使用由脂质纳米颗粒递送的crispr/cas系统在动物中进行的转录调控
1.相关申请的交叉引用
2.本技术要求于2019年9月13日提交的美国申请第62/900,080号和于2020年6月23日提交的美国申请第63/042,762号的权益,这些美国申请中的每一个以全文引用的方式并入本文用于所有目的。
3.通过efs web作为文本文件提交的序列表的引用
4.写入文件693746seqlist.txt中的序列表为122千字节,创建于2020年9月11日,并且特此通过引用并入。
背景技术:
5.基因表达在许多生物过程(如发育和疾病)中受到严格控制。转录因子通过与靶基因的增强子和启动子区域处的特定dna序列结合来调控基因表达,并通过它们的效应子域调节转录。基于相同的原理,已经通过将各种功能域与dna结合域融合而生成人工转录因子(atf),该dna结合域被设计为与感兴趣的基因结合,从而调节它们的表达。然而,这些atf的结合特异性通常是退化的并且难以预测,并且atf的复杂和耗时的设计和生成限制了它们的应用。
6.基于crispr/cas的激活是功能基因探询的强大工具,但递送困难限制了其在体内的应用。体内的一个限制是需要同时将所有组分引入活生物体中,使得所有组分到达同一细胞并诱导靶基因转录的稳健和持续增加。需要更好的方法和工具来在体内引入crispr/cas试剂。
技术实现要素:
7.提供了在同一脂质纳米颗粒中一起包含crispr/cas协同激活介体系统组分的脂质纳米颗粒,以及使用此类脂质纳米颗粒来增加真核基因组、细胞和生物体中体内和离体靶基因表达和评估crispr/cas协同激活介体系统增加真核基因组、细胞和生物体中体内和离体靶基因表达的能力的方法。
8.在一个方面,提供了用于将负荷物递送至靶基因以增加靶基因在动物或细胞中的表达的脂质纳米颗粒(lnp)。在一些此类lnp中,该负荷物包含:(a)编码嵌合规律间隔成簇短回文重复序列(crispr)相关联(cas)蛋白的核酸,该蛋白包含与一个或多个转录激活域融合的无核酸酶活性的cas蛋白;(b)编码嵌合衔接蛋白的核酸,该嵌合衔接蛋白包含与一个或多个转录激活域融合的衔接蛋白;以及(c)一个或多个向导rna或编码该一个或多个向导rna的一个或多个核酸,每个向导rna包含嵌合衔接蛋白可特异性结合的一个或多个衔接结合元件,并且其中一个或多个向导rna中的每一个能够与cas蛋白形成复合物并将其导向至靶基因内的靶序列,从而增加靶基因的表达。
9.在一些此类lnp中,多顺反子或双顺反子核酸包含(a)和(b)。任选地,(a)和(b)通过多顺反子或双顺反子核酸中的2a蛋白编码序列链接。在一些此类lnp中,(a)和(b)是单独
的核酸。在一些此类lnp中,(a)和(b)各自均为信使rna(mrna)的形式。任选地,mrna被修饰为被假尿苷完全取代。任选地,mrna是包含(a)和(b)的多顺反子或双顺反子核酸,其中该mrna包含seq id no:61所示的序列或包含与seq id no:61所示序列至少90%、95%、96%、97%、98%、99%或100%相同的序列(并且任选地编码与seq id no:61相同的蛋白)。在一些此类lnp中,(c)为rna的形式。任选地,一个或多个向导rna中的每一个被修饰以在5'末端和/或3'末端包含一个或多个稳定末端修饰。任选地,一个或多个向导rna中的每一个的5'末端和/或3'末端被修饰以包含一个或多个硫代磷酸酯键。任选地,一个或多个向导rna中的每一个的5'末端和/或3'末端被修饰以包含一个或多个2'-o-甲基修饰。
10.在一些此类lnp中,靶序列包含靶基因内的调控序列。任选地,调控序列包含启动子或增强子。在一些此类lnp中,靶序列在靶基因的转录起始位点的200个碱基对内。任选地,靶序列在转录起始位点上游200个碱基对和转录起始位点下游1个碱基对的区域内。
11.在一些此类lnp中,一个或多个向导rna中的每一个均包含嵌合衔接蛋白可以特异性结合的两个衔接结合元件。任选地,第一衔接结合元件处于所述一个或多个向导rna中的每个向导rna的第一环内,并且第二衔接结合元件处于所述一个或多个向导rna中的每个向导rna的第二环内。任选地,一个或多个向导rna中的每一个是包含与反式激活crispr rna(tracrrna)部分融合的crispr rna(crrna)部分的单个向导rna,并且第一环是对应于seq id no:12、14、52或53的残基13至16的四环,并且第二环是对应于seq id no:12、14、52或53的残基53至56的茎环2。
12.在一些此类lnp中,衔接结合元件包含seq id no:16所示的序列。任选地,一个或多个向导rna中的每一个包含seq id no:40、45、56或57所示的序列。
