1.本发明属于分子生物学技术领域,涉及一种木薯吡哆醛激酶基因及其在提高植物耐盐性的应用。
背景技术:
2.由于盐碱土中含有大量可溶性的盐离子(特别是大量的可交换性钠)。从而抑制了植物的正常生长,同时由于自然和人为因素的影响,盐碱地面积仍在不断增加。因此,对盐碱土地的改良与应用具有重大的意义。到目前为止,治理盐碱地主要由物理改良、化学改良和生物改良,其中物理改良和化学改良土壤不仅耗资巨大,还有可能会造成土壤次生盐碱化问题,而通过种植耐盐碱植物的生物改良不仅耗资少,而且效果显著。因此,利用转基因技术培育耐盐转基因植物新品种成为利用盐碱地的重要途径之一,提高植物的耐盐碱能力是缓解盐碱地对植物影响的一个有效生物措施,同时可以产生较好的生态和经济效益,促进农业的可持续发展。
3.木薯是大戟科木薯属植物,是一种重要的粮食作物。据统计,世界上约八亿人以富含淀粉的木薯块根作为主食。木薯富含淀粉、蛋白质、纤维素、矿物质和维生素等营养物质,从块根提取的木薯粉,是小麦和其它谷物的最佳替代品之一。木薯原产于巴西和美洲的热带地区,生长在热带地区,对各种环境条件有较强的耐受性,是生产力最强的食用植物之一。因此,发掘木薯耐盐基因是目前木薯抗逆遗传资源与品种改良研究的热点。
技术实现要素:
4.本发明的目的是提供一种木薯吡哆醛激酶基因及其在提高植物耐盐性的应用,该基因具有提高植物耐盐性的功能。
5.上述木薯吡哆醛激酶基因,所述基因的序列如seq id no.1所示,具有1011个碱基,其编码的氨基酸序列如seq id no.2所示。
6.本发明还提供了一种表达载体,其含有所述的木薯吡哆醛激酶基因;所述表达载体如 pet-28a、pcambia2301、psp72、prokii、pbin438、pcambia1302、pcambia1301、 pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb等。
7.本发明还提供一种宿主细胞,其含有所述的表达载体转化的原核细胞或真核细胞。
8.本发明的另一个目的是提供上述木薯吡哆醛激酶基因在提高植物耐盐性中的用途。
9.本发明还提供了一种提高植物耐盐性的方法,将seq id no.1所示的木薯吡哆醛激酶基因构建重组表达载体导入受体植物中,可以获得过表达木薯吡哆醛激酶基因的耐盐转基因植物。
10.其中,所述重组表达载体可通过使用农杆菌介导、ti质粒、植物病毒载体、直接dna 转化、微注射、电穿孔等常规生物技术方法导入植物细胞或组织。
11.其中,所述方法还包括从导入seq id no.1所示的基因的受体植物中筛选所述木薯吡哆醛激酶基因表达的植物,得到转基因植物的步骤。
12.其中,所述受体植物为木薯。
13.其中,所述转基因植物理解为不仅包含将所述基因转化受体植物得到的第一代转基因植物,也包括其子代。对于转基因植物,可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述转基因植物包括种子、愈伤组织、完整植株和细胞。
14.本发明的有益效果
15.本发明提供了一种木薯吡哆醛激酶基因、该基因编码的蛋白及其用于提高植物耐盐性的用途。在木薯植株中沉默编码该基因的表达会降低木薯的耐盐能力,这说明在木薯植株中过表达该基因可以增强植株的耐盐能力,通过将基因导入受体植物中进行过表达,获得转基因植物,可以提高转基因植物的耐盐性。本发明提供的木薯吡哆醛激酶基因是一种新的提高植物耐盐性的基因,为改良耐盐性较差的木薯品种提供了新的思路,也为其他作物利用异源基因技术提高耐盐性提供了理论支持,可用于植物分子育种,能够解决传统选育的效率低和周期长的难题。
附图说明
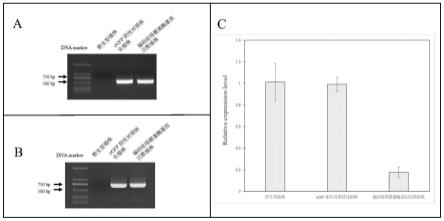
16.图1为木薯植株的pcr扩增验证和荧光定量pcr实验图;
17.图2为基因功能鉴定结果图。
具体实施方式
18.以下实施例进一步说明本发明的内容,但不应理解为对本发明的限制。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。
19.下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所使用的材料、试剂等,如无特殊说明,均可从商业途径得到。
20.实施例1木薯吡哆醛激酶基因序列的克隆
21.1、根据拟南芥atsos4基因信息在木薯基因组中筛选同源基因
22.从phytozome数据库(https://phytozome-next.jgi.doe.gov/)中下载拟南芥atsos4基因 (at5g37850.2)的氨基酸序列,然后利用phytozome数据库的blast search功能进行同源比对,鉴定获得一个木薯基因组中atsos4的同源序列为木薯吡哆醛激酶基因 (manes.07g054300.2)。
