:
1.本发明属于食品保鲜技术领域,特别涉及一种筛选自发酵核桃粕的抗菌多肽及其应用。
背景技术:
2.研究报道果蔬在供应链中采后部分的损失和浪费高达13%~38%,因此,亟待进行相关研究以延长果蔬的货架期,从而减少采后损失。目前关于果蔬采后保鲜的研究侧重于加入一些抗菌剂如有机酸(苯甲酸和山梨酸等)、酶(溶菌酶)、细菌素(nisin)和多酚(姜黄素和槲皮素等)等,但现有的天然抗菌剂存在成本高和作用靶向不明确等问题。
3.基于实验室前期研究,核桃粕发酵可以通过微生物和酶产生代谢产物如有机酸、挥发物、多肽和游离氨基酸等,这些代谢物可以穿过微生物的细胞膜,在细胞质中积累以发挥抗菌活性。另外,通常植物提取物发挥生物活性并不是因为单一成分的存在,而是由多种活性成分相互作用而产生的效果。
4.因此,本技术将制备对刺梨腐败优势菌具有抗菌活性的发酵物,并利用其开发天然且低成本的抗菌产品,以期延长食品特别是果蔬的贮藏期,为发酵核桃粕在采后果蔬保鲜中的应用提供理论基础。
技术实现要素:
5.本研究采用复合益生菌发酵核桃粕以提高对刺梨腐败优势菌的抗菌活性,探究发酵核桃粕中具有抗菌活性的多肽的变化,并从中筛选具有抗菌活性的多肽,分析抗菌多肽与酚类化合物之间的互作机理,并获得最优的抗菌组合。
6.为解决上述技术问题,本发明以对刺梨腐败优势菌维多利亚青霉(penicillium.victoriae)的抑菌效果为指标,采用枯草芽孢杆菌(bacillus subtilis)与植物乳杆菌(lactiplantibacillus plantarum)复合发酵核桃粕。探究发酵前后酚类化合物的变化,筛选发酵核桃粕中对p.victoriae抗菌活性最强的酚类化合物。比较不同分子量发酵核桃粕的抗菌活性,采用lc-ms/ms对抗菌活性最强的发酵核桃粕组分鉴定多肽序列,pyrx软件虚拟筛选,人工合成肽段并验证抗菌活性。通过maestro软件分子对接探究抗菌肽与rho1gtpase晶体结构(pdb id:3a58)的结合方式。采用二倍稀释法复配抗菌肽与酚类化合物,测定复合物对p.victoriae的抗菌活性,并通过傅里叶红外光谱(ftir)、荧光光谱、分子对接探究抗菌多肽与酚类化合物之间的互作机理。
7.本发明提供的技术方案之一,是提取自发酵核桃粕的抗菌肽,所述抗菌肽自n端至c端的氨基酸序列分别为phe-gly-gly-asp-ser-thr-his-pro(fggdsthp),ala-leu-gly-gly-gly-tyr(algggy),tyr-val-val-pro-trp(yvvpw)和pro-leu-leu-arg-trp(pllrw);
8.进一步地,抗菌肽yvvpw对p.victoriae的抗菌活性最强,mic为16.0mg/ml;同时对s.aureus具有抗菌活性;
9.进一步地,抗菌肽fggdsthp,对p.victoriae具有抗菌活性;
10.进一步地,抗菌肽algggy,对p.victoriae和s.aureus均具有抗菌活性;
11.进一步地,抗菌肽pllrw,对s.aureus具有抗菌活性。
12.本发明提供的技术方案之二,是一种抗菌组合物,所述抗菌组合物包括抗菌肽yvvpw和水杨酸(sa);
13.进一步地,组合物中抗菌肽yvvpw的浓度为1.0mg/ml~4.0mg/ml,sa的浓度为0.27mg/ml~1.10mg/ml;
14.优选地,组合物中抗菌肽yvvpw的浓度为1.0mg/ml,sa的浓度为0.27mg/ml,此时二者组合的协同效果最好,分级抑菌浓度指数为0.07,对p.victoriae的抑菌圈达到7.35
±
0.20mm。
15.优选地,组合物中抗菌肽yvvpw的浓度为2.0mg/ml,sa的浓度为0.55mg/ml,分级抑菌浓度指数fic为0.25,此时二者组合对p.victoriae的抗菌活性最高,对p.victoriae的抑菌圈达到11.50
±
0.41mm。
16.本发明提供的技术方案之三,是技术方案一所述抗菌肽或技术方案二所述抗菌组合物的应用,特别是在食品保鲜,更特别地是蔬果保鲜中的应用,更进一步地,是在刺梨保鲜中的应用。
17.有益效果:
