一种水曲柳u6基因启动子profmu6.7及其克隆与应用
技术领域
1.本发明属生物技术领域,特别是植物转基因技术领域,具体涉及一种水曲柳rna聚合酶ⅲ型启动子,更具体涉及一种水曲柳u6基因启动子profmu6.7,并进一步公开其克隆方法和应用。
背景技术:
2.水曲柳是木犀科白蜡属落叶大乔木,被列为国家二级保护渐危种,木材总体强度高、抗震性好、木质优良,常用于建筑、家具等,具有重要的经济价值。自1983年首次获得烟草转基因植株后,木本植物的遗传转化也得到重视,遗传转化作为林木基因工程的一个重要组成部分,在木本植物抗病、抗虫、改良性状以及基因工程育种中发挥重要作用。在生命科学领域,突变体对于基因功能的研究具有至关重要的作用。然而,林木世代周期长、遗传杂合度高、基因组倍性复杂,传统的随机诱变方法往往需要构建大群体的突变体库,并进行大规模筛选才能获得目的基因功能丧失的突变体,这一过程需要大量的人力和物力。相比之下,基因组定点编辑技术具有巨大优势,能够直接在基因组特定位置引入突变。
3.crispr/cas9系统是目前公认的最具有发展潜力的基因编辑技术,cas9酶在sgrna引导下定向切割目标位点,对目标基因进行精准编辑。相较于锌指核酸酶(zinc finger nucleases,zfns)、类转录激活因子效应物核酸酶(transcription activator-like effector nucleases,talens)技术,crispr/cas9系统设计流程简单、操作方便、基因编辑效率高,一经开发被迅速广泛应用于动物、植物、微生物的研究中。自2013年crispr/cas9首次用于模式植物拟南芥和烟草以来,该基因编辑系统已运用于包括水稻(nekrasov等,nat biotechnol,2013,31(8):691
–
693.)、小麦(wang等,nature biotechnology,2014,32(9):947-951.)、玉米(zhen等,journal of genetics and genomics,2014,41(2):63-68.)、高粱(jiang等,nucleic acids research,2013,41(20):e188.)、二穗短柄草、番茄(brooks等,plant physiology,2014,166(3):1292-1297)等内的24个科45个属的植物(shan等,applications in plant sciences,2020,8(1):e11314.),在提高作物产量、品质、抗性等方面具有良好的应用前景。crispr/cas9系统的出现同样也为林木基因功能研究和遗传改良起到了很大的促进作用。但相较于草本植物和粮食作物,crispr/cas9系统在木本植物的相关研究及应用较为滞后,多数仍处于基因编辑体系的建立阶段。柑橘属植物是最早尝试应用crispr/cas9进行基因编辑的木本植物,此外该技术还成功应用于猕猴桃属、葡萄属、苹果属、石榴属、咖啡属、可可属、木薯属等。同时,其在杨树等经济木本植物中的成功应用展现了crispr/cas9系统在调控植物逆境耐受性和缩短林木育种周期等方面的巨大潜力,实现了抗旱、抗病等林木的新品种培育,为林木基础研究和分子育种提供了新的途径,为精准改良植物性状和选育新品种提供了新的思路。
4.为提高crispr/cas9系统的基因编辑效率,人们不断对载体进行优化。决定靶点特异性的sgrna是一段具有特定二级结构的小rna,通常由u3/u6系列启动子驱动。u3/u6启动子是crispr/cas9基因编辑系统的重要元件之一,其转录起始位点分别为a和g,且转录活性
较高。其中u3多用于单子叶植物,u6多用于双子叶植物。选择具有明确起始位点的u3或u6启动子,能够精确引导sgrna的转录,从而减少无关dna转录带来的脱靶效应。虽然u3/u6已在多个物种的基因编辑中成功运用,但同一启动子在同源关系较远的物种间并不一定适用,且同一物种基因中常存在多个u3或u6启动子,其活性及转录效率存在一定差异。因此克隆出更多目标植物的内源u3/u6启动子,有利于crispr/cas9基因编辑系统的完善。凡惠金等人在毛竹中克隆出两个peu3启动子并进行不同长度的截短,发现不同启动子及同一启动子不同截短长度时转录活性不同(凡惠金等,植物学报,2020,55(03):299-307.)。浦艳等在番茄中验证启动子转录活性时发现克隆的u3启动子长度在250bp以内仍具有转录活性(蒲艳,华北农学报,2019(1):33-39.)