养基和补料批次方法,来改善抗体的糖基化水平,但该方法流程较为复杂,受培养基限 制较大,很难广泛的应用到实际生产中去。根据janike ehret[1]、michael j.gramer[2]、 yuzhou fan[3]、rodriguez[4]等研究结果,发现添加800μm 2-f-peracetyl fucose可降低 抗体的76.1%的核心岩藻糖基化类型;添加kifunensine可增加抗体的高甘露糖类型;添 加20
×
umg(尿嘧啶,锰离子,半乳糖)可以提高抗体23%的半乳糖基化类型;dmso (二甲基亚砜)的添加可降低γ-干扰素的唾液酸化糖型;添加高浓度的n
4
可降低重组肿 瘤坏死因子受体(tnfr)-igg融合蛋白的半乳糖化以及唾液酸化的糖型结构。根据patrickhossler[5]、stanley chung[6]等研究结果,发现补加生物类黄酮类和芦丁等抗氧化剂可降 低酸性峰比例。但这些都是以糖型和电荷异构体其中一种质量属性的调节为目的的方法, 对另外一种的调节效果不显著。
[0007]
电荷异质性中酸性峰和碱性峰具有相似的化学性质,通过后期纯化分离控制电荷异 质性具有一定的难度,而通过控制细胞培养工艺流程来控制抗体电荷异质性的方法也具 有一定的挑战性,成为本领域一直难于解决的问题。
[0008]
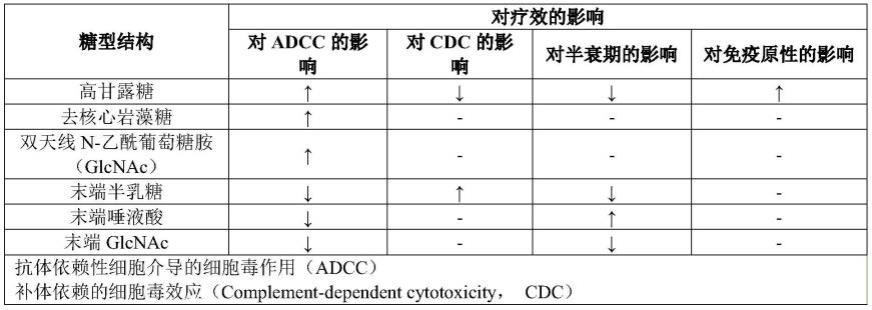
表1糖型结构对临床疗效的影响
[0009][0010]
因抗体及其它治疗性蛋白质的糖型结构和电荷异质性在产品的疗效方面都有非常重 要的作用,对于目标糖型和电荷异构体的调控就显得尤为重要。目前关于糖型和电荷异 质性分开调控的方法有很多,大部分调控的目标是集中于调节某种糖型结构或调节某种 电荷异质性,对于同时调控糖型和电荷异构体的方法研究的较少。
技术实现要素:
[0011]
本发明旨在提供一种同时调控糖型(半乳糖化糖型)和电荷异质性(酸性异构体)的 方法,使得关注产品半乳糖糖型和酸性异构体含量的质量属性通过本发明方法进行调控 后,蛋白质量较好,可快速地满足目标需求。且本发明的工艺简单,经济高效,适宜工业 生产。
[0012]
据报道,n-乙酰葡萄糖胺对n-乙酰葡萄糖胺类糖基化分布有影响;芦丁、迷迭香酸 (ra)和没食子儿茶素没食子酸酯(egcg)等仅对电荷异质性有影响。mn
2
、尿苷和 半乳糖对半乳糖基化分布有影响。
[0013]
本发明的目的是提供一种可同时调节半乳糖基化类型和酸性峰电荷异构体的方法, 添加二甲基亚砜(dmso)可提高半乳糖基化含量,降低的酸性峰含量;添加nh4cl可 降低半乳糖基化含量,同时降低的酸性峰含量。
[0014]
本发明提供一种同时调控糖型和电荷异质性的方法,即在细胞培养过程中添加二甲 基亚砜(dmso)、nh4cl等小分子化合物,来调节重组cho细胞表达产物的电荷异质 性和糖型结构。该方法能在降低抗体酸性峰的同时改良抗体的糖基化水平,该方法可以 应用于生物类似药的开发以及抗体药物的质量调节上。相比于glcnac、芦丁、迷迭香酸 和egcg等添加物对糖基化和电荷异构体的单一调节作用,采用本技术可对糖基化结构 和电荷异构体进行同时调控。
[0015]
为了克服现有技术的缺陷和不足,本发明的技术方案提供了一种调节细胞表达产物 的糖型和/或电荷异质性的方法,在细胞培养的过程中添加调节剂,所述调节剂针对调节 蛋白酸性峰和/或碱性峰,所述调节剂为dmso和/或nh4cl;优选地,所述调节剂还可 以调节糖型;更优选地,所述糖型为半乳糖化糖型。
[0016]
在某些具体实施方案中,所述细胞为cho-k1细胞;优选地,所述cho-k1细胞包 括含gs基因的表达载体,并且所述gs基因下游插入了外源基因。
[0017]
在某些具体实施方案中,当调节剂为dmso时,其浓度为1-5%;其中%为体积百分 比;当调节剂为nh4cl时,其浓度为20-55mm。
[0018]
在某些具体实施方案中,当调节剂为dmso时,其浓度为2%-4%;当调节剂为nh4cl 时,其浓度为25-50mm。