13.在一些此类lnp中,一个或多个向导rna中的至少一个靶向ttr基因,任选地,其中靶向ttr的向导rna靶向包含seq id no:34至36中任一个所示的序列的序列,或者任选地,其中靶向ttr的向导rna包含seq id no:37至39和55中任一个所示的序列。
14.在一些此类lnp中,一个或多个向导rna靶向两个或更多个靶基因。在一些此类lnp中,一个或多个向导rna包含靶向单个靶基因的多个向导rna。在一些此类lnp中,一个或多个向导rna包含靶向单个靶基因的至少三个向导rna。任选地,该至少三个向导rna靶向小鼠ttr基因座,并且其中第一向导rna靶向包含seq id no:34的序列或者包含seq id no:37所示的序列,第二向导rna靶向包含序列seq id no:35的序列或者包含seq id no:38所示的序列,并且第三向导rna靶向包含seq id no:36的序列或者包含seq id no:39或55所示的序列。
15.在一些此类lnp中,cas蛋白是cas9蛋白。任选地,cas9蛋白为酿脓链球菌cas9蛋白、空肠弯曲杆菌cas9蛋白或金黄色葡萄球菌cas9蛋白。任选地,当与酿脓链球菌cas9蛋白最佳比对时,cas9蛋白包含对应于d10a和n863a或d10a和h840a的突变。
16.在一些此类lnp中,编码cas蛋白的序列经过密码子优化以在动物或细胞中表达。
17.在一些此类lnp中,嵌合cas蛋白中的一个或多个转录激活子域选自:vp16、vp64、p65、myod1、hsf1、rta、set7/9及其组合。任选地,嵌合cas蛋白中的一个或多个转录激活子域包含vp64。任选地,嵌合cas蛋白从n端到c端包含:无催化活性的cas蛋白;核定位信号;以及vp64转录激活子域。任选地,嵌合cas蛋白包含与seq id no:1所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。任选地,编码嵌合cas蛋白的核酸包含与
seq id no:25所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。
18.在一些此类lnp中,衔接蛋白位于嵌合衔接蛋白的n末端,并且一个或多个转录激活域位于嵌合衔接蛋白的c末端。在一些此类lnp中,衔接蛋白包含ms2外壳蛋白或其功能片段或变体。在一些此类lnp中,嵌合衔接蛋白中的一个或多个转录激活域选自:vp16、vp64、p65、myod1、hsf1、rta、set7/9及其组合。任选地,嵌合衔接蛋白中的一个或多个转录激活域包含p65和hsf1。任选地,嵌合衔接蛋白从n端到c端包含:ms2外壳蛋白;核定位信号;p65转录激活域;以及hsf1转录激活域。任选地,嵌合衔接蛋白包含与seq id no:6所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。任选地,编码嵌合衔接蛋白的核酸包含与seq id no:27所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。
19.在一些此类lnp中,动物是非人动物。在一些此类lnp中,动物是哺乳动物。任选地,所述哺乳动物是啮齿动物。任选地,该啮齿动物是大鼠或小鼠。任选地,该啮齿动物是小鼠。在一些此类lnp中,动物是人。在一些此类lnp中,靶基因是在肝脏中表达的基因。
20.在一些此类lnp中,靶基因是疾病相关联基因。在一些此类lnp中,靶基因的表达或活性降低与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。在一些此类lnp中,靶基因是单倍体不足基因或者是otc、hbg1或hbg2。任选地,单倍体不足基因是kcnq4、pink1、tp73、glut1、myh、abca4、lrh-1、pax8、slc40a1、bmpr2、pkd2、pik3r1、hmga1、gck、eln、gtf3、gata3、bub3、pax6、fli1、hnf1a、pkd1、mc4r、dmpk或myh9。任选地,该单倍体不足基因为表2或表3中的任一种基因。在一些此类lnp中,靶基因的表达或活性增加与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。
21.一些此类lnp包含阳离子脂质、中性脂质、辅助脂质和隐形脂质。任选地,阳离子脂质是mc3和/或中性脂质是dspc和/或辅助脂质是胆固醇和/或隐形脂质是peg-dmg。任选地,lnp包含摩尔比为约50:10:38.5:1.5的mc3、dspc、胆固醇和peg-dmg。