。基于构建crispr/cas9基因编辑载体的需要,所用的u3/u6启动子在保证其具有较高转录活性的基础上,长度要求尽量短,以保证尽可能不含有酶切位点,已有研究表明,用于crispr/cas9基因编辑技术的u6启动子通常只有200-400bp长(fauser等,the plant journal,2014,79(2):348-359.),甚至短于100bp(vladimir等,nature biotechnology,2013,31(8):691-693.)。long等利用棉花内源u6启动子将sgrna的表达水平提高了6-7倍,基因编辑效率也提高了4-5倍(longs等,plant methods.2018,14(1):80.)。刘春霞等人利用番茄u6启动子驱动sgrna的表达,与拟南芥的u6启动子驱动的sgrna相比较,番茄基因编辑效率从63%提高到73%(刘春霞等,分子植物育种,2020,18(20):6716-6724.)。
5.成功建立应用于水曲柳的crispr/cas9基因编辑体系将为水曲柳定向突变体库的获取提供高效可靠的技术支持,为深入研究水曲柳基因功能及其基因资源的开发利用提供宝贵的材料基础。然而,迄今为止,在水曲柳上对u6启动子仍然缺乏研究,缺少适用的、尽可能短且具有较高转录活性的内源u6启动子已成为水曲柳crispr/cas9基因编辑系统构建的限制因子,也限制了crispr/cas9系统在水曲柳遗传育种及种质创新等方面的应用。因此,优先克隆出具有高效转录活性的水曲柳内源fmu6启动子,在此基础上对其长度进行截短,构建以水曲柳内源u6基因启动子驱动水曲柳靶基因sgrna转录的crispr/cas9载体,应用于水曲柳功能基因研究及遗传育种,具有重要的研究意义及应用价值。
技术实现要素:
6.针对目前研究现状,本发明的目的是提供一种水曲柳u6基因启动子fmu6.7和该启动子的克隆方法及应用。
7.本发明解决其技术问题所采用的技术方案之一是:提供了一种水曲柳内源u6基因启动子profmu6.7,其核苷酸序列如seq id no:1所示。
8.优选地,所述水曲柳u6基因启动子profmu6.7属于水曲柳u6 snrna基因的rna聚合酶ⅲ型启动子。
9.优选地,所述水曲柳u6基因启动子profmu6.7的核苷酸序列中包含102bp u6 snrna。
10.本发明解决其技术问题所采用的技术方案之二是:提供了一种克隆所述的水曲柳fmu6基因启动子profmu6.7的方法,包括如下步骤:
11.(1)以水曲柳无菌苗dna为模板,设计如下特异引物:
12.profmu6.7-f:tataccaaccattccttctcactg
13.profmu6.7-r:ttctggacaaaaccctggc
14.(2)使用la taq酶在50μl体系中进行pcr克隆,pcr扩增的反应程序为:95℃预变性2min,95℃变性30s,58℃退火30s,72℃延伸2min,35个循环,70℃终延伸10min。
15.(3)将扩增产物克隆到pcloneez-topo载体上,转化大肠杆菌dh5α,挑取重组单克隆测序,即分别获取长度为1506bp的水曲柳u6基因启动子profmu6.7。
16.本发明解决其技术问题所采用的技术方案之三是:提供了一种水曲柳u6基因截短启动子的克隆方法,包括如下步骤:
17.(1)以水曲柳u6基因启动子profmu6.7阳性重组单克隆质粒为模板,分别设计不同序列长度的如下特异引物:
18.profmu6.7.4-f:taacatcgttgggtaaatg
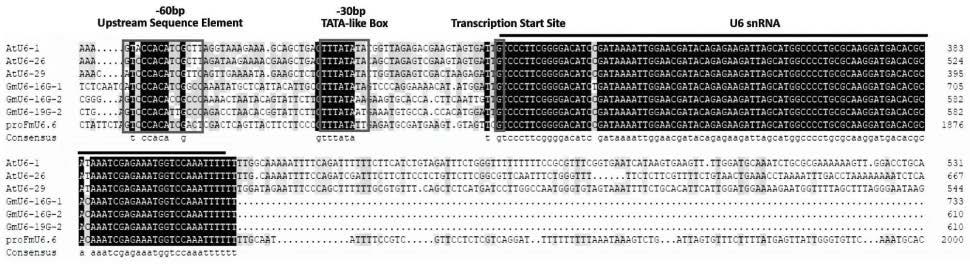
19.profmu6.7.4-r:aattttatcggatgtccccg
20.(2)使用phanta max master mix高保真酶在50μl体系中进行pcr克隆,pcr扩增的反应程序为:95℃预变性3min,95℃变性15s,57℃退火15s,72℃延伸30-60sec/kb,30个循环,72℃终延伸7min。
21.(3)将扩增产物克隆到pcloneez-topo载体上,转化大肠杆菌dh5α,挑取重组单克隆测序,即分别获取长度为334bp的水曲柳u6基因截短启动子profmu6.7.4。
22.优选地,所述水曲柳u6基因截短启动子profmu6.7.4含有27bp snrna,其核苷酸序列选自seq id no:1中937-1270位所示核苷酸序列;