18.本发明提供了一组对刺梨腐败优势菌有抑制效果的发酵核桃粕抗菌肽,抗菌作用明显,且采用核桃粕为原料,生产成本低,经济效益高。
19.基于试验结果,发酵核桃粕中的酚类化合物中sa对p.victoriae抗菌活性最高(mic为4.40mg/ml)。超滤发酵核桃粕,对p.victoriae抗菌活性测定结果为分子量在0~3kda的发酵核桃粕的抗菌活性最高(冻干粉mic为10μg/ml),fggdsthp、algggy和yvvpw等多肽均对p.victoriae具有抗菌活性,pllrw对s.aureus具有抗菌活性。其中合成的短肽序列yvvpw被证明对p.victoriae有较高的抗菌活性(mic为16.0mg/ml)。yvvpw与sa联合发挥协同抑菌作用时的浓度分别为1.0~4.0mg/ml的yvvpw与0.27~1.10mg/ml的sa,二者对p.victoriae有协同抗菌活性。sa的加入使yvvpw的trp和tyr残基发生荧光猝灭,氢键、疏水相互作用、π-π堆积是二者之间的主要相互作用力。
附图说明:
20.图1未发酵核桃粕冻干粉负离子(a)和正离子(b)色谱图;
21.其中,1.没食子酸;2.水杨酸;3.原儿茶酸;4.水杨苷;5.绿原酸;6.原花青素;7.(-)-表儿茶素;8.异槲皮苷;9.金丝桃苷;10.咖啡酸;11.鞣花酸;12.反式阿魏酸;13.根皮苷;14.柚皮素-7-o-葡萄糖苷;15.柚皮素。
22.图2发酵核桃粕冻干粉负离子(a)和正离子(b)色谱图;
23.其中,1.没食子酸;2.水杨酸;3.原儿茶酸;4.水杨苷;5.绿原酸;6.原花青素;7.(-)-表儿茶素;8.异槲皮苷;9.金丝桃苷;10.咖啡酸;11.鞣花酸;12.反式阿魏酸;13.根皮苷;14.柚皮素-7-o-葡萄糖苷;15.柚皮素。
24.图3 0~3kda发酵核桃粕冻干粉的液质分析基峰图。
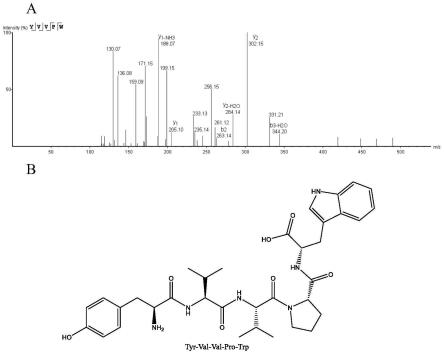
25.图4肽段yvvpw的二级质谱图(a)和化学结构(b)。
26.图5肽段yvvpw与rho1 gtpase结合的三维结构(a),活性位点的结合方式(b)和最
佳构象相互作用示意平面图(c)。
27.图6不同浓度sa存在时多肽yvvpw的荧光光谱图;
28.图7yvvpw、sa及yvvpw-sa的傅里叶红外光谱。
29.图8yvvpw与sa的结合方式,(a)复合体的静电表面;(b)活性位点的结合方式。
具体实施方式:
30.为了使本专利的目的、技术方案及优点更加清楚明白,以下结合具体实施例,对本专利进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本专利,并不用于限定本发明。
31.本发明所述的多肽,本领域技术人员可通过人工合成的方式获得,也可通过对核桃粕发酵筛选获得。
32.本发明发酵核桃粕所使用的植物乳杆菌为植物乳杆菌(lactiplantibacillus plantarum)cicc 23121,本发明所使用的枯草芽孢杆菌为枯草芽孢杆菌(bacillus subtilis)cicc 10002。
33.本发明以b.subtilis与l.plantarum在发酵温度35℃的条件下发酵核桃粕26h,定性与定量分析发酵核桃粕酚类化合物结构,检测其对维多利亚青霉(penicillium victoriae),金黄色葡萄球菌(staphylococcus aureus)和大肠杆菌(escherichia coli)的抗菌活性,得到抗真菌活性最高的sa;发酵核桃粕经过超滤分离,得到不同分子量范围(0~3kda、3~5kda、5~10kda、》10kda)的发酵核桃粕组分,检测其对p.victoriae,s.aureus和e.coli的抗菌活性,得到超滤后抗菌活性最高的0~3kda发酵核桃粕;测定发酵核桃粕冻干粉多肽的结构,通过分子对接技术从143条多肽中筛选出与rho1 gtpase晶体结构(pdb id:3a58)对接时对接能较低的多肽序列,继而对相应的多肽进行人工合成,检测其对p.victoriae,s.aureus和e.coli的抗菌活性,得到发酵核桃粕中高抗菌短肽序列fggdsthp、algggy、yvvpw和pllrw。采用分子对接技术将抗菌活性最高的多肽yvvpw与rho1 gtpase进行分子对接,结果表明二者紧密结合,相互作用力主要包括氢键、疏水相互作用和π-π堆积。