[0019]
在某些具体实施方案中,当调节剂为dmso和nh4cl时,其浓度分别为1-2%的dmso和25-50mm的nh4cl。
[0020]
在某一些具体实施方案中,当调节剂为dmso和nh4cl时,其浓度分别为25mmnh4cl 1%dmso或50mm nh4cl 2%dmso。
[0021]
在某些具体实施方案中,所述调节剂还包括umg。
[0022]
在某一些具体实施方案中,当调节剂包括umg时,所述调节剂为1-5%umg 1-2% dmso;更优选地,所述调节剂为4%umg 1%dmso或4%umg 2%dmso。
[0023]
在某些具体实施方案中,所述方法包括:将所述细胞接种至适合细胞生长的培养基 中,在第5、7、9、11天时添加调节剂。
[0024]
上述方法可以采用本领域常见的化学成分限定的培养基进行分批补料(fed-batch)模 式的细胞培养,在细胞培养中后期添加dmso和nh4cl的培养模式。例如,所述培养基 可为适用于cho细胞培养的无血清化学成分限定的,无动物源成分的本领域常见的培养 基。所述cho细胞培养采取分批补料(fed-batch)模式。
[0025]
在某些具体实施方案中,当调节剂为dmso时,在第5、7、9、11天时添加所述调 节剂,优选在第7天时添加所述调节剂;当调节剂为nh4cl时,在第5、7、9、11天时 添加所述调节剂。
[0026]
在某些具体实施方案中,所述方法的培养条件为37℃、125rpm、80%湿度和5.5% co2;优选地,所述培养条件还包括培养基中葡萄糖浓度为6-8g/l。
[0027]
本发明第二方面提供一种如本发明第一方面所述的调节剂在培养细胞中的应用。
[0028]
在符合本领域常识的基础上,上述各优选条件,可任意组合,即得本发明各较佳实 例。
[0029]
本发明所用试剂和原料均市售可得。
[0030]
本发明的积极进步效果在于:
[0031]
本发明提供的方法既可以降低酸性异构体的比例,还可以提高半乳糖糖型,而且对 抗体表达量基本没有影响。总地来说,本发明操作简单,成本较低,能够广泛地适用于治 疗性抗体和治疗性蛋白类生物类似药和新药的开发和商业化生产。
附图说明
[0032]
图1展示了n末端糖型分布图。图1的a表示高甘露糖化糖型;图1的b表示核 心岩藻糖化糖型;图1的c表示末端n-乙酰葡萄糖胺化糖型结构;图1的d表示半乳糖 化糖型;图1的e表示唾液酸化糖型。
[0033]
图2a是实施例1中培养天数和活细胞密度结果图;图2b是实施例1中培养天数 和细胞活率结果图;图2c是实施例1中归一化的蛋白表达量。
[0034]
图3是实施例1中补加dmso的糖基化分布结果。
[0035]
图4是实施例1中补加dmso的电荷异质性结果。
[0036]
图5a是实施例2中培养天数和活细胞密度结果图;图5b是实施例2中培养天数 和细胞活率结果图;图5c是实施例2中归一化的蛋白表达量。
[0037]
图6是实施例2中补加nh4cl的糖基化分布结果。
[0038]
图7是实施例2中补加nh4cl的电荷异质性结果。
[0039]
图8a是实施例3中培养天数和活细胞密度结果图;图8b是实施例3中培养天数 和细胞活率结果图;图8c是实施例3中归一化的蛋白表达量。
[0040]
图9是实施例3中补加不同浓度dmso的糖基化分布结果。
[0041]
图10是实施例3中补加不同浓度dmso电荷异质性结果。
[0042]
图11a是实施例4中培养天数和活细胞密度结果图;图11b是实施例4中培养天数 和细胞活率结果图;图11c是实施例3中归一化的蛋白表达量。
[0043]
图12是实施例4中补加不同浓度nh4cl的糖基化分布结果。
[0044]
图13是实施例4中补加不同浓度nh4cl的电荷异质性结果。
[0045]
图14a是实施例5中培养天数和活细胞密度结果图;图14b是实施例5中培养天 数和细胞活率结果图;图14c是实施例5中归一化的蛋白表达量。
