用于眼部基因递送的aav载体变体
1.本发明涉及腺相关病毒衣壳多肽序列及其在靶向光感受器或视网膜色素上皮细胞的向眼睛的治疗性转基因递送中的用途。
背景技术:
2.感光视网膜感光细胞(pr)的永久性退化会导致失明。光感受器死亡可能由主要位于pr或视网膜色素上皮(rpe)细胞中的遗传突变引起,诸如在色素性视网膜炎(rp)中;或由影响光感受器健康的多因素条件引起,例如像在年龄相关性黄斑变性(amd)中。鉴定出了导致许多形式的光感受器退化的基因突变,从而使得能够通过施用治疗性转基因进行干预。现有的基于核酸的靶向pr的疗法仅在对pr的损害最小的早期疾病阶段有效。采用向功能性内层视网膜细胞赋予光敏性的合适光遗传蛋白的基因疗法即使在后期疾病阶段也可能恢复视力。为了使此类方法适用于临床环境,需要具有更好的视网膜渗透以进行基因递送的改进型载体。
3.基于腺相关病毒(aav)的重组载体是在眼睛中进行治疗性基因转移的候选物。目前,典型地视网膜下应用aav载体。然而,已经证明视网膜下应用会导致视网膜厚度和视敏度下降。另外,视网膜下应用仅仅在紧邻注射液主体的细胞中实现基因转移和表达,而有效的施用方法应当沿着视网膜的整个宽度到达细胞。玻璃体内注射到眼睛的果冻状填充物(玻璃体液)中不仅有可能将载体递送到整个视网膜中,而且比视网膜下注射安全得多,并且技术要求低得多。然而,将神经视网膜与玻璃体液分开的内界膜(ilm)在许多aav血清型的天然受体中是丰富的,因此为aav载体提供了巨大的屏障。需要在玻璃体内递送时更好地渗透穿过ilm和其他视网膜屏障的工程化aav载体,以提高眼部aav基因疗法的功效和安全性。
4.基于上文提到的现有技术,本发明的目的是提供用于将基因转移到视网膜中的手段和方法,从而使得能够靶向所有视网膜细胞类型、特别是内层视网膜细胞和光感受器。此目的通过本说明书的独立权利要求的主题来实现。
技术实现要素:
5.本发明的第一方面涉及腺相关病毒(aav)衣壳多肽,其包含在aav血清型2衣壳的位置453或587/588或者另一种血清型的aav衣壳中与其同源的位置处的肽插入物(表3:#1,#2),位置453或587/588分别位于最高和第二高衣壳突起处。该肽插入片段选自以下序列:
6.saseast(cap3;seq id no 10)、dtrphdq(cap5;seq id no11)、ehynstc(cap7;seq id no 12)、pnpnctl(cap9;seq id no 13)、tppsita(cap11;seq id no 14)、cgessyl(cap12;seq id no 15)、prtphta(cap13;seq id no 16)和elcdgfa(cap14;seq id no 17)。
7.该肽的侧翼可以为短段的其他氨基酸(1、2、3、4、5或甚至6个aa)。使所指示序列嵌入衣壳序列内的所指示位置处的侧翼氨基酸的具体实例可以选自(但不限于)ala、leu、
gly、ser和thr。
8.至少对于所测试的细胞类型和组织,此肽插入片段提高了腺相关病毒的感染效力和/或腺相关载体的转导功效,该腺相关病毒和/或腺相关载体将该肽插入片段展示在衣壳表面上所指示位置之上的i-587处。
9.由于其细胞表面暴露,这可能在i-588或i-453处也奏效。不希望受理论束缚,诸位发明人相信至少部分作用是由于在体内与硫酸乙酰肝素蛋白聚糖的结合较低。
10.本发明的第二方面涉及编码根据第一方面的aav衣壳多肽的核酸序列。具体实施例包括将此核酸序列包含在aav cap序列中、特别是aav血清型2衣壳序列中,以用于产生衣壳工程化的aav载体。
11.本发明的第三方面涉及用于治疗影响视网膜或rpe细胞的病症的药剂,其选自该aav衣壳多肽、aav载体和该核酸序列。此方面的替代形式体现为影响光感受器或rpe细胞的病症的治疗方法,这些治疗方法包括向有需要的患者施用根据本发明的药剂。
12.包含本发明的药剂的施用形式是本发明的进一步的方面。
附图说明
13.图1所选择衣壳变体的视网膜层表达特异性。
14.图2玻璃体内注射后整个鼠外层视网膜的全视网膜转基因表达的实例。
15.图3在玻璃体内注射到小鼠眼睛中之后,新型衣壳与aav2(m6)和aav2(7m8)所递送的转基因表达的比较。
具体实施方式
16.术语和定义
17.在本说明书的上下文中缩写aav是指腺相关病毒。
18.在本说明书的上下文中术语aav载体是指由60种aav衣壳蛋白和衣壳化的aav核酸构成的病毒载体。aav载体源自aav病毒体,但是通过从aav基因组中去除rep和cap基因,aav载体被工程化为在辅助病毒存在下无法复制。衣壳化的aav核酸可以包含有待被递送到靶细胞中的转基因。
19.除非另外定义,否则本文使用的所有技术和科学术语都具有与本领域(例如,细胞培养、分子遗传学、核酸化学、杂交技术和生物化学)的普通技术人员通常所理解的相同的含义。使用标准技术进行分子、遗传和生物化学方法(通常参见,sambrook等人,molecular cloning:a laboratory manual[分子克隆:实验室手册],第2版(1989)cold spring harbor laboratory press[冷泉港实验室出版社],cold spring harbor[冷泉港],n.y.[纽约州]和ausubel等人,short protocols in molecular biology[精编分子生物学实验指南](1999)第4版,john wiley&sons,inc.[约翰