34.本发明涉及的部分试验方法如下:
35.(1)抑菌圈试验:
36.刺梨腐败优势菌为维多利亚青霉(penicillium victoriae),以对维多利亚青霉(p.victoriae)、大肠杆菌(escherichia coli)和金黄色葡萄球菌(staphylococcus aureus)的抑菌效果为筛选指标,测定不同组分的抑菌活性。吸取100μl浓度105cfu/ml的p.victoriae孢子悬液于pda培养基涂布,100μl浓度105cfu/ml的s.aureus与e.coli孢子悬液于lb培养基涂布。利用无菌牛津杯打孔并加入100μl发酵核桃粕(测定其他成分的抑菌圈时进行替换即可),以无菌水作阴性对照和纳他霉素作为阳性对照。将pda和lb平板置于28℃培养箱分别培养5d和24h后,测量抑菌圈直径。
37.(2)发酵核桃粕的多酚含量测定
38.参考folin和ciocalteu(1927)方法,精密称取没食子酸10mg,用超纯水溶解并定容至100ml容量瓶中,摇匀备用,即为没食子酸标准溶液。准确吸取标准溶液0、0.5、1.0、1.5、2.0和2.5ml于10ml比色管中,依次加入1.0ml(50%)福林酚试剂和2ml(15%)naco3溶
液,定容至10ml,充分混匀后,室温放置1h,于760nm波长下测定吸光度值,建立标准曲线。吸取1.0ml发酵核桃粕按照标准曲线测定方法测定多酚含量,以不加反应液的发酵核桃粕为空白样品。
39.(3)发酵核桃粕的多肽含量测定
40.参考鲁伟等(2005),采用双缩脲法测定多肽得率。取1ml不同浓度gly-gly-tyr-arg四肽与4ml双缩脲试剂混合,静置60min,于540nm波长下测定吸光度值,建立标准曲线。取1ml发酵核桃粕,加入1ml 10%(w/v)的三氯乙酸溶液,摇匀后室温静置10min,6000r/min离心10min得上清液。取1ml上清液与4ml双缩脲试剂混合,静置60min,于540nm波长下测定吸光度值。由标准曲线方程求得发酵核桃粕的多肽浓度并计算多肽含量。
41.(4)发酵前后游离氨基酸的组成与变化
42.核桃粕发酵前后游离氨基酸的组成与变化测定依据为《食品中氨基酸的测定》(gb5009.124-2016),使用的仪器为氨基酸分析仪(茚三酮柱后衍生离子交换色谱仪)。
43.(5)发酵前后酚类化合物的组成与变化
44.代谢物提取:
45.取100mg发酵核桃粕冻干粉,加入900μl去离子水,冰浴条件下超声15min后,12000r/min转速下离心10min,取上清液0.45μm膜过滤后进样检测。取0.1g未发酵核桃粕冻干粉,加入1ml去离子水,冰浴条件下超声15min,12000r/min转速下离心10min,取上清液0.45μm膜过滤后进样检测。
46.液相色谱-质谱分析条件:
47.本试验所用的仪器为岛津超高效液相色谱仪(lc-30)连接sciex 5600 质谱仪。具体参数如下:色谱柱shimadzu inersustain c 18(100
×
2.1mm,2μm),柱温35℃,流速0.300ml/min。流动相a为乙腈,流动相b为0.1%乙酸溶液。洗脱梯度为:0~2min,5%a;2~4min,5~20%a;4~12min,20~25%a;12~14min,25~46%a;14~26min,46~100%a;26~28min,100%a;28~30min,5%a。质谱分别采用电喷雾电离(esi)正离子和负离子模式进行检测。电喷雾离子源(esi)条件:离子源温度500℃(正离子)和450℃(负离子),离子喷雾电压(isvf)5500v(正离子)和4400v(负离子),tof ms扫描范围100~1200da,子离子扫描范围50~1000da,tof ms扫描累积时间0.2s,子离子扫描累积时间0.01s。二级质谱去簇电压
±
60v,碰撞能量35
±
15ev。
48.以下通过具体实施例对本发明作进一步地解释说明。
49.实施例1发酵核桃粕
