胞用1μm荧光钙黄绿素-am(molecular probes)在37℃下标记30分钟。洗涤后,将这些钙黄绿素-am标记的粒细胞以2.0x106个细胞/ml重悬于补充有10%热灭活fcs(sigma-aldrich)的rpmi 1640(gibco)中,且随后将100μl钙黄绿素-am标记的粒细胞(即,0.2x106个细胞/孔)与滴定的(在rpmi/10% fcs中)纯化的嵌合小鼠/人抗人cd89抗体(即,
8f3、10e7、16d6、20b4和30c7)在4℃下孵育20分钟。在此之后(即,不洗涤),将100μl的这些细胞(即,0.2x106个细胞/孔)添加至96孔平底elisa板(nunc-immuno maxisorp),所述板先前用100μl/孔的10μg/ml的纯化的人(血清来源的)iga (mp biomedicals)或100μl/孔的10μg/ml的bsa(用作阴性对照;sigma-aldrich)包被。在37℃下孵育30分钟后,收获上清液(180μl/孔)以除去非结合的粒细胞,并使用这些上清液(在若干离心清除步骤后)来测量乳铁蛋白生产水平(用作脱粒标志物;参见下文)。为了比较,商业小鼠抗人cd89抗体克隆mip8a、克隆a59和克隆a3并行运行,如实施例3(d)中所述。
[0484]
测量用板结合的血清人iga刺激的原代人嗜中性粒细胞的上清液中的乳铁蛋白产生(代表脱粒程度)(参见上文)。为此,在4℃-8℃下将96孔平底elisa板(nunc-immuno maxisorp)用100μl/孔兔抗人乳铁蛋白抗体(1:5000;sigma-aldrich)包被16-24小时。在pbs/0.05% tween 20中充分洗涤后,在室温下将板用200μl/孔pbs/0.05% tween 20/0.5% bsa (sigma-aldrich)封闭1小时。然后将板与100μl/孔的1:2稀释度的上清液(在封闭缓冲液中)在37℃孵育1小时。在pbs/0.05% tween 20中充分洗涤后,将板与碱性磷酸酶标记的兔抗人乳铁蛋白检测抗体(1:2500;mp biomedicals)在37℃下孵育1小时。在添加磷酸对硝基苯酯(sigma-aldrich)后,使用微板读数器(imark;biorad)在450nm波长下测量光密度。纯化的人乳铁蛋白(sigma-aldrich)用作标准物,以计算由血清人iga刺激的离体人嗜中性粒细胞释放的乳铁蛋白的量。
[0485]
如图21所示,所有我们的纯化的cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体(除16d6外)都剂量依赖性地抑制了血清人iga介导的分离自两个健康个体的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生,尽管观察到一些供体间变化。
[0486]
这些结果证明,cd89/iga阻断嵌合小鼠/人抗人cd89特异性抗体8f3、10e7、20b4和30c7抑制了iga介导的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生(脱粒标志物)。
[0487]
实施例9.cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7的产生
[0488]
基于cd89/iga阻断小鼠抗人cd89抗体10e7的确定的小鼠v区(参见以上实施例5),产生了cd89/iga阻断人源化抗人cd89抗体10e7型式。
[0489]
使用pdl技术(由panorama research institute,sunnyvale,ca,usa进行)获得了cd89/iga阻断小鼠抗人cd89抗体10e7的计算机人源化可变重链序列(3)和人源化可变轻链序列(4)。对于cd89/iga阻断小鼠抗人cd89抗体10e7的人源化可变重链和可变轻链氨基酸序列,参见seq id no:117(vh1)、118(vh2)、119(vh3)、120(vl1)、121(vl2)、122(vl3)和123(vl4)。
[0490]
在这种设计后,在geneart(regensburg,germany)订购了灰仓鼠优化的cdna序列seq id no:124、125、126(编码全长人源化重igg4链10e7型式,即分别vh1、vh2、vh3)和seq id no:127、128、129、130(编码全长人源化轻κ链10e7型式,即分别vl1、vl2、vl3、vl4),所述cdna序列编码人信号肽,随后是与人稳定的(即,s239p;根据angal等人在mol.immunol.,第30卷,第1期,第105-108页中,1993)igg4恒定区连接的人源化可变重链,或随后是与人κ恒定区连接的人源化可变轻链。使用合适的限制性酶,将产生的cdna亚克隆到pcdna3.1来源的表达质粒中。随后使用freestyle
tm 293表达系统(invitrogen)在293-f细胞中瞬时表达cd89/iga阻断人源化抗人cd89抗体10e7型式。分别使用常规elisa和流式细胞术(实施例
10,参见下文)检查含有这些表达的cd89/iga阻断人源化抗人cd89抗体10e7型式的上清液的针对rhucd89和膜结合的人cd89的结合。
[0491]
以这种方式,产生了cd89/iga阻断人源化抗人cd89抗体10e7的十二种人源化型式,即,vh1vl1、vh1vl2、vh1vl3、vh1vl4、vh2vl1、vh2vl2、vh2vl3、vh2vl4、vh3vl1、vh3vl2、vh3vl3和vh3vl4。
[0492]
对于cd89/iga阻断人源化抗人cd89抗体10e7的人源化氨基酸序列,参见seq id no:131、132、133(编码全长人源化重igg4链10e7型式,即,分别vh1、vh2、vh3)和seq id no:134、135、136、137(编码全长人源化轻κ链10e7型式,即,分别vl1、vl2、vl3、vl4)。
[0493]
实施例10.cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7的结合表征
[0494]
(a).cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式对人cd89的相对结合亲和力
[0495]
为了确定cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式对人cd89的相对结合亲和力,使用了elisa和facs分析。
[0496]
elisa:在4℃-8℃下在16-24小时期间,使用96孔maxisorp板(nunc),将rhucd89(sino biological)以0.5μg/ml包被在pbs中(50ng/100μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1%bsa(roche)封闭1小时。随后,将板与和不与100μl滴定的(在freestyle
tm 293表达培养基(invitrogen)中;通过使用生物层干涉法设备与蛋白a生物传感器尖端的组合预先测定上清液(参见实施例9)中针对内部人igg4κ标准物的抗体浓度)人源化抗人cd89抗体10e7/孔在室温下孵育1小时。嵌合抗人cd89抗体10e7(参见实施例6)并行运行,以供参考。在pbs/0.05% tween 20中充分洗涤后,在室温下用1:5000稀释的辣根过氧化物酶(hrp)缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)测定抗体在rhucd89上的结合,持续1小时,随后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m hcl后,使用微板读数器(synergy htx;biotek)在450nm波长(参考波长为655nm)下测量抗体在rhucd89上的结合(光密度)。
[0497]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与和不与100μl滴定的(在freestyle
tm 293表达培养基(invitrogen)中;通过使用生物层干涉法设备与蛋白a生物传感器尖端的组合预先测定上清液(参见实施例9)中针对内部人igg4κ标准物的抗体浓度)人源化抗人cd89抗体10e7/管在4℃下孵育30分钟。嵌合抗人cd89抗体10e7(参见实施例6)并行运行,以供参考。在pbs/bsa/nan3中充分洗涤后,随后将细胞在4℃下与1:200稀释的pe缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的4%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量抗体在膜人cd89上的结合(几何平均荧光强度)。
[0498]
如图25a所示,所有cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式(即,vh1vl1、vh1vl2、vh1vl3、vh1vl4、vh2vl1、vh2vl2、vh2vl3、vh2vl4、vh3vl1、vh3vl2、vh3vl3和vh3vl4)和cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7都剂量依赖性地与
rhucd89结合。cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,参考)和cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3vl3和vh3vl4显示相似的s形滴定曲线,这表明它们针对rhucd89的结合亲和力相似(半最大结合ec
50
≈20ng/ml),而cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式vh1vl1、vh1vl2、vh1vl3、vh1vl4、vh2vl1、vh2vl2、vh2vl3、vh2vl4、vh3vl1和vh3vl2显示显著更低的结合亲和力,这通过较低的最大光密度信号和不相似的滴定曲线来说明。
[0499]
如图25b所示,所有cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式(即,vh1vl1、vh1vl2、vh1vl3、vh1vl4、vh2vl1、vh2vl2、vh2vl3、vh2vl4、vh3vl1、vh3vl2、vh3vl3和vh3vl4)和cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7都剂量依赖性地与膜人cd89结合。cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,参考)和cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3vl3和vh3vl4显示相似的s形滴定曲线,这表明它们针对膜人cd89的结合亲和力相似(半最大结合ec50≈1000ng/ml),而cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7型式vh1vl1、vh1vl2、vh1vl3、vh1vl4、vh2vl1、vh2vl2、vh2vl3、vh2vl4、vh3vl1和vh3vl2显示显著更低的结合亲和力,这通过较低的最大荧光信号和不相似的滴定曲线来说明。
[0500]
在这种上清液筛选后,选择了cd89/iga阻断人源化igg4/κ抗人cd89单克隆抗体10e7 vh3vl3。然而,cd89/iga阻断抗人cd89单克隆抗体10e7的可变重链中的cdr2(参见seq id no:46)含有两个脱酰胺“天冬酰胺-甘氨酸”(“asn-gly”)基序,所述基序易于脱酰胺(sydow等人plos one 2014;9(6):e100736)。脱酰胺是化学反应,其中氨基酸天冬酰胺的侧链中的酰胺官能团被转化为另一个官能团。通常,天冬酰胺被转化为天冬氨酸或异天冬氨酸。在抗体中,这种反应可形成严重风险因子,因为这种反应可改变抗体结构、稳定性或功能(即,抗体-抗原结合),并且可导致抗体降解。因此,使用fv结构模型(由applied protein services,lonza biologics,cambridge,uk进行)获得了cd89/iga阻断小鼠抗人cd89抗体10e7的计算机“cdr2脱酰胺修复的”人源化可变重链序列。基于这一详细评估,vh:asn52和vh:asn54分别被(1)vh:ser52和vh:gln54的(vh3sq)组合以及(2)vh:ser52和vh:thr54的(vh3st)组合取代。对于cd89/iga阻断小鼠抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化可变重链氨基酸序列,参见seq id no:138(vh3sq)和139(vh3st)。
[0501]
在这种设计后,在geneart(regensburg,germany)订购了灰仓鼠优化的cdna序列seq id no:140、141(编码全长“cdr2脱酰胺修复的”人源化重igg4链10e7型式,即分别vh3sq、vh3st)和seq id no:129(编码全长人源化轻κ链10e7型式,即vl3),所述cdna序列编码人信号肽,随后是与人稳定的(即,s239p;根据angal等人在mol.immunol.,第30卷,第1期,第105-108页,1993)igg4恒定区连接的“cdr2脱酰胺修复的”人源化可变重链,或随后是与人κ恒定区连接的人源化可变轻链。使用合适的限制性酶,将产生的cdna亚克隆到pcdna3.1来源的表达质粒中。随后使用freestyle
tm 293表达系统(invitrogen)在293-f细胞中瞬时表达cd89/iga阻断人源化抗人cd89抗体10e7型式。收获上清液后,使用常规亲和色谱法蛋白a柱纯化cd89/iga阻断“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7型式vh3sqvl3和vh3stvl3,并且随后分别使用常规elisa和流式细胞术(参见下文的实施例10(b))检查其针对rhucd89和膜结合的人cd89的结合。此外,使用lal显色终点测定(hycult biotech)测定lps水平,并且cd89/iga阻断“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7
型式vh3sqvl3和vh3stvl3含有《0.002eu lps/μg人源化igg。
[0502]
通过这种方式,产生了cd89/iga阻断人源化抗人cd89抗体10e7的两种“cdr2脱酰胺修复的”人源化型式,即vh3sqvl3和vh3stvl3。
[0503]
对于cd89/iga阻断人源化抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化氨基酸序列,参见seq id no:142、143(编码全长“cdr2脱酰胺修复的”人源化重igg4链10e7型式,即,分别vh3sq、vh3st)和seq id no:136(编码全长人源化轻κ链10e7型式,即,vl3)。
[0504]
(b).cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对人cd89的相对结合亲和力
[0505]
为了确定cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对人cd89的相对结合亲和力,使用了elisa和facs分析。
[0506]