23.2、木薯吡哆醛激酶基因
24.从phytozome数据库上下载木薯吡哆醛激酶基因的cds序列如seq id no.1所示,进行人工合成克隆,该基因序列具有1011个碱基,编码的蛋白长为336个氨基酸残基,其序列如seq id no.2所示。
25.实施例2基因功能鉴定
26.1、烟草脆裂病毒(trv)诱导木薯编码吡哆醛激酶基因的沉默
27.(1)使用在线分析网站sng-vigs(https://vigs.solgenomics.net/)进行分析,得
到预测沉默效率最高的300bp长度的序列,其序列如seq id no.3所示,然后在bamhi酶切位点进行单酶切,将该序列克隆到瞬时表达载体ptrv2上,得到重组瞬时表达载体 ptrv2::seq id no.3。
28.(2)利用热激法将重组瞬时表达载体载体ptrv2::seq id no.3导入根癌农杆菌菌株lba4404,获得携带表达载体ptrv2::seq id no.3的转化菌株;将空载配套载体质粒 ptrv1导入根癌农杆菌菌株lba4404的转化菌株,获得携带ptrv1的转化菌株;将空载载体ptrv2-egfp导入根癌农杆菌菌株lba4404的转化菌株,获得携带ptrv2-egfp的转化菌株。
29.(3)使用木薯辐选01作为材料,将20cm长的茎段种植在含有100%珍珠岩的盆中, 26℃培养30d。将携带ptrv1的根癌农杆菌菌株lba4404培养液,分别与携带ptrv1 的根癌农杆菌菌株lba4404培养液和携带ptrv2-egfp的根癌农杆菌菌株lba4404培养液(菌液浓度od值分别为0.5)按照v:v=1:1混合,制备成两种混合接种菌液。通过压渗法分别将两种混合菌液导入木薯植株,每株选择3-4片叶进行接种转化。以不接种的植株为野生型对照,接种由含有ptrv1和ptrv2-egfp菌液制备的混合菌液的植株作为egfp 阴性对照转化植株,接种由含有ptrv1和表达载体ptrv2::seq id no.3菌液制备的混合菌液的植株作为编码吡哆醛激酶基因沉默植株。
30.(4)继续培养10d,采集野生型叶片及转化植株的接种叶片,然后使用商业化试剂盒,分别提取木薯叶片的dna;
31.使用ptrv1特异性引物对木薯dna进行pcr扩增验证,所述ptrv1特异性引物的上游引物序列为:5
’‑
agcagcaaccgacgactt-3’(seq id no.4),下游引物序列为: 5
’‑
accaactccttcttctcagact-3’(seq id no.5),pcr扩增产物的电泳图如图1-a 所示;
32.使用ptrv2特异性引物对木薯dna进行pcr扩增验证,所述ptrv2特异性引物的上游引物序列为:5
’‑
tgcgctaatcaacatgggaga-3’(seq id no.6),下游引物序列为:5
’‑
ggtatcacccaccctctgaa-3’(seq id no.7),pcr扩增产物的电泳图如图 1-b所示;
33.(5)利用商业化试剂盒,提取野生型植株、egfp阴性对照转化植株和编码吡哆醛激酶基因沉默植株转化后新生叶片的rna,再反转录合成cdna,以cdna为模板,利用荧光定量pcr分析验证野生型植株、egfp阴性对照转化植株与编码吡哆醛激酶基因沉默植株中编码吡哆醛激酶基因的表达(图1-c)。从图1-c中可以看出,编码吡哆醛激酶基因沉默植株中编码吡哆醛激酶基因的表达量远远低于野生型植株和egfp阴性对照转化植株。
34.经过上述检测分析后,表明trv病毒成功诱导木薯编码吡哆醛激酶基因的沉默。
35.2、功能鉴定和结果
36.使用200mm nacl溶液浇灌处理野生型植株、egfp阴性对照转化植株和编码吡哆醛激酶基因沉默植株3d,然后改用250mm nacl溶液浇灌处理6d,进行观察。结果表明,野生型和egfp阴性对照转化植株在250mm nacl的胁迫下没有明显变化,而编码吡哆醛激酶基因沉默植株在250mm nacl的胁迫下叶片出现明显萎焉(图2-a)。同时向野生型植株、egfp阴性对照转化植株和编码吡哆醛激酶基因沉默植株浇灌不含nacl的蒸馏水,培养9d,作为对照,结果显示野生型植株、egfp阴性对照转化植株和编码吡哆醛激酶基因沉默植株生长无明显变化(图2-b)。
37.这些试验显示了沉默编码吡哆醛激酶基因的表达会降低木薯的耐盐能力,说明编码吡哆醛激酶基因在木薯耐盐方面发挥重要作用。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。