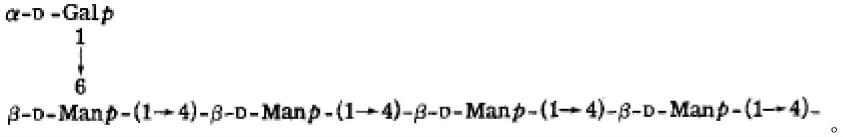
分支半乳糖基的甘露寡糖也产生非还原末端带有α-(1
→
6)-分支半乳糖基的甘露寡糖,以及不带有或末端不带有α-(1
→
6)-分支半乳糖基的甘露寡糖。这些多种不同类型的带有或不带有α-(1
→
6)-分支半乳糖基的甘露寡糖结构的表征,可用于β-半乳甘露聚糖精细结构鉴定的应用;同时主要形成终产物甘露二糖,可用于水解β-半乳甘露聚糖制备甘露二糖中的应用。
7.技术方案:为了实现上述目的,本发明所述灰盖拟鬼伞甘露聚糖酶ccman5c在β-半乳甘露聚糖精细结构鉴定和在水解β-半乳甘露聚糖制备甘露二糖中的应用。
8.其中,所述甘露聚糖酶为灰盖拟鬼伞甘露聚糖酶ccman5c,能够以内切方式水解β-半乳甘露聚糖主链上由带有α-(1
→
6)-分支半乳糖基的甘露糖基c1位(还原端)半缩醛羟基或c4位(非还原端)羟基与其相邻的甘露糖基相应羟基形成的β-(1
→
4)-甘露糖苷键以及内部的不带有α-(1
→
6)-分支半乳糖基的β-(1
→
4)-甘露糖苷键,产生还原末端或非还原末端带有α-(1
→
6)-分支半乳糖基的甘露寡糖,以及不带有或末端不带有α-(1
→
6)-分支半乳糖基的甘露寡糖,寡糖的聚合度(dp)为2-5,基本上不产生甘露糖。
9.作为优选,所述灰盖拟鬼伞甘露聚糖酶ccman5c在角豆胶β-半乳甘露聚糖的水解和水解产物的分析和鉴定中的应用。
10.其中,将灰盖拟鬼伞甘露聚糖酶作为糖链分析工具酶应用于角豆胶β-半乳甘露聚糖(locust bean gum glactomannan polysaccharides,lbg)的水解,结合高效阴离子色谱偶联质谱方法分析和鉴定水解产物寡糖结构,得出角豆胶半乳甘露聚糖中2个带有分支的半乳糖基的甘露糖基之间间隔的不带分支半乳糖基的甘露糖基数目(m)是变化的,m可以从1至5,角豆胶β-半乳甘露聚糖具有如下精细结构:
[0011][0012]
其中,所述灰盖拟鬼伞甘露聚糖酶水解角豆胶β-半乳甘露聚糖主要产物为甘露二糖(约占水解产物寡糖量的50%),经分子排阻色谱法分离纯化获得甘露二糖。
[0013]
本发明所制得甘露二糖对dpph自由基的清除活性是同样浓度甘露聚糖5-7倍,是其他纤维二糖、昆布二糖、乳糖、龙胆二糖、蔗糖的dpph自由基清除率3.5-8.0倍,可用作优质功能性食品或膳食益生元。
[0014]
作为优选,所述甘露聚糖酶为灰盖拟鬼伞甘露聚糖酶ccman5c,其仅水解β-(1
→
4)-甘露聚糖,而不水解α-(1
→
2)-甘露聚糖或其他类型的葡聚糖或木聚糖,并且能够以内切方式水解由带有α-(1
→
6)-分支半乳糖基的甘露糖基还原端或非还原端形成的β-(1
→
4)-甘露糖苷键以及内部的β-(1
→
4)-甘露糖苷键。
[0015]
其中,所述灰盖拟鬼伞甘露聚糖酶ccman5c为成熟的甘露聚糖酶蛋白形式(mccman5c),由基因工程构建的灰盖拟鬼伞甘露聚糖酶异源重组表达毕赤酵母菌发酵获得。其中,成熟甘露聚糖酶mccman5c编码基因的cdna片段以灰盖拟鬼伞菌体中提取的mrna为模板通过rt-pcr合成,或者根据编码基因的cdna序列由人工直接合成,然后将mccman5c的cdna片段整合到表达质粒上,导入毕赤酵母菌中获得重组表达成熟甘露聚糖酶mccman5c的异源重组表达毕赤酵母菌菌株。
[0016]
作为优选,所述灰盖拟鬼伞甘露聚糖酶异源重组表达毕赤酵母由毕赤酵母菌为宿主细胞,导入灰盖拟鬼伞成熟甘露聚糖酶mccman5c编码基因获得,所述灰盖拟鬼伞成熟甘露聚糖酶mccman5c编码基因是指甘露聚糖酶ccman5c cdna的64-1329碱基对的核苷酸序列,即不含编码1-21位氨基酸的1-63碱基对的核苷酸序列及终止密码子的成熟甘露聚糖酶cdna(mccman5c cdna)。
[0017]
所述甘露聚糖酶ccman5c编码基因cdna在ncbi数据库中为cc1g_09314基因、eau91632.1基因、xp_001830154.1基因、xm_001830102.1基因或者nw_003307542.1基因,或其同源蛋白基因kag2015513.1、kag2015514.1、kag2015515.1中的任意一种;所述毕赤酵母菌为毕赤酵母菌gs115菌株或者其他毕赤酵母菌菌株。
[0018]
进一步地,所述灰盖拟鬼伞甘露聚糖酶异源重组表达菌中使用的灰盖拟鬼伞甘露聚糖酶异源重组表达质粒,由灰盖拟鬼伞ccman5c成熟蛋白mccman5c的编码基因cdna片段(mccman5c cdna)和质粒ppiczαa经酶切处理后混合连接,并转化至感受态细胞,提取质粒,获得重组的表达质粒ppiczαa-mccman5c导入毕赤酵母菌。
[0019]
其中,所述灰盖拟鬼伞甘露聚糖酶异源重组表达毕赤酵母菌的构建方法,包括如下步骤:
[0020]
(1)将固体或液体培养基上生长的灰盖拟鬼伞菌丝体或子实体,冷冻后充分研磨成粉末提取制备总rna样品,将mrna逆转录合成cdna;
[0021]
(2)设计引物,以步骤(1)合成的cdna为模板pcr扩增得到成熟ccman5c cdna片段(mccman5c cdna),将mccman5c cdna片段产物和质粒ppiczαa分别经酶切处理后混合连接,并转化至感受态细胞,提取质粒,获得重组的表达质粒ppiczαa-mccman5c;
[0022]
(3)将上述所获得的重组表达质粒ppiczαa-mccman5c线性化处理后电转至毕赤酵母菌细胞中,得到含有质粒ppiczαa-mccman5c的毕赤酵母菌即为重组表达成熟甘露聚糖酶mccman5c的异源重组表达毕赤酵母菌。
[0023]
其中,步骤(2)所述引物为用带ecor i酶切位点上游引物(5
’‑
agagaggctgaagctgaattcgtaggcccttggggccagt-3’)和带not i酶切位点下游引物(5
’‑
tgttctagaaagctggcggccgccccgcgagccttcatgg-3’)。
[0024]
其中,所述甘露聚糖酶的制备包括如下步骤:
[0025]
(1)将灰盖拟鬼伞甘露聚糖酶异源重组表达菌即重组表达成熟甘露聚糖酶mccman5c的异源重组表达毕赤酵母菌,接种bmgy培养基中,过夜培养,离心收集菌体,将菌体悬浮后接种至bmmy培养基中,扩大培养,加入甲醇进行诱导表达,同时取样测定培养基上清液的酶活,当酶活不再增加即终止培养;
[0026]