·
威利父子公司])以及化学方法。
[0020]
在本说明书的上下文中术语aav衣壳是指由诸位发明人产生并且在下文列出的工程化衣壳(cap)基因所编码的多肽。可以使用本文披露的aav衣壳来组装用于基因疗法的重组腺相关病毒载体。
[0021]
在本说明书的上下文中对aav衣壳中的氨基酸位置的提及是指具有seq id no.1的序列的腺相关病毒2衣壳蛋白vp1的衣壳氨基酸序列(表2)(对应核酸序列的genbank登录
号为j01901.1)。其他同源腺相关病毒血清型的对应氨基酸位置示于表3中。
[0022]
在本说明书的上下文中术语同源是指源自不同aav血清型的核酸或氨基酸序列。不同腺相关病毒血清型的对应位置示于表3中。
[0023]
在本说明书的上下文中术语转基因是指已经从一种生物体转移到另一种生物体中的基因或遗传物质。在本发明的上下文中,该术语还可以指代将基因序列的天然或生理上完整的变体转移到缺失它的患者组织中。它可以进一步指代转移天然编码序列,其表达由靶向组织中缺失或沉默的启动子驱动。
[0024]
在本说明书的上下文中术语重组是指这样的核酸,其是克隆、限制和/或连接中的一个或几个步骤的产物并且不同于天然存在的核酸。重组病毒颗粒包含重组核酸。
[0025]
在本说明书的上下文中术语玻璃体内施用是指药剂(例如病毒载体)的施用途径,其中药剂被递送到眼睛的玻璃体中。玻璃体内施用是将药物直接放入眼睛后部称为玻璃体腔的空间中的程序,该空间充满称为玻璃体液凝胶的果冻状液体。
[0026]
在本说明书的上下文中术语视网膜下施用是指将药剂、特别是在本说明书的上下文中的病毒载体施用到视网膜色素上皮(rpe)细胞与光感受器之间的空间中的途径。
[0027]
在本说明书的上下文中术语多肽是指由50个或更多个形成直链的氨基酸组成的分子,其中氨基酸通过肽键连接。多肽的氨基酸序列可以代表整个(如在生理上发现的)蛋白质或其片段的氨基酸序列。术语“多肽”和“蛋白质”在本文中可互换地使用,并且包括蛋白质及其片段。多肽在本文中披露为氨基酸残基序列。
[0028]
从氨基到羧基末端给出氨基酸残基序列。序列位置的大写字母是指单字母密码的l-氨基酸(stryer,biochemistry[生物化学],第3版第21页)。氨基酸序列位置的小写字母是指对应的d-或(2r)-氨基酸。以从氨基到羧基末端的方向从左到右书写序列。根据标准命名法,氨基酸残基序列由三字母或单字母密码命名,如下所指示:丙氨酸(ala,a)、精氨酸(arg,r)、天冬酰胺(asn,n)、天冬氨酸(asp,d)、半胱氨酸(cys,c)、谷氨酰胺(gln,q)、谷氨酸(glu,e)、甘氨酸(gly,g)、组氨酸(his,h)、异亮氨酸(ile,i)、亮氨酸(leu,l)、赖氨酸(lys,k)、甲硫氨酸(met,m)、苯丙氨酸(phe,f)、脯氨酸(pro,p)、丝氨酸(ser,s)、苏氨酸(thr,t)、色氨酸(trp,w)、酪氨酸(tyr,y)和缬氨酸(val,v)。
[0029]
术语变体是指不同于参考多肽但保留基本特性的多肽。多肽的典型变体在其一级氨基酸序列方面与用作参考的另一种多肽不同。通常,差异是有限的,因此参考多肽和变体的序列总体上非常类似,并且在许多区域中是相同的。变体和参考多肽在氨基酸序列方面的不同可以在于一种或多种修饰(例如,取代、添加和/或缺失)。多肽的变体可以是天然存在的,诸如等位基因变体;或者它可以是已知不是天然存在的变体。
[0030]
在本说明书的上下文中术语生物活性是指本文所附多肽的某些变体所展示的具体可测量的量。例如,在病毒载体促进基因转移到视网膜细胞中的能力的背景下,衣壳变体的生物活性可以通过在标准测定中测量转基因(例如萤火虫萤光素酶)在培养细胞中或在体内模式生物体中的表达来测定。
[0031]
在本说明书的上下文中,术语序列同一性和序列同一性百分比是指通过比较两个比对序列所确定的值。用于比较的序列比对方法是本领域熟知的。用于比较的序列比对可以通过smith和waterman,adv.appl.math.[应用数学进展]2:482(1981)的局部同源算法、通过needleman和wunsch,j.mol.biol.[分子生物学杂志]48:443(1970)的全局比对算法、
通过pearson和lipman,proc.nat.acad.sci.[国家科学院院刊]85:2444(1988)的相似性检索方法或通过这些算法的计算机化实现(包括但不限于:clustal、gap、bestfit、blast、fasta和tfasta)来进行。用于执行blast分析的软件是公开可用的,例如通过国家生物技术信息中心(http://blast.ncbi.nlm.nih.gov/)。
[0032]