50.(1)菌种活化:挑取植物乳杆菌cicc 23121转接于100ml mrs液体培养基中,37℃培养箱内培养24h。挑取枯草芽孢杆菌cicc 10002转接于100ml lb液体培养基中,28℃培养箱内培养24h。
51.(2)核桃粕发酵基质:取已过60目筛并紫外线杀菌的核桃粕粉与无菌水混合(8.0%,w/v),反复冻融后,与经115℃灭菌15min的葡萄糖溶液于无菌操作下混合,使得葡萄糖终浓度为2.0%(w/v)。
52.(3)发酵核桃粕:
53.①
经活化的菌种植物乳杆菌、枯草芽孢杆菌在种子培养基中培养18h;
54.②
按各自6.5%(v/v)的接种量量取以上2种菌液,于4000rpm/min离心10min,弃上
清,将沉淀的菌体重悬于等体积的0.9%(w/v)的生理盐水中;
55.③
将以上菌悬液混合接种于核桃粕发酵基质中,置于35℃的培养箱内摇菌发酵(180r/min)培养26h。
56.④
取摇匀后的核桃粕发酵液进行冰浴超声破碎,设置超声破碎条件为工作频率20khz、功率375w、时间30min、时间间隔(工作时间:间隔时间)5:5(s/s)。超声完成后,8000r/min离心20min(4℃),所得上清液即为发酵核桃粕,上清液冻干粉即为发酵核桃粕冻干粉。也即,在本发明中核桃粕发酵液超声破碎后的上清液即发酵核桃粕。将发酵核桃粕置于真空冷冻干燥得到发酵核桃粕冻干粉,并保存于-80℃。
57.(4)抑菌性能测定
58.取核桃粕发酵液超声破碎后的上清液进行抑菌圈试验,其对p.victoria的抑菌圈为13.70
±
0.10mm,对s.aureus的抑菌圈为25.25
±
0.50mm。
59.(5)发酵核桃粕中多酚和多肽的测定
60.核桃粕发酵0h的超声破碎上清液中多酚和多肽含量分别为50.53
±
0.22mg/100ml和7.64
±
0.15mg/ml,发酵结束后的超声破碎上清液中多酚和多肽含量分别为74.25
±
0.09mg/100ml和15.27
±
0.15mg/ml。
61.实施例2多酚鉴定
62.(1)酚类测定
63.采用lc-ms/ms对未发酵核桃粕粉和发酵核桃粕冻干粉中酚类化合物进行检测,共鉴定出15种酚类化合物(表1,图1和2),包括7种酚酸类化合物、7种黄酮类化合物和1种酚苷类化合物。其中酚酸类化合物包括没食子酸、水杨酸(sa)、绿原酸、原儿茶酸、咖啡酸、鞣花酸、反式阿魏酸,黄酮类化合物包括原花青素、(-)-表儿茶素、异槲皮苷、金丝桃苷、根皮苷、柚皮素-7-o-葡萄糖苷和柚皮素,酚苷类化合物水杨苷,而这些化合物的结构之间存在一定的联系。
64.其中,益生菌复合发酵核桃粕后,没食子酸、咖啡酸、sa和绿原酸增加最多,是发酵前的125倍、119倍、117倍和112倍,但异槲皮苷含量降低了约50%,金丝桃苷含量降低了90%,柚皮素-7-o-葡萄糖苷消失。
65.表1 lc-ms/ms分析未发酵和发酵核桃粕冻干粉中的酚类化合物
[0066][0067][0068]
(2)不同酚类化合物的抗菌水平
[0069]
文献查阅发酵核桃粕中15种酚类化合物的抗菌活性,发现没食子酸、sa、柚皮素、咖啡酸、阿魏酸、原儿茶酸和绿原酸可能有抗菌效果。将以上7种酚类化合物对e.coli、s.aureus和p.victoriae处理(表2),发现没食子酸在浓度为5.0mg/ml时,对s.aureus有抗菌效果,抑菌圈达到15.0
±
0.50mm,但对e.coli和p.victoriae没有抗菌效果。sa在浓度为5.0mg/ml时,对s.aureus有抗菌效果,抑菌圈达到13.0
±
0.58mm,对p.victoriae也有抗菌效果,抑菌圈达到10.0
±
0.29mm,但对e.coli没有抑制效果。其余5种酚类化合物包括柚皮素、咖啡酸、阿魏酸、原儿茶酸和绿原酸在浓度为5.0mg/ml时,对s.aureus、e.coli和p.victoriae均无抗菌效果。对sa进行最小抑菌浓度(mic)测定,发现sa对p.victoriae的mic为4.40mg/ml。
[0070]
表2发酵核桃粕冻干粉中酚类化合物的抗菌水平
[0071][0072]
注:不同小写字母(a~c)和大写字母(a~b)分别表示5.0mg/ml的不同酚类化合物对s.aureus和p.victoriae的抗菌活性显著性差异(p《0.05)。采用无菌水和纳他霉素分别作为阴性和阳性对照。