22.在另一方面,提供了用于在动物体内或在离体或在体动物细胞中增加靶基因的表达的方法。同样地,提供了用于在体外在动物细胞中增加靶基因的表达的方法。一些此类方法包含向动物或细胞中引入:(a)编码嵌合规律间隔成簇短回文重复序列(crispr)相关联(cas)蛋白的核酸,该蛋白包含与一个或多个转录激活域融合的无核酸酶活性的cas蛋白;(b)编码嵌合衔接蛋白的核酸,该嵌合衔接蛋白包含与一个或多个转录激活域融合的衔接蛋白;以及(c)一个或多个向导rna或编码该一个或多个向导rna的一个或多个核酸,每个向导rna包含嵌合衔接蛋白可特异性结合的一个或多个衔接结合元件,并且其中一个或多个向导rna中的每一个能够与cas蛋白形成复合物并将其导向至靶基因内的靶序列,从而增加靶基因的表达,其中(a)、(b)和(c)一起在同一脂质纳米颗粒(lnp)中递送。
23.在一些此类方法中,多顺反子或双顺反子核酸包含(a)和(b)。任选地,(a)和(b)通过多顺反子或双顺反子核酸中的2a蛋白编码序列链接。在一些此类方法中,(a)和(b)是单独的核酸。在一些此类方法中,(a)和(b)各自均以信使rna(mrna)的形式引入。任选地,mrna被修饰为被假尿苷完全取代。任选地,mrna是包含(a)和(b)的多顺反子或双顺反子核酸,其中该mrna包含seq id no:61所示的序列或包含与seq id no:61所示序列至少90%、95%、96%、97%、98%、99%或100%相同的序列(并且任选地编码与seq id no:61相同的蛋白)。在一些此类方法中,(c)以rna的形式引入。任选地,一个或多个向导rna中的每一个被修饰
以在5'末端和/或3'末端包含一个或多个稳定末端修饰。任选地,一个或多个向导rna中的每一个的5'末端和/或3'末端被修饰以包含一个或多个硫代磷酸酯键。任选地,一个或多个向导rna中的每一个的5'末端和/或3'末端被修饰以包含一个或多个2'-o-甲基修饰。
24.在一些此类方法中,靶序列包含靶基因内的调控序列。任选地,调控序列包含启动子或增强子。在一些此类方法中,靶序列在靶基因的转录起始位点的200个碱基对内。任选地,靶序列在转录起始位点上游200个碱基对和转录起始位点下游1个碱基对的区域内。
25.在一些此类方法中,一个或多个向导rna中的每一个均包含嵌合衔接蛋白可以特异性结合的两个衔接结合元件。任选地,第一衔接结合元件处于所述一个或多个向导rna中的每个向导rna的第一环内,并且第二衔接结合元件处于所述一个或多个向导rna中的每个向导rna的第二环内。任选地,一个或多个向导rna中的每一个是包含与反式激活crispr rna(tracrrna)部分融合的crispr rna(crrna)部分的单个向导rna,并且第一环是对应于seq id no:12、14、52或53的残基13至16的四环,并且第二环是对应于seq id no:12、14、52或53的残基53至56的茎环2。
26.在一些此类方法中,衔接结合元件包含seq id no:16所示的序列。任选地,一个或多个向导rna中的每一个包含seq id no:40、45、56或57所示的序列。
27.在一些此类方法中,一个或多个向导rna中的至少一个靶向ttr基因,任选地,其中靶向ttr的向导rna靶向包含seq id no:34至36中任一个所示的序列的序列,或者任选地,其中靶向ttr的向导rna包含seq id no:37至39和55中任一个所示的序列。
28.在一些此类方法中,一个或多个向导rna靶向两个或更多个靶基因。在一些此类方法中,一个或多个向导rna包含靶向单个靶基因的多个向导rna。在一些此类方法中,一个或多个向导rna包含靶向单个靶基因的至少三个向导rna。任选地,该至少三个向导rna靶向小鼠ttr基因座,并且其中第一向导rna靶向包含seq id no:34的序列或者包含seq id no:37所示的序列,第二向导rna靶向包含序列seq id no:35的序列或者包含seq id no:38所示的序列,并且第三向导rna靶向包含seq id no:36的序列或者包含seq id no:39或55所示的序列。
29.在一些此类方法中,所述cas蛋白是cas9蛋白。任选地,cas9蛋白为酿脓链球菌cas9蛋白、空肠弯曲杆菌cas9蛋白或金黄色葡萄球菌cas9蛋白。任选地,当与酿脓链球菌cas9蛋白最佳比对时,cas9蛋白包含对应于d10a和n863a或d10a和h840a的突变。
30.在一些此类方法中,编码cas蛋白的序列经过密码子优化以在动物中表达。