(2)将停止培养的重组菌发酵液离心收集上清,将上清液与等体积结合缓冲液混合后进行柱层析,收集合并有活性的重组蛋白组分,对透析缓冲液透析过夜,得到纯化的甘露聚糖酶制剂。
[0027]
作为优选,步骤(1)将灰盖拟鬼伞甘露聚糖酶异源重组表达菌接种bmgy培养基中,于28℃,180rpm,过夜培养至od
600
=2~6,离心收集菌体,用1-2ml灭菌水将菌体悬浮后按0.5-1%体积接种至bmmy培养基中,于28℃,180rpm扩大培养,每24h加入甲醇至终浓度为0.5%进行诱导表达,同时取样测定培养基上清液的酶活,当酶活不再增加即终止培养。
[0028]
本发明首次提出了甘露聚糖酶(重组表达的灰盖拟鬼伞成熟甘露聚糖酶
mccman5c)在β-半乳甘露聚糖的水解和水解产物的分析和鉴定中的应用,结合高效阴离子色谱偶联质谱方法(hpaec-pad-ms)分析和鉴定水解产物寡糖结构,首次得出角豆胶β-半乳甘露聚糖精细结构;同时本发明所述重组表达的灰盖拟鬼伞成熟甘露聚糖酶(mccman5c)水解角豆胶半乳甘露聚糖可以制备主要产物甘露二糖。
[0029]
本发明还提出一种重组表达菌株,所述重组表达菌株包括灰盖拟鬼伞成熟甘露聚糖酶mccman5c的基因mccman5c cdna。本发明将所获得的重组的分泌型表达质粒ppiczαa-mccman5c用限制性内切酶pme i单酶切使其线性化,然后电转至毕赤酵母pichia pastoris gs115细胞中,得到一种表达分泌型内切β-1,4-甘露聚糖酶(ccman5c)的毕赤酵母菌,即为重组表达菌株。所含成熟甘露聚糖酶mccman5c的cdna的5’端与ppiczαa上的paox1启动子、α信号肽序列融合,3’端与c-myc-标签、6xhis-标签和终止子编码序列融合,表达产生分泌型、带有6xhis-纯化标签的成熟β-1,4-甘露聚糖酶mccman5c。
[0030]
本发明提供的一种灰盖拟鬼伞成熟的β-1,4-甘露聚糖酶(mccman5c)重组表达质粒和重组表达毕赤酵母菌株、重组表达的一种灰盖拟鬼伞内切β-1,4-甘露聚糖酶,所述毕赤酵母菌含有并能够表达mccman5c基因,产生超糖基化的、分子量大于天然酶蛋白的重组表达的高活性的成熟β-1,4-甘露聚糖酶(mccman5c)。
[0031]
本发明所述的灰盖拟鬼伞甘露聚糖酶ccman5c蛋白质序列和mrna序列在美国国立生物信息研究中心(ncbi)互联网免费公开数据库中收索获得,灰盖鬼伞甘露聚糖酶ccman5c蛋白质序列和mrna序列在美国国立生物信息研究中心互联网免费公开数据库中的登录号为cc1g_09314或eau91632.1或xp_001830154.1或xm_001830102.1或nw_003307542.1,或它的同源蛋白登录号(kag2015513.1或kag2015514.1或kag2015515.1)。ccman5c cdna全长共1332碱基对,编码443个氨基酸,signalp 5.0server软件分析确认其蛋白质n端1-21个氨基酸(mklsaglvslaiavtsasaqa)为信号肽序列,不含信号肽的成熟蛋白质mccman5c共有422个氨基酸,理论分子量为45.8kda。
[0032]
本发明首先以coprinopsis cinerea endo-1,4-beta-mannosidase作关键词在美国国立生物信息研究中心互联网免费公开数据库(ncbi)收索,找到本发明所提出的甘露聚糖酶基因ccman5c,然后根据该基因的cdna序列,使用设计和合成的带ecor i酶切位点的上游引物(5
’‑
agagaggctgaagctgaattcgtaggcccttggggccagt-3’)和带not i酶切位点的下游引物(5
’‑
tgttctagaaagctggcggccgccccgcgagccttcatgg-3’),以从灰盖拟鬼伞菌制备的mrna逆转录合成的cdna为模板,通过pcr扩增出mccman5c cdna的64-1329碱基对的核苷酸序列,即不含编码1-21位氨基酸的1-63碱基对的核苷酸序列及终止密码子的成熟甘露聚糖酶cdna(mccman5c cdna)。所含成熟甘露聚糖酶mccman5c的cdna的5’端与ppiczαa上的paox1启动子、α信号肽序列融合,3’端与c-myc-标签、6xhis-标签和终止子编码序列融合,以便表达产生分泌型、带有6xhis-纯化标签的mccman5c。
[0033]
本发明来源于灰盖拟鬼伞的甘露聚糖酶编码基因序列虽然已经存在于公用、免费基因资源库中,并且被自动基因组注释预测为编码某种甘露聚糖酶的序列,但是该基因所编码蛋白质的功能并没有通过任何实验进行证实。而本发明中根据毕赤酵母菌作为表达宿主的特性,将所述的成熟的甘露聚糖酶mccman5基因导入毕赤酵母菌后进行发酵培养和提纯,得到重组表达的高活性的成熟内切β-1,4-甘露聚糖酶(mccman5c),其甘露聚糖酶纯化形式及催化特性在本发明中均为第一次证实。由本发明提供的甘露聚糖酶可用于生物技术
应用领域。
[0034]
本发明首次发现灰盖拟鬼伞成熟的内切β-1,4-甘露聚糖酶(mccman5c)能够以内切方式水解半乳甘露聚糖主链上由带有α-(1
→
6)-分支半乳糖基的甘露糖基还原端或非还原端形成的β-(1
→
4)-甘露糖苷键,既产生还原末端带分支半乳糖基的甘露寡糖又产生非还原末端带分支半乳糖基的甘露寡糖,以及无分支的、或无末端分支半乳糖基的甘露寡糖,本发明利用重组表达成熟甘露聚糖酶mccman5c应用于β-半乳甘露聚糖的水解,水解的特定产物的产生可以有效用于β-半乳甘露聚糖精细结构鉴定,并提出β-半乳甘露聚糖精细结构。如二糖产物中包括gal1-6man和man1-4man两种同分异构体(图3b,c);三糖产物中包括gal1-6man1-4man、man1-4(gal1-6)man和man1-4man1-4man三种同分异构体(图3d,1,2,3);四糖产物中包括gal1-6man1-4man1-4man、man1-4man1-4(gal1-6)man和man1-4man1-4man1-4man三种同分异构体(图3e,1,2,3);五糖产物中包括gal1-6man1-4man1-4man1-4man、man1-4man1-4man1-4(gal1-6)man、man1-4(gal1-6)man1-4man1-4man和man1-4man1-4man1-4man1-4man四种同分异构体(图3f,1,2,3,4),但水解产物中基本上不含单糖甘露糖。由于灰盖拟鬼伞成熟的内切β-1-4-甘露聚糖酶(mccman5c)水解角豆胶半乳甘露聚糖产物产生一系列不同聚合度(dp2-dp5)的还原末端或非还原末端带1个分支半乳糖基的甘露寡糖,以及不带有、或末端不带有分支半乳糖基的甘露寡糖,这些多种不同类型的带有或不带有α-(1