23.本发明解决其技术问题所采用的技术方案之四是:提供了水曲柳u6基因启动子profmu6.7在水曲柳分子育种技术领域中的应用。
24.本发明具有如下有益效果:本发明首次在水曲柳中克隆获得了水曲柳u6 snrna基因的rna聚合酶ⅲ型启动子——水曲柳内源u6启动子profmu6.7,将其与gus基因融合,瞬时转化水曲柳无菌幼苗,通过gus瞬时表达验证了该启动子能够在水曲柳中高效表达,为水曲柳及近缘植物的转化研究提供了高效的启动子序列。此外,本发明首次将具有高效启动活性的水曲柳u6基因启动子profmu6.7进行截短,获得水曲柳u6基因截短启动子profmu6.7.4,通过与gus基因融合表达载体的构建,验证了水曲柳u6基因截短启动子仍具有高效启动活性,并将水曲柳u6基因截短启动子profmu6.7.4构建到基因编辑载体中,获得水曲柳内源u6基因启动子启动向导rna转录的基因编辑载体psc1-fmu6.7.4-cas9-hyp。因此,本发明克隆水曲柳u6基因截短启动子fmu6.7.4可应用于水曲柳crispr/cas9基因编辑体系,从而实现对水曲柳高效精准的种质创新与品种遗传改良。
附图说明
25.图1为水曲柳u6基因启动子profmu6.7与拟南芥及大豆u6启动子序列比对,其中,框线位置分别为u6 snrna转录的关键元件use(upstream sequence element)、tata-like box及转录起始位点,横线标注位置为u6 snrna;
26.图2为水曲柳u6基因启动子profmu6.7的克隆电泳图,可见经过pcr扩增分别得到了1506bp的水曲柳u6基因启动子profmu6.7片段;
27.图3为水曲柳u6基因截短启动子profmu6.7.4示意图,其中短线位置为caat-box;
28.图4为水曲柳u6基因截短启动子profmu6.7.4的克隆电泳图,可见经过pcr扩增分
别得到了334bp的水曲柳u6基因截短启动子profmu6.7.4片段;
29.图5为水曲柳无菌苗gus染色情况,control是使用农杆菌gv3101瞬时侵染的对照组。
30.图6为水曲柳重组基因编辑载体psc1-fmu6.7.4-cas9-hyp结构示意图,其中lgui酶切位点为靶基因sgrna序列插入位置。
31.图7为sgrna在水曲柳u6基因截短启动子profmu6.7.4启动下的转录水平
具体实施方式
32.为了更好地理解本发明,下面结合实施例和附图对本发明做进一步的详细说明,但本领域技术人员应当了解,下述实施例不是对本发明保护范围的限制,任何在本发明基础上做出的改变和变化,都属于本发明的保护范围之内。
33.下述实施例中,所使用的实验方法如无特别说明,均为常规方法。
34.下述实例中启动子活性检测载体pnc-121-pro为nc biotech公司的pnc系列载体,该生物材料只为重复本发明的相关实验所用,不作为其他用途使用。
35.下述实施例中所使用的材料、试剂等,如无特殊说明,均可从商业途径得到。
36.实施例1:
37.水曲柳u6基因启动子profmu6.7的获得,具体操作如下:
38.(1)根据u6 snrna序列在不同物种间的保守性,利用拟南芥atu6启动子的snrna序列(gtcccttcggggacatccgataaaattggaacgatacagagaagattagcatggcccctgcgcaaggatgacacgcataaatcgagaaatggtccaaatttt)及大豆gmu6启动子的snrna序列(gtcccttcggggacatccgataaaattggaacgatacagagaagattagcatggcccctgcgcaaggatgacacgcacaaatcgagaaatggtccaaatttt)与水曲柳基因组序列进行比对(blast)。检查比对结果,从中挑选序列同源性高于或等于99%的位置,利用plant care在线分析软件对其上游1800bp序列进行启动子顺式作用元件分析。分析结果发现,use(upstream sequence element)及基本转录相关相关的tata-like box分别位于这些u6基因的转录起始位点上游60bp、30bp处。选择其中一条启动子序列命名为profmu6.7。
39.(2)以水曲柳无菌苗基因组dna为模板,在水曲柳u6基因启动子profmu6.7上下游设计特异引物:
40.profmu6.7-f:tataccaaccattccttctcactg
41.profmu6.7-r:ttctggacaaaaccctggc
42.其中下游引物位于基因组序列中水曲柳snrna下游300bp处,以保证克隆序列中包含完整的102bp的snrna。
43.(3)使用la taq酶在50μl体系中进行pcr克隆,pcr扩增的反应程序为:95℃预变性2min,95℃变性30s,58℃退火30s,72℃延伸2min,35个循环,70℃终延伸10min。
44.(4)将扩增产物克隆到pcloneez-topo载体上,转化大肠杆菌dh5α,挑取重组单克隆测序,最后分别获取如图2所示长度为1506bp的水曲柳u6基因启动子profmu6.7。
45.(5)使用dnaman将该启动子序列与拟南芥atu6-1、atu6-26、atu6-29及大豆gmu6-16g-1、gmu6-16g-2、gmu6-19g-2碱基序列比对分析,发现水曲柳u6基因启动子profmu6.7序列含有use元件及u6 snrna转录关键位点tata-like box,如图1所示,并且水曲柳fmu6基因
启动子序列中这两个元件相对于转录起始位点的位置与拟南芥atu6-1、atu6-26、atu6-29及大豆gmu6-16g-1、gmu6-16g-2、gmu6-19g-2启动子序列位置一致,这对其功能的行使具有重要意义。
46.(6)以水曲柳u6基因启动子profmu6.7阳性重组单克隆质粒为模板,设计如下特异引物:
47.profmu6.7.4-f:taacatcgttgggtaaatg
48.profmu6.7.4-r:aattttatcggatgtccccg
49.其中,水曲柳u6基因截短启动子profmu6.7.4片段位于u6 snrna上,以保证序列中包含27bp snrna,截短位置如图3所示。
50.(7)使用phanta max master mix高保真酶在50μl体系中进行pcr克隆,pcr扩增的反应程序为:95℃预变性3min,95℃变性15s,57℃退火15s,72℃延伸30-60sec/kb,30个循环,72℃终延伸7min。(8)将扩增产物克隆到pcloneez-topo载体上,转化大肠杆菌dh5α,挑取重组单克隆测序,即分别获取如图4所示长度为334bp的水曲柳u6基因截短启动子profmu6.7.4。
51.实施例2:
52.水曲柳u6基因启动子启动活性检测,具体操作如下:
53.(1)水曲柳u6基因启动子活性检测载体的构建:
54.分别设计如下用于与pnc-121-pro(pbi121框架,gus报告基因,将nc克隆框取代pbi121的35s启动子而成)同源重组克隆的含有同源臂的特异引物:
55.pnc-profmu6.7-f:cagtggtctctgtccagtccttataccaaccattccttctcactg
56.pnc-profmu6.7-r:cggtctcagcagaccacaagtttctggacaaaaccctggc
57.pnc-profmu6.7.4-f:cagtggtctctgtccagtccttaacatcgttgggtaaatg
58.pnc-profmu6.7.4-r:cggtctcagcagaccacaagtaattttatcggatgtccccg(下划线所示为与pnc-121-pro载体同源序列)
59.以水曲柳u6基因启动子fmu6.7、水曲柳u6基因截短启动子profmu6.7.4阳性重组单克隆质粒为模板,pcr扩增获得两端含有同源臂的水曲柳u6基因启动子dna片段,用nimble mix(a)将上述片段分别与pnc-121-pro质粒混合,吸打混匀10-20次后,用pcr仪50℃处理45分钟后,4℃恒温,最终获得pnc-121-pro:profmu6.7、pnc-121-pro:profmu6.7.4两种启动子活性检测载体。
60.(2)水曲柳u6基因启动子农杆菌转化验证:
61.将构建的五种启动子活性检测载体分别转化到农杆菌gv3101菌株,以接种15d的水曲柳无菌幼苗为瞬时转化外植体材料,进行农杆菌瞬时侵染,共培养3天后取出水曲柳幼苗进行gus染色观察,以评估水曲柳u6基因启动子的启动能力和表达活性。
62.将用于检测的水曲柳组培苗加入gus染液,真空抽滤半小时后于37℃摇床中避光过夜染色,倒掉染色液,95%酒精脱色3d,观察gus染色情况。以空载gv3101农杆菌侵染的水曲柳幼苗做对照,