[0073]
实施例3多肽鉴定
[0074]
本实施例利用超滤和lc-ms/ms纯化鉴定多肽序列,pyrx虚拟筛选抗菌活性特征肽段,人工合成验证其抗菌活性,并采用分子对接技术分析其与受体蛋白的相互作用力。
[0075]
(1)不同分子量发酵核桃粕冻干粉的抗菌活性
[0076]
取未发酵核桃粕和发酵核桃粕冻干粉紫外杀菌并转移至离心管,无菌水溶解,备用。
[0077]
采用截留分子量分别为3kda、5kda、10kda的超滤膜对发酵核桃粕进行超滤处理,以纯化和富集制备分子量分别为0~3kda、3~5kda、5~10kda和大于10kda的发酵核桃粕。结束后,将各分子量发酵核桃粕冷冻干燥,紫外灭菌后,转移至离心管并无菌水溶解,备用。
[0078]
测定上述不同成分与不同分子量发酵核桃粕冻干粉的抑菌活性。
[0079]
结果如下:对比不同成分与分子量发酵核桃粕冻干粉对p.victoriae的抗菌效果,如表3所示。结果表明,未发酵核桃粕对p.victoriae没有抗菌活性,发酵核桃粕冻干粉对p.victoriae的mic为40μg/ml,抑菌圈达到12.23
±
0.25mm。
[0080]
发酵核桃粕经过10kda、5kda和3kda膜包超滤和冻干后,得到大于10kda、5~10kda、3~5kda和0~3kd发酵核桃粕冻干粉。对比4组不同分子量发酵核桃粕冻干粉对p.victoriae的抗菌活性,发现5~10kda和3~5kda发酵核桃粕冻干粉对p.victoriae没有抗菌活性。大于10kda发酵核桃粕冻干粉对p.victoriae的mic为20μg/ml,抑菌圈达到10.33
±
0.26mm。0~3kda发酵核桃粕冻干粉对p.victoriae的mic为10μg/ml,抑菌圈达到13.18
±
0.28mm,低于发酵核桃粕冻干粉对p.victoriae的mic,这说明发酵核桃粕冻干粉对p.victoriae的抗菌活性主要来源于0~3kda的组分。后续将对0~3kda冻干粉组分进行多肽的鉴定分析。
[0081]
表3不同发酵核桃粕组分冻干粉对p.victoriae的抗菌水平
[0082][0083][0084]
注:同一列不同大写字母(a~b)表示不同发酵核桃粕组分冻干粉在同一浓度下对p.victoriae的抗菌活性显著性差异(p《0.05)。采用无菌水和纳他霉素分别作为阴性和阳性对照。
[0085]
(2)lc-ms/ms多肽组成鉴定
[0086]
将0~3kda的发酵核桃粕冻干粉进行还原烷基化预处理,过预柱(acclaim pepmap rplc c 18,300μm
×
5mm,5μm)后以600nl/min的流速注入分析柱(acclaim pepmap rplc c 18,150μm
×
150mm,1.9μm)。流动相a为0.1%甲酸,流动相b为0.1%甲酸和80%乙腈。洗脱梯度为:0~2min,4~8%b;2~45min,8~28%b;45~55min,28~40%b;55~56min,40~95%b;56~66min,95~95%b。一级质谱参数:一级质谱分辨率为70000,agc目标为3e6,maximum it为40ms,扫描范围为100~1500m/z。二级质谱参数:二级质谱分辨率为17500,agc目标为1e5,maximum it为60ms,topn为20,nce/stepped nce为27。使用软件peaks studio8.5 de novo对多肽序列解析。
[0087]
结果:将0~3kda的发酵核桃粕冻干粉进行lc-ms/ms(图3)并数据库搜索比对,共得到143个多肽序列,其中氨基酸数量≤10的多肽序列122个。
[0088]
(3)虚拟筛选肽段
[0089]
利用chem3d ultra 14.0软件绘制肽段的3d结构图,格式转化以及能量最小化。rho1 gtpase晶体结构(pdb id:3a58)来自rcsb数据库(https://www.rcsb.org)。蛋白结构用autodocktools 1.5.6软件进行加氢预处理,并保存为pdbqt格式。使用pyrx软件将所有肽段设为配体,与酶依次进行对接打分和虚拟筛选,通过对接能大小判断其相互作用力的强弱以筛选肽段。在uniprot蛋白质数据库(https://www.uniprot.org/proteomes)下载核桃蛋白序列,通过geneious软件对比肽段是否属于核桃蛋白。