→
6)-分支半乳糖基的甘露寡糖结构的表征,首次揭示了角豆胶半乳甘露聚糖中2个带有分支的半乳糖基的甘露糖基之间间隔的不带分支半乳糖基的甘露糖基数目(m)是变化的,m可以从1至5,而不是现有技术中报道的间隔3个不带分支半乳糖基的甘露糖基(dea and morrison,1975,adv carbohydr chem biochem,31:242
–
312.)。所以本发明首次揭示角豆胶β-半乳甘露聚糖具有如下精细结构:
[0035]
另外,首次发现,由于灰盖拟鬼伞成熟的内切β-1-4-甘露聚糖酶(mccman5c)水解角豆胶半乳甘露聚糖形成的终产物中主要产物是甘露二糖(约占水解寡糖产物量的50%),可用于水解β-半乳甘露聚糖制备甘露二糖中的应用。
[0036]
以往报道或公开的β-甘露聚糖酶水解半乳甘露聚糖只产生一种类型的末端半乳糖基取代的甘露寡糖,即一种还原末端带分支半乳糖基的甘露寡糖,或一种非还原末端带分支半乳糖基的甘露寡糖,以及产生无分支半乳糖基取代的甘露寡糖和带有非末端分支半乳糖基的不同聚合度的甘露寡糖。如一种黑曲霉β-甘露聚糖酶和一种产紫青霉甘露聚糖酶水解半乳甘露聚糖只产生一种还原末端带分支半乳糖基的甘露寡糖,以及非末端带有分支半乳糖基的不同聚合度甘露寡糖、甘露二糖和甘露三糖(mccleary et al,1983,carbohydr res,119:191-219;park et al,j microbiol biotechnol,1992,2:204-208)。而一种芽孢杆菌内切β-1-4-甘露聚糖酶manb-1601水解半乳甘露聚糖只产生一种非还原末端带分支半乳糖基的甘露二糖、α-(1
→
6)-半乳糖基甘露糖,以及甘露二糖和一个未鉴定的五糖(srivastava et al,2017,j agric food chem 65:2827-2838),并且这些已报道的酶都没能水解半乳甘露聚糖产生一种占优势的水解产物寡糖。
[0037]
有益效果:与现有技术相比,本发明具有以下优点:
[0038]
(1)本发明构建的异源重组表达甘露聚糖酶ccman5的毕赤酵母工程菌可以方便地进行发酵培养和提纯,得到重组表达的成熟的β-1-4-甘露聚糖酶(mccman5c),用于生物技术应用领域。
[0039]
(2)本发明重组表达的成熟的β-1,4-甘露聚糖酶(mccman5c)具有特殊的催化特性和储藏稳定性,有利于生物技术领域应用。
[0040]
(3)本发明首次发现重组表达的成熟的β-1,4-甘露聚糖酶(mccman5c)能够以内切方式水解半乳甘露聚糖主链上由带有α-(1
→
6)-分支半乳糖基的甘露糖基还原端或非还原端形成的β-1-4-甘露糖苷键,产生还原末端或非还原末端带分支半乳糖基的甘露寡糖,以及不带有或末端不带有分支半乳糖基的甘露寡糖,但基本上不产生单糖甘露糖,这样首次用于β-半乳甘露聚糖精细结构鉴定的应用,首次揭示了β-半乳甘露聚糖精细结构:
[0041][0042]
该精细结构的鉴定为mccman5c降解角豆胶β-半乳甘露聚糖主要产生甘露二糖(作为产物的优势糖)提供了理论支撑作用和后续制备生产基础。
[0043]
(4)本发明首次发现重组表达的成熟的β-1,4-甘露聚糖酶(mccman5c)水解角豆胶半乳甘露聚糖形成的终产物中主要产物是甘露二糖,并首次用于水解β-半乳甘露聚糖制备出高产率和高纯度的甘露二糖。
[0044]
(5)本发明首次发现制备的甘露二糖对dpph自由基的清除活性是同样浓度甘露聚糖5-7倍,是其他二糖如纤维二糖、昆布二糖、乳糖、龙胆二糖、蔗糖的dpph自由基清除率3.5-8.0倍,可用作优质功能性食品或膳食益生元。
附图说明
[0045]
图1中a构建的表达载体ppiczαa-mccman5c;b制备的毕赤酵母重组表达的灰盖拟鬼伞成熟甘露聚糖酶mccman5c的蛋白的sds-page分析,其中m,分子量标准蛋白;1,含有空载质粒ppiczαa的毕赤酵母菌gs115菌株的发酵上清液;2,含有质粒ppiczαa-mccman5c的毕赤酵母菌gs115菌株的发酵上清液;3,纯化的灰盖拟鬼伞成熟甘露聚糖酶mccman5c蛋白;c纯化所得66kd蛋白的胰蛋白酶解肽片段的maldi-tof/tof ms分析的两个氨基酸序列(c1和c2)及在ncbi数据库中显示的ccman5c氨基酸序列中的位置(下划线)(c3);d纯化所得64kd蛋白的胰蛋白酶解肽片段的maldi-tof/tof ms分析的两个氨基酸序列(d1和d2)及在ncbi数据库中显示的ccman5c氨基酸序列中的位置(d3),方框线表示ccman5c信号肽序列。
[0046]
图2为制备的毕赤酵母重组表达的成熟甘露聚糖酶mccman5c对不同多糖底物的水解活性(a);对不同浓度角豆胶半乳甘露聚糖的水解速率(b);在50℃、不同ph条件下的水解活性(c)和ph稳定性(d);在ph8.0、不同温度条件下的水解活性(e)和温度稳定性(f);在不同有机溶剂(g)和表面活性剂(h)存在时酶的水解活性;在液态(i)或冷冻干燥状态(j)下在不同储存温度保藏一定时间后残留的酶活性。
[0047]
图3中a为角豆胶半乳甘露聚糖(lbg)经mccman5c酶解24h后的产物的hpaec-pad色谱图;b为图a中p1二糖ms/ms质谱图及结构;c为图a中p2二糖ms/ms质谱图及结构;d为图a中p3三糖ms/ms质谱图及结构;e为图a中p4四糖ms/ms质谱图及结构;f为图a中p5五糖ms/ms质谱图及结构。
[0048]
图4中a为角豆胶半乳甘露聚糖(lbg)的mccman5c酶解液的分子尺寸排阻色谱图;b为a中p1~p4组分的一级质谱图;c为纯化获得的甘露寡糖酸解后单糖组分的hpaec-pad图;d为纯化获得的p2二糖ms/ms质谱图;e中e1为5mg/ml及10mg/ml的甘露糖(m),分子尺寸排阻色谱分离得到的聚合度为dp2-dp5的甘露寡糖组分(dp2-dp5),酶解lbg(dlbg)和未酶解lbg的dpph自由基清除率比较;e2为5mg/ml的不同二糖蔗糖、乳糖、龙胆二糖、纤维二糖、昆布二糖及与本发明纯化所得甘露二糖的dpph自由基清除率比较。
具体实施方式
[0049]