31.在一些此类方法中,嵌合cas蛋白中的一个或多个转录激活子域选自:vp16、vp64、p65、myod1、hsf1、rta、set7/9及其组合。任选地,嵌合cas蛋白中的一个或多个转录激活子域包含vp64。任选地,嵌合cas蛋白从n端到c端包含:无催化活性的cas蛋白;核定位信号;以及vp64转录激活子域。任选地,嵌合cas蛋白包含与seq id no:1所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。任选地,编码嵌合cas蛋白的核酸包含与seq id no:25所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。
32.在一些此类方法中,衔接蛋白位于嵌合衔接蛋白的n末端,并且一个或多个转录激活域位于嵌合衔接蛋白的c末端。在一些此类方法中,衔接蛋白包含ms2外壳蛋白或其功能片段或变体。在一些此类方法中,嵌合衔接蛋白中的一个或多个转录激活域选自:vp16、vp64、p65、myod1、hsf1、rta、set7/9及其组合。任选地,嵌合衔接蛋白中的一个或多个转录
激活域包含p65和hsf1。任选地,嵌合衔接蛋白从n端到c端包含:ms2外壳蛋白;核定位信号;p65转录激活域;以及hsf1转录激活域。任选地,嵌合衔接蛋白包含与seq id no:6所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。任选地,编码嵌合衔接蛋白的核酸包含与seq id no:27所示的序列至少90%、95%、96%、97%、98%、99%或100%相同的序列。
33.在一些此类方法中,动物是非人动物。在一些此类方法中,动物是哺乳动物。任选地,所述哺乳动物是啮齿动物。任选地,该啮齿动物是大鼠或小鼠。任选地,该啮齿动物是小鼠。在一些此类方法中,动物是人。在一些此类方法中,动物是需要增加靶基因表达的对象,其中靶基因在对象中低表达,并且该低表达与对象中的疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。在一些此类方法中,靶基因是在肝脏中表达的基因。在一些此类方法中,将一个或多个向导rna施用于动物的途径是静脉内注射、实质内注射、腹膜内注射、经鼻安装或玻璃体内注射。
34.在一些此类方法中,靶基因是疾病相关联基因。在一些此类方法中,靶基因的表达或活性降低与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。在一些此类方法中,靶基因是单倍体不足基因或者是otc、hbg1或hbg2。任选地,单倍体不足基因是kcnq4、pink1、tp73、glut1、myh、abca4、lrh-1、pax8、slc40a1、bmpr2、pkd2、pik3r1、hmga1、gck、eln、gtf3、gata3、bub3、pax6、fli1、hnf1a、pkd1、mc4r、dmpk或myh9。任选地,该单倍体不足基因为表2或表3中的任一种基因。在一些此类方法中,靶基因的表达或活性增加与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。
35.在一些此类方法中,脂质纳米颗粒包含阳离子脂质、中性脂质、辅助脂质和隐形脂质。任选地,阳离子脂质是mc3和/或中性脂质是dspc和/或辅助脂质是胆固醇和/或隐形脂质是peg-dmg。任选地,脂质纳米颗粒包含摩尔比为约50:10:38.5:1.5的mc3、dspc、胆固醇和peg-dmg。
36.在一些此类方法中,靶基因的表达增加相对于对照动物或细胞高出至少0.5倍、1倍、2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍或20倍。在一些此类方法中,靶基因表达增加的持续时间为至少约1天、至少约2天、至少约3天、至少约4天、至少约5天、至少约6天、至少约1周、至少约2周、至少约3周、至少约4周、至少约1个月或至少约2个月。
37.在一些此类方法中,将包含(a)、(b)和(c)的脂质纳米颗粒循序引入动物或细胞中两次或更多次。在一些此类方法中,将包含(a)、(b)和(c)的脂质纳米颗粒循序引入动物或细胞中三次或更多次。任选地,在每次循序引入脂质纳米颗粒后,靶基因的表达增加至至少相同的水平。任选地,与仅引入脂质纳米颗粒一次的方法相比,靶基因的表达增加至更高的水平。
附图说明
38.图1(未按比例)示出了ttr向导rna阵列的示意图。向导rna阵列等位基因从5'到3'包含:第一u6启动子;第一向导rna编码序列;第二u6启动子;第二向导rna编码序列;第三u6启动子;和第三向导rna编码序列。
39.图2(未按比例)示出了设计三个向导rna的示意图,这些向导rna靶向ttr转录起始位点的上游。
40.图3示出了通用单个向导rna(seq id no:45)的示意图,其中四环和茎环2已被ms2结合适体替换以促进融合到转录激活域的嵌合ms2外壳蛋白(mcp)的募集。