[0090]
结果:β-1,3-葡聚糖合成酶负责合成真菌细胞壁组成成分β-1,3-葡聚糖,其由至少两个亚基组成。其中调节亚基rho1 gtpase具有两个开关,这两个开关都参与了β-1,3-葡聚糖合成酶的激活。因此,rho1 gtpase对真菌细胞壁生物合成至关重要,且该亚基通常是抗真菌新药开发的靶点。
[0091]
将lc-ms/ms后得到的122个氨基酸数量≤10的多肽序列与rho1 gtpase进行虚拟筛选,并根据对接能高低对小分子多肽进行排序。对接能通常为负数,其值越小,表明肽段与rho1 gtpase之间的结合越稳定,即越可能具有抗菌活性。因为b.subtilis自身的发酵产物含有具有一定抗菌作用的环脂肽,包括伊枯草菌素和丰原素等。因此,需要通过geneious软件对比多肽和核桃蛋白序列以确定多肽是否属于核桃蛋白序列。经过序列对比,发现对接能排名前10的多肽中有4个肽段属于核桃蛋白序列,分别是fggdsthp、algggy、yvvpw和pllrw(表4)。
[0092]
表4对接能排名前10且属于核桃蛋白序列的肽段
[0093][0094]
(4)人工合成肽段
[0095]
对fggdsthp、algggy、yvvpw和pllrw肽段进行人工合成。肽段的合成采用fmoc固相合成法,根据氨基酸序列合成肽段,经切割、析出、纯化后得到粉末状多肽。合成过程委托南京源肽生物科技有限公司,纯度均大于95%。
[0096]
(5)合成肽段的抗菌活性
[0097]
结果:通过抑菌圈的直径大小评价合成肽段的抗菌活性(表5)。研究发现4种肽段对e.coli都没有抗菌活性,这与前述发酵核桃粕及其冻干粉对e.coli无抗菌活性的结果一致。对于p.victoriae,采用16.0mg/ml的4种多肽对其依次处理,发现多肽yvvpw的抗菌活性最好,而且yvvpw对p.victoriae的mic为16.0mg/ml。algggy和fggdsthp抗菌活性次之,pllrw对p.victoriae没有抗菌活性。对于s.aureus,采用16.0mg/ml的4种多肽对其依次处理,发现没有抗菌活性。增加多肽浓度,采用20.0mg/ml多肽对s.aureus依次处理,发现algggy的抗菌活性大于yvvpw和pllrw,fggdsthp没有抗菌活性。由于导致果蔬腐败的微生物主要为真菌,且刺梨的致腐优势菌为p.victoriae,因此后续实验将对多肽yvvpw展开研究。yvvpw的二级质谱图与化学结构如图4所示。
[0098]
表5不同肽段的抗菌活性
[0099]
[0100]
注:不同小写字母表示16.0mg/ml的不同多肽对p.victoriae的抗菌活性显著性差异(p《0.05),不同大写字母表示20.0mg/ml的不同多肽对s.aureus的抗菌活性显著性差异(p《0.05)。采用无菌水和纳他霉素分别作为阴性和阳性对照。
[0101]
(6)分子对接肽段yvvpw与rho1 gtpase
[0102]
将肽段作为配体,导入schrodinger软件,加氢、结构优化、能量最小化。将rho1 gtpase作为受体,在maestro11.9平台处理,schrodinger软件去除结晶水,加氢,能量最小化。采用maestro软件的glide模块分子对接rho1gtpase和肽段。利用protein preparation wizard模块对rho1 gtpase进行预处理、优化和最小化。导入rho1 gtpase,确定坐标和盒子大小,选取rho1 gtpase的预测活性位点作为盒子的质心,使用sp方法分子对接。
[0103]