[0046]
图15是实施例5中补加dmso、nh4cl与umg两两相互组合添加物的糖基化分 布结果。
[0047]
图16是实施例5中补加dmso、nh4cl与umg两两相互组合添加物的电荷异质 性结果。
具体实施方式
[0048]
下面通过实施例的方式进一步说明本发明,但并不因此将本发明限制在所述的实施 例范围之中。下列实施例中未注明具体条件的实验方法,按照常规方法和条件,或按照 商品说明书选择。
[0049]
图1为n末端糖型分布图。a表示高甘露糖化糖型;b表示核心岩藻糖化糖型;c 表示末端n-乙酰葡萄糖胺化糖型结构;d表示半乳糖化糖型;e表示唾液酸化糖型。
[0050]
实施例1在cho细胞系表达单克隆抗体igg的细胞培养过程中添加dmso等添加 物进行质量调节
[0051]
1.1细胞培养
[0052]
将重组的cho-k1细胞株复苏后,经过2-3次传代扩增后的细胞,以0.5
×
106细胞 /ml的密度接种至化学限定的基础培养基(本领域常规的适合cho-k1细胞株生长的基 础培养基)中,以初始培养体积为15ml的50ml摇管中培养。培养条件为:37℃、180 rpm、80%湿度和5.5%co2。在培养第5天补加培养基后降温至32.5℃。
[0053]
dmso实验组:第7天添加dmso;终浓度为2%。
[0054]
n-乙酰葡萄糖胺对照组:第7天添加n-乙酰葡萄糖胺;终浓度为20mm。
[0055]
芦丁对照组:第5、7、9、11天添加芦丁;终浓度为0.2g/l。
[0056]
迷迭香酸(ra)对照组:第5、7、9、11天添加迷迭香酸;终浓度0.1g/l。
[0057]
egcg对照组:第11天添加egcg。终浓度为0.13g/l。
[0058]
并设置2组除补料培养基和葡萄糖外不补加其它任何添加物的空白组作为空白对照。
[0059]
同时,在第3、5、7、9、11、13和15天取样检测细胞密度和活率以及营养物质,如 果葡萄糖浓度低于5g/l,通过补加400g/kg的葡萄糖溶液使细胞培养液中的葡萄糖浓度 维持在6-8g/l。当活率低于70%或培养周期达到15天结束培养(以先到者为准),通过 离心和过滤收获上清液。
[0060]
1.2检测方法与仪器
[0061]
通过proa-hplc(蛋白a-高效液相色谱)法检测上清液的抗体表达量。用于电荷异 质性和糖型检测的质量分析的上清液样品,首先需经过蛋白a亲和层析一步纯化法进行 纯化。电荷异质性通过iec-hplc(离子交换-高效液相色谱)法检测一步纯化后的样品 进行分析。n段糖基化结构的分析方法为:首先用内切糖苷酶(pngase f)在37℃处理 样品,再用冰乙醇沉淀蛋白得到含寡糖的上清液,经真空干燥去除上清液中的冰乙醇。 然后用2-氨基-苯甲酰胺(2-ab)标记干燥后的多糖凝胶物,使其带上荧光基团,最后用 uplc-fld法分析鉴定。通过比较样品和标准品的色谱图,鉴定样品蛋白的糖型种类, 并根据不同类型的寡糖吸收峰面积计算其相对百分比。
[0062]
本实施例中,细胞密度和活率的检测采用贝克曼的细胞计数仪检测,葡萄糖和乳酸 等营养物质采用希尔曼的生化分析仪检测。hplc仪器的型号为:waters alliance 2695带 waters 2998光电二极管(pda)检测器。uplc仪器的型号为:waters alliance 2695h class。
[0063]
1.3结果
[0064]
图2a-图2c展示了细胞培养表现的结果,细胞生长基本正常。相比于对照组,添加 dmso的这一组,尽管细胞活率下降得较快,但可提前2天结束培养,提高培养效率, 且最终的蛋白表达量基本不受影响。
[0065]
从糖型分析的结果来看,如图3所示,添加2%dmso会改变糖基化分布(相对比), 提高半乳糖基化含量其中,半乳糖绝对占比为11%。n-乙酰葡萄糖胺(glcnac)会降低 半乳糖化糖型的比例,与glcnac对照组相比,dmso对半乳糖基化水平有正向调控作 用。其中,图中也涉及不同对照组对高甘露糖化的改变,但是由于高甘露糖化结构本身 占比为4%左右,即使相对值变化50%引起高甘露糖绝对值的变化仍小于2%。图3中引 起高甘露糖基化比例变化最大的组别是添加芦丁的对照组,使得高甘露糖基化的相对百 分比增加了43%,