41.图4示出了通过elisa测定的在未处理的dcas9 sam小鼠、用aav8-gfp处理的dcas9sam小鼠,以及用包含ttr向导rna阵列的aav8处理的dcas9 sam小鼠中的ttr循环血清水平。示出了注射后5天、19天、2个月、3个月、4个月、5个月、6个月、7个月和8个月的结果。
42.图5a和5b示出了通过elisa测定的在未处理的dcas9 sam小鼠和用包含ttr向导rna的lnp(r-lnp 277)处理的dcas9 sam小鼠中,ttr循环血清水平(图5a)和ttr循环血清水平相对于基线的百分比变化(图5b)。示出了注射后1、3、6、8、10、13、17、20、27、34和67天的结果。
43.图6示出了通过elisa测定的在未处理的dcas9 sam小鼠和用包含ttr向导rna的lnp(r-lnp 277)处理的dcas9 sam小鼠中的ttr循环水平。示出了0.5mpk、1mpk和2mpk剂量在注射后1周、2周、3周、4周、5周、6周和7周的结果。所有值均绘制为平均值 /-sd。星号指示显著性,并且星号数指示小数点后0的数量(ttest)。
44.图7a示出了在dcas9 sam小鼠(r26
sam/sam
)中循序四周给药lnp
ttrga2
时的ttr循环水平。用0.5mpk的合成ttr ga2 sam向导配制lnp颗粒,并在零周和/或四周时引入到纯合dcas9 sam小鼠(r26
sam/sam
)(n=5)中。每周取血通过elisa确定蛋白表达水平。所有值均绘制为平均值 /-sd。星号指示显著性,并且星号数指示小数点后0的数量(ttest)。
45.图7b示出了在dcas9 sam小鼠(r26
sam/sam
)中循序两周给药lnp
ttrga2
时的ttr循环水平。用0.5mpk的合成ttr ga2 sam向导配制lnp颗粒,并在零周和/或两周时引入到纯合dcas9 sam小鼠(r26
sam/sam
)(n=5)中。每周取血通过elisa确定蛋白表达水平。所有值均绘制为平均值 /-sd。星号指示显著性,并且星号数指示小数点后0的数量(ttest)。
46.图8示出了在dcas9 sam小鼠(r26
sam/sam
)中循序零周、两周和四周给药lnp
ttrga2
时的ttr循环水平。用0.5mpk的合成ttr ga2 sam向导配制lnp颗粒,并在零周、两周和四周或仅在零周时引入到纯合dcas9 sam小鼠(r26
sam/sam
)(n=5)中。每周取血通过elisa确定蛋白表达水平。所有值均绘制为平均值 /-sd。
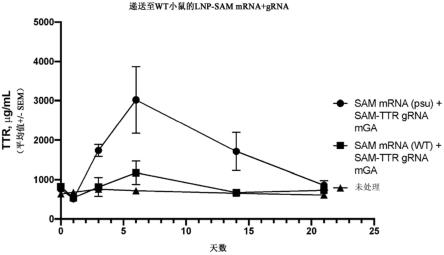
47.图9示出了用合成ttr sam向导和sam mrna(经假尿苷修饰或未经修饰)配制的lnp颗粒向野生型小鼠给药后的ttr循环水平。使用未处理小鼠作为阴性对照。在指示的时间点取血通过elisa确定蛋白表达水平。所有值均绘制为平均值 /-sem。
48.定义
49.本文可互换使用的术语“蛋白质”、“多肽”、和“肽”包含任何长度的聚合形式的氨基酸,包含编码氨基酸和非编码氨基酸以及以化学方式或生物化学方式修饰的氨基酸或以化学方式或生物化学方式衍生的氨基酸。这些术语还包含已经修饰的聚合物,如具有经修饰的肽骨架的多肽。术语“结构域”是指具有特定功能或结构的蛋白质或多肽的任何部分。
50.蛋白被视为具有“n端”和“c端”。术语“n端”涉及蛋白或多肽的起点,其终止于具有游离胺基(-nh2)的氨基酸。术语“c端”是指氨基酸链(蛋白质或多肽)的末端,其终止于游离羧基(-cooh)。
51.本文可互换使用的术语“核酸”和“多核苷酸”包含任何长度的聚合形式的核苷酸,包含核糖核苷酸、脱氧核糖核苷酸、或其类似物或经过修饰的形式。所述核苷酸包含单链、双链和多链dna或rna、基因组dna、cdna、dna-rna杂交体、和包括嘌呤碱基、嘧啶碱基或其它
天然的、以化学方式修饰的、以生物化学方式修饰的、非天然的或衍生的核苷酸碱基的聚合物。
52.核酸被视为具有“5'末端”和“3'末端”,因为以使得一个单核苷酸戊糖环的5'磷酸通过磷酸二酯键在一个方向上与其相邻的单核苷酸戊糖环的3'氧附着的方式使单核苷酸反应以形成寡核苷酸。如果寡核苷酸的5'磷酸不与单核苷酸戊糖环的3'氧相连,那么将寡核苷酸的端称为“5'末端”。如果寡核苷酸的3'氧不与另一个单核苷酸戊糖环的5'磷酸相连,那么将寡核苷酸的端称为“3'末端”。即使核酸序列处于更大的寡核苷酸的内部,所述核酸序列也可以被视为具有5'末端和3'末端。在线性或环状dna分子中,离散元件被称为“下游”或3'元件的“上游”或5'。