以下结合附图和实施例对本发明作进一步说明。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
[0050]
实施例中所使用的试剂和培养基:
[0051]
结合缓冲液的ph值为7.0,结合缓冲液包括以下摩尔浓度的组分:500mm的nacl、5mm的咪唑和20mm的k2hpo4。
[0052]
洗脱缓冲溶液ph为7.0,所述洗脱缓冲液包括以下摩尔浓度的组分:500mm的nacl、500mm的咪唑和20mm的k2hpo4。
[0053]
透析缓冲液的ph值为7.0,所述透析缓冲液包括以下摩尔浓度的组分:50mm的k2hpo4。
[0054]
bmgy培养基:酵母提取物10g,胰蛋白胨20g,双蒸水700ml,混合后115℃灭菌20分钟,待冷却至室温后,分别加入过滤除菌的100ml的13.4%酵母含氮碱和2ml的200μg/ml生物素,以及115℃灭菌20分钟后的100ml的1m k2hpo4/kh2po4(ph6.0)和100ml的10%甘油。
[0055]
bmmy培养基:酵母提取物10g,胰蛋白胨20g,双蒸水700ml,混合后115℃灭菌20分钟,待冷却至室温后,分别加入过滤除菌的100ml的13.4%酵母含氮碱、2ml的200μg/ml生物素和100ml的5%甲醇,以及115℃灭菌20分钟后的100ml的1m k2hpo4/kh2po4(ph6.0)。
[0056]
实施例1
[0057]
灰盖拟鬼伞内切β-1,4-甘露聚糖酶(ccman5c)重组表达质粒和重组表达毕赤酵母菌株构建、重组表达的甘露聚糖酶的纯化与鉴定
[0058]
1、重组表达质粒的制备方法,包括以下步骤:
[0059]
将固体或液体培养基上生长的灰盖拟鬼伞(灰盖拟鬼伞菌株为atcc 56838,从美国典型菌种保藏中心atcc购买)菌丝体或子实体,经液氮冷冻后在液氮中充分研磨成粉末,使用生工(sangon biotech)公司生产的total rna extractor(trizol)试剂盒,按照说明书操作制备总rna样品。使用vazyme公司生产的ⅱq rt supermix for qpcr( gdnawiper)试剂盒,按照说明书要求操作将总rna样品中基因组dna降解去除后,将mrna逆转录合成cdna。
[0060]
设计带ecor i酶切位点上游引物seq id no.3:(5
’‑
agagaggctgaagctgaattcgta
ggcccttggggccagt-3’)和带not i酶切位点下游引物seq id no.:4:(5
’‑
tgttctagaaagctggcggccgccccgcgagccttcatgg-3’),以上述步骤合成的cdna为模板,使用primestar mix(takara,china),按以下条件进行pcr扩增:
①
98℃,5min;
②
98℃,20sec;
③
55℃,20sec;
④
72℃,1.5min;
⑤
重复
②‑④
30个循环;
⑥
72℃,10min,得到灰盖拟鬼伞ccman5c成熟蛋白的cdna片段(mccman5c cdna)。
[0061]
将mccman5c cdna片段产物和质粒ppiczαa分别经ecor i和not i两种限制性核酸内切酶处理后,使用entry one step cloning kit试剂盒(vazyme,chian)按说明书混合连接,并转化至e.coli dh5α,用sanprep柱式质粒dna小量抽提试剂盒(生工生物公司,中国)提取质粒,获得重组的表达质粒ppiczαa-mccman5c。图1a为实施例1构建的表达载体ppiczαa-mccman5c图,图中mccman5c基因插入到ecor i和not i两个酶切位点之间,替换了其他多克隆位点。
[0062]
阳性的克隆质粒送生工生物技术公司(上海)测序鉴定,mccman5c cdna的核酸序列如seq id no.1所示,与ncbi数据库中登录序列相一致,用于后续实验,其表达的蛋白的氨基酸序列如seq id no.2所示。
[0063]
2、重组表达菌株的制备方法,包括以下步骤:
[0064]
将上述所获得的重组的分泌型表达质粒ppiczαa-mccman5c用限制性内切酶pme i单酶切使其线性化,然后根据厂家操作手册电转至毕赤酵母pichia pastoris gs115细胞(invitrogen,美国)。得到一种表达内切β-1,4-甘露聚糖酶(ccman5c)的毕赤酵母菌,即为重组表达菌株。所含成熟甘露聚糖酶mccman5c的cdna的5’端与ppiczαa上的paox1启动子、α信号肽序列融合,3’端与c-myc-标签、6xhis-标签和终止子编码序列融合,表达产生分泌型、带有6xhis-纯化标签的成熟的β-1,4-甘露聚糖酶mccman5c(图1a)。
[0065]
3、甘露聚糖酶的制备方法,包括如下步骤:
[0066]
将重组表达菌株接种到10ml bmgy培养基中,于28℃,180rpm,过夜培养至od
600
=4,离心收集菌体,用1ml灭菌水将菌体悬浮后按1%体积接种至bmmy培养基中,于28℃,180rpm扩大培养,每24h加入甲醇至终浓度为0.5%进行诱导表达,同时取样测定培养基上清液的酶活,当酶活不再增加即终止培养;
[0067]
将停止培养的重组菌发酵液离心收集上清,在4℃下,将上清液与等体积的2
×
结合缓冲液混合后上样至proteinisotm ni-nta树脂(transgen biotech公司),也可以采用probond
tm
nickel-chelating resin树脂(invitrogen公司)或其他类似含ni-亲和树脂装填的亲和层析柱,上样结束后用1
×
结合缓冲液冲洗柱子至流出液紫外吸收(od280)为0,然后采用5-100%洗脱缓冲液梯度,在bio-rad低压层析系统上梯度洗脱层析柱上的重组蛋白,收集合并有活性的重组蛋白组分,使用14kda截留透析袋(biosharp公司,中国)对透析缓冲液透析过夜,得到纯化的甘露聚糖酶制剂(即为重组表达的灰盖拟鬼伞成熟甘露聚糖酶mccman5c)。
[0068]
本实施例通过重组毕赤酵母菌表达的灰盖拟鬼伞成熟的甘露聚糖酶mccman5c,在聚丙烯凝胶电泳(sds-page)上显示其分子量比灰盖拟鬼伞蛋白质甘露聚糖酶理论分子量48kd大,是由重组表达的甘露聚糖酶被毕赤酵母菌糖基化修饰所引起,且具有两个糖基化程度不同分子量变体,分子量分别为66kd和64kd。图1b为实施例1制备的毕赤酵母重组表达的灰盖拟鬼伞甘露聚糖酶mccman5c的蛋白的sds-page分析结果图。m,分子量标准蛋白;1,