由此可知不同对照组对高甘露糖化的改变均可忽略不计。结合上述论 述,可认为除dmso和n-乙酰葡萄糖胺对照组外,其它添加物对照组对糖基化分布没有 显著的影响。
[0066]
分析dmso对电荷异质性的影响,结果见图4。与空白对照组相比,添加2%dmso 会导致电荷异构体发生变化,降低酸性峰,提高碱性峰。与之前报道的迷迭香酸和芦丁 等添加物对照组相比,添加dmso对酸性峰的负调控作用更明显。
[0067]
综上,补加2%的dmso可以同时调节糖基化分布和电荷异质性,在提高半乳糖基 化比例的同时降低酸性峰比例,而且对产品产量基本没有影响。
[0068]
实施例2在cho细胞系表达单克隆抗体的细胞培养过程中添加nh4cl等添加物进 行质量调节
[0069]
2.1细胞培养
[0070]
将重组的cho-k1细胞株复苏后,经过2-3次传代扩增后的细胞,以0.5
×
106细胞 /ml的密度接种至化学成分限定的基础培养基(同实施例1)中,以初始培养体积为15 ml的50ml摇管中培养。培养条件为:37℃、180rpm、80%湿度和5.5%co2。在培养 第5天补加培养基后降温至32.5℃。
[0071]
50mm nh4cl实验组:第5、7、9、11天添加nh4cl;终浓度为50mm。
[0072]
100mm nh4cl实验组:第5、7、9、11天添加nh4cl;浓度为100mm。
[0073]
mncl2对照组:第5、7、9、11天添加mncl2;终浓度20μm。
[0074]
umg对照组:第5、7、9、11天添加umg;母液中尿苷、氯化锰和半乳糖的浓度 分别为24mm、48μm和120mm,总的添加比例为4%。
[0075]
芦丁对照组:第5、7、9、11天添加芦丁;终浓度0.2g/l。
[0076]
迷迭香酸(ra)对照组:第5、7、9、11天添加迷迭香酸;终浓度0.1g/l。
[0077]
egcg对照组:第5、7、9、11天添加egcg;终浓度0.13g/l。
[0078]
并设置2组除补料培养基和葡萄糖外不补加其它任何添加物的空白组作为空白对照。
[0079]
在第3、5、7、9、11、13和15天取样检测细胞密度和活率以及营养物质,如果葡萄 糖浓度低于5g/l,通过补加400g/kg的葡萄糖溶液使细胞培养液中的葡萄糖浓度维持在 6-8g/l。当活率低于70%或培养周期达到15天结束培养(以先到为准),通过离心和过 滤收获上清液。
[0080]
2.2检测方法与仪器
[0081]
同实施例1的1.2。
[0082]
2.3结果
[0083]
图5a-图5c展示了细胞培养表现的结果,补加nh4cl在一定程度上降低了蛋白表 达量。
[0084]
从糖基化分析的结果来看,如图6所示,添加50mm-100mm nh4cl会影响糖型分 布结果(相对比),降低半乳糖基化比例,增加高甘露糖。nh4cl的浓度越低,半乳糖基 化的降低幅度越大,高甘露糖增加的幅度越小。
[0085]
结合图7电荷异质性分析的结果来看,与其他对照组相比,添加nh4cl会影响电荷 异构体分布,添加低浓度的nh4cl,其酸性峰的降低比例越大,相应的碱性峰的增加比 例也越大。与之前报道的迷迭香酸、芦丁等添加物对照组相比,50mm nh4cl对酸性峰 的负调控
作用更明显。结合图6和图7,在细胞培养中后期额外补加nh4cl,可对糖基化 结构和电荷异构体进行同时调控。
[0086]
综上,补加nh4cl可在调节糖型的同时改变电荷异质性,降低半乳糖基化含量并减 小酸性峰比例。50mm的nh4cl对产品产量的负影响为30%左右,且nh4cl浓度越低对 产品产量的负影响越小。
[0087]
实施例3在cho细胞系表达单克隆抗体的细胞培养过程中添加不同浓度的dmso 进行质量调节