elisa:在4℃-8℃下在16-24小时期间,使用96孔maxisorp板(nunc),将rhucd89(10414-h08h,sino biological,inc)以0.5μg/ml包被在pbs中(50ng/100μl/孔)。在用pbs/0.05% tween 20充分洗涤后,在室温下将板用pbs/0.05% tween 20/1% bsa (roche)封闭1小时。随后,将板与或不与100μl滴定的(在pbs/bsa/nan3中)纯化的“cdr2脱酰胺修复的”人源抗人cd89抗体10e7/孔在室温下孵育1小时。嵌合抗人cd89抗体10e7(参见实施例6)并行运行,以供参考。在pbs/0.05% tween 20中充分洗涤后,在室温下用1:5000稀释的辣根过氧化物酶(hrp)缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)测定抗体在rhucd89上的结合1小时,随后用tmb底物的即用型溶液(invitrogen)进行比色检测。在添加1m hcl后,使用微板读数器(synergy htx;biotek)在450nm波长(参考波长为655nm)下测量抗体在rhucd89上的结合(光密度)。
[0507]
facs:将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))在4℃下以10x106个细胞/ml置于冰冷冻了的pbs中10分钟,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3),补充有50μg/ml人igg(阻断可能的fcγ受体;sigma)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与和不与100μl滴定的(在pbs/bsa/nan3中)纯化的“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7/管在4℃下孵育30分钟。嵌合抗人cd89抗体10e7(参见实施例6)并行运行,以供参考。在pbs/bsa/nan3中充分洗涤后,随后在4℃下将细胞与1:200稀释的pe缀合的山羊抗人igg fcγ特异性抗体(jackson immunoresearch)孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的4%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量抗体在膜人cd89上的结合(几何平均荧光强度)。
[0508]
如图26a所示,“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者以及嵌合igg4/κ抗人cd89单克隆抗体10e7均剂量依赖性地与rhucd89结合。cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,参考)和cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3显示相似的s形滴定曲线,这表明它们针对板结合的rhucd89的结合亲和力相似(半最大结合ec
50
≈10ng/ml)。
[0509]
如图26b所示,“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者以及嵌合igg4/κ抗人cd89单克隆抗体10e7均剂量依赖性地与膜人cd89结合。出人意料地,cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,参考)以
及cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者显示不相似的s形滴定曲线,这表明cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3针对膜人cd89的结合亲和力(半最大结合ec50≈600ng/ml)高于cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7的结合亲和力(半最大结合ec50≈1300ng/ml)。综上所述,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者似乎在“cdr2脱酰胺修复”过程中增加它们对膜结合的人cd89的结合亲和力。
[0510]
(c).cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7形式vh3sqvl3和vh3stvl3的cd89/iga阻断能力的程度
[0511]
为了分析纯化的“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3的cd89/iga阻断的程度,通过使用facs分析来测定纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3在空间上阻碍血清或分泌型人iga与人cd89的相互作用的能力。
[0512]
将稳定的人全长cd89转染的hek293f细胞(克隆2号;参见以上实施例1(b))以10x106个细胞/ml置于冰冷冻了的pbs中,所述pbs含有0.1% bsa(sigma)/0.05% nan3(pbs/bsa/nan3)。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与50μl滴定的(在pbs/bsa/nan3中)纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3/管在4℃下孵育30分钟。并行地,运行50μl滴定的(在pbs/bsa/nan3中)纯化的嵌合抗人cd89抗体10e7(参见实施例6)以供参考。在此之后(即,不洗涤),将50μl的0.20μm纯化的人血清来源的iga(bethyl laboratories)(在pbs/bsa/nan3中稀释)或0.16μm的初乳来源的iga(biorad)(在pbs/bsa/nan3中稀释)添加至这些细胞,并在4℃下再孵育30分钟。在pbs/bsa/nan3中充分洗涤后,在4℃下用5μg/ml的生物素缀合的f(ab')2片段山羊抗人血清igaα链特异性抗体(jackson immunoresearch)测定血清人iga或分泌型人iga在膜人cd89上的结合,持续30分钟。在pbs/bsa/nan3中充分洗涤后,添加1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch),并在4℃下孵育30分钟。在pbs/bsa/nan3中充分洗涤后,将细胞在4℃下在pbs/bsa/nan3中的4%甲醛中固定30分钟。使用流式细胞仪(facscalibur;bd biosciences)测量血清人iga或分泌型人iga在膜人cd89上的结合(几何平均荧光强度)。
[0513]
如图27a和27b所示,“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者以及cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7均剂量依赖性地阻止了血清和分泌型人iga与膜人cd89的结合。出人意料地,cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,参考)以及cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者显示不相似的抑制滴定曲线,这表明“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者的cd89/iga抑制强于(分别地,对血清人iga和分泌型人iga的半最大抑制ic50≈900ng/ml和ic50≈600ng/ml)嵌合igg4/κ抗人cd89单克隆抗体10e7的cd89/iga抑制(分别地,对血清人iga和分泌型人iga的半最大抑制ic50≈1600ng/ml和ic50≈1000ng/ml)。综上所述,“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3似乎在“cdr2脱酰胺修复”过程中增加其cd89/iga阻断能力程度,这与它们对膜
结合的人cd89的结合亲和力增加一致(参见以上实施例10(b))。
[0514]
总之,这些结果证明,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3阻止了单体血清人iga和二聚体分泌型人iga与膜人cd89的结合。
[0515]
(d).cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对人cd89的平衡解离常数(kd)
[0516]
为了测定cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3对人cd89的平衡解离常数(kd),使用了基于bli/的分析。
[0517]
在30℃下在5分钟过程中将生物素化的(使用来自pierce的n-羟基琥珀酰亚胺基-生物素)纯化的“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7型式vh3sqvl3和vh3stvl3以在pbs/0.1% bsa/0.02% tween 20(ph 7.4)中2.5μg/ml负载到链霉亲和素生物传感器尖端(fortebio)上。随后,在30℃下测量0nm(用于监测基线漂移的参考样品)和0.31-20nm(在pbs/0.1% bsa/0.02%tween 20中的2倍稀释步骤)下的单体重组c末端聚组氨酸标记的人(细胞外)cd89分析物(rhucd89(10414-h08h,sino biological,inc))的缔合,持续5分钟,然后是使用red96设备(fortebio)在30℃下20分钟的解离期。出于比较目的对生物素化的(使用来自pierce的n-羟基琥珀酰亚胺基-生物素)纯化的小鼠和嵌合抗人cd89抗体10e7对应物(分别参见实施例2和实施例6)进行了测试。在red96软件(fortebio)中使用1:1langmuir结合模型测定平衡解离常数(kd)。
[0518]“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者、嵌合igg4/κ抗人cd89单克隆抗体10e7和小鼠抗人cd89单克隆抗体10e7显示与可溶性单体rhucd89的高亲和力结合。出人意料地,与cd89/iga阻断嵌合igg4/κ抗人cd89单克隆抗体10e7(即,kd值为1.28nm)和cd89/iga阻断小鼠抗人cd89单克隆抗体10e7(即,kd值1.67nm)的结合亲和力相比,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3显示与可溶性单体rhucd89的更高亲和力结合(即,kd值分别为0.82nm和0.92nm)。
[0519]
总之,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者似乎在“cdr2脱酰胺修复”过程中增加它们对可溶性单体rhucd89的结合亲和力。关于总结,参见表9。
[0520]
表9.来自用纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3、嵌合igg4/κ抗人cd89单克隆抗体10e7以及小鼠抗人cd89单克隆抗体10e7对可溶性单体rhucd89进行的动力学测定的检查的结合动力学参数的结果。
[0521]
结合动力学参数
[0522]
[0523][0524]
*来自2个独立实验的结果(平均值
±
sd),**来自1个独立实验的结果。
[0525]
实施例11.使用离体表达人cd89的原代人嗜中性粒细胞进行的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3的生物学表征
[0526]
(a).cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3在表达人cd89的原代人嗜中性粒细胞上的结合
[0527]
为了确定纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3在表达人cd89的原代人嗜中性粒细胞上的结合,使用了facs分析。
[0528]
使用lymphoprep
tm
(axis-shield)梯度离心、然后在nh4cl裂解缓冲溶液中裂解红细胞,从健康供体(在知情同意后)外周血中分离原代人嗜中性粒细胞。在pbs中洗涤后,将粒细胞以10x106个细胞/ml置于含有0.1% bsa的冰冷冻了的pbs(sigma-aldrich;pbs/bsa)中。然后,将10μl/管(即,0.1x106个细胞)的这些细胞与或不与100μl的10μl/ml纯化的“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7(在pbs/bsa中)在4℃下孵育30分钟。并行地,100μl的10μg/ml纯化的人igg4同种型对照抗体(sigma-aldrich)(在pbs/bsa中)作为阴性对照运行。此外,运行100μl的10μl/ml纯化的嵌合抗人cd89抗体10e7(参见实施例6)(在pbs/bsa中)以供参考。在pbs/bsa中充分洗涤后,随后将细胞与1:200稀释的生物素缀合的小鼠抗人igg4特异性抗体(invitrogen)在4℃下孵育30分钟。在pbs/bsa中充分洗涤后,将细胞与1:200稀释的pe缀合的链霉亲和素(jackson immunoresearch)在4℃下孵育30分钟。在pbs/bsa中充分洗涤后,将细胞在4℃下在pbs/bsa中的2%甲醛中固定30分钟。使用流式细胞仪(attune;thermo fisher)测量抗体在来自离体人嗜中性粒细胞的膜人cd89上的结合(几何平均荧光强度)。
[0529]
如图28所示,纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者(在10μg/ml下)结合至来自分离自多个供体(n=3)的人嗜中性粒细胞的膜人cd89。
[0530]
这些结果还证明,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3识别来自离体人嗜中性粒细胞的膜人cd89上的非多态表位,因为这些抗体明确地与从所有3个经检查的供体中分离的人嗜中性粒细胞结合。
[0531]
(b).使用cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3阻断血清人iga介导的表达人cd89的原代人嗜中性粒细胞的吞噬作用
[0532]
为了分析纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3的生物活性,测定了所产生的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3抑制表达人
maxisorp),所述板先前用100μl/孔10μg/ml的纯化的人(血清来源的)iga(mp biomedicals)或100μl/孔10μg/ml的bsa(用作阴性对照;sigma-aldrich)包被。在37℃下孵育30分钟后,收获上清液(180μl/孔)以除去非结合的粒细胞,并使用这些上清液(在若干离心清除步骤后)来测量乳铁蛋白生产水平(用作脱粒标志物;参见下文)。为了比较,商业小鼠抗人cd89抗体克隆mip8a并行运行,如实施例3(d)中所述。
[0539]
测量用板结合的血清人iga刺激的原代人嗜中性粒细胞的上清液中的乳铁蛋白产生(代表脱粒程度)(参见上文)。为此,在4℃-8℃下将96孔平底elisa板(nunc-immuno maxisorp)用100μl/孔兔抗人乳铁蛋白抗体(1:5000;sigma-aldrich)包被16-24小时。在pbs/0.05% tween 20中充分洗涤后,在室温下将板用200μl/孔pbs/0.05% tween 20/0.5% bsa(sigma-aldrich)封闭1小时。然后将板与100μl/孔1:2稀释度的上清液(在封闭缓冲液中)在37℃孵育1小时。在pbs/0.05% tween 20中充分洗涤后,将板与碱性磷酸酶标记的兔抗人乳铁蛋白检测抗体(1:2500;mp biomedicals)在37℃下孵育1小时。在添加磷酸对硝基苯酯(sigma-aldrich)后,使用微板读数器(imark;biorad)在450nm波长下测量光密度。纯化的人乳铁蛋白(sigma-aldrich)用作标准物,以计算由血清人iga刺激的离体人嗜中性粒细胞释放的乳铁蛋白的量。