含有空载质粒ppiczαa的毕赤酵母菌gs115菌株的发酵上清液;2,含有质粒ppiczαa-mccman5c的毕赤酵母菌gs115菌株的发酵上清液;3,纯化的灰盖拟鬼伞成熟的甘露聚糖酶mccman5c蛋白。
[0069]
将上述的重组毕赤酵母菌表达的灰盖拟鬼伞成熟的甘露聚糖酶mccman5c用胰蛋白酶水解,产生的肽片段分离纯化后经maldi-tof/tof ms分析鉴定,66kd和64kd蛋白两个变体蛋白均为毕赤酵母gs115菌株表达的mccman5c蛋白,氨基酸序列上并无差异,这是常见的毕赤酵母重组表达蛋白不同程度的糖基化修饰所引起的分子量大小差异。图1c表示纯化所得66kd蛋白的胰蛋白酶解肽片段的maldi-tof/tof ms分析的两个氨基酸序列(c1和c2)及在ncbi数据库中显示的ccman5c氨基酸序列中的位置(下划线)(c3)。图1d表示纯化所得64kd蛋白的胰蛋白酶解肽片段的maldi-tof/tof ms分析的两个氨基酸序列(d1和d2)及在ncbi数据库中显示的ccman5c氨基酸序列中的位置(d3)。方框线表示为man5c信号肽序列。说明灰盖拟鬼伞成熟甘露聚糖酶mccman5c可以经重组毕赤酵母菌表达和ni-亲和树脂层析柱分离纯化获得。
[0070]
实施例2
[0071]
酶催化特性
[0072]
本发明发现实施例1所制得的重组表达的灰盖拟鬼伞成熟甘露聚糖酶具有特殊的催化特性,耐高温、耐碱、耐表面活性剂、耐有机溶剂、耐长期储存。
[0073]
对实施例1制备的重组表达、纯化获得的灰盖拟鬼伞成熟甘露聚糖酶mccman5c的酶活、底物特异性、最适反应条件、酶促动力学、酶的稳定性及对有机溶剂和表面活性剂的耐受性等特性进行分析,测定方法及结果如下:
[0074]
1、甘露聚糖酶的酶活及底物特异性分析(还原糖法,也即dns法)
[0075]
标准的甘露聚糖酶的酶活测定反应体系、条件和操作步骤为,100μl反应溶液中含50mm pbs缓冲液(ph8.0),2.5mg/mlβ-半乳甘露聚糖或其他多糖,250ng/ml重组表达纯化的β-甘露聚糖酶mccman5c(实施例1制备得到纯化的甘露聚糖酶制剂),反应溶液在50℃温度、800rpm条件下孵育10min后,加入100μl的dns试剂(3,5-二硝基水杨酸),混匀后100℃加热10min;冷却至室温后,离心取上清测od
520
nm。1个酶活力单位定义为在该反应条件下每分钟释放1.0μmol还原糖(以甘露糖为标准)所需要的酶量。
[0076]
由图2a可以看出,mccman5c只对角豆胶β-半乳甘露聚糖(locust bean gum glactomannan polysaccharides from seeds of ceratoria silique,lbg,含β-d-man-1,4-β-d-man和α-d-gal-1,6-β-d-man糖苷键,购自美国sigma公司)有水解活性,比活为312.68u/mg,而不能降解酿酒酵母α-甘露聚糖(含α-d-man-1,2-α-d-man和α-d-man-1,6-α-d-man糖苷键,购自美国sigma公司),以及微晶纤维素avicel(含β-d-glc-1,4-β-d-glc糖苷键,购自日本tci公司)、燕麦木聚糖(含β-d-xyl-1,4-β-d-xyl糖苷键,购自美国sigma公司)和掌狀昆布昆布多糖(含β-d-glc-1,3-β-d-glc和β-d-glc-1,6-β-d-glc糖苷键,购自美国sigma公司)。
[0077]
2、甘露聚糖酶最适反应ph分析
[0078]
将上述的甘露聚糖酶的酶活测定的反应溶液中50mm pbs缓冲溶液(ph8.0)替换成不同ph的缓冲液,50mm naac-hac缓冲液(ph 3.0~6.0),或50mm pbs缓冲液(ph 6.0~8.0),或50mm tris-hcl缓冲液(ph 8.0~9.0),其他成分和含量及反应条件和操作步骤保
持不变,分别通过上述dns法测定酶活,以测得的最高酶活值设定为100%,计算其他各ph反应区间的相对酶活。
[0079]
如图2c所示,本发明实施例1制备的成熟的甘露聚糖酶mccman5c在ph3.0~9.0之间随着ph升高,酶活性增加,ph 8.5-9.0时达到最高酶活,说明该酶偏好碱性的反应条件。
[0080]
3、甘露聚糖酶的ph稳定性分析
[0081]
将前述的甘露聚糖酶配制在50μl不同上述不同ph(ph值为3~9)的50mm缓冲液中,于室温孵育1h后,调ph至8.0,再与50μl的50mm pbs缓冲液(ph8.0)配制的5mg/mlβ-半乳甘露聚糖混合后,通过上述dns法测定酶活,以未孵育储藏酶测得的值设定为100%,计算不同ph孵育后酶的相对酶活。
[0082]
由图2d可知,本发明实施例1制备的成熟的甘露聚糖酶mccman5c在ph为8-9条件下,孵育1h后能够维持大于90%的酶活。说明本发明重组表达的灰盖鬼伞成熟的甘露聚糖酶mccman5c的ph稳定性好。
[0083]
4、甘露聚糖酶最适反应温度分析
[0084]
将上述的甘露聚糖酶酶活测定的反应体系中指定的50℃温度条件分别变换成20℃、30℃、40℃、50℃、60℃、70℃、或80℃温度,其他反应溶液成分和含量及反应条件和操作步骤保持不变,分别通过上述dns法测定酶活,以测得的最高酶活值设定为100%,计算其他各温度反应区间的相对酶活。
[0085]
结果如图2e所示,本发明实施例1制备的成熟的甘露聚糖酶mccman5c在20℃~70℃范围内,酶活随着反应温度的上升而逐渐升高,在70℃时酶活力最大,当反应温度超过70℃时,酶活力急剧下降,说明酶最适温度为70℃。
[0086]
5、甘露聚糖酶的温度稳定性分析
[0087]
先将前述的配制在50mmpbs缓冲液(ph8.0)成熟的甘露聚糖酶mccman5c溶液50μl于20-80℃不同的温度条件下孵育60min后,再与50μl的50mm pbs缓冲液(ph8.0)配制的5mg/mlβ-半乳甘露聚糖混合后,通过上述dns法测定酶活,以未孵育储藏酶测得的值设定为100%,计算不同温度孵育后酶的相对酶活。
[0088]
由图2f可以看出,该酶在20~50℃之间时较为稳定,随着温度升高超过50℃后,酶活快速下降。本发明实施例1制备的甘露聚糖酶在50℃温度下孵育1h后仍保持80%以上的酶活。说明本发明重组表达的灰盖鬼伞成熟的甘露聚糖酶mccman5c的温度稳定性好。