[0088]
3.1细胞培养
[0089]
为确定dmso对糖型和电荷异质性的调控的适用浓度,将重组的cho-k1细胞株复 苏后,经过2-3次传代扩增后的细胞,以0.5
×
106细胞/ml的密度接种至化学成分限定 的基础培养基(同实施例1)中,以初始培养体积为50ml的250ml摇瓶中培养。培养 条件为:37℃、125rpm、80%湿度和5.5%co2。在培养第5天补加培养基后降温至32.5℃。
[0090]
0.1%dmso实验组:第7天添加dmso,dmso的终浓度为0.1%。
[0091]
0.5%dmso实验组:第7天添加dmso,dmso的终浓度为0.5%。
[0092]
1%dmso实验组:第7天添加dmso,dmso的终浓度为1%。
[0093]
2%dmso实验组:第7天添加dmso,dmso的终浓度为2%。
[0094]
4%dmso实验组:第7、9天分两次添加dmso,dmso的终浓度为4%。
[0095]
并设置1个除补料培养基和葡萄糖外不补加其它任何添加物的空白组作为空白对照。
[0096]
在第3、5、7、9、11、13和15天取样检测细胞密度和活率以及营养物质,如果葡萄 糖浓度低于5g/l,通过补加400g/kg的葡萄糖溶液使细胞培养液中的葡萄糖浓度维持在 6-8g/l。当活率低于70%或培养周期达到15天结束培养(以先到者为准),通过离心和 过滤收获上清液。
[0097]
3.2检测方法与仪器
[0098]
同实施例1的1.2。
[0099]
3.3结果
[0100]
图8a-图8c展示了细胞培养表现的结果,4%高浓度的dmso对细胞生长的负影响 较大,细胞活率下降得较快。从蛋白表达量结果来看,单独添加4%dmso降低了20% 的蛋白表达量。dmso的添加浓度在0.1%-2%范围内,对蛋白表达量没有负影响。
[0101]
从糖基化分析的结果来看,如图9所示,添加2%-4%dmso会影响糖型分布结果, 提高了21%-32%半乳糖基化比例(相对比)。dmso的浓度越高,半乳糖基化的提高的幅 度越大。
[0102]
结合图10电荷异质性分析的结果来看,与对照组相比,添加2%-4%的dmso对电 荷异质性的影响较大,降低了15%-23%的酸性电荷异构体(相对比)。dmso浓度越高, 其酸性峰的降低比例越大,相应的碱性峰或主峰的增加比例也越大。结合图8a-图8c和 图9,在细胞培养中后期额外补加不同浓度的dmso(0.1%-4%),可对糖基化结构和电 荷异构体进行同时调控。
[0103]
综上,补加2%-4%dmso可在调节糖型的同时改变电荷异质性,降低半乳糖基化含 量并减小酸性峰比例。2%的dmso对产品产量没有影响,4%的dmso对蛋白产量的负 影响为
20%,dmso浓度越低对产品产量的负影响越小。
[0104]
实施例4在cho细胞系表达单克隆抗体的细胞培养过程中添加不同浓度的nh4cl 进行质量调节
[0105]
4.1细胞培养
[0106]
为确定nh4cl对糖型和电荷异质性的调控的适用浓度,将重组的cho-k1细胞株复 苏后,经过2-3次传代扩增后的细胞,以0.5
×
106细胞/ml的密度接种至化学成分限定 的基础培养基(同实施例1)中,以初始培养体积为50ml的250ml摇瓶中培养。培养 条件为:37℃、125rpm、80%湿度和5.5%co2。在培养第5天补加培养基后降温至32.5℃。
[0107]
5mm nh4cl实验组:第5、7、9、11天均匀添加nh4cl,nh4cl的终浓度为5 mm;
[0108]
10mm nh4cl实验组:第5、7、9、11天均匀添加nh4cl,nh4cl的终浓度为10 mm;
[0109]
25mm nh4cl实验组:第5、7、9、11天均匀添加nh4cl,nh4cl的终浓度为25 mm;
[0110]
50mm nh4cl实验组:第5、7、9、11天均匀添加nh4cl,nh4cl的终浓度为50 mm;
[0111]