[0540]
如图30所示,纯化的cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3两者剂量依赖性地抑制了血清人iga介导的分离自五个健康个体的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生,尽管观察到一些供体间变化。
[0541]
这些结果证明,cd89/iga阻断“cdr2脱酰胺修复的”人源化igg4/κ抗人cd89单克隆抗体10e7型式vh3sqvl3和vh3stvl3抑制了iga介导的表达人cd89的离体原代人嗜中性粒细胞的乳铁蛋白产生(脱粒标志物)。
[0542]
本发明的某些方面
[0543]
方面1一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89(人fcαri)的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且与抗体mip8a相比时诱导所述表达人cd89的细胞的更少细胞死亡。
[0544]
方面2一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且不会使所述细胞在37℃下过夜孵育后的细胞活力降低超过60%。
[0545]
方面3一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且不会使所述细胞在37℃下过夜孵育后的磷脂酰丝氨酸表达增加超过20%。
[0546]
方面4一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与重组人cd89分子结合,其中人cd89的氨基酸22-46被替换为食蟹猴cd89的氨基酸22-46。
[0547]
方面5一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸47-71被替换为食蟹猴cd89的氨基酸47-71。
[0548]
方面6一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,
并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸72-96被替换为食蟹猴cd89的氨基酸72-96。
[0549]
方面7一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且其与嵌合cd89分子的结合未降低20%或更少,其中人cd89的氨基酸97-121被替换为食蟹猴cd89的氨基酸97-121。
[0550]
方面8一种抗体,所述抗体能够结合表达人cd89的细胞上的人cd89的细胞外部分,并且在所述抗体与所述细胞结合时能够阻止人iga与人cd89的结合,并且20%或更少地与嵌合cd89分子结合,其中人cd89的氨基酸58、59、73、74、76、106和107被替换为食蟹猴cd89的相应氨基酸58、59、73、74、76、106和107。
[0551]
方面9根据方面1至8中任一项所述的抗体,其中所述细胞是表达人cd89的hek293f细胞(以编号dsm acc3341保藏)。
[0552]
方面10一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:29-31的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:32-34的cdr1、cdr2和cdr3序列的轻链可变区。
[0553]
方面11一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:27的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:28的氨基酸序列的轻链可变区。
[0554]
方面12一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:45-47的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:48-50的cdr1、cdr2和cdr3序列的轻链可变区。
[0555]
方面13一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:43的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:44的氨基酸序列的轻链可变区。
[0556]
方面14一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:69-71的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:72-74的cdr1、cdr2和cdr3序列的轻链可变区。
[0557]
方面15一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:67的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:68的氨基酸序列的轻链可变区。
[0558]
方面16一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:77-79的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:80-82的cdr1、cdr2和cdr3
序列的轻链可变区。
[0559]
方面17一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:75的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:76的氨基酸序列的轻链可变区。
[0560]
方面18一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:53-55的cdr1、cdr2和cdr3序列的重链可变区,和具有含0、1或2个氨基酸插入、缺失、取代或添加的seq id no:56-58的cdr1、cdr2和cdr3序列的轻链可变区。
[0561]
方面19一种能够结合人cd89的细胞外部分的抗体,所述抗体包含具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:51的氨基酸序列的重链可变区,和具有含0、1、2、3、4、5、6、7、8、9或10个氨基酸插入、缺失、取代或添加的seq id no:52的氨基酸序列的轻链可变区。
[0562]
序列说明
[0563]
seq id no:1:
[0564]
全长人cd89的氨基酸序列(swiss-prot号p24071.1;aa 287)
[0565]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayltqlmiiknstyreigrrlkfwnetdpefvidhmdankagryqcqyrighyrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0566]
信号肽(aa序列1-21),包含ig样ec1结构域(aa序列22-121)、短铰链区(aa序列122-125)、ig样ec2结构域(aa序列126-220)和膜近端“接头”区域(aa序列221-227)的细胞外结构域(aa序列22-227),然后是跨膜结构域(aa序列228-246)和短细胞质尾(aa序列247-287),根据ding等人j biol chem2003;278:27966-27970。
[0567]
seq id no:2:
[0568]
编码全长人cd89蛋白的cdna序列(优化用于哺乳动物表达)
[0569]
atggaccccaagcagaccaccctgctgtgcctggtgctgtgtctgggccagagaatccaggcccaggaaggcgacttccccatgcccttcatcagcgccaagagcagccccgtgatccccctggatggcagcgtgaagatccagtgccaggccatcagagaggcctacctgacccagctgatgatcattaagaacagcacctaccgcgagatcggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttccggtacagcgacaccctggaactggtcgtgaccggcctgtacggcaagcctttcctgagcgccgatcggggactggtgctgatgcccggcgagaacatcagcctgacctgtagcagcgcccacatccccttcgacagattcagcctggccaaagagggcgagctgagcctgcctcagcatcagtctggcgagcaccccgccaactttagcctgggccctgtggacctgaacgtgtccggcatctaccggtgctacggctggtacaaccggtccccctacctgtggtccttccccagcaacgctctggaactggtcgtgacagacagcatccaccaggactacaccacccagaacctgatccggatggccgtggctgggctggtgctggtggctctgctggccattctggtggaaaactggcacagccacaccgccctgaacaaagaggccagcgccgatgtggccgagccttcttggagccagcagatgtgtcagcccggcctgaccttcgccagaaccccttctgtgtgcaag
[0570]
seq id no:3:
[0571]
包含来自人cd89的ec1和来自牛fcγ2r的ec2的嵌合fcr的氨基酸序列(牛tm和牛ic)
[0572]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayltqlmiiknstyreigrrlkfwnetdpefvidhmdankagryqcqyrighyrfrysdtlelvvtgeepagrlrdrpslsvrpspsvapgenvtllcqsgnrtdtfllskegaahrplrlrsqdqdgwyqaefslspvtsahggtyrcyrslstnpyllsqpseplallvadytmqnlirmglaasvllllgillcqarhdhggareaars
[0573]
来自人cd89的信号肽(aa序列1-21),由来自人cd89的ec1结构域(aa序列22-121)和来自牛fcγ2r的ec2结构域(aa序列122-232)组成的嵌合人/牛fcr细胞外结构域(aa序列22-232),跨膜结构域和来自牛fcγ2r的短细胞质尾(aa序列233-266)。
[0574]
seq id no:4:
[0575]
包含来自人cd89的ec1和来自牛fcγ2r的ec2的嵌合fcr的氨基酸序列(人tm和人ic)
[0576]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayltqlmiiknstyreigrrlkfwnetdpefvidhmdankagryqcqyrighyrfrysdtlelvvtgeepagrlrdrpslsvrpspsvapgenvtllcqsgnrtdtfllskegaahrplrlrsqdqdgwyqaefslspvtsahggtyrcyrslstnpyllsqpseplallvadytmqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0577]
来自人cd89的信号肽(aa序列1-21),由来自人cd89的ec1结构域(aa序列22-121)和来自牛fcγ2r的ec2结构域(aa序列122-232)组成的嵌合人/牛fcr细胞外结构域(aa序列22-232),来自人cd89的跨膜结构域(aa序列233-251),和来自人cd89的短细胞质尾(aa序列252-292)。
[0578]
seq id no:5:
[0579]
编码包含来自人cd89的ec1和来自牛fcγ2r的ec2的嵌合fcr的cdna序列(牛tm和牛ic;优化用于哺乳动物表达)
[0580]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccagtgccaggccatcagagaggcctacctgacacagctgatgatcattaagaacagcacctaccgcgagatcggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttcagatacagcgacaccctggaactggtggtcaccggcgaagaacctgctggcagactgagagatagacccagcctgtctgtgcggccttctccttctgttgcccctggcgagaatgtgaccctgctctgtcagagcggcaaccggaccgataccttcctgctgtctaaagaaggcgccgctcacagacccctgagactgagatcacaggaccaggacggatggtatcaggccgagttctctctgagcccagtgacatctgctcacggcggcacctacagatgctacagaagcctgagcacaaacccctatctgctgagccagcctagcgagcctctggctctgctggtggccgattacaccatgcagaacctgatcagaatgggcctcgccgcctctgttctgctgctgctgggaatcctgctgtgtcaagccagacacgatcacggcggagccagagaagctgccagatct
[0581]
seq id no:6:
[0582]
编码包含来自人cd89的ec1和来自牛fcγ2r的ec2的嵌合fcr的cdna序列(人tm和人ic;优化用于哺乳动物表达)
[0583]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccag
tgccaggccatcagagaggcctacctgacacagctgatgatcattaagaacagcacctaccgcgagatcggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttcagatacagcgacaccctggaactggtggtcaccggcgaagaacctgctggcagactgagagatagacccagcctgtctgtgcggccttctccttctgttgcccctggcgagaatgtgaccctgctctgtcagagcggcaaccggaccgataccttcctgctgtctaaagaaggcgccgctcacagacccctgagactgagatcacaggaccaggacggatggtatcaggccgagttctctctgagcccagtgacatctgctcacggcggcacctacagatgctacagaagcctgagcacaaacccctatctgctgagccagcctagcgagcctctggctctgctggtggccgattacaccatgcagaacctgatcagaatggccgtggccggactggtgctggttgcactgctggctatcctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0584]