比较氨基酸序列的一个实例是使用以下默认设置的blastp算法:期望阈值:10;字长:3;查询范围内的最大匹配:0;矩阵:blosum62;空位罚分:存在11,延伸1;成分调整:条件性成分得分矩阵调整。比较核酸序列的一个这样的实例是使用以下默认设置的blastn算法:期望阈值:10;字长:28;查询范围内的最大匹配:0;匹配/错配得分:1.-2;空位罚分:线性。除非另外说明,否则本文提供的序列同一性值是指使用blast程序套件(altschul等人,j.mol.biol.[分子生物学杂志]215:403-410(1990))使用分别用于蛋白质和核酸比较的上文鉴定的默认参数获得的值。
[0033]
在本说明书的上下文中,术语氨基酸接头是指用于连接两种多肽以产生单链多肽的可变长度的寡肽。可用于实施本文所指明的发明的接头的示例性实施例是由1至6个氨基酸组成的寡肽链。氨基酸接头的非限制性实例是aaa或aa,如下文所例示。
[0034]
aav是可感染人和其他灵长类动物物种的小的非致病性病毒。
[0035]
aav2感染通过与细胞表面受体硫酸乙酰肝素蛋白聚糖(hspg)对接而开始。其与聚糖的低亲和力结合诱导衣壳的可逆结构重排,这促进与辅助受体αvβ5或α5β1整合素的结合,从而诱导形成网格蛋白包被的凹坑。网格蛋白包被的凹坑经由胞吞作用被内化,并且病毒颗粒被转运到细胞核中。由于内体区室的酸化,ph下降,这是内体囊泡成熟的特征。酸化触发的构象变化发生在衣壳中,并且病毒通过脂解孔形成而从晚期内体中逃逸。
[0036]
在野生型aav中,基因组由4.7千碱基长的单链dna(ssdna)(正义的或反义的)构成。基因组包含三个开放阅读框(orf),其侧翼为反向末端重复序列(itr)。itr是在aav基因组的5'-端和3'-端的自身互补的富含cg的t形发夹,并且是重组载体基因组中唯一必需的病毒组分。itr包括末端解离位点(trs)和rep结合元件(rbe),它们促进病毒基因组的复制和衣壳化。orf编码基因rep、cap、aap。aav生命周期需要四种由rep编码的多功能非结构rep蛋白。cap编码衣壳蛋白vp1、vp2和vp3,它们一起相互作用以形成二十面体对称的衣壳;以及组装激活蛋白(aap),它是稳定和将新产生的vp蛋白从细胞质转运到细胞核中所需的。所有三种vp都是从一种mrna翻译而来,并且不同地剪接。最大的90kda的vp1是未剪接的转录物,72kda的vp2是从非常规acg起始密码子翻译而来,而最小的60kda的vp3是从aug密码子翻译而来。所有三种vp具有重叠的c-末端。
[0037]
vp3构成衣壳的90%,并且vp1和vp2与vp3具有共同的c-末端氨基酸序列,但是具有埋在衣壳内的n-末端延伸。vp1特有的n-末端序列含有感染所需的磷脂酶a2(pla2)活性以及核定位信号。三种vp单体组装成二倍、三倍和五倍对称轴,以产生aav衣壳的60个亚基。本发明的第一方面涉及腺相关病毒(aav)衣壳多肽,其包含在aav血清型2衣壳的位置453或587处的顶点或尖突起处的肽插入片段。肽插入片段选自以下序列:
[0038]
saseast(cap3;seq id no 10)、dtrphdq(cap5;seq id no11)、ehynstc(cap7;seq id no 12)、pnpnctl(cap9;seq id no 13)、tppsita(cap11;seq id no 14)、cgessyl(cap12;seq id no 15)、prtphta(cap13;seq id no 16)和elcdgfa(cap14;seq id no 17),
[0039]
特别地,插入片段选自saseast(cap3;seq id no 10)、tppsita(cap11;seq id no 14)、prtphta(cap13;seq id no 16)和elcdgfa(cap14;seq id no 17)。
[0040]
如上文所表达的肽插入片段可以包含在由上文序列之一组成的7-13聚体中,侧翼为0-6个插入在上一段中给出的插入片段序列的n-末端和/或c-末端的选自但不限于ala、leu、gly、ser和thr的接头氨基酸(6是n末端和c末端侧翼氨基酸总数的最大数目)。
[0041]
针对本文给出的aav2序列给出的任何位置都是关于可从genbank条目号j01901.1(腺相关病毒2,完整基因组)获得的参考序列。对应的氨基酸序列在表2中作为seq id no 1给出。
[0042]
尖突起(顶点)代表衣壳最暴露的区域。最高顶点位于aav2衣壳上的氨基酸位置453处,并且第二高顶点位于位置587处。这些顶点在不干扰衣壳组装的情况下接受肽插入物,并且为靶向不允许的细胞提供机会。同样地,突起代表aav宿主相互作用、受体结合和免疫原性的关键位点。值得注意的是,在某些实施例中,如果插入片段插入在位置453处,则r585和588发生突变以获得最佳效率。
[0043]