结果:采用分子对接模拟构象yvvpw与rho1 gtpase的相互作用,对接结果如图5所示。抗菌肽yvvpw与rho1 gtpase紧密结合且匹配度高,氢键、疏水相互作用和π-π堆积是抗菌肽yvvpw与rho1 gtpase之间的主要相互作用力。将对接后抗菌肽yvvpw与rho1 gtpase形成的复合物利用pymol2.1软件进行可视化,得到抗菌肽yvvpw与rho1 gtpase的结合模式。根据结合模式可以清晰的看到yvvpw与rho1 gtpase活性位点相互作用的氨基酸残基有pro-36、cys-25、val-38、tyr-39、ala-20、lys-123、phe-35、lys-167、leu-126等。抗菌肽yvvpw与rho1 gtpase活性位点氨基酸形成4个氢键,包括抗菌肽的tyr残基与lys-167、pro残基与lys-123、trp残基与pro-36、trp残基与val-38。氢键距离较短,结合力较强,氢键对锚定蛋白rho1 gtpase空腔中的yvvpw有着重要作用。另外,抗菌肽yvvpw的苯环可以与tyr-39、cys-25和phe-35形成π-π共轭相互作用,与lys-167、lys-123、ala-20、tyr-39、pro-36和cys-25形成疏水相互作用力(烷基键和π-烷基键两种),对稳定复合物也有着重要贡献。综上,分子对接模拟构象从分子水平和理论角度阐明了二者的结合模式,抗菌肽yvvpw与rho1 gtpase紧密结合且匹配度高。
[0104]
实施例4yvvpw和sa联合抗菌活性
[0105]
本实施例将抗菌肽与sa联合,测定其混合物对p.victoriae的抗菌活性,并利用荧光光谱、傅里叶红外光谱、分子对接探究两者在结构上的相互作用。
[0106]
(1)采用二倍稀释法测定sa和yvvpw对p.victoriae的最小抑菌浓度(mic)。在吸取100μl浓度约为105cfu/ml的p.victoriae孢子悬液涂布于pda培养基利用无菌牛津杯打孔,第一组加入100μl一定浓度的sa,第二组加入1/2第一组浓度的sa,第三组加入1/2第二组浓度的sa,依次,一共测定5组。以无菌水为阴性对照,以添加纳他霉素为阳性对照。将平板置于28℃培养箱中培养5d,观察结果,以不出现肉眼可见菌落的浓度为sa对p.victoriae的mic。yvvpw的mic依同样方法测定。
[0107]
根据sa和yvvpw单独的mic确定抗菌剂稀释浓度,抗菌剂最高浓度为mic,依次用无菌水倍比稀释成1/2、1/4、1/8、1/16mic进行5
×
5联合。将平板置于28℃培养箱中培养5d,观察结果,以不出现肉眼可见菌落的浓度为sa和yvvpw对p.victoriae的联合mic。联合抗菌实验采用分级抑菌浓度指数(fic)作为判定依据:
[0108][0109]
fic≤0.5,协同作用,fic越小,协同效果越好;0.5《fic≤1,相加作用;1《fic≤2,
无关作用;fic》2,拮抗作用。
[0110]
结果:抗菌肽yvvpw对p.victoriae单独的mic为16.0mg/ml,sa对p.victoriae单独的mic为4.40mg/ml。组合物中抗菌肽yvvpw的浓度范围为1.0mg/ml~4.0mg/ml,sa的浓度范围为0.27mg/ml~1.10mg/ml时二者具有协同抑菌活性(表6)。当组合物中抗菌肽yvvpw的浓度为2.0mg/ml,sa的浓度为0.55mg/ml时,fic值为0.25,对p.victoriae的抗菌活性最高,抑菌圈达到11.50
±
0.41mm。当组合物中抗菌肽yvvpw的浓度为1.0mg/ml,sa的浓度为0.27mg/ml时,对p.victoriae的抑菌圈达到7.35
±
0.20mm,fic值最低,协同效果最好。
[0111]
表6 sa与yvvpw对p.victoriae的协同抑菌活性
[0112][0113]
注:不同小写字母(a~h)表示不同协同工作浓度的sa与yvvpw对p.victoriae的抗菌活性显著性差异(p《0.05)。不同大写字母(a~g)表示不同协同工作浓度sa与yvvpw的分级抑菌浓度指数显著性差异(p《0.05)。
[0114]
(2)荧光光谱探究yvvpw与sa相互作用
[0115]
选择一定浓度的肽段yvvpw与sa混合,通过激发加发射稳态谱测定激发波长。再将1.0mg/ml的yvvpw与不同浓度(0、0.27、0.55、1.10、2.20、4.40mg/ml)的sa混合,记录270nm至500nm的发射光谱。