seq id no:7:
[0585]
包含来自牛fcγ2r的ec1和来自人cd89的ec2的嵌合fcr的氨基酸序列(人tm和人ic)
[0586]
maptlpallclglsvglrtqvqagtfpkpiiwaepssvvplgssvtilcqgppntksfslnkegdstpwnihpslepwdkanffisnvreqqagryhcshfigvnwsepsepldllvaglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0587]
来自牛fcγ2r的信号肽(aa序列1-23),由来自牛fcγ2r的ec1结构域(aa序列24-119)、来自人cd89的短铰链区(aa序列aa 120-123)、来自人cd89的ec2结构域(aa序列124-218)和来自人cd89的膜近端“接头”区(aa序列219-225)组成的嵌合牛/人fcr细胞外结构域(aa序列24-225),然后是来自人cd89的跨膜结构域(aa序列226-244),和来自人cd89的短细胞质尾(aa序列245-285),根据ding等人j biol chem 2003;278:27966-27970。
[0588]
seq id no:8
[0589]
编码包含牛fcγ2r的ec1和人cd89的ec2的嵌合fcr的cdna序列(优化用于哺乳动物表达)
[0590]
atggcccctacactgcctgctctgctgtgtctgggactgtctgtgggcctgagaacacaggtgcaggccggcacattccccaagcctatcatttgggccgagcctagctctgtggtgcctctgggaagcagcgtgaccatcctgtgtcagggccctccaaacaccaagagcttcagcctgaacaaagagggcgacagcaccccttggaacattcaccctagcctggaaccttgggacaaagccaacttcttcatcagcaacgtgcgcgagcagcaggccggaagataccactgctctcacttcatcggagtgaattggagcgagcccagcgagcctctggatctgcttgtggctggcctgtacggcaagccttttctgtctgccgatagaggcctggtgctgatgcccggcgagaatatcagcctgacctgtagcagcgctcacatccccttcgacagattctccctggccaaagaaggcgagctgagcctgcctcagcatcagtctggcgaacaccccgccaacttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctggaactggtggtcaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggactggtgctggttgcactgctggctattctggtggaaaactggcacagccacaccgctctcaacaaagaagcctctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0591]
seq id no:9:
[0592]
全长牛fcγ2r的氨基酸序列(swiss-prot号q28109;aa 264)
[0593]
maptlpallclglsvglrtqvqagtfpkpiiwaepssvvplgssvtilcqgppntksfslnkegdstpwnihpslepwdkanffisnvreqqagryhcshfigvnwsepsepldllvageepagrlrdrpslsvrpspsvapgenvtllcqsgnrtdtfllskegaahrplrlrsqdqdgwyqaefslspvtsahggtyrcyrslstnpyllsqpseplallvadytmqnlirmglaasvllllgillcqarhdhggareaars
[0594]
信号肽(aa序列1-23),包含ig样ec1结构域(aa序列24-119)和ig样ec2结构域(aa序列120-230)的细胞外结构域(aa序列24-230),然后是跨膜结构域和短细胞质尾(aa序列231-264)。
[0595]
seq id no:10:
[0596]
编码全长牛fcγ2r蛋白的cdna序列(优化用于哺乳动物表达)
[0597]
atggcccctacactgcctgctctgctgtgtctgggactgtctgtgggcctgagaacacaggtgcaggccggcacattccccaagcctatcatttgggccgagcctagctctgtggtgcctctgggaagcagcgtgaccatcctgtgtcagggccctccaaacaccaagagcttcagcctgaacaaagagggcgacagcaccccttggaacattcaccctagcctggaaccttgggacaaagccaacttcttcatcagcaacgtgcgcgagcagcaggccggaagataccactgctctcacttcatcggagtgaattggagcgagcccagcgagcctctggatctgcttgttgctggcgaagaaccagccggcagactgagagatagaccctctctgagtgtgcggccctctccttctgttgcccctggcgaaaatgtgaccctgctgtgccagagcggcaacaggaccgataccttcctgctgagcaaagaaggcgccgctcacagacccctgagactgagatcacaggaccaggacggatggtatcaggccgagttcagcctgtctcctgtgacatctgctcacggcggcacctacagatgctacagaagcctgagcacaaacccctacctgctgtcccagccttctgagcctttggctctgctggtggccgactacaccatgcagaacctgatcagaatgggcctcgccgcctctgttctgctgctgctgggaatcctgctctgtcaggccagacacgatcatggcggagccagagaagccgccagatct
[0598]
seq id no:11:
[0599]
全长食蟹猴cd89的氨基酸序列(ncbi参考序列:xp_005590398.1;aa 287)
[0600]
mdpkettllclvlclgqriqaqegnfstpfistrsspvvpwggsvriqcqaipdayliwlmmlknstyekrdeklgfwndttpefvidhmdankagryrcryriglsrfrysdtlelvvtglygkpslsvdrgpvlmpgenisvtcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgsyrcygwynrspylwsfpsnalelvvtdsinrdyttqnlirmamaglvlvallailvenwhshkalnkeasadvaepswshqmcqpgwtfartpsvck
[0601]
seq id no:12:
[0602]
编码全长食蟹猴cd89的cdna序列(优化用于哺乳动物表达)
[0603]
atggaccccaaagaaaccaccctgctgtgcctggtgctgtgtctgggccagagaatccaggcccaggaaggcaacttcagcacccccttcatcagcaccagatccagccccgtggtgccttggggaggctctgtgcggattcagtgccaggccatccccgacgcctacctgatctggctgatgatgctgaagaacagcacctacgagaagcgggacgagaagctgggcttctggaacgacaccacccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccggtgccggtacagaatcggcctgagccggttcagatacagcgacaccctggaactggtcgtgaccggcctgtacggcaagcctagcctgtccgtggatagaggccccgtgctgatgcccggcgagaacatcagcgtgacctgtagcagcgcccacatccccttcgacagattcagcctggccaaagagggcgagctgagcctgcctcagcatcagtctggcgagcaccccgccaactttagcctgggccctgtggacctgaacgtgtccggcagctacagatgctacggctggtacaaccggtccccctacctgtggtccttccccagcaacgctctggaactggtcgtgacagacagcatcaaccgggactacaccacccagaacctgatccggatggctatggccggactggtgctggtggccctgctggccatcctggtggaaaactggcacagccacaaggccctgaacaaagaggccagcgccgatgtggccgagccttcttggagccaccagatgtgtcagcccggc
tggaccttcgccagaaccccttctgtgtgcaag
[0604]
seq id no:13:
[0605]
通过将来自人cd89的ec1部分(gln22-lys46)与来自食蟹猴cd89的ec1部分(gln22-arg46)交换的嵌合cd89的氨基酸序列
[0606]
mdpkqttllclvlclgqriqaqegnfstpfistrsspvvpwggsvriqcqaireayltqlmiiknstyreigrrlkfwnetdpefvidhmdankagryqcqyrighyrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0607]
来自人cd89的信号肽(aa序列1-21),包含来自食蟹猴cd89的ig样ec1结构域部分(aa序列22-46;ncbi参考序列:xp_005590398.1)和来自人cd89的ig样ec1结构域部分(aa序列47-121;swiss-prot号p24071.1)、来自人cd89的短铰链区(aa序列122-125)、来自人cd89的ig样ec2结构域(aa序列126-220)和来自人cd89的膜近端“接头”区域(aa序列221-227)的嵌合人/食蟹猴cd89细胞外结构域(aa序列22-227),然后是来自人cd89的跨膜结构域(aa序列228-246),以及来自人cd89的短细胞质尾(aa序列247-287),根据ding等人j biol chem 2003;278:27966-27970。
[0608]
seq id no:14:
[0609]
编码通过将来自人cd89的ec1部分(gln22-lys46)与来自食蟹猴cd89的ec1部分(gln22-arg46)交换的嵌合cd89的cdna序列(优化用于哺乳动物表达)
[0610]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaggctcaagagggcaacttcagcacccctttcatcagcaccagatctagccccgtggtgccttggggcggctctgttagaatccagtgccaggccatcagagaggcctacctgacacagctgatgatcattaagaacagcacctaccgcgagatcggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttcagatacagcgacaccctggaactggtggtcaccggcctgtacggcaagccttttctgagcgccgatagaggcctggtcctgatgcctggcgagaacatcagcctgacctgtagcagcgctcacatccccttcgacagattcagcctggccaaagagggcgagctgtctctgcctcagcatcagtctggcgagcaccccgccaatttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctcgaactggtcgtgaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggactggtgctggttgctctgctggctattctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0611]
seq id no:15:
[0612]
通过将来自人cd89的ec1部分(ile47-ile71)与来自食蟹猴cd89的ec1部分(ile47-arg71)交换的嵌合cd89的氨基酸序列
[0613]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaipdayliwlmmlknstyekrgrrlkfwnetdpefvidhmdankagryqcqyrighyrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0614]
来自人cd89的信号肽(aa序列1-21),包含来自人cd89的ig样ec1结构域部分(aa序列22-46;swiss-prot号p24071.1)、来自食蟹猴cd89的ig样ec1结构域部分(aa序列47-71;
ncbi参考序列:xp_005590398.1)和来自人cd89的ig样ec1结构域部分(aa序列48-121;swiss-prot号p24071.1)、来自人cd89的短铰链区(aa序列122-125)、来自人cd89的ig样ec2结构域(aa序列126-220)和来自人cd89的膜近端“接头”区(aa序列221
ꢀ‑
227)的嵌合人/食蟹猴cd89细胞外结构域(aa序列22-227),然后是来自人cd89的跨膜结构域(aa序列228-246),以及来自人cd89的短细胞质尾(aa序列247-287),根据ding等人j biol chem 2003;278:27966-27970。
[0615]
seq id no:16:
[0616]
编码通过将来自人cd89的ec1部分(ile47-ile71)与来自食蟹猴cd89的ec1部分(ile47-arg71)交换的嵌合cd89的cdna序列(优化用于哺乳动物表达)
[0617]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccagtgccaggccattcctgacgcctacctgatctggctgatgatgctgaagaacagcacctacgagaagagaggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttcagatacagcgacaccctggaactggtggtcaccggcctgtacggcaagccttttctgtctgccgatagaggactggtgctgatgcccggcgagaacatcagcctgacctgtagctctgctcacatccccttcgacagattcagcctggccaaagaaggcgagctgagcctgcctcagcatcagtctggcgaacaccccgccaacttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctcgaactggtcgtgaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggcctggttctggttgctctgctggctattctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0618]
seq id no:17:
[0619]
通过将来自人cd89的ec1部分(gly72-gly96)与来自食蟹猴cd89的ec1部分(asp72-gly96)交换的嵌合cd89的氨基酸序列