[0089]
6、甘露聚糖酶的酶促动力学测定
[0090]
以50mm pbs缓冲液(ph8.0)分别配制0.25mg/ml、0.5mg/ml、1mg/ml、1.5mg/ml、2mg/ml、2.5mg/ml、3mg/ml、3.5mg/ml、4mg/ml、4.5mg/ml、5mg/ml角豆胶半乳糖甘露聚糖溶液,分别将上述不同浓度的角豆胶半乳糖甘露聚糖溶液50μl替换前述的甘露聚糖酶的酶活测定的反应体系中5mg/mlβ-半乳甘露聚糖溶液,其他成分和含量及反应条件和操作步骤保持不变,分别通过上述dns法测定酶活,利用origin软件绘制km曲线(图2b)并计算km及vm。经计算得出成熟的甘露聚糖酶mccman5c km和vmax值分别为1.23mg.ml-1
和531.77mol.min-1
.mg-1
。说明在1-3mg/ml甘露聚糖浓度范围内,灰盖鬼伞成熟的甘露聚糖酶mccman5c的催化活性随甘露聚糖浓度增加而上升,超过3mg/ml浓度后,甘露聚糖酶催化活性基本上不再增加。
[0091]
7、甘露聚糖酶对表面活性剂的耐受性
[0092]
将表面活性剂sds、或tritonx-100、或tween-20按终浓度1%或10%分别加入至上述的标准的甘露聚糖酶的酶活测定反应溶液中,其他成分和含量及反应条件和操作步骤保持不变,分别通过上述dns法测定酶活,以未加表面活性剂的空白组的酶活为100%,计算其他各加入表面活性剂的实验组的相对酶活。如图2h所示,sds抑制mccman5c酶活,但1%或10%tween20和tritonx-100增强酶的催化活性,与对照相比酶活提高了26.2%~45.6%。说明mccman5c不仅耐受、而且其催化活性还能被表面活性剂所增强。
[0093]
8、甘露聚糖酶对有机溶剂的耐受性
[0094]
将有机溶剂甲醇、或乙醇、或异丙醇、或丙酮按终浓度10%或20%分别加入至上述的标准的甘露聚糖酶的酶活测定反应溶液中,其他成分和含量及反应条件和操作步骤保持不变,分别通过上述dns法测定酶活,以未加有上述溶剂的空白组的酶活为100%,计算其他各加入有机溶剂的实验组的相对酶活。如图2g所示,10%或20%甲醇、乙醇、异丙醇和丙酮等有机溶剂对ccman5c活性均无明显抑制作用,相反,10-20%异丙醇和丙酮、以及20%的甲醇、乙醇却能够增强酶的催化活性,与对照相比酶活提高了9.20%~34.98%。说明mccman5c不仅耐受而且其催化活性还能被较高浓度的有机溶剂中所增强。
[0095]
9、甘露聚糖酶的储存稳定性
[0096]
对实施例1制备的重组表达、纯化获得的灰盖拟鬼伞成熟的甘露聚糖酶mccman5c溶液样品分装后直接分别放置于4℃、-20℃、-80℃冰箱中或冷冻真空干燥后后分别放置于4℃、-20℃、-80℃冰箱中保存,分别于保藏半个月、1个月、3个月、6个月、12个月后取出样品,按上述的标准的甘露聚糖酶的酶活测定反应体系测定不同保存条件下酶样品的酶活。以保存起始时测得的mccman5c的初始酶活为100%,计算其他不同保藏条件、不同保藏时间实验组样品的相对酶活。如图2i所示,冷冻干燥后的甘露聚糖酶mccman5c在-20℃或-80℃冰箱储存的前3个月内活性有所下降,但在保存6个月后mccman5c的酶活相比初始酶活增加了40%和60%,保藏12个月后的酶活仍比初始酶活高28%;如图2j所示,纯化透析后液态的甘露聚糖酶mccman5c在-20℃或-80℃冰箱储存的前3个月内,mccman5c的酶活与对照相比也有所下降,但在保存6个月、甚至保存12个月后mccman5c的酶活则与初始酶活相比几乎没有变化;另外,冷冻干燥的或液态的甘露聚糖酶mccman5c样品在4℃保存的12个月期间,其酶活与对照相比始终维持50%以上。说明甘露聚糖酶mccman5c具有很高的存储稳定性。
[0097]
实施例3
[0098]
甘露聚糖酶应用于β-半乳甘露聚糖的水解和水解产物的分析和鉴定
[0099]
200μl角豆胶β-半乳甘露聚糖酶解反应液的组成成分和含量为:50mm pbs缓冲液(ph8.0)中含终浓度1.5μg/ml灰盖拟鬼伞成熟的甘露聚糖酶mccman5c(实施例1制备)和终浓度2.5mg/ml角豆胶β-半乳甘露聚糖(lbg,购自美国sigma公司)。酶解反应液于50℃条件下反应24h,然后100℃加热15min,冷却至室温,离心取上清液通过0.22μm滤头过滤,备用。
[0100]
上述过滤后的酶解液上样carbopac pa1色谱柱(4
×
250mm,dionex,sunnyvale,ca),利用高效阴离子液相色谱仪(professional ic vario 940)进行高效阴离子液相色谱结合脉冲安培检测器(hpaec-pad)色谱分析,柱温30℃,流速1ml/min,定量环为20μl。按以下程序进行洗脱,0~30min:100%a等梯度;30~60min:100%a—60%a 40%b;60~65min:60%a 40%b—100%b;65~70min:100%b等梯度;70~71min:100%b—100%a 0%b;71~80min:100%a等梯度;流动相a:12mm naoh,流动相b:12mm naoh 300mm naoac。图3a为lbg
酶解产物的hpaec-pad色谱图,显示共有p1、p2、p3、p4、p5五个水解产物寡糖峰。
[0101]
采用高效阴离子液相色谱结合脉冲安培检测器-质谱仪(hpaec-pad-q-tof-ms)对酶解液寡糖产物峰进行分析鉴定。分析时样品溶液经pa-1分析柱分离后,洗脱液经一个三通分流器分流,除了一部分洗脱液直接流入脉冲安培检测器(pad)检测外,另一部分洗脱液经脱盐柱处理后流经质谱仪进行分析。q-tof-ms分析参数为,雾化气压:40psi;干燥气流速/温度:10l/min,350℃;毛细管电压:3500v;扫描质量数范围:100m/z~3000m/z,碎裂电压:100v;碰撞能根据分子量大小在10v~50v之间适当调整;采用负离子模式。q-tof-ms分析结果显示,p1和p2均为二糖,p3为三糖,p4为四糖,p5为五糖,没有检测到明显的单糖甘露糖。图3b是产物峰p1二糖的q tof-ms/ms二级质谱图,在它的ms/ms碎片离子峰中,除了来自糖苷键断裂产生的典型的碎片离子b1,c1,跨环断裂离子
0,2
a2,
0,3
a2,
0,4