63.染色情况如图5所示。
64.实施例3:
65.水曲柳u6基因截短启动子profmu6.7.4基因编辑重组载体的构建,具体操作如下:
66.(1)水曲柳u6基因截短启动子profmu6.7.4基因编辑载体的构建
67.使用asc i和lgu i双酶切psc1-gmu6-gmubi3载体,去掉该载体上大豆gmu6启动子,回收15472bp载体骨架大片段。
68.设计用于同源重组克隆的含同源臂的引物:
69.psc1-profmu6.7.4-f:atctttcactggcgcgcctaacatcgttgggtaaatg
70.psc1-profmu6.7.4-r:tctagctctaaaacagaagagcaattttatcggatgtccccg(下划线所示为与psc1-gmu6-gmubi3载体同源序列)
71.以水曲柳u6基因截短启动子fmu6.7.4阳性重组单克隆质粒为模板,pcr扩增获得两端含有同源臂的水曲柳u6基因截短启动子fmu6.7.4的dna片段,用2
×
master assembly mix将上述片段同源重组到线性化的psc1-gmu6-gmubi3载体上,获得水曲柳基因编辑载体psc1-fmu6.7.4-cas9-hyp,载体结构示意图如图6,其中箭头为靶基因sgrna插入位点。
72.(2)水曲柳u6基因截短启动子profmu6.7.4基因编辑重组载体的构建
73.为了验证水曲柳u6基因截短启动子profmu6.7.4在水曲柳基因编辑中的功能活性,选择水曲柳内源糖基转移酶gt8家族基因fmgaut9作为crispr/cas9编辑的目标基因,设计合适的靶位点:
74.gttaatccgcgggcttgtgca。
75.针对靶点,在靶序列两端加上接头序列,设计的靶位点克隆引物如下:
76.psc1-gaut9-f:gttgttaatccgcgggcttgtgca
77.psc1-gaut9-r:aactgcacaagcccgcggattaac(下划线所示为用于载体连接的接头序列)
78.分别将上述psc1-gaut9-f与psc1-gaut9-r靶点引物混匀,用pcr仪95℃处理3分钟后,1℃/20s降温至16℃退火,形成带有与载体互补粘性末端的双链退火产物,同时用lgu i酶切psc1-fmu6.7.4-cas9-hyp载体,然后使用t4 dna连接酶将靶点片段连接到酶切载体上,最终得到psc1-fmu6.7.4:gaut9-cas9-hyp水曲柳基因编辑重组载体。
79.(3)水曲柳u6基因截短启动子profmu6.7.4启动sgrna的转录
80.将编辑载体psc1-fmu6-7-4:gaut2-cas9-hyp转化到农杆菌gv3101菌株,以水曲柳幼苗为瞬时转化外植体材料,进行农杆菌瞬时侵染,分别置于28℃及35℃恒温培养箱,共培养3天后取出水曲柳幼苗蒸馏水反复冲洗,液氮速冻后提取rna,反转录为cdna后进行荧光定量pcr检测验证。
81.根据靶向fmgaut9基因的sgrna及sgrna骨架设计如下特异性定量引物:
82.sgrna9-f:cgggcttgtgcagtttta
83.sgrna9-r:cgactcggtgccactttt
84.actin-f:aggacgctgccaacaacttt
85.actin-r:ttgaggggaagggtaaatagtg
86.其中actin为水曲柳管家基因,其sgrna相对表达量如图7所示。
87.可见,本发明在水曲柳中获得了水曲柳u6 snrna基因的rna聚合酶ⅲ型启动子——水曲柳内源u6启动子fmu6.7,并被验证具有启动活性,一种水曲柳u6基因截短启动子profmu6.7.4,且具有启动活性,能够启动sgrna转录,并首次实现了将水曲柳内源u6启动子应用于crispr/cas9系统。因此,本发明所述启动子可应用于水曲柳crispr/cas9基因编
辑体系,从而实现对水曲柳高效精准的新种质创制与品种改良。
88.虽然以上描述了本发明的具体实施方式,但是熟悉本技术领域的技术人员应当理解,我们所描述的具体实施例只是说明性的,而不是用于对本发明的范围的限定,对于本技术领域的技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的权力要求所保护的范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。