[0620]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayltqlmiiknstyreideklgfwndttpefvidhmdankagryqcqyrighyrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0621]
来自人cd89的信号肽(aa序列1-21),包含来自人cd89的ig样ec1结构域部分(aa序列22-71;swiss-prot号p24071.1)、来自食蟹猴cd89的ig样ec1结构域部分(aa序列72-96;ncbi参考序列:xp_005590398.1)和来自人cd89的ig样ec1结构域部分(aa序列97-121;swiss-prot号p24071.1)、来自人cd89的短铰链区(aa序列122-125)、来自人cd89的ig样ec2结构域(aa序列126-220)和来自人cd89的膜近端“接头”区域(aa序列221-227)的嵌合人/食蟹猴cd89细胞外结构域(aa序列22-227),然后是来自人cd89的跨膜结构域(aa序列228-246),以及来自人cd89的短细胞质尾(aa序列247-287),根据ding等人j biol chem 2003;278:27966-27970。
[0622]
seq id no:18:
[0623]
编码通过将来自人cd89的ec1部分(gly72-gly96)与来自食蟹猴cd89的ec1部分(asp72-gly96)交换的嵌合cd89的cdna序列(优化用于哺乳动物表达)
[0624]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccagtgccaggccatcagagaggcctacctgacacagctgatgatcattaagaacagcacctaccgcgagatcgacgagaagctcggcttctggaacgacaccacacctgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggccactaccggttcagatacagcgacaccctggaactggtggtcaccggcctgtacggcaagccttttctgtctgccgatagaggactggtgctgatgcccggcgagaacatcagcctgacctgtagctctgctcacatccccttcgacagattcagcctggccaaagaaggcgagctgagcctgcctcagcatcagtctggcgaacaccccgccaacttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctcgaactggtcgtgaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggcctggttctggttgctctgctggctattctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0625]
seq id no:19:
[0626]
通过将来自人cd89的ec1部分(arg97-gly12)与来自食蟹猴cd89的ec1部分(arg97-gly121)交换的嵌合cd89的氨基酸序列
[0627]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayltqlmiiknstyreigrrlkfwnetdpefvidhmdankagryrcryriglsrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0628]
来自人cd89的信号肽(aa序列1-21),包含来自人cd89的ig样ec1结构域部分(aa序列22-96;swiss-prot号p24071.1)和来自食蟹猴cd89的ig样ec1结构域部分(aa序列97-121;ncbi参考序列:xp_005590398.1)、来自人cd89的短铰链区(aa序列122-125)、来自人cd89的ig样ec2结构域(aa序列126-220)和来自人cd89的膜近端“接头”区域(aa序列221-227)的嵌合人/食蟹猴cd89细胞外结构域(aa序列22-227),然后是来自人cd89的跨膜结构域(aa序列228-246),以及来自人cd89的短细胞质尾(aa序列247-287),根据ding等人j biol chem 2003;278:27966-27970。
[0629]
seq id no:20:编码通过将来自人cd89的ec1部分(arg97-gly121)与来自食蟹猴cd89的ec1部分(arg97-gly121)交换的嵌合cd89的cdna序列(优化用于哺乳动物表达)
[0630]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccagtgccaggccatcagagaggcctacctgacacagctgatgatcattaagaacagcacctaccgcgagatcggcagacggctgaagttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccggtgcagatacagaatcggcctgagccggttccggtacagcgatacactggaactggtggtcaccggcctgtacggcaagccttttctgagcgccgatagaggactggtgctgatgcccggcgagaacatcagcctgacctgtagctctgctcacatccccttcgacagattcagcctggccaaagaaggcgagctgagcctgcctcagcatcagtctggcgaacaccccgccaacttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctggaactcgtcgtgaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggcctggttctggttgctctgctggctattctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggc
ctgaccttcgccagaacacctagcgtgtgcaag
[0631]
seq id no:21:
[0632]
通过将来自人cd89的接触ec1部分内的氨基酸残基(thr58、gln59、arg73、arg74、lys76、his106和tyr107)的人iga与来自食蟹猴cd89的ec1部分的相互氨基酸(ile58、trp59、glu73、lys74、gly76、leu106和ser107)交换的嵌合cd89的氨基酸序列
[0633]
mdpkqttllclvlclgqriqaqegdfpmpfisaksspvipldgsvkiqcqaireayliwlmiiknstyreigeklgfwnetdpefvidhmdankagryqcqyriglsrfrysdtlelvvtglygkpflsadrglvlmpgenisltcssahipfdrfslakegelslpqhqsgehpanfslgpvdlnvsgiyrcygwynrspylwsfpsnalelvvtdsihqdyttqnlirmavaglvlvallailvenwhshtalnkeasadvaepswsqqmcqpgltfartpsvck
[0634]
来自人cd89的信号肽(aa序列1-21),包含来自人cd89的ig样ec1结构域(aa序列22-121;swiss-prot号p24071.1)、除来自食蟹猴cd89的ig样ec1结构域的ile58、trp59、glu73、lys74、gly76、leu106和ser107之外(ncbi参考序列:xp_005590398.1)、来自人cd89的短铰链区(aa序列122-125)、来自人cd89的ig样ec2结构域(aa序列126-220)和来自人cd89的膜近端“接头”区域(aa序列221-227)的嵌合人/食蟹猴cd89细胞外结构域(aa序列22-227),然后是来自人cd89的跨膜结构域(aa序列228-246),以及来自人cd89的短细胞质尾(aa序列247-287),根据ding等人j biol chem 2003;278:27966-27970。
[0635]
seq id no:22:
[0636]
编码通过将来自人cd89的接触ec1部分内的氨基酸残基(thr58、gln59、arg73、arg74、lys76、his106和tyr107)的人iga与来自食蟹猴cd89的ec1部分的相互氨基酸(ile58、trp59、glu73、lys74、gly76、leu106和ser107)交换的嵌合cd89的cdna序列(优化用于哺乳动物表达)
[0637]
atggaccccaagcagaccacactgctgtgcctggtgctgtgtctcggccagagaatccaagctcaagagggcgacttccccatgcctttcatcagcgccaagagcagccctgtgatccctctggatggcagcgtgaagatccagtgccaggccatcagagaggcctacctgatctggctgatgatcattaagaacagcacctaccgcgagatcggcgagaagctcggcttctggaacgagacagaccccgagttcgtgatcgaccacatggacgccaacaaggccggcagataccagtgtcagtaccggatcggcctgagccggttcagatacagcgataccctggaactggtggtcaccggcctgtacggcaagccttttctgtctgccgatagaggactggtgctgatgcccggcgagaacatcagcctgacctgtagctctgctcacatccccttcgacagattcagcctggccaaagaaggcgagctgagcctgcctcagcatcagtctggcgaacaccccgccaacttttctctgggccctgtggacctgaacgtgtccggcatctacagatgctacggctggtacaatcggagcccctacctgtggtctttccccagcaatgccctcgaactggtcgtgaccgatagcatccaccaggactacaccacacagaacctgatcagaatggccgtggccggcctggttctggttgctctgctggctattctggtggaaaactggcacagccacacagccctgaacaaagaggcttctgccgacgtcgccgagccttcttggagtcagcagatgtgtcagcccggcctgaccttcgccagaacacctagcgtgtgcaag
[0638]
seq id no:23:
[0639]
人cd89的ec1部分内的氨基酸序列i(gln22-lys46)
[0640]
qegdfpmpfisaksspvipldgsvk
[0641]
seq id no:24:
[0642]
人cd89的ec1部分内的氨基酸序列ii(ile47-ile71)
[0643]
iqcqaireayltqlmiiknstyrei
[0644]
seq id no:25:
[0645]
人cd89的ec1部分内的氨基酸序列iii(gly72-gly96)
[0646]
grrlkfwnetdpefvidhmdankag
[0647]
seq id no:26:
[0648]
人cd89的ec1部分内的氨基酸序列iv(arg97-gly121)
[0649]
ryqcqyrighyrfrysdtlelvvtg
[0650]
seq id no:27:
[0651]
小鼠抗人cd89特异性抗体8f3的重链可变区的共有氨基酸序列
[0652]
evqlvesggglvkpggslklscaasgftfsnygmswvrqtpdkrlewvatmhsggtytyypdnvkgrftisrdnaknnlylqmsslrsedtamyycaretglydglfdfwgqgttltvss
[0653]
seq id no:28:
[0654]
小鼠抗人cd89特异性抗体8f3的轻链可变区的共有氨基酸序列
[0655]
dvqitqspsylaaspgetitincrasktiskylawyqekpgktnmlliysgstlqsgvpsrfsgsgsgtdftltisslepedlamyycqqhdeypwtfgggtkleik
[0656]
小鼠抗人cd89特异性抗体8f3的互补决定区(cdr):seq id no:29-34
[0657]
seq id no:29:
[0658]
8f3的氨基酸序列重链cdr1
[0659]
nygms
[0660]
seq id no:30:
[0661]
8f3的氨基酸序列重链cdr2
[0662]
tmhsggtytyypdnvkg
[0663]
seq id no:31:
[0664]
8f3的氨基酸序列重链cdr3
[0665]
etglydglfdf
[0666]
seq id no:32:
[0667]
8f3的氨基酸序列轻链cdr1
[0668]
rasktiskyla
[0669]
seq id no:33:
[0670]
8f3的氨基酸序列轻链cdr2
[0671]
sgstlqs
[0672]
seq id no:34:
[0673]
8f3的氨基酸序列轻链cdr3
[0674]
qqhdeypwt
[0675]
seq id no:35:
[0676]
小鼠抗人cd89特异性抗体9h7的重链可变区的共有氨基酸序列
[0677]
qiqlvqsgpelkkpgetvkisckasgytfknygmnwvkqapgkglkwmawintytgepthtddfkgrfvfsletsastaylqinnlknedtatyfcarwglgtteeaardywgqgttltvss
[0678]
seq id no:36:
[0679]
小鼠抗人cd89特异性抗体9h7的轻链可变区的共有氨基酸序列
[0680]
dvvmtqtpltlsvtigqpasmsckssqslldsdgktylnwllqrpgqspksliylvskqdsgvpdrftgsgsgtdftlkisrveaedlgvyycwqgthfpqtfgggtkleik
[0681]
小鼠抗人cd89特异性抗体9h7的互补决定区(cdr):seq id no:37-42
[0682]
seq id no:37:
[0683]
9h7的氨基酸序列重链cdr1
[0684]
nygmn
[0685]
seq id no:38:
[0686]
9h7的氨基酸序列重链cdr2
[0687]
wintytgepthtddfkg
[0688]
seq id no:39:
[0689]
9h7的氨基酸序列重链cdr3
[0690]
wglgtteeaardy
[0691]
seq id no:40:
[0692]
9h7的氨基酸序列轻链cdr1
[0693]
kssqslldsdgktyln
[0694]
seq id no:41:
[0695]
9h7的氨基酸序列轻链cdr2
[0696]
lvskqds
[0697]
seq id no:42:
[0698]
9h7的氨基酸序列轻链cdr3
[0699]
wqgthfpqt
[0700]
seq id no:43:
[0701]
小鼠抗人cd89特异性抗体10e7的重链可变区的共有氨基酸序列
[0702]
evqlvesggglvqpggslklscaasgltfssygmswvrqtpdkrlelvatingngdityypdsvkgrftisrdnakntlylqmsslksedtamyycardydydyamdywgqgtsvtvss
[0703]
seq id no:44:
[0704]
小鼠抗人cd89特异性抗体10e7的轻链可变区的共有氨基酸序列
[0705]