在提及引入肽插入片段的某个顶点或尖突起位置时,应理解肽插入片段可以直接在此位置之后(在c-末端方向上)或在c-末端方向上更远的一至六个氨基酸之后。例如,定义为在位置587处的肽插入片段(定义为在插入前位置587与588之间的插入物)可以在位置588或589(在c-末端方向上更远的一个或两个氨基酸)或590(在c-末端方向上更远的三个氨基酸)处开始。所有这些插入位置都允许不干扰总体衣壳结构而是可能提高转导功效的肽插入片段。
[0044]
此肽插入片段在玻璃体内递送后提高了病毒的视网膜渗透和转导功效。
[0045]
在某些实施例中,肽插入片段选自saseast(cap3;seq id no10)、dtrphdq(cap5;seq id no 11)、ehynstc(cap7;seq id no 12)、pnpnctl(cap9;seq id no 13)、tppsita(cap11;seq id no 14)、cgessyl(cap12;seq id no 15)、prtphta(cap13;seq id no 16)和elcdgfa(cap14;seq id no 17)。
[0046]
在某些实施例中,肽插入片段选自saseast(cap3;seq id no10)、tppsita(cap11;seq id no 14)、prtphta(cap13;seq id no 16)和elcdgfa(cap14;seq id no 17)。
[0047]
肽插入片段可以按原样存在于aav衣壳多肽中,或者在1、2、3或4个侧翼间隔氨基酸的背景下,存在于氨基末端和/或羧基末端。合适的间隔aa包括但不限于丙氨酸、亮氨酸、甘氨酸、苏氨酸和丝氨酸。
[0048]
在诸位发明人的实例中,由于所使用的克隆策略,aav2 vp1的n
587
与r
588
之间的插入物是r
585
g n a a a x
1 x
2 x
3 x
4 x
5 x
6 x
7-a a r
588 q a a形式的12聚体,而x
1-x7代表所插入的七聚体寡聚物,并且a代表侧翼接头。应理解,这仅是一个实例,并且本发明不限于恰好在恰好具有这些侧翼序列的此位置处的插入片段。
[0049]
在涉及aav2中指定为587的插入位置的实施例中,寡聚物总是插入在aav2的vp1上的位置587与588之间。它是插入物,因此vp1aav2编号不会改变,并且后续位置在取代等方面在“无插入片段”的情况下进行计数。所插入的肽含有如本文所指明的七聚体,但是也可能更短以发挥功能或更长。在实例中表示的实施例中,诸位发明人插入了呈aaa-x1x2x3x4x5x7-aa形式的12聚体,而接头氨基酸可以自由地选自ala、leu、gly、ser、thr。n-末端和c-末端接头可以具有0-5个aa的长度。在某些实施例中,插入肽可以具有5-13个氨基
酸的长度,包含0-6个接头氨基酸。
[0050]
在某些实施例中,肽插入物(在本说明书中的其他地方称为“寡聚物(oligo)”)可以将1-5个aa进一步移向c-末端。对于aav2,插入位点也可以在氨基酸453处(例如,紧接其c-末端)。
[0051]
根据本发明的此第一方面的替代方案,上文所列举的任何肽序列都插入seq id no 001的位置453而不是位置587处。同样,插入的位置可以变化,并且肽序列的侧翼可以为如上文所讨论的插入片段,并且可以由5-13个aa组成。除了位置453处的肽插入物外,还可以将r587a和r588a突变引入aav2的vp1中(boucas j等人j gene med.[基因医学杂志]2009年12月;11(12):1103-13.doi:10.1002/jgm.1392)。
[0052]
根据本发明的此方面的另一个替代方案,可以选择与aav血清型2中的位置587/588或453同源的位置(表3:#1,#2),以构建另一种血清型的具有上文所列举的所插入的肽序列之一的aav。
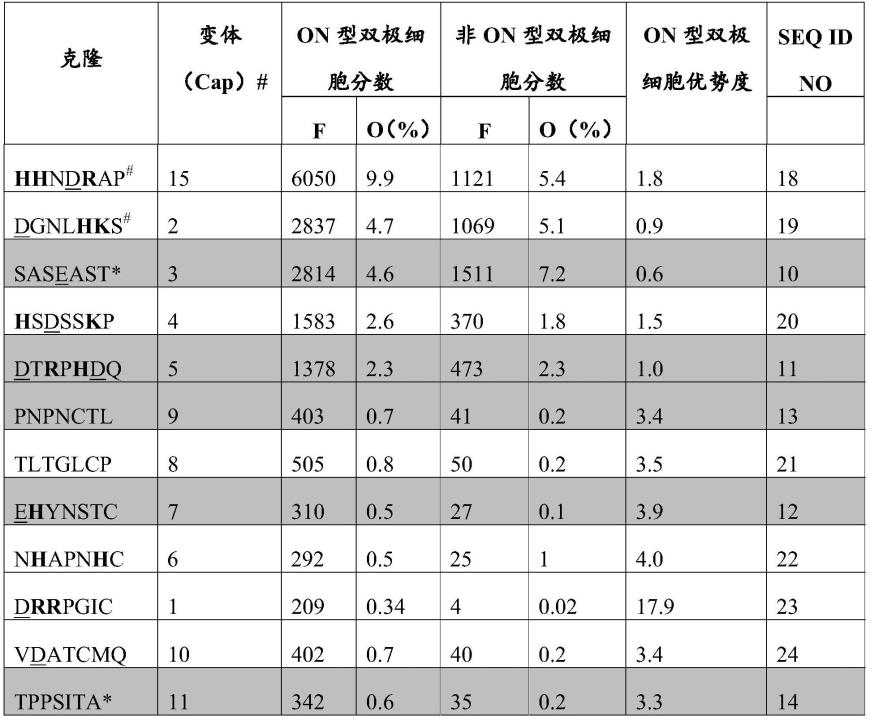
[0053]
在某些实施例中,aav衣壳蛋白的特征在于酪氨酸残基的一个或几个酪氨酸至苯丙氨酸取代,其中酪氨酸残基出现在野生型衣壳序列中的位置252、272、444、500、700、704和730处。
[0054]
在酪氨酸被修饰的衣壳的背景下,通过进化方法选择本文中的某些衣壳变体。
[0055]
在某些实施例中,aav衣壳蛋白的特征在于位置252、272、444、500、704和730处的一个或几个酪氨酸至苯丙氨酸取代。
[0056]
在某些甚至更具体的实施例中,aav衣壳蛋白的特征在于在位置252、272、444、500、700和730中的全部处的几个酪氨酸至苯丙氨酸取代。
[0057]
根据本发明的此方面的另一个替代方案,可以选择与aav血清型2中的位置252、272、444、500、700、704和730中的任一个同源的酪氨酸位置以取代为苯丙氨酸(表3:#3-#9),以构建另一种血清型的具有酪氨酸至苯丙氨酸取代的aav。此外,其他酪氨酸位置可以取代为苯丙氨酸。
[0058]
在某些实施例中,aav衣壳蛋白的特征在于一个或几个苏氨酸至缬氨酸取代、特别是t491v。
[0059]
根据本发明的此方面的另一个替代方案,可以选择与aav血清型2中的位置491同源的苏氨酸位置以取代为缬氨酸(表3:#10),以构建另一种血清型的具有苏氨酸至缬氨酸取代的aav。