a2来自于距非还原末端的第二个糖环上,确认糖苷键是1,6连接。已知lbg是由以β-(1
→
4)-糖苷键连接的甘露聚糖为主链,以α-(1
→
6)-糖苷键连接的d-半乳糖为分支侧链按半乳糖和甘露糖约1:4的比例组成的杂多糖,所以该p1二糖为gal1-6man。图3c是p2二糖q tof-ms/ms质谱图,在ms/ms碎片离子峰中,除了来自糖苷键断裂产生的典型的碎片离子b1和c1,还存在来自于距非还原末端的第二个糖环上跨环断裂离子
0.2
a2,
2.4
a2,
2.5
a2,因而它们是(1
→
4)-糖苷键连接,根据已知的lbg结构可以判断p2二糖为man1-4man。图3d是p3三糖q tof-ms/ms质谱图,图中可见在碎片离子峰中,除了有来自糖苷键断裂产生的典型的碎片离子b1,c1,b2,c2,还有1,6-糖苷键跨糖环断裂诊断性离子峰
0.3
a2和1,4-糖苷键跨糖环断裂诊断性离子峰
2.5
a2,说明非还原末端的第一个糖基与距非还原端第二个糖基的连接方式有(1
→
6)-糖苷键和(1
→
4)-糖苷键两种,提示三糖分子非还原末端为半乳糖基以(1
→
6)-糖苷键与甘露糖基相连,或非还原末端为甘露糖基以(1
→
4)-糖苷键另一甘露糖基相连;另外,存在距非还原末端的第三个糖环产生的跨环断裂离子峰
0.2
a3,
2.4
a3,
2.5
a3,说明距非还原末端的第二个糖基与第三个糖基以-(1
→
4)-糖苷键连接;还发现由距非还原端第三个糖基的c4、c6上同时各连接一个糖环时产生的一个特殊的跨环断裂离子峰
3.5
a3,考虑到角豆胶β-半乳甘露聚糖的β-(1
→
4)-主链中存在α-(1
→
6)-半乳糖分支侧链,且甘露糖与半乳糖比值约为4:1,一分子三糖中不可能同时存在两分子半乳糖,由此判断dp3的产物中存在一种甘露二糖的还原末端的甘露糖基上带有一个α-(1
→
6)-半乳糖基分支侧链的结构。因而dp3三糖存在三个同分异构体,man1-4man1-4man(1)、gal1-6man1-4man(2)和man1-4(gal1-6)man(3)。图3e是p4四糖q tof-ms/ms图谱,图中可见在碎片离子峰中,有来自糖苷键断裂产生的典型的碎片离子b1,c1,b2,c2,b3,c3,诊断性跨糖环断裂离子
0.3
a2和
2.5
a2在距非还原末端的第二个糖基处均存在,认定距非还原末端的第一个糖基和第二糖基或者以(1
→
6)-糖苷键连接,或者以(1
→
4)-糖苷键连接;跨糖环断裂离子
0.2
a,
2.4
a,
2.5
a在距非还原末端的第三和第四糖基处产生,确认它们是(1
→
4)-连接;另外,跨糖环断裂离子
3.5
a4存在,再结合lbg中甘露糖与半乳糖的比值约为4:1,一分子寡糖中不可能同时存在两分子半乳糖,最多只能存在一个,由此可以判断dp4的产物中存在一种其还原末端甘露糖基带有一个α-(1
→
6)-分支半乳糖基的结构。因而产物p4四糖存在三个同分异构体,man1-4man1-4man1-4man(1)、gal1-6man1-4man1-4man(2)、man1-4man1-4(gal1-6)man(3)。图3f是p5五糖q tof-ms/ms质谱图,在碎片离子峰中,有来自糖苷键断裂产生的典型的碎片离子b1,c1,b2,c3,b4,c4,诊断性跨糖环断裂离子
0.3
a2和
2.5
a2在距非还原末端的第二个糖环处均存在,认定既有(1
→
6)-糖苷键连接又有(1
→
4)-糖苷键连接,提示五糖分子非还原末端为半乳糖基以α-(1
→
6)-糖苷键与甘露糖基相连,或为甘露糖基以β-(1
→
4)-糖苷键与另一甘露糖基相连;跨环断裂离子
0.2
a,
2.4
a,
2.5
a在距非还原末端的第三、第四和第五糖环处产生,确认它们是以β-1,4糖苷键连接。另外,距非还原末端第三糖环跨环断裂离子
3,5
a3和第五糖环跨环断裂离子
3.5
a5也分别存在,由于lbg中甘露糖与半乳糖的比值约为4:1,一分子寡糖中不可能同时存在两分子半乳糖,最多只能存在一个半乳糖基,因此提示在甘露五糖分子中距非还原端第三个糖基(甘露糖基)或第五个糖基(还原末端甘露糖基)上分别带有一个α-(1
→
6)-分支的半乳糖基。这样,五糖共有四个同分异构体,分别为man1-4man1-4man1-4man1-4man(1)、gal1-6man1-4man1-4man1-4man(2)、man1-4(gal1-6)man1-4man1-4man(3)、man1-4man1-4man1-4(gal1-6)man(4)。
[0102]
根据上述水解产物寡糖的结构分析,由于水解产物中二糖结构分别为gal1-6man和man1-4man,三糖结构分别为man1-4man1-4man、gal1-6man1-4man和man1-4(gal1-6)man,四糖结构分别为man1-4man1-4man1-4man、gal1-6man1-4man1-4man、man1-4man1-4(gal1-6)man,五糖结构分别为man1-4man1-4man1-4man1-4man、gal1-6man1-4man1-4man1-4man、man1-4(gal1-6)man1-4man1-4man和man1-4man1-4man1-4(gal1-6)man,但没有检测到单糖甘露糖,可以得出角豆胶半乳甘露聚糖中2个带有分支的半乳糖基的甘露糖基之间间隔的不带分支半乳糖基的甘露糖基数目(m)是变化的,m可以从1至5,这样角豆胶β-半乳甘露聚糖具有如下精细结构:
[0103][0104]
而不是原先报道的间隔3个不带分支半乳糖基的甘露糖基(dea and morrison,1975,adv carbohydr chem biochem,31:242
–
312.):
[0105][0106]
本实施例首次发现一种灰盖拟鬼伞内切β-(1
→
4)-甘露聚糖酶(mccman5c)能够以内切方式水解半乳甘露聚糖主链上由带有α-(1
→
6)-分支半乳糖基的甘露糖基还原端或非还原端形成的β-(1
→
4)-甘露糖苷键以及内部的β-(1
→
4)-甘露糖苷键,产生还原末端或非还原末端带有α-(1
→
6)-分支半乳糖基的甘露寡糖,以及不带有分支的或末端不带有α-(1
→
6)-分支半乳糖基的甘露寡糖。这些多种不同类型的带有或不带有α-(1
→