53.术语“表达载体”或“表达构建体”或“表达盒”是指含有期望编码序列的重组核酸,所述期望编码序列可操作地连接到在特定宿主细胞或生物体中表达可操作地连接的编码序列所必需的适当核酸序列。在原核生物中表达所必需的核酸序列通常包含启动子、操纵子(任选的)和核糖体结合位点以及其它序列。众所周知,真核细胞利用启动子、增强子以及终止信号和多聚腺苷酸化信号,但是在不牺牲必要表达的情况下可以删除一些元素并添加其它元素。
54.术语“靶向载体”是指可以通过同源重组、非同源末端连接介导的连结或任何其它重组方式引入到细胞基因组中的靶位置的重组核酸。
55.关于蛋白、核酸和细胞的术语“分离的”包括相对于通常可以原位存在的其它细胞或生物体组分相对纯化的蛋白、核酸和细胞,直至并且包括蛋白、核酸或细胞的基本上纯的制备。术语“分离的”还包括不具有天然存在的对应物的蛋白和核酸,或者已经化学合成并且因此基本上未被其它蛋白或核酸污染的蛋白或核酸。术语“分离的”还包含已经从与蛋白质、核酸或细胞天然伴随的大多数其它细胞组分或生物体组分(例如,其它细胞蛋白、核酸或细胞或胞外组分)中分离或纯化的蛋白质、核酸或细胞。
56.术语“野生型”包含具有在正常(与突变、患病、改变等相比)状态或情况下发现的结构和/或活性的实体。野生型基因和多肽通常以多种不同形式(例如,等位基因)存在。
57.术语“内源序列”是指在细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)内天然存在的核酸序列。例如,非人动物的内源性ttr序列是指天然存在于非人动物的ttr基因座处的天然ttr序列。
[0058]“外源性”分子或序列包含通常不以所述形式存在于细胞中的分子或序列。正常存在包含关于细胞的特定发育阶段和环境条件的存在。例如,外源性分子或序列可以包含细胞内对应的内源性序列的突变版本(如内源性序列的人源化版本),或者可以包含与细胞内的内源性序列相对应但形式不同(即,不在染色体内)的序列。相比之下,内源性分子或序列包含在特定环境条件下在特定发育阶段在特定细胞中通常以所述形式存在的分子或序列。
[0059]
当在核酸或蛋白质的上下文中使用时,术语“异源的”指示核酸或蛋白质包括在同一分子中并非天然地一起出现的至少两个区段。例如,当关于核酸的段或蛋白质的区段使用时,术语“异源的”指示核酸或蛋白质包括在自然界中未发现彼此处于相同关系(例如,连接在一起)的两个或更多个子序列。举例来说,核酸载体的“异源”区域是在自然界中未发现与其它分子缔合的另一个核酸分子内或与其附着的核酸区段。例如,核酸载体的异源区域可以包含侧接有在自然界中未发现与编码序列缔合的序列的编码序列。同样地,蛋白质的“异源”区域是在自然界中未发现与其它肽分子缔合的另一个肽分子(例如,融合蛋白或具有标签的蛋白质)内或与其附着的氨基酸的区段。相似地,核酸或蛋白质可以包括异源标记或异源分泌或定位序列。
[0060]“密码子优化”利用密码子的简并性,如指定氨基酸的三碱基对密码子组合的多样性所展示的,并且通常包含通过用宿主细胞的基因中更频繁或最频繁使用的密码子置换天然序列的至少一个密码子同时维持天然氨基酸序列来修饰核酸序列以在特定宿主细胞中增强表达的过程。例如,与天然存在的核酸序列相比,可以修饰对cas9蛋白进行编码的核酸以置换在给定的原核或真核细胞中具有更高使用频率的密码子,包含细菌细胞、酵母细胞、人细胞、非人细胞、哺乳动物细胞、啮齿动物细胞、小鼠细胞、大鼠细胞、仓鼠细胞或任何其它宿主细胞。密码子使用表很容易获得,例如,在“密码子使用数据库”中获得。这些表可以通过多种方式进行调整。参见nakamura等人,(2000)《核酸研究(nucleic acids research)》28(1):292,该文献以全文引用的方式并入本文用于所有目的。也可获得用于在特定宿主中表达的特定序列的密码子优化的计算机算法(参见例如,《基因伪造(gene forge)》)。
[0061]
术语“基因座”是指基因(或显著序列)、dna序列、多肽编码序列或生物体的基因组的染色体上的位置的特异性定位。例如,“ttr基因座”可以指ttr基因、ttr dna序列、ttr编码序列或在生物体的基因组的染色体上的ttr位置的特定位置,该ttr位置已经被鉴别为此类序列存在的位置。“ttr基因座”可以包含ttr基因的调控元件,包括例如增强子、启动子、5'和/或3'非翻译区(utr)或其组合。
[0062]
术语“基因”是指染色体中的对产物(例如,rna产物和/或多肽产物)进行编码并且包含被非编码内含子中断的编码区和定位于5'末端和3'末端两者上的编码区附近而使得基因与全长mrna(包含5'和3'非翻译序列)相对应的序列的dna序列。术语“基因”还包含其它非编码序列,包含调节序列(例如,启动子、增强子和转录因子结合位点)、聚腺苷酸化信号、内部核糖体进入位点、沉默子、绝缘序列和基质附着区域。这些序列可以靠近基因的编码区(例如,在10kb内)或处于远处的位点,并且所述序列影响基因的转录和翻译的水平或速率。