[0060]
在某些实施例中,将上文提到的酪氨酸至苯丙氨酸取代与苏氨酸至缬氨酸取代、特别是t491v组合。
[0061]
y-f和t-v突变减少泛素化,因而减少病毒颗粒一旦被细胞内化后的蛋白酶体降解,从而提高转导功效。
[0062]
在某些实施例中,腺相关病毒衣壳多肽包含选自seq id no 2-seq id no 9的氨基酸序列,或与其具有至少85%同一性并且具有选自seq id no 2-seq id no 9的序列的至少90%生物活性的序列。
[0063]
生物活性测定:
[0064]
无论在本文的什么地方提及“具有(参考)序列的至少一定百分比的生物活性”的多肽时,此生物活性都可以如下测量:
[0065]
使用磷酸钙沉淀法用编码测试或参考多肽的质粒(该质粒具有辅助腺病毒基因)和作为报告物的选自sccmv-mcitrine、scef1a-mcitrine、sccmv-egfp、scef1a-egfp的转基因质粒共转染hek293细胞。通过以下方式浓缩含有转基因的载体:利用碘克沙醇梯度(阿克西斯-希尔德公司(axis-shield),奥斯陆)进行密度纯化,随后通过例如amicon过滤(密理博公司(millipore))对40%碘克沙醇级分进行缓冲液交换。通过实时pcr相对于标准载体针对抗dna酶载体基因组对aav级分进行滴定。将3μl病毒载体玻璃体内地注射到麻醉的(100mg/kg氯胺酮和10mg/kg甲苯噻嗪)野生型c57bl/6j小鼠中。对于注射,病毒的滴度与e11 gc/ml匹配,并且每只眼睛注射3ul,即3xe8 vg。注射后3周取出小鼠视网膜进行免疫组织化学分析。将小鼠眼睛视网膜外植块(转导后7天)在室温(rt)下用在pbs中的4%(wt/vol)多聚甲醛固定40min,在分等级的蔗糖溶液(在pbs中的10%、20%和30%蔗糖)中在4℃下连续低温保护三个晚上,包埋在具有o.c.t.化合物的低温模具(樱花检验仪器株式会社(sakura finetek))中,并且在液氮冷却的2-甲基丁烷中冷冻。在低温恒温器上切割10μm厚的纵向切片,将其安装在superfrost载玻片(门采尔公司(menzel))上,并且用合适的一抗和二抗进行免疫组织化学标记,以检测egfp、yfp、mcitrine或turbo635荧光以及特定的视网膜细胞类型。典型地通过荧光定量靶细胞类型中的报告物表达,并且将其与其余部分的组织中的报告物的量进行比较,以评估工程化病毒衣壳的生物活性。工程化病毒衣壳的生物活性的另一种量度是所转导靶细胞的百分比和组织表达面积。为免生疑问,在测量生物活性时,采用定量靶细胞类型中的报告物表达并且将其与其余部分的组织中的报告物的量进行比较的测定。
[0066]
本发明的第二方面涉及编码根据第一方面的aav衣壳多肽的核酸序列。
[0067]
在具体实施例中,根据本发明的aav序列不含有整合到宿主基因组中所需的rep元件。非整合病毒对于在人中应用更安全。在某些实施例中,核酸序列是根据自身互补的aav载体基因组概念设计的。在所有情况下,都使用单链dna序列。
[0068]
在某些实施例中,核酸序列包含具有或不具有调控序列的转基因。
[0069]
在某些实施例中,转基因编码正确蛋白质(即rpe65)、sirna或shrna(设计用于靶向mrna区域以降解显性致病基因的野生型和突变型rna)或用于基因编辑的crispr/cas-grna盒的序列。
[0070]
在某些实施例中,转基因编码光敏蛋白,诸如无脊椎动物或脊椎动物视蛋白或其变体。
[0071]
在某些实施例中,转基因编码光敏感通道蛋白-2或其变体。
[0072]
在某些实施例中,转基因编码微生物光门控抑制性离子泵,例如像嗜盐菌视紫红质(enphr)或古细菌视紫红质(archt)。
[0073]
在某些实施例中,转基因编码gpcr视蛋白,例如像人视紫红质、视黑蛋白或视锥视蛋白。
[0074]
在某些实施例中,转基因编码光可切换配体的受体蛋白。
[0075]
在某些实施例中,转基因受在哺乳动物细胞中可操作的启动子序列的控制。在某些实施例中,启动子序列在人视网膜细胞中是可操作的。
[0076]
在某些实施例中,启动子是遍在启动子。在某些实施例中,启动子是细胞特异性启动子。
[0077]
在某些实施例中,启动子是cmv立即早期启动子。在某些实施例中,启动子是人ef1a启动子。在某些实施例中,启动子是光感受器特异性启动子。
[0078]
本发明的第三方面涉及选自根据第一方面的aav衣壳多肽、包含根据第一方面的衣壳多肽的aav载体和根据第二方面的核酸序列的药剂,用于在医学中使用/用于作为药物使用。
[0079]
在某些实施例中,使用aav载体来转导眼睛细胞。
[0080]
眼睛由三层组成,这三层由各种解剖结构构成。纤维膜是最外层,并且由角膜和巩膜构成。血管膜或葡萄膜是中间层,并且由脉络膜、睫状体、色素上皮和虹膜组成。视网膜是最内层,它从脉络膜(在后)和视网膜血管(在前)的血管中获得氧合。
[0081]