并设置1个除补料培养基和葡萄糖外不补加其它任何添加物的空白组作为空白对照。
[0112]
在第3、5、7、9、11、13和15天取样检测细胞密度和活率以及营养物质,如果葡萄 糖浓度低于5g/l,通过补加400g/kg的葡萄糖溶液使细胞培养液中的葡萄糖浓度维持在 6-8g/l。当活率低于70%或培养周期达到15天结束培养(以先到者为准),通过离心和 过滤收获上清液。
[0113]
4.2检测方法与仪器
[0114]
同实施例1的1.2。
[0115]
4.3结果
[0116]
图11a-图11c展示了细胞培养表现的结果,50mm高浓度的nh4cl对细胞生长的 负影响较大,细胞活率下降得较快。从蛋白表达量的结果来看,单独添加50mm nh4cl 降低了22%的蛋白表达量。nh4cl的添加浓度在5mm-25mm范围内,对蛋白表达量基 本没有影响。
[0117]
图13是本实施例中补加不同浓度nh4cl的电荷异质性结果。从糖基化分析的结果 来看,如图12所示,添加5mm-50mm nh4cl会影响糖型分布结果,降低12%-43%半 乳糖基化比例(相对比),增加高甘露糖。nh4cl的浓度越高,半乳糖基化的降低幅度越 大,当浓度达到25mm时,半乳糖基化含量降低的幅度基本趋于饱和,相对比为40%。 进一步增加nh4cl的浓度至50mm,半乳糖基化结构降低的比例仅为43%,同时大幅度 提高了高甘露糖基化的比例。结合图11a-图11c和图12,在细胞培养中后期额外补加一 系列不同浓度nh4cl(5mm-50mm)),可对糖基化结构和电荷异构体进行同时调控。
[0118]
结合图7电荷异质性分析的结果来看,与对照组相比,添加nh4cl会影响电荷异构 体分布,添加较低浓度的nh4cl(5mm-25mm),其酸性峰的降低比例较小,相对比在10%以内。nh4cl的添加浓度达到25mm-50mm时,酸性峰含量降低的相对比例为10%
‑ꢀ
26%。
[0119]
综上,补加25mm-50mm的nh4cl可在调节糖型的同时改变电荷异质性,降低半 乳糖基化含量并减小酸性峰比例。50mm的nh4cl对产品产量的负影响为22%,且nh4cl 浓度越低对产品产量的负影响越小。
[0120]
实施例5在cho细胞系表达单克隆抗体的细胞培养过程中添加不同浓度的dmso、 nh4cl与umg两两相互组合的添加物进行质量调节
[0121]
5.1细胞培养
[0122]
为确定dmso、nh4cl与umg相互组合对糖型和电荷异质性的调控的有效组合添 加物,将重组的cho-k1细胞株复苏后,经过2-3次传代扩增后的细胞,以0.5
×
106细 胞/ml的密度接种至化学成分限定的基础培养基(同实施例1)中,以初始培养体积为50 ml的250ml摇瓶中培养。培养条件为:37℃、125rpm、80%湿度和5.5%co2。在培养 第5天补加培养基后降温至32.5℃。
[0123]
根据表3的组合情况,在第5天、第7天、第9天和第11天补加dmso、nh4cl和 umg两两相互组合的添加物。相应组合及终浓度具体见表3。同时设置1个除补料培养 基和葡萄糖外不补加其它任何添加物的空白组作为空白对照。实施例3-5中dmso、 nh4cl和umg汇总的添加方案如表3所示。
[0124]
表3dmso、nh4cl及umg添加方案
[0125][0126]
在第3、5、7、9、11、13和15天取样检测细胞密度和活率以及营养物质,如果葡萄 糖浓度低于5g/l,通过补加400g/kg的葡萄糖溶液使细胞培养液中的葡萄糖浓度维持在 6-8g/l。当活率低于70%或培养周期达到15天结束培养(以先到者为准),通过离心和 过滤收获上清液。
[0127]
5.2检测方法与仪器
[0128]
同实施例1的1.2。
[0129]
5.3结果
[0130]