diqmtqstsslsaslgdsvtiscrasqdiinylnwyqqkpdgtvklliyytsrlhsgvpsrfsgsgsgteysltvsnlekediatyfcqqgktlpytfgggtkleik
[0706]
小鼠抗人cd89特异性抗体10e7的互补决定区(cdr):seq id no:45-50
[0707]
seq id no:45:
[0708]
10e7的氨基酸序列重链cdr1
[0709]
sygms
[0710]
seq id no:46:
[0711]
10e7的氨基酸序列重链cdr2
[0712]
tingngdityypdsvkg
[0713]
seq id no:47:
[0714]
10e7的氨基酸序列重链cdr3
[0715]
dydydyamdy
[0716]
seq id no:48:
[0717]
10e7的氨基酸序列轻链cdr1
[0718]
rasqdiinyln
[0719]
seq id no:49:
[0720]
10e7的氨基酸序列轻链cdr2
[0721]
ytsrlhs
[0722]
seq id no:50:
[0723]
10e7的氨基酸序列轻链cdr3
[0724]
qqgktlpyt
[0725]
seq id no:51:
[0726]
小鼠抗人cd89特异性抗体16d6的重链可变区的共有氨基酸序列
[0727]
qvqlqqsgaelmkpgasvkisckatgytfstywiewvkqrpghglewigeilpgsgsanynekfkgkatftadtssntaymqlssltfedsavyycakgfgspyyyamdywgqgtsvtvss
[0728]
seq id no:52:
[0729]
小鼠抗人cd89特异性抗体16d6的轻链可变区的共有氨基酸序列
[0730]
eivltqspalmaaspgekvtitcsvsssisssnlhwyqqksetspkpwiygtsklasgvpvrfsgsgsgtsysltisnmeaedaatyycqqwssfpltfgggtkleik
[0731]
小鼠抗人cd89特异性抗体16d6的互补决定区(cdr):seq id no:53-58
[0732]
seq id no:53:
[0733]
16d6的氨基酸序列重链cdr1
[0734]
tywie
[0735]
seq id no:54:
[0736]
16d6的氨基酸序列重链cdr2
[0737]
eilpgsgsanynekfkg
[0738]
seq id no:55:
[0739]
16d6的氨基酸序列重链cdr3
[0740]
gfgspyyyamdy
[0741]
seq id no:56:
[0742]
16d6的氨基酸序列轻链cdr1
[0743]
svsssisssnlh
[0744]
seq id no:57:
[0745]
16d6的氨基酸序列轻链cdr2
[0746]
gtsklas
[0747]
seq id no:58:
[0748]
16d6的氨基酸序列轻链cdr3
[0749]
qqwssfplt
[0750]
seq id no:59:
[0751]
小鼠抗人cd89特异性抗体26d6的重链可变区的共有氨基酸序列
[0752]
qiqlvqsgpelkkpgetvkisckasgytltkygmnwvkqapgkglkwmgwintytgeptyaddfkgrfa
fsletsastaylqinnlknedtatyfcaslplfgrdsfaywgqgtlvtvsa
[0753]
seq id no:60:
[0754]
小鼠抗人cd89特异性抗体26d6的轻链可变区的共有氨基酸序列
[0755]
dvvmtqtpltlsvtigqpasisckssqslldsdgktylnwllqrpgqspkrliylvskldsgvpdrftgsgsgtdftlkisrveaedlgvyycwqvthfpqtfgggtkleik
[0756]
小鼠抗人cd89特异性抗体26d6的互补决定区(cdr):seq id no:61-66
[0757]
seq id no:61:
[0758]
26d6的氨基酸序列重链cdr1
[0759]
kygmn
[0760]
seq id no:62:
[0761]
26d6的氨基酸序列重链cdr2
[0762]
wintytgeptyaddfkg
[0763]
seq id no:63:
[0764]
26d6的氨基酸序列重链cdr3
[0765]
lplfgrdsfay
[0766]
seq id no:64:
[0767]
26d6的氨基酸序列轻链cdr1
[0768]
kssqslldsdgktyln
[0769]
seq id no:65:
[0770]
26d6的氨基酸序列轻链cdr2
[0771]
lvsklds
[0772]
seq id no:66:
[0773]
26d6的氨基酸序列轻链cdr3
[0774]
wqvthfpqt
[0775]
seq id no:67:
[0776]
小鼠抗人cd89特异性抗体20b4的重链可变区的共有氨基酸序列
[0777]
qvqlkesgpglvapsqslsitctvsgfsltsygvhwvrqppgkglewlgiiwaggstsynsalmsrlsiskdnsksqvflkmnslqtddtamyycardhydlfaywgqgtlvtvsa
[0778]
seq id no:68:
[0779]
小鼠抗人cd89特异性抗体20b4的轻链可变区的共有氨基酸序列
[0780]
dilmtqspssmsvslgdtvsitchasqdisrnigwlqqkpgksfkgliyhgtnledgvpsrfsgsgsgadysltisslesedfadyycvqyaqfprtfgggtkleik
[0781]
小鼠抗人cd89特异性抗体20b4的互补决定区(cdr):seq id no:69-74
[0782]
seq id no:69:
[0783]
20b4的氨基酸序列重链cdr1
[0784]
sygvh
[0785]
seq id no:70:
[0786]
20b4的氨基酸序列重链cdr2
[0787]
iiwaggstsynsalms
[0788]
seq id no:71:
[0789]
20b4的氨基酸序列重链cdr3
[0790]
dhydlfay
[0791]
seq id no:72:
[0792]
20b4的氨基酸序列轻链cdr1
[0793]
hasqdisrnig
[0794]
seq id no:73:
[0795]
20b4的氨基酸序列轻链cdr2
[0796]
hgtnled
[0797]
seq id no:74:
[0798]
20b4的氨基酸序列轻链cdr3
[0799]
vqyaqfprt
[0800]
seq id no:75:
[0801]
小鼠抗人cd89特异性抗体30c7的重链可变区的共有氨基酸序列
[0802]
qvqlqqsgpelvrpgvsvkisckgsgytftdyvmhwvkqshakslewigvistysgntnynqkfkgkatmtvdkssstaymelarltsedsaiyycaresdgyyfdywgqgttltvss
[0803]
seq id no:76:
[0804]
小鼠抗人cd89特异性抗体30c7的轻链可变区的共有氨基酸序列
[0805]
divmsqspsslavsagekvtmsckssqsllnsrtrknylawyqqkpgqspklliywastresgvpdrftgsgsgtdftltissvqaedlavyyckqsynlmytfgggtkleik
[0806]
小鼠抗人cd89特异性抗体30c7的互补决定区(cdr):seq id no:77-82
[0807]
seq id no:77:
[0808]
30c7的氨基酸序列重链cdr1
[0809]
dyvmh
[0810]
seq id no:78:
[0811]
30c7的氨基酸序列重链cdr2
[0812]
vistysgntnynqkfkg
[0813]
seq id no:79:
[0814]
30c7的氨基酸序列重链cdr3
[0815]
esdgyyfdy
[0816]
seq id no:80:
[0817]
30c7的氨基酸序列轻链cdr1
[0818]
kssqsllnsrtrknyla
[0819]
seq id no:81:
[0820]
30c7的氨基酸序列轻链cdr2
[0821]
wastres
[0822]
seq id no:82:
[0823]
30c7的氨基酸序列轻链cdr3
[0824]
kqsynlmyt
[0825]
seq id no:83:
[0826]
编码嵌合小鼠vh 8f3人重igg4链的cdna序列
[0827]
atggagctgggcctgagctggatttttctgctggccatcctgaagggcgtgcagtgcgaagtgcagctggttgaatctggcggcggactggttaagcctggcggatctctgaagctgagctgtgccgccagcggcttcaccttcagcaattacggcatgagctgggtccgacagacccctgacaagagactggaatgggtcgccacaatgcacagcggcggcacctacacctactatcccgacaacgtgaagggcagattcaccatcagccgggacaacgccaagaacaacctgtacctgcagatgagcagcctgcggagcgaggataccgccatgtactactgcgccagagaaaccggcctgtacgacggcctgttcgatttttggggccagggcaccacactgaccgtgtctagcgcctctacaaagggccctagcgtgttccctctggctccttgtagcagaagcaccagcgagtctacagccgctctgggctgtctggtcaaggactactttcccgagcctgtgacagtgtcctggaactctggcgctctgacaagcggcgtgcacacatttccagccgtgctgcaaagcagcggcctgtattctctgagcagcgtggtcacagtgcccagctctagcctgggcaccaagacctacacatgcaatgtggaccacaagcctagcaacaccaaggtggacaagcgcgtggaatctaagtacggccctccttgtcctccatgtcctgcacctgagtttctcggcggaccctccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggacgtttcccaagaggaccctgaggtgcagttcaattggtacgtggacggcgtggaagtgcacaatgccaagaccaagcctagagaggaacagttcaacagcacctacagagtggtgtccgtgctgacagtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgcctagcagcatcgagaaaaccatcagcaaggccaagggccagccaagagaaccccaggtgtacacactgcctccaagccaagaggaaatgaccaagaaccaggtgtccctgacctgcctggttaagggcttctacccctccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagaccacacctcctgtgctggacagcgacggctcattcttcctgtacagcaggctgaccgtggacaagagcagatggcaagagggcaacgtgttcagctgcagcgtgatgcacgaggccctgcacaaccactacacccagaagtctctgagcctgtctctgggcaag
[0828]
seq id no:84:
[0829]
编码嵌合小鼠vh 10e7人重igg4链的cdna序列
[0830]
atggagctgggcctgagctggatttttctgctggccatcctgaagggcgtgcagtgcgaagtgcagctggttgaatctggcggaggactggttcagcctggcggatctctgaagctgtcttgtgccgcctctggcctgacctttagcagctacggcatgagctgggtccgacagacccctgacaagagactggaactggtggccacaatcaacggcaacggcgacatcacctactatcccgacagcgtgaagggcagattcaccatcagccgggacaacgccaagaacaccctgtacctgcagatgagcagcctgaagtccgaggacaccgccatgtactactgcgccagagactacgactacgattacgctatggactactggggccagggcaccagcgtgacagttagctctgcctctacaaagggccctagcgtgttccctctggctccttgtagcagaagcaccagcgagtctacagccgctctgggctgtctggtcaaggactactttcccgagcctgtgaccgtgtcctggaattctggcgctctgacaagcggcgtgcacacctttccagctgtgctgcaaagcagcggcctgtactctctgagcagcgtggtcacagtgcctagctctagcctgggcaccaagacctacacctgtaatgtggaccacaagcctagcaacaccaaggtggacaagcgcgtggaatctaagtacggccctccttgtcctccatgtcctgctccagagtttctcggcggaccctccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggacgtttcccaagaggaccctgaggtgcagttcaattggtacgtggacggcgtggaagtgcacaatgccaagaccaagcctagagaggaacagttcaacagcacctacagagtggtgtccgtgctgaccgtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgcctagcagcatcgagaaaaccatcagcaaggccaagggccagccaagagaaccccaggtgtacacactgcctccaagccaagaggaaatgaccaagaaccaggtgtccctgacctgcctggttaagggcttctacccctccgatatcgccgtggaatgggagagcaatgg
ccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcagactgaccgtggacaagagcagatggcaagagggcaacgtgttcagctgcagcgtgatgcacgaggccctgcacaaccactacacccagaagtctctgagcctgtctctgggcaag
[0831]
seq id no:85:
[0832]
编码嵌合小鼠vh 16d6人重igg4链的cdna序列
[0833]