角膜与晶状体之间的空间充满房水,并且晶状体后面的整个后腔都充满玻璃体,一种果冻状物质。玻璃体由水、胶原蛋白、原纤维、透明质酸和离子构成。视网膜中未被神经元或血管占据的空隙空间被跨越所有视网膜细胞层的m
ü
ller胶质细胞的端突填充。
[0082]
视网膜的外界膜(olm)由m
ü
ller细胞(mc)与感光细胞内段之间的连接形成,并且充当视网膜下间隙之间的代谢屏障,从而限制大分子的通过。视网膜的内界膜(ilm)通过mc端脚(end-feet)与基膜之间的横向连接形成,并且充当玻璃体液与神经视网膜之间的扩散屏障。
[0083]
视网膜由黄斑、视盘、中央凹和周边视网膜构成。感光细胞是在视网膜中发现的专门类型的神经上皮细胞。已知三种类型的感光细胞:视杆细胞、视锥细胞和感光性视网膜节细胞。视杆细胞分布在视网膜的周边区域;而称为黄斑的中央色素区域富含视锥感光细胞。视网膜色素上皮(rpe)提供营养,并且维持感光细胞的健康。感光细胞的细胞核构成外核层(onl),而双极细胞、无长突细胞和水平细胞的细胞核位于内核层(inl)。
[0084]
在某些实施例中,选自根据第一方面的aav衣壳多肽、包含根据第一方面的衣壳多肽的aav载体和根据第二方面的核酸序列的药剂用于在治疗影响以下细胞的病症中使用:
[0085]
a.视网膜或视网膜色素上皮细胞,和/或
[0086]
b.视网膜的光感受器、双极细胞、无长突细胞或节细胞。
[0087]
在某些实施例中,药剂用于在治疗以下疾病中使用:青光眼、色素性视网膜炎、黄斑变性、视网膜劈裂症、莱伯先天性黑朦、糖尿病视网膜病变、全色盲或色盲、黑色素瘤相关性视网膜病变、先天性静止性夜盲、视锥视杆营养不良、晚期年龄相关性黄斑变性、黄斑病变、早发性严重视网膜营养不良、全色盲、眼白化病、眼皮肤白化病、眼底黄色斑点症、无脉络膜、脊髓小脑性共济失调7型(scat)、影响角膜的溶酶体贮积症(诸如粘多糖贮积症(mps)iv和mps vii)、视网膜母细胞瘤、眼部黑色素瘤、高血压性视网膜病变。
[0088]
在某些实施例中,本发明的药剂可以用于治疗或预防影响内耳的疾病。
[0089]
在某些实施例中,通过以下方式施用药剂:
[0090]
a.玻璃体内施用、特别是玻璃体内注射,或者
[0091]
b.视网膜下施用。
[0092]
在某些实施例中,将药剂递送至后段、前段、巩膜、脉络膜、结膜、虹膜、晶状体或角膜。
[0093]
类似地,在本发明范围内的是用于治疗有需要的患者的选自包括色素性视网膜炎在内的视杆视锥营养不良、包括黄斑变性在内的视锥视杆营养不良和先天性静止性夜盲
(csbn1)的病症的方法,该方法包括向该患者施用包含根据上文描述的aav衣壳和/或核酸序列的病毒载体。
[0094]
类似地,提供了用于预防或治疗选自视锥视杆营养不良、视杆视锥营养不良和先天性静止性夜盲的病症的剂型,其包含根据本发明的上文方面之一的aav衣壳和/或核酸序列。
[0095]
本文披露的药剂和方法在对pr的损害小的早期疾病阶段确实也提供了显著优势,因为由本发明所促进的载体比任何现有的替代物都更有效。
[0096]
无论在本文的什么地方将单独的可分离特征(例如像aav血清型蛋白或衣壳肽插入片段序列或医学适应症)的替代方案作为“实施例”表达,都应理解此类替代方案可以自由组合以形成本文披露的本发明的离散实施例。
[0097]
通过以下实例和附图进一步说明本发明,从实例和附图中可以得出进一步的实施例和优势。这些实例意在说明本发明,而不是意在限制其范围。
[0098]
附图描述
[0099]
图1所选择衣壳变体与选择骨架aav2(y252、272、444、500、700、730f)(在下面称为aav2(m6))和现有技术的合成衣壳aav2(7m8)相比的细胞表达特异性。所有aav都已经被包装为自身互补的(scaav),从而在两种遍在启动子hef1a和cmv下表达egfp或mcitrine,以消除一种启动子对某种视网膜细胞类型的可能偏倚。与aav2(m6)和aav2(7m8)相比,玻璃体内注射后新型变体在光感受器(onl)中的表达显著增强。玻璃体内地注射的2.5μl的滴度与1e 11vg/ml匹配(cap14除外,为3e 10vg/ml)。计数了4个视网膜,平均值
±
标准差。inl,内核层;gcl,节细胞层;dapi,被核染料dapi染色的所有细胞体的数目。
[0100]
图2在降低100倍的功能滴度下如所指示的新型衣壳变体与现有技术的aav2(7m8)相比在整个onl中的全视网膜强表达。所有注射的滴度都是匹配的,并且将7.4e 7vg递送到c57bl/6小鼠的玻璃体中。a.在用sccap5-cmv-egfp转导后,聚焦于小鼠视网膜全样载片的onl的低放大率图片。b-c.纵向视网膜冷冻切片。根据显微照片,与aav2(7m8)相比,新型衣壳在低滴度下的效力是显而易见的。
[0101]
图3将2.5μl scaav玻璃体内注射到c57bl/6小鼠眼睛中后的转导比较(如例如van wyk等人,plos biol[公共科学图书馆
·