图14a-图14c展示了细胞培养表现的结果,50mm nh4cl 2%dmso和4%umg 50mm nh4cl对细胞生长的负影响较大,细胞活率下降得较快,于第13天结束培养。 结合蛋白表达量结果来看,50mm nh4cl 2%dmso和4%umg 50mm nh4cl对蛋 白产量的影响最大,降低了33%-55%的蛋白表达量;4%umg 25mm nh4cl的影响次 之,减少了14%的蛋白表达量。其它3种组合添加物对蛋白表达量没有负影响。
[0131]
从糖基化分析的结果来看,如图15所示,添加25mm nh4cl 1%dmso、50mmnh4cl 2%dmso、4%umg 1%dmso、4%umg 2%dmso、4%umg 25mmnh4cl和4%umg 50mm nh4cl会影响糖型分布结果(相对比)。其中,25mm nh4cl 1%dmso和50mm nh4cl 2%dmso降低35%左右的半乳糖基化比例(相对比), 50mm nh4cl 2%dmso大幅度地增加高甘露糖比例;4%umg 1%dmso和4%umg 2%dmso大幅度提高半乳糖基化比例,相对比增加了118%-142%,高甘露糖基化的含 量基本不受影响;4%umg 25mm nh4cl和4%umg 50mm nh4cl对半乳糖基化的 影响不明显,半乳糖基化含量的变化幅度在14%(相对比)以内。
[0132]
从图16电荷异质性分析的结果来看,与对照组相比,添加25mm nh4cl 1%dmso、 50mm nh4cl 2%dmso、4%umg 1%dmso、4%umg 2%dmso、4%umg 25 mm nh4cl和4%umg 50mm nh4cl会影响电荷异构体分布。其中,添加25mm nh4cl 1%dmso和50mm nh4cl 2%dmso,降低24%-36%的酸性峰比例,相应的增加了 碱性峰的比例;补加50mm nh4cl 1%dmso和4%umg 2%dmso,降低16%-22% 的酸性峰含量;4%umg 25mm nh4cl和4%umg 50mm nh4cl对电荷异质性的影 响呈现出相反的方向,4%umg 25mm nh4cl降低27%的酸性异构体比例,而4%umg 50mm nh4cl提高56%的酸性峰比例。上述降低或提高的比例均是相对百分比。在细 胞培养中后期额外补加dmso(1%,2%)、nh4cl(25mm,50mm)与umg两两相互 组合的添加物,可对糖基化结构和电荷异构体进行同时调控。
[0133]
综上所述,补加25mm nh4cl 1%dmso、50mm nh4cl 2%dmso、4%umg 1%dmso和4%umg 2%dmso可在调节糖型的同时改变电荷异质性。其中,25mmnh4cl 1%dmso和50mm nh4cl 2%dmso降低半乳糖基化含量并减小酸性峰比 例。25mm nh4cl 1%dmso降低35%左右的半乳糖比例,同时减小24%的酸性峰比 例。50mm nh4cl 2%dmso在降低35%半乳糖基化含量的同时,对酸性峰的负调控 更显著,降低的比例为36%,但对产品产量的负影响达到33%。4%umg 1%dmso和4%umg 2%dmso提高半乳糖基化含量同时降低酸性峰比例。dmso浓度由1%增加 至2%,半乳糖基化含量的增加幅度从118%提高至142%,酸性峰比例的降低幅度由16% 增加至22%,且这两种组对蛋白表达量均没有负影响。
[0134]
综上实施例1-5,添加2%-4%二甲基亚砜(dmso)可提高21%-32%的半乳糖基化 含量,同时降低15%-31%的酸性峰含量;添加25-50mm nh4cl可降低38%-48%的半乳 糖基化含量,同时降低10-33%的酸性峰含量。在dmso、nh4cl和umg相互组合添加 的情况下,1%-2%dmso umg可提高118%-142%的半乳糖基化比例,同时降低16%
‑ꢀ
21%的酸性峰含量;25mm nh4cl 1%dmso和50mm nh4cl 2%dmso可降低35% 左右的半乳糖基化含量,同时减小24%-34%的酸性峰比例。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。