6)-分支半乳糖基的甘露寡糖结构的表征,有利于β-半乳甘露聚糖精细结构鉴定的应用。本实施例将重组甘露聚糖酶作为糖链分析工具酶应用于角豆胶β-半乳甘露聚糖的水解,结合高效阴离子色谱偶联质谱方法分析和鉴定水解产物寡糖结构,推导出角豆胶β-半乳甘露聚糖精细结构。之前没有将甘露聚糖酶用于水解β-半乳甘露聚糖并通过高效阴离子色谱分析偶联质谱方法分析寡糖结构推导出β-半乳甘露聚糖精细结构报道。本发明中甘露聚糖酶水解β-半乳甘露聚糖产生的寡糖通过高效阴离子色谱分析偶联质谱方法分析寡糖结构,与以往采用核磁共振或衍生物法结合gc-ms等方法相比具有样品用量少,能够精确分析糖基及其糖苷键的
排列顺序等优点。
[0107]
实施例4
[0108]
甘露聚糖酶应用于甘露二糖的制备
[0109]
1.2ml角豆胶半乳甘露聚糖的甘露聚糖酶解反应液的组成和含量为:50mm醋酸铵(nh4ac)缓冲液(ph 7.12)含终浓度30μg/ml灰盖拟鬼伞成熟的甘露聚糖酶mccman5c(实施例1制备)和终浓度2.5mg/ml角豆胶β-半乳甘露聚糖(lbg,购自美国sigma公司)。酶解反应液于50℃条件下,800rpm孵育24h,然后100℃加热15min灭活,冷却至室温,离心后上清液用0.22μm滤头过滤,备用。
[0110]
酶解产物甘露寡糖的分离纯化。取1.0ml上述酶解液上清液上样至安装在agilent 1100系统上的分子尺寸排阻色谱柱(superdex
tm
30xk26三联柱,事先已用0.15m nh4ac,ph 7.12,平衡处理),使用流动相0.15m nh4ac(ph 7.12)以0.5-2.0ml/min流速洗脱,折光率检测器(agilent 1100系列g1362 arid)监测,每1-5min 1管分步收集所有洗脱组分,分离纯化酶解产物β-甘露寡糖。如图4a所示,分子尺寸排阻色谱图显示5个峰面积较大且分离良好的峰,p1、p2、p3、p4和p5。如图4b经一级质谱鉴定峰p1-p5所产生的阴离子化分子[m-h]-、[m 2h2o-h]-以及[m hso4]-,确定p1为二糖(dp2),p2为三糖(dp3),p3为四糖(dp4),p4为五糖(dp5),p5不是糖类物质。经分子尺寸排阻色谱分离纯化后,从5mg lbg酶解液中可分离获得0.56mg二糖、0.24mg三糖、0.14mg四糖、0.24mg五糖,其中二糖得率为11.2%,二糖占水解产物寡糖量约50%。如图4c所示,纯化mos经酸解后采用hpaec-pad单糖组分分析,lbg中甘露糖与半乳糖的摩尔比为3.98:1,纯化得到的dp2、dp3、dp4、dp5寡糖中甘露糖与半乳糖的摩尔比分别为40.33:1、7.17:1、7.70:1、15.23:1。dp2二糖中甘露二糖含量达95.15%。如图4d所示,p1二糖经ms/ms质谱分析,显示来自糖苷键断裂产生的碎片离子b1和c1,以及来自于距非还原末端的第二个糖环上跨环断裂碎片离子
0.2
a2,
2.4
a2,
2.5
a2,因而它们是1,4糖苷键连接,为man1-4man。
[0111]
测定甘露二糖或其他己糖二糖对dpph的自由基清除能力。向100μl 5mg/ml或10mg/ml的甘露二糖(上述纯化得到的dp2)或其他己糖二糖水溶液中加入100μl 0.15mm dpph乙醇溶液,于37℃避光孵育30min后,测定517nm处的吸光度(ab)。100μl分子尺寸排阻色谱洗脱液50mm乙酸铵充分冻干重溶后水溶液分别与100μl 0.15mm dpph混合反应后测定值做空白对照。其他己糖二糖分别溶解于50mm乙酸铵中充分冻干后再重溶于超纯水中用作对比样品测试。另制备100μl mccman5c酶解角豆胶半乳甘露聚糖(dlbg)溶液,向100μl 50mm(ph 8.0)tris缓冲液中含5mg/ml lbg和15μg/ml mccman5c,或含10mg/ml lbg和30μg/ml mccman5c,在37℃,800rpm,反应24h后,100℃高温灭活,降至室温,与同样浓度未酶解角豆胶半乳甘露聚糖(lbg)作对照,同上加入100μl 0.15mm dpph乙醇溶液,测试酶降解作用对角豆胶β-半乳甘露聚糖的dpph自由基清除能力的影响。自由基清除活性计算为dpph的抑制百分数:inhibition%=(abscontrol-abssample)/abscontrol
×
100。
[0112]
如图4e1所示,5mg/ml、10mg/ml的lbg酶解液(dlbg)对dpph自由基的清除活性分别是未酶解lbg溶液的2.4倍和2.7倍。说明相较长链甘露聚糖,短链甘露寡糖具有较高抗氧化活性。同时发现本发明首次制备的甘露二糖(dp2)有着比dp3甘露寡糖、dp4甘露寡糖、dp5甘露寡糖更高更显著的抗氧化活性,其中5mg/ml甘露二糖表现出54%dpph自由基清除活性,10mg/ml甘露二糖则表现出73%dpph自由基清除活性,对dpph自由基的清除活性是同样浓
度半乳甘露聚糖5-7倍。进一步比较各种二糖与甘露二糖的dpph自由基清除活性如图4e2所示,5mg/ml纤维二糖、昆布二糖、乳糖、龙胆二糖、蔗糖的dpph自由基清除率仅依次为15.44%、13.76%、10.67%、8.03%、6.59%,而甘露二糖的dpph自由基清除率是纤维二糖、昆布二糖、乳糖、龙胆二糖、蔗糖的dpph自由基清除率3.5-8.0倍。首次揭示甘露二糖对dpph自由基的清除活性极大地高于是其他己糖二糖,可用作优质功能性食品或膳食益生元。
[0113]
β-甘露聚糖酶可通过酶解法、化学法或物理法或这些方法结合的策略,部分水解甘露聚糖的β-(1
→
4)-糖苷键产生甘露寡糖,用作功能性食品或膳食益生元,具有调节肠道菌群和增强免疫调节活性功能,还具有抗癌、抗氧化、抗炎、抗冻保护剂、抗应激和抗糖尿病等作用。但纯化的甘露二糖功能及其与其他己糖二塘的抗氧化活性未有报道。本实施例首次发现,灰盖拟鬼伞内切β-1,4-甘露聚糖酶(ccman5c)水解角豆胶半乳甘露聚糖形成的终产物中主要产物是甘露二糖(约占水解寡糖产物量的50%),可应用于水解β-半乳甘露聚糖制备甘露二糖。而以往报道的真菌β-甘露聚糖酶水解半乳甘露聚糖都不能产生一种占优势的水解产物寡糖。本发明实施例1提出重组的成熟的甘露聚糖酶mccman5c水解角豆胶半乳甘露聚糖经分子排阻色谱法分离纯化获得甘露二糖的方法,所制得甘露二糖对dpph自由基的清除活性是同样浓度甘露聚糖5-7倍,是其他纤维二糖、昆布二糖、乳糖、龙胆二糖、蔗糖的dpph自由基清除率3.5-8.0倍,可用作优质功能性食品或膳食益生元。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。