[0116]
结果:荧光光谱可以用来探索活性物质之间的猝灭行为和结合常数,已被广泛应用于研究小分子与蛋白质的相互作用。抗菌肽yvvpw因含有trp和tyr残基而具有内在荧光,因此可以通过内在荧光猝灭研究yvvpw与sa的相互作用。将一定浓度的yvvpw与sa混合,通过激发加发射稳态谱确定激发波长为261nm。测定不同浓度sa(0、0.27、0.55、1.10、2.20、4.40mg/ml)存在时yvvpw荧光光谱的变化,如图6所示。未添加sa时,yvvpw在368nm处出现最大吸收波长,随着sa添加浓度的增大,yvvpw的荧光强度逐渐降低,表明荧光猝灭发生,且yvvpw的最大吸收波长发生红移。当sa添加浓度为4.40mg/ml时,最大波长红移至397nm。这些结果证实了与sa的相互作用可能改变了抗菌肽yvvpw的分子构象,yvvpw的trp和tyr残基参与了相互作用。
[0117]
(3)傅立叶红外光谱探究yvvpw与sa相互作用
[0118]
为了对yvvpw、sa及yvvpw-sa混合物进行官能团鉴定,采用ftir-atr测量模式进行分析。光谱以400~4000cm-1
的透射率模式记录,扫描次数为32次,分辨率为4cm-1
。
[0119]
结果:为进一步探究抗菌肽yvvpw与sa的相互作用,yvvpw、sa和yvvpw-sa的ftir结果如图7所示。yvvpw在3294.57cm-1
处的吸收峰归属为-oh伸缩振动峰,在2968.92cm-1
处的吸收峰对应饱和碳上的c-h伸缩振动,在1658.18cm-1
处的吸收峰对应酰胺i带,主要由c=o伸缩振动引起,在1518.26cm-1
处的吸收峰对应酰胺酰胺ⅱ带结构,主要由n-h弯曲振动引起。sa在3231.97cm-1
处的吸收峰主要与-oh的伸缩振动有关,2851.82cm-1
处的吸收峰主要与饱和碳上的c-h的伸缩振动有关,1657.04cm-1
处的吸收峰主要由c=o的伸缩振动引起,1609.05cm-1
和1557.0cm-1
处的吸收峰主要由苯环的骨架振动引起,755.54cm-1
和690.77cm-1
处的吸收峰主要与苯环的c-h面外弯曲振动有关。与yvvpw相比,yvvpw-sa在3403.26cm-1
处出现一个窄峰,表明二者之间存在氢键。复合物酰胺i带和酰胺ⅱ带的吸收峰发生了轻微的红移,且随着sa的加入相对强度增大。与sa相比,yvvpw-sa在1609.05cm-1
,755.54cm-1
和690.77cm-1
处的吸收峰分别红移至1613.02cm-1
,756.44cm-1
和698.59cm-1
,且相对强度随着yvvpw的加入减小,说明yvvpw-sa之间存在分子间相互作用。因此,ftir的结果表明,yvvpw与sa之间存在氢键和分子间作用力。
[0120]
(4)分子对接探究yvvpw与sa相互作用
[0121]
利用分子对接进一步验证yvvpw与sa的互作机理,对接结果如图8所示,氢键、疏水相互作用、π-π堆积是抗菌肽yvvpw与sa之间的主要相互作用力。抗菌肽yvvpw与sa紧密结合且匹配度高,结合能为-5.27kcal/mol。利用pymol2.1软件将对接后抗菌肽yvvpw与sa形成的复合物进行可视化,发现sa与yvvpw形成3个强氢键,包括与tyr残基形成键长为的氢键,与抗菌肽n端的第2个和第3个val残基形成键长都为的氢键。氢键距离短,远小于传统氢键的表明sa与yvvpw之间的氢键结合力强,氢键对抗菌肽yvvpw锚定sa有着重要作用。另外,sa的苯环可以与yvvpw的trp残基的五元环形成很强的π-π共轭相互作用,对稳定复合物也有着重要贡献。综上,sa与yvvpw靶点结合紧密,能够形成稳定的复合物,这与本研究的荧光光谱和ftir结果一致。
[0122]
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本专利构思的前提下,上述各实施方式还可以做出若干变形、组合和改进,这些都属于本专利的保护范围。因此,本专利的保护范围应以权利要求为准。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。