atggagctgggcctgagctggatttttctgctggccatcctgaagggcgtgcagtgtcaggttcagctgcagcagtctggcgccgagcttatgaagcctggcgcctctgtgaagatcagctgcaaggccaccggctacaccttcagcacctactggatcgagtgggtcaagcagaggcctggccacggactggaatggatcggagagatcctgcctggcagcggcagcgccaactacaacgagaagttcaagggcaaagccaccttcaccgccgacaccagcagcaacacagcctacatgcagctgagcagcctgaccttcgaggacagcgccgtgtactactgcgccaaaggcttcggcagcccctactactacgctatggattactggggccagggcaccagcgtgacagtgtctagcgcctctacaaagggccctagcgtgttccctctggctccttgtagcagaagcaccagcgagtctacagccgctctgggctgtctggtcaaggactactttcccgagcctgtgaccgtgtcctggaattctggcgctctgacaagcggcgtgcacacctttccagctgtgctgcaaagcagcggcctgtactctctgagcagcgtggtcacagtgcctagctctagcctgggcaccaagacctacacctgtaatgtggaccacaagcctagcaacaccaaggtggacaagcgcgtggaatctaagtacggccctccttgtcctccatgtcctgctccagagtttctcggcggaccctccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggacgtttcccaagaggaccctgaggtgcagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagttcaactccacctacagagtggtgtccgtgctgaccgtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgcctagcagcatcgagaaaaccatcagcaaggccaagggccagccaagagaaccccaggtgtacacactgcctccaagccaagaggaaatgaccaagaaccaggtgtccctgacctgcctcgtgaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcagactgaccgtggacaagagcagatggcaagagggcaacgtgttctcctgcagcgtgatgcacgaggccctgcacaaccactacacccagaagtctctgagcctgtctctgggcaag
[0834]
seq id no:86:
[0835]
编码嵌合小鼠vh 20b4人重igg4链的cdna序列
[0836]
atggagctgggcctgagctggatttttctgctggccatcctgaagggcgtgcagtgcagagtgcagctgaaagagtctggccctggactggtggccccaagccagtctctgagcatcacctgtaccgtgtccggcttcagcctgacaagctatggcgtgcactgggtccgacagcctccaggcaaaggactggaatggctgggcatcatttgggctggcggcagcaccagctacaacagcgctctgatgagccggctgagcatctccaaggacaacagcaagagccaggtgttcctgaagatgaacagcctgcagaccgacgacaccgccatgtactactgcgccagagatcactacgacctgttcgcctattggggccagggcacactggttacagtgtccgccgcctctacaaagggccctagtgtgtttcctctggctccctgcagcagaagcaccagcgaatctacagccgctctgggctgcctggtcaaggactactttcctgagccagtgaccgtgtcctggaactctggcgctctgacaagcggcgtgcacacatttccagccgtgctgcaaagcagcggcctgtactctctgtccagcgtggtcacagtgcctagctctagcctgggcaccaagacctacacctgtaatgtggaccacaagcctagcaacaccaaggtggacaagcgcgtggaatctaagtacggccctccttgtcctccatgtcctgcacctgagtttctcggcggaccctccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggacgtttcccaagaggaccctgaggtgcagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagttcaacagcacctacagagtggtgtccgtgctgacagtgctgcaccagga
ttggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgcctagcagcatcgagaaaaccatcagcaaggccaagggccagccaagagaaccccaggtgtacacactgcctccaagccaagaggaaatgaccaagaatcaggtgtccctgacctgcctcgtgaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcagactgaccgtggacaagagcagatggcaagagggcaacgtgttcagctgctccgtgatgcacgaggccctgcacaaccactacacccagaaaagcctgagcctgtctctgggcaag
[0837]
seq id no:87:
[0838]
编码嵌合小鼠vh 30c7人重igg4链的cdna序列
[0839]
atggagctgggcctgagctggatttttctgctggccatcctgaagggcgtgcagtgtcaggttcagctgcagcagtctggccccgaacttgtcagacctggcgtgtccgtgaagatcagctgtaaaggcagcggctacaccttcaccgactacgtgatgcactgggtcaagcagagccacgccaagagcctggaatggatcggcgtgatcagcacctacagcggcaacaccaactacaaccagaagttcaagggcaaagccaccatgaccgtggacaagagcagcagcaccgcctacatggaactggccaggctgacctctgaggacagcgccatctactactgcgccagagagagcgacggctactacttcgattattggggccagggcaccacactgaccgtgtctagcgcctctacaaagggccctagcgtgttccctctggctccttgtagcagaagcaccagcgagtctacagccgctctgggctgtctggtcaaggactactttcccgagcctgtgacagtgtcctggaactctggcgctctgacaagcggcgtgcacacatttccagccgtgctgcaaagcagcggcctgtactctctgagcagcgtggtcacagtgcctagctctagcctgggcaccaagacctacacctgtaatgtggaccacaagccttccaacaccaaggtggacaagcgcgtggaatctaagtacggccctccttgtcctccatgtcctgcacctgagtttctcggcggaccctccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggacgtttcccaagaggaccctgaggtgcagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagttcaactccacctacagagtggtgtccgtgctgacagtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgcctagcagcatcgagaaaaccatcagcaaggccaagggccagccaagagaaccccaggtgtacacactgcctccaagccaagaggaaatgaccaagaaccaggtgtccctgacctgcctcgtgaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcagactgacagtggataagagccggtggcaagagggcaacgtgttctcctgctctgtgatgcacgaggccctgcacaaccactacacccagaaaagcctgagcctgtctctgggcaag
[0840]
seq id no:88:
[0841]
编码嵌合小鼠vl 8f3人轻κ链的cdna序列
[0842]
atggacatgagagttcccgctcagctgctgggactgctgctgctttggtttcctggcgctagatgcgacgtgcagatcacacagagccctagctacctggctgcctctcctggcgagacaatcaccatcaactgccgggccagcaagaccatcagcaagtacctggcctggtatcaagagaagcccggcaagaccaacatgctgctgatctacagcggcagcacactgcagagcggagtgcctagcagattttccggctctggcagcggcaccgatttcaccctgaccataagcagcctggaacctgaggacctggccatgtactactgccagcagcacgacgagtacccctggacatttggcggaggcaccaagctggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctgcagtccggcaatagccaagagagcgtgaccgagcaggacagcaaggactctacctacagcctgagcagcaccctgacactgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgaatgt
[0843]
seq id no:89:
[0844]
编码嵌合小鼠vl 10e7人轻κ链的cdna序列
[0845]
atggacatgagagttcccgctcagctgctgggactgctgctgctttggtttcctggcgctagatgcgacatccagatgacccagagcaccagcagcctgtctgcctctctgggcgatagcgtgaccatcagctgtagagccagccaggacatcatcaactacctgaactggtatcagcagaaacccgacggcaccgtgaagctgctgatctactacaccagcagactgcacagcggcgtgcccagcagattttctggctctggaagcggcaccgagtacagcctgaccgtgtccaacctggaaaaagaggatatcgctacctacttctgccagcaaggcaagaccctgccttacacctttggcggaggcaccaagctggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctgcagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactctacctatagcctgagcagcaccctgacactgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgaatgt
[0846]
seq id no:90:
[0847]
编码嵌合小鼠vl 16d6人轻κ链的cdna序列
[0848]
atggacatgagagttcccgctcagctgctgggactgctgctgctttggtttcctggcgctagatgcgagatcgtgctgacacagagccctgctctgatggctgcttcccctggcgagaaagtgaccatcacctgtagcgtgtccagcagcatcagcagctccaacctgcactggtatcagcagaagtccgagacaagccccaagccttggatctacggcacaagcaaactggccagcggcgtgccagtcagattttctggctctggcagcggcaccagctacagcctgaccatcagcaacatggaagccgaggatgccgccacctactactgccagcagtggtccagctttccactgacctttggcggaggcaccaagctggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcaatggaaggtggacaacgccctgcagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactccacctatagcctgagcagcaccctgacactgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgaatgt
[0849]
seq id no:91:
[0850]
编码嵌合小鼠vl 20b4人轻κ链的cdna序列
[0851]
atggacatgagagttcccgctcagctgctgggactgctgctgctttggtttcctggcgctagatgcgacatcctgatgacacagagccccagctccatgtccgtgtctctgggcgataccgtgtccatcacatgtcacgccagccaggacatcagccggaatatcggatggctgcagcagaagcccggcaagagctttaagggcctgatctaccacggcaccaacctggaagatggcgtgcccagcagattttccggctctggatctggcgccgactacagcctgacaatcagcagcctggaaagcgaggacttcgccgattactactgcgtgcagtacgcccagtttcctcggacatttggcggaggcacaaagctggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctgcagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactccacctatagcctgagcagcaccctgacactgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgaatgt
[0852]
seq id no:92:
[0853]
编码嵌合小鼠vl 30c7人轻κ链的cdna序列
[0854]
atggacatgagagttcccgctcagctgctgggactgctgctgctttggtttcctggcgctagatgcga
catcgtgatgtctcagagccctagcagcctggctgtgtctgccggcgagaaagtgaccatgagctgcaagagcagccagagcctgctgaacagccggaccagaaagaactacctggcctggtatcagcagaagcccggacagtctcccaagctgctgatctactgggccagcaccagagaaagcggcgtgcccgatagattcacaggcagcggcagcggaaccgacttcaccctgacaatcagctctgtgcaggccgaggatctggccgtgtactactgcaagcagagctacaacctgatgtacaccttcggcggaggcaccaagctggaaatcaagagaacagtggccgctcctagcgtgttcatcttcccaccttccgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctcaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctgcagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactctacctacagcctgagcagcacactgaccctgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgaatgt
[0855]
seq id no:93:
[0856]
嵌合小鼠vh 8f3人重igg4链的氨基酸序列
[0857]
melglswifllailkgvqcevqlvesggglvkpggslklscaasgftfsnygmswvrqtpdkrlewvatmhsggtytyypdnvkgrftisrdnaknnlylqmsslrsedtamyycaretglydglfdfwgqgttltvssastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[0858]
seq id no:94:
[0859]
嵌合小鼠vh 10e7人重igg4链的氨基酸序列
[0860]
melglswifllailkgvqcevqlvesggglvqpggslklscaasgltfssygmswvrqtpdkrlelvatingngdityypdsvkgrftisrdnakntlylqmsslksedtamyycardydydyamdywgqgtsvtvssastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[0861]
seq id no:95:
[0862]
嵌合小鼠vh 16d6人重igg4链的氨基酸序列
[0863]
melglswifllailkgvqcqvqlqqsgaelmkpgasvkisckatgytfstywiewvkqrpghglewigeilpgsgsanynekfkgkatftadtssntaymqlssltfedsavyycakgfgspyyyamdywgqgtsvtvssastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[0864]
seq id no:96:
[0865]
嵌合小鼠vh 20b4人重igg4链的氨基酸序列
[0866]
melglswifllailkgvqcrvqlkesgpglvapsqslsitctvsgfsltsygvhwvrqppgkglewlgi
iwaggstsynsalmsrlsiskdnsksqvflkmnslqtddtamyycardhydlfaywgqgtlvtvsaastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[0867]
seq id no:97:
[0868]
嵌合小鼠vh 30c7人重igg4链的氨基酸序列
[0869]
melglswifllailkgvqcqvqlqqsgpelvrpgvsvkisckgsgytftdyvmhwvkqshakslewigvistysgntnynqkfkgkatmtvdkssstaymelarltsedsaiyycaresdgyyfdywgqgttltvssastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[0870]
seq id no:98:
[0871]
嵌合小鼠vl 8f3人轻κ链的氨基酸序列
[0872]
mdmrvpaqllgllllwfpgarcdvqitqspsylaaspgetitincrasktiskylawyqekpgktnmlliysgstlqsgvpsrfsgsgsgtdftltisslepedlamyycqqhdeypwtfgggtkleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0873]
seq id no:99:
[0874]
嵌合小鼠vl 10e7人轻κ链的氨基酸序列
[0875]
mdmrvpaqllgllllwfpgarcdiqmtqstsslsaslgdsvtiscrasqdiinylnwyqqkpdgtvklliyytsrlhsgvpsrfsgsgsgteysltvsnlekediatyfcqqgktlpytfgggtkleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0876]
seq id no:100:
[0877]
嵌合小鼠vl 16d6人轻κ链的氨基酸序列
[0878]
mdmrvpaqllgllllwfpgarceivltqspalmaaspgekvtitcsvsssisssnlhwyqqksetspkpwiygtsklasgvpvrfsgsgsgtsysltisnmeaedaatyycqqwssfpltfgggtkleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0879]
seq id no:101:
[0880]
嵌合小鼠vl 20b4人轻κ链的氨基酸序列
[0881]
mdmrvpaqllgllllwfpgarcdilmtqspssmsvslgdtvsitchasqdisrnigwlqqkpgksfkgliyhgtnledgvpsrfsgsgsgadysltisslesedfadyycvqyaqfprtfgggtkleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0882]
seq id no:102:
[0883]
嵌合小鼠vl 30c7人轻κ链的氨基酸序列
[0884]
mdmrvpaqllgllllwfpgarcdivmsqspsslavsagekvtmsckssqsllnsrtrknylawyqqkpgqspklliywastresgvpdrftgsgsgtdftltissvqaedlavyyckqsynlmytfgggtkleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0885]
seq id no:103:pcr引物
[0886]
atgagtgtgc tcactcaggt cctggsgttg
[0887]
seq id no:104:pcr引物
[0888]
atgaggrccc ctgctcagwt tyttggmwtc ttg
[0889]
seq id no:105:pcr引物
[0890]
atggatttwc aggtgcagat twtcagcttc
[0891]
seq id no:106:pcr引物
[0892]
atgggcwtca aagatggagt caca
[0893]
seq id no:107:pcr引物
[0894]
atggtrtccw casctcagtt ccttg
[0895]
seq id no:108:pcr引物
[0896]
actggatggt gggaagatgg
[0897]
seq id no:109:pcr引物
[0898]
atgggatgga gctrtatcat sytctt
[0899]
seq id no:110:pcr引物
[0900]
atgractttg ggytcagctt grttt
[0901]
seq id no:111:pcr引物
[0902]
atggcttgtc yttrgsgctr ctcttctgc
[0903]
seq id no:112:pcr引物
[0904]
atggratgga gckgggtctt tmtctt
[0905]
seq id no:113:pcr引物
[0906]
atggmttggg tgtggamctt gct
[0907]
seq id no:114:pcr引物
[0908]
cagtggatag acagatgggg g
[0909]
seq id no:115:pcr引物
[0910]
aagatggata cagttggtgc
[0911]
seq id no:116:pcr引物
[0912]
gasrthstga tgacccagac ncc
[0913]
seq id no:117:
[0914]
人源化抗人cd89抗体10e7重链可变区vh1的氨基酸序列
[0915]
1evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglewvst ingngdityy
[0916]
61pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0917]
seq id no:118:
[0918]
人源化抗人cd89抗体10e7重链可变区vh2的氨基酸序列
[0919]
1evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglewvat ingngdityy
[0920]
61pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0921]
seq id no:119:
[0922]
人源化抗人cd89抗体10e7重链可变区vh3的氨基酸序列
[0923]
1evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglelvat ingngdityy
[0924]
61pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0925]
seq id no:120:
[0926]
人源化抗人cd89抗体10e7轻链可变区vl1的氨基酸序列
[0927]
1diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gkapklliyy tsrlhsgvps
[0928]
61rfsgsgsgtd ftltisslqp edfatyycqq gktlpytfgq gtkleik
[0929]
seq id no:121:
[0930]
人源化抗人cd89抗体10e7轻链可变区vl2的氨基酸序列
[0931]
1diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gkapklliyy tsrlhsgvps
[0932]
61rfsgsgsgtd ytltisslqp edfatyycqq gktlpytfgq gtkleik
[0933]
seq id no:122:
[0934]
人源化抗人cd89抗体10e7轻链可变区vl3的氨基酸序列
[0935]
1diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gkavklliyy tsrlhsgvps
[0936]
61rfsgsgsgtd ytltisslqp edfatyycqq gktlpytfgq gtkleik
[0937]
seq id no:123:
[0938]
人源化抗人cd89抗体10e7轻链可变区vl4的氨基酸序列
[0939]
1diqmtqspss lsasvgdrvt itcrasqdii nylnwyqqkp gktvklliyy tsrlhsgvps
[0940]
61rfsgsgsgtd ytltisslqp edfatyfcqq gktlpytfgq gtkleik
[0941]
seq id no:124:
[0942]
编码覆盖人源化vh1的抗人cd89抗体10e7的人源化igg4链的cdna序列
[0943]
[0944][0945]
seq id no:125:
[0946]
编码覆盖人源化vh2的抗人cd89抗体10e7的人源化igg4链的cdna序列
[0947]
[0948][0949]
seq id no:126:
[0950]
编码覆盖人源化vh3的抗人cd89抗体10e7的人源化igg4链的cdna序列
[0951]
[0952][0953]
seq id no:127:
[0954]
编码覆盖人源化vl1的抗人cd89抗体10e7的人源化κ链的cdna序列
[0955][0956]
seq id no:128:
[0957]
编码覆盖人源化vl2的抗人cd89抗体10e7的人源化κ链的cdna序列
[0958][0959]
[0960]
seq id no:129:
[0961]
编码覆盖人源化vl3的抗人cd89抗体10e7的人源化κ链的cdna序列
[0962][0963]
seq id no:130:
[0964]
编码覆盖人源化vl4的抗人cd89抗体10e7的人源化κ链的cdna序列
[0965][0966][0967]
seq id no:131:
[0968]
编码覆盖人源化vh1的抗人cd89抗体10e7的人源化igg4链的氨基酸序列
[0969][0970]
seq id no:132:
[0971]
编码覆盖人源化vh2的抗人cd89抗体10e7的人源化igg4链的氨基酸序列
[0972][0973]
seq id no:133:
[0974]
编码覆盖人源化vh3的抗人cd89抗体10e7的人源化igg4链的氨基酸序列
[0975][0976][0977]
seq id no:134:
[0978]
编码覆盖人源化vl1的抗人cd89抗体10e7的人源化κ链的氨基酸序列
[0979][0980]
seq id no:135:
[0981]
编码覆盖人源化vl2的抗人cd89抗体10e7的人源化κ链的氨基酸序列
[0982][0983]
seq id no:136:
[0984]
编码覆盖人源化vl3的抗人cd89抗体10e7的人源化κ链的氨基酸序列
[0985][0986]
seq id no:137:
[0987]
编码覆盖人源化vl4的抗人cd89抗体10e7的人源化κ链的氨基酸序列
[0988][0989][0990]
seq id no:138:
[0991]“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7重链可变区vh3sq的氨基酸序列
[0992]
1evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglelvat isgqgdityy
[0993]
61pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0994]
seq id no:139:
[0995]“cdr2脱酰胺修复的”人源化抗人cd89抗体10e7重链可变区vh3st的氨基酸序列
[0996]
1evqllesggg lvqpggslrl scaasgltfs sygmswvrqa pgkglelvat isgtgdityy
[0997]
61pdsvkgrfti srdnskntly lqmnslraed tavyycardy dydyamdywg qgtlvtvss
[0998]
seq id no:140:
[0999]
编码覆盖人源化vh3sq的抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化igg4链的cdna序列
[1000][1001][1002]
seq id no:141:
[1003]
编码覆盖人源化vh3st的抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化igg4链的cdna序列
[1004][1005][1006]
seq id no:142:
[1007]
编码覆盖人源化vh3sq的抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化igg4链的氨基酸序列
[1008]
[1009]
seq id no:143:
[1010]
编码覆盖人源化vh3st的抗人cd89抗体10e7的“cdr2脱酰胺修复的”人源化igg4链的氨基酸序列
[1011][1012]
pct
[1013]
输出(原始电子形式)
[1014]
[1015][1016]
由受理局填写
[1017][1018]
由国际局填写
[1019]
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。