[0063]
术语“等位基因”是指基因的变体形式。一些基因具有多种不同的形式,所述基因定位于染色体上的相同位置或基因位点处。二倍体生物体在每个基因座处具有两个等位基因。每对等位基因表示特异性基因座的基因型。如果在特定基因座处有两个相同的等位基因,则基因型被描述为纯合的,如果两个等位基因不同,则基因型被描述为杂合的。
[0064]“启动子”是dna的调控区,其通常包括能够引导rna聚合酶ii在特定多核苷酸序列的适当转录起始位点处起始rna合成的tata盒。启动子可以另外包括影响转录起始速率的其它区域。本文所公开的启动子序列调节可操作连接的多核苷酸的转录。启动子可以在本文所公开的细胞类型(例如,真核细胞、非人哺乳动物细胞、人细胞、啮齿动物细胞、多能性细胞、单细胞期胚胎、分化细胞或其组合)中的一种或多种细胞类型中具有活性。启动子可以是例如组成型活性启动子、条件型启动子、诱导型启动子、时间受限启动子(例如,受发育调控的启动子)或空间受限启动子(例如,细胞特异性或组织特异性启动子)。启动子的实例可以例如在wo 2013/176772中找到,所述文献出于所有目的通过引用整体并入本文。
[0065]
组成型启动子是在所有发育阶段的所有组织或特定组织中都具有活性的启动子。
harbor,n.y.),1989),该文献以全文引用的方式并入本文用于所有目的。
[0072]
杂交要求两个核酸含有互补序列,但是碱基之间有可能错配。两个核酸之间杂交的适当条件取决于核酸的长度和互补程度,这些变量是众所周知的。两个核苷酸序列之间的互补程度越大,具有这些序列的核酸杂交体的解链温度(tm)值就越大。对于具有较短互补段(例如,在35个或更少、30个或更少、25个或更少、22个或更少、20个或更少或18个或更少的核苷酸上互补)的核酸之间的杂交,错配位置变得尤为重要(参见sambrook等人,同上,11.7-11.8)。通常,可杂交核酸的长度为至少约10个核苷酸。可杂交核酸的说明性最小长度包含至少约15个核苷酸、至少约20个核苷酸、至少约22个核苷酸、至少约25个核苷酸和至少约30个核苷酸。此外,可以根据如互补区的长度和互补程度等因素,根据需要调整温度和洗涤溶液盐浓度。
[0073]
多核苷酸序列不必与其可特异性杂交的靶核酸具有100%互补性。此外,多核苷酸可以在一个或多个段上杂交,使得插入或相邻段不参与杂交事件(例如,环结构或发夹结构)。多核苷酸(例如,grna)可以包括与其靶向的靶核酸序列内的靶区域的至少70%、至少80%、至少90%、至少95%、至少99%或100%的序列互补性。例如,其中20个核苷酸中的18个与靶区域互补并因此特异性杂交的grna将表示90%的互补性。在此实施例中,剩余的非互补核苷酸可以与互补核苷酸成簇或散布并且不需要彼此相邻或与互补核苷酸相邻。
[0074]
核酸内特定核酸序列段之间的互补性百分比可以通过以下方式常规地确定:使用blast程序(基本局部比对搜索工具)和powerblast程序(altschul等人(1990)《分子生物学杂志(j.mol.biol.)》215(3):403-410;zhang和madden(1997)《基因组研究(genome res.)》7(6):649-656)或使用gap程序(威斯康星州麦迪逊大学研究园(university research park,madison wis.),遗传学计算机组,unix第8版,威斯康星序列分析包),该程序使用默认设置,其使用史密斯-沃特曼(smith and waterman)(1981)算法《应用数学进展(adv.appl.math.)》2(4):482-489。
[0075]
本文所提供的方法和组合物采用多种不同的组分。贯穿说明书的一些组分可以具有活性变体和片段。此类组分包含例如cas蛋白、crispr rna、tracrrna和向导rna。这些组分中的每个组分的生物活性在本文其它地方描述。术语“功能性”是指蛋白质或核酸(或其片段或变体)表现出生物活性或功能的先天能力。此类生物活性或功能可以包含例如cas蛋白结合向导rna和靶dna序列的能力。与原始分子相比,功能性片段或变体的生物学功能可以相同或实际上可以改变(例如,关于其特异性或选择性或功效)但保留分子的基本生物学功能。
[0076]
术语“变体”是指与群体中最普遍的序列不同的核苷酸序列(例如,相差一个核苷酸)或与群体中最普遍的序列不同的蛋白质序列(例如,相差一个氨基酸)。
[0077]