生物学],2015.13(5):第e1002143页中所述)。在人ef1a启动子控制下的mcitrine表达,滴度与1e 11vg/ml匹配。根据纵向且经免疫标记的视网膜冷冻切片的显微照片,显而易见的是scaav2(cap3)和scaav2(cap5)更好地渗透通过ilm和视网膜层,并且在与现有技术的aav(7m8)或粗文库骨架aav2(m6)相比时,在onl的光感受器中显示出无与伦比的表达功效。
[0102]
实例
[0103]
实例1:肽展示文库的产生
[0104]
aav衣壳是具有60个亚基的二十面体。在aav衣壳的当衣壳亚基组装时形成的3倍轴处,形成尖突起(顶点),它们代表衣壳最暴露的区域。在aav2衣壳上,最高顶点位于氨基酸(aa)位置453处,并且第二高顶点位于aa位置587/588处(aav2-vp1编号)。这些顶点在不干扰病毒衣壳组装的情况下接受肽插入物,并且代表宿主相互作用、受体结合和免疫原性的关键位点。为了产生肽展示文库,诸位发明人在aav2的vp1的开放阅读框的n
587
与r
588
之间插入随机七肽,其含有六个另外的酪氨酸至苯丙氨酸突变(y252、272、444、500、700、730f),
称为aav2(m6)(表2)。先前已证明y-f和t-v突变(b
ü
ning和srivastava,mol ther methods clin dev.[分子疗法-方法与临床开发]2019年1月26日;12:248-265.doi:10.1016/j.omtm.2019.01.008)会减少泛素化,因而减少病毒颗粒一旦被细胞内化后的蛋白酶体降解(petrs-silva等人,mol ther[分子疗法],2011.19:第293-301页.),从而提高转导功效,并且可能有利于变体处于以非常低的计数表示的初始轮次的体内进化中的选择过程。可以在其他衣壳血清型的gh环(环iv)中的同源位点处进行随机插入,如表3所示。诸位发明人如何开发文库的方法改编自perabo和同事(perabo等人,mol ther[分子疗法],2003.8:第151-157页)。在aav2(m6)基因组的氨基酸587与588之间引入两个独特的限制性位点asci和noti(表2)。所引入的形成asci和noti位点的dna片段编码侧翼为丙氨酸接头的终止密码子(aaa终止密码子aa)。接下来,合成作为以下序列的具有nnb密码子的单链随机7聚体库:
[0105]
5'-ttggcgcgccgcvnnvnnvnnvnnvnnvnnvnnggcggc cgcttttttccttga-3'(seq id no 25)
[0106]
(欧陆基因组学公司(eurofins genomics)),并且使用反义引物5'-ctcaaggaaaaaagc-3'(seq id no 26)通过第二链合成将其转化为dsdna片段库。为了产生7聚体展示文库,将dsdna随机寡核苷酸克隆到经修饰的aav(m6)基因组的asci-noti位点中,从而替换终止密码子。产生病毒文库,使得每个病毒基因组都被包装或衣壳化在该基因组所编码的衣壳蛋白变体内(基因型-表型结合)。另外,所有衣壳都是以在60个亚基中的每一个中仅存在一种肽的方式产生的。以此方式,通过选择鉴定出的功能改进可以与编码病毒衣壳内所含的这种改进功能的基因组序列相联系。
[0107]
实例2:肽变体的选择
[0108]
在由我们的实验室产生的c57bl/6_opto-mglur6小鼠(van wyk等人,plos biol[公共科学图书馆
·
生物学],2015.13(5):第e1002143页)内对此文库进行正向选择,这些小鼠在视网膜on型双极细胞上表达红色荧光标记fp635。基于以下基本原理选择此小鼠品系:on型双极细胞位于视网膜深处,并且已知不允许aav转导,使得选择将有利于具有有利特性以转导on型双极细胞的渗透良好的aav和变体。简而言之,为5-8只4-6周龄的转基因小鼠玻璃体内地注射2.5μl经碘克沙醇纯化的文库(其基因组滴度为大约5x 10
11
个病毒基因组(vg)/ml),注射到双眼中,如(van wyk等人,plos biol[公共科学图书馆
·
生物学],2015.13(5):第e1002143页;van wyk等人,front neurosci[神经科学前沿],2017.11:第161页)中所述。
[0109]
10天后,摘除眼睛的细胞核,并且使用光木瓜蛋白酶处理分离出视网膜,然后对on型双极细胞群进行荧光激活细胞分选(facs)。然后从dna提取物中对成功的病毒体进行pcr扩增,并且对其进行进一步克隆和重新包装以进行后续轮次的注射。
[0110]
在将7聚体寡聚物克隆到aav2(m6)基因组中的每个步骤中,运行四个平行的连接,并且对每个库的12个克隆铺板并测序,以估计每个体内选择步骤后文库的收敛。此体内定向进化过程通过应用类似于自然进化过程的正向选择产生on型双极细胞允许的aav变体。
[0111]
从体内选择的文库中,通过下一代测序鉴定出81'738种变体的衣壳基因。结合高总体转导功效(在ngs中的积累)、对on型双极细胞靶向的偏好和在选择轮次的早期出现或在氨基酸序列方面看起来有希望(即含有带负电荷的残基或脯氨酸残基)的基序,选择来自前67名的15个七肽序列进行功能评价和表征,如表1中所概述。所有文库变体的关键特征是
通过插入分离aav2的硫酸乙酰肝素蛋白聚糖结合基序的正电荷(r
585
和r
588
)。然而,肽序列与接头序列的两个丙氨酸一起可以重新设置与硫酸乙酰肝素蛋白聚糖结合的能力(uhrig
…bü
ning,gene therapy[基因疗法])。在7聚体插入片段序列中,在特定位置处存在适度的偏好,例如位置1或2和/或位置5或6处的带电荷的氨基酸以及位置7处的极性氨基酸(诸如cys或ala)(表1)。
[0112]
实例3:新型衣壳变体的评价
[0113]
克隆表1的所有15种所选择肽的aav2(m6)~7聚体~的重组形式。诸位发明人用sccmv-egfp转基因(cmv:巨细胞病毒启动子)包装。除两种变体(cap8和cap10)外,所有衣壳都包装良好,滴度在10
11-10
12
vg/ml之间。在玻璃体内注射到成年小鼠中三周后,主要在光感受器中观察到cap3、cap5、cap7、cap9、cap 11、cap 13和cap 14的稳健表达(表1,图1-图3),而且在内层视网膜细胞(诸如双极细胞、无长突细胞和m
ü
ller细胞)中观察到一些表达,并且在视网膜节细胞中观察到非常少的表达。在与也是进化的aav2变体(dalkara等人,sci transl med[科学转化医学],2013.5:第189ra76页.)并且目前被认为是玻璃体内注射的现有技术的aav2(7m8)的性能的直接比较中,诸位发明人的新型变体显著更好地渗透通过视网膜,并且转导显著更多的光感受器和显著更少的节细胞(图1,图1-图3)。通过以下方式证实上文的结果:也用schef1a-egfp-mcitrine转基因(hef1a:人ef1a启动子,“强”天然哺乳动物遍在启动子)包装8种预选变体(cap3、cap5、cap7、cap9、cap 11、cap 13和cap 14;表1),并且以1x10
11 vg/m的匹配滴度向成年小鼠玻璃体内地注射。
[0114]
同样地,新变体所转导的光感受器的数目显著大于滴度匹配的对照aav2(m6)和aav(7m8)。因此,与aav2(m6)和aav2(7m8)相比,所选择变体似乎具有优越的内界膜和视网膜渗透特性,并且在转导外核层(onl)中的光感受器方面显著更有效(图1)。
[0115]
用sccmv-egfp和schef1a-egfp包装并且以3e 10vg/ml注射的cap14的性能与以相同低剂量注射的aav2(7m8)相比显著更好(图1、图2),并且甚至与以典型地在小鼠中使用的1e 12vg/ml的最小剂量注射的aav2(7m8)更好。鉴于在患者中使用较低剂量以减少对毒性和免疫反应的担忧,这是有希望的。
[0116]
表1:来自定向进化的on型双极细胞分离变体的下一代测序揭示了病毒文库的高收敛度。7聚体插入片段的侧翼为丙氨酸间隔物,呈~n
587
aaasaseastaa r
588
~(seq id no 28)的形式,在这里以变体#3为例示。
[0117]
on型双极细胞优势度:=出现[on型双极细胞/非on双极细胞]
[0118][0119][0120]
f=频率;o=出现
[0121]
#
出现在第2轮选择中
[0122]
*从体内筛选中选出的顶级候选物
[0123]
具有插入片段nr.8和nr.10的衣壳未包装(灰色字体)
[0124]
带正电荷
[0125]
带负电荷
[0126]
所有15个所选择克隆都排在从c57bl/6_opto-mglur6小鼠(van wyk等人,plos biol[公共科学图书馆
·
生物学],2015.13(5):第e1002143页)的分离的on型双极细胞部分测序的60'884种肽的前67名中。
[0127]
表2:具有不同插入物的aav2衣壳蛋白vp1的氨基酸序列。
[0128]
vp3(灰色序列)与vp1重叠,酪氨酸至苯丙氨酸(y-f)突变以深灰色突出显示并加下划线,氨基酸编号参考整个vp1序列。插入随机7聚体的g
453
处的最高顶点和n587处的第二高顶点加下划线并用白色衬底指示。插入物是斜体且加框的,包括丙氨酸接头。
[0129]
si:肽插入序列的seq id no;fs:完整衣壳序列的seq id no
[0130]
[0131]
[0132]
[0133]
[0134]
[0135][0136]
表3:不同aav血清型的vp3衣壳蛋白上相应的潜在寡聚体插入位点以及y-f和t-v点突变位点的同源位置。
[0137]
[0138][0139]
加下划线的位置展示出相对于aav2不同的氨基酸。
[0140]
表4:腺相关病毒2(y252、272、444、500、700、730f)(称为aav2(m6))的衣壳基因序列(vp1)
[0141]
[0142]
[0143][0144]
基于aav 2完整基因组序列,genbank登录号j01901.1
[0145]
文库的cap基因由vp3(灰色序列)组成,酪氨酸至苯丙氨酸(y-f)突变加下划线,碱基对编号参考整个vp1序列。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