当提及蛋白时,术语“片段”意指比全长蛋白更短或具有更少氨基酸的蛋白。当提及核酸时,术语“片段”意指比全长核酸更短或具有更少核苷酸的核酸。当提及蛋白质片段时,片段可以是例如n端片段(即,去除蛋白质的c末端的一部分)、c端片段(即,去除蛋白质的n末端的一部分)或内部片段(即,去除蛋白质的n末端和c末端中的每个末端的一部分)。当提及核酸片段时,片段可以是例如5'片段(即,去除核酸的3'末端的一部分)、3'片段(即,去除核酸的5'末端的一部分)或内部片段(即,去除核酸的5'末端和3'末端中的每个末端的一部分)。
[0078]
在两个多核苷酸或多肽序列的上下文中,“序列同一性”或“同一性”是指当在指定的比较窗口上针对最大对应性进行比对时两个序列中相同的残基。当提及蛋白质的序列同一性的百分比时,不相同的残基位置通常因保守性氨基酸取代而不同,其中氨基酸残基被具有相似化学性质(例如,电荷或疏水性)的其它氨基酸残基取代,因此不改变分子的功能性质。当序列的保守性取代不同时,可以将百分比序列同一性向上调整以校正取代的保守性质。因此类保守性取代而不同的序列被视为具有“序列类似性”或“类似性”。用于进行这种调整的方法众所周知。通常,这涉及将保守性取代计为部分错配而不是完全错配,从而增加百分比序列同一性。因此,例如,当相同氨基酸的所得得分为1,非保守性取代的所得得分为零时,保守性取代的所得得分介于零与1之间。例如,通过在项目pc/gene(加利福尼亚州山景城的intelligenetics公司(intelligenetics,mountain view,california))中的实施方式计算保守性取代的得分。
[0079]“序列同一性百分比”包含通过在比较窗口上比较两个最佳比对序列测定的值(完全匹配残基的最大数量),其中在比较窗口中的多核苷酸序列部分与参考序列(不包括添加物或缺失部分)相比可以包括添加物或缺失部分(即缺口),以实现两个序列的最佳比对。通过测定在两个序列中出现相同核酸碱基或氨基酸残基的位置数计算百分比来得到匹配位置数,用匹配位置数除以比较窗口中的位置总数,并将结果乘以100以得到序列同一性的百分比。除非另有说明(例如,较短的序列包含连接的异源序列),否则所述比较窗口为两个所比较序列中较短序列的全长。
[0080]
除非另有说明,否则序列同一性/相似性值包含使用以下参数使用第10版gap获得的值:使用gap权重50、长度权重3以及nwsgapdna.cmp得分矩阵的核苷酸序列的同一性百分比和相似性百分比;使用gap权重8和长度权重2以及blosum62得分矩阵的氨基酸序列的同一性百分比和相似性百分比;或其任何等效程序。“等效程序”包含当与第10版gap生成的对应比对进行比较时针对所讨论的任何两个序列产生具有相同核苷酸或氨基酸残基匹配和相同百分比序列同一性的比对的任何序列比较程序。
[0081]
术语“保守性氨基酸取代”是指用具有相似大小、电荷或极性的不同氨基酸取代序列中正常存在的氨基酸。保守性取代的实例包含用非极性(疏水性)残基(如异亮氨酸、缬氨酸或亮氨酸)取代另一种非极性残基。同样地,保守性取代的实例包含用一种极性(亲水性)残基取代另一种极性残基,如精氨酸与赖氨酸之间的极性残基、谷氨酰胺与天冬酰胺之间的极性残基或甘氨酸与丝氨酸之间的极性残基。另外,用碱性残基(如赖氨酸、精氨酸或组氨酸)取代另一种碱性残基或者用一种酸性残基(如天冬氨酸或谷氨酸)取代另一种酸性残基是保守性取代另外的实例。非保守性取代的实例包含用非极性(疏水性)氨基酸残基(如异亮氨酸、缬氨酸、亮氨酸、丙氨酸或甲硫氨酸)取代极性(亲水性)残基(如半胱氨酸、谷氨酰胺、谷氨酸或赖氨酸)和/或用极性残基取代非极性残基。典型的氨基酸分类总结在下表1中。
[0082]
表1:氨基酸分类。
[0083][0084]“同源”序列(例如,核酸序列)包含与已知参考序列相同或基本上相似的序列,使得其例如与已知参考序列具有至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或100%同一性。同源序列可以包含例如直系同源序列和旁系同源序列。例如,同源基因通常通过物种形成事件(直系同源基因)或基因复制事件(旁系同源基因)从共同的祖先dna序列下降。“直系同源”基因包含不同物种中通过物种形成从共同祖先基因进化而来的基因。直系同源物通常在进化过程中保留相同的功能。“旁系同源”基因包含通过基因组内的复制相关的基因。旁系同源物可以在进化过程中进化出新的功能。
[0085]
术语“体外(in vitro)”包括人工环境和在人工环境(例如,试管或分离的细胞或细胞系)内发生的过程或反应。术语“体内(in vivo)”包含自然环境(例如,细胞、生物体或身体)以及在自然环境内发生的过程或反应。术语“离体”包含已从个体体内取出的细胞以及在此类细胞内发生的过程或反应。
[0086]
术语“报告基因”是指具有对基因产物(通常是酶)进行编码的序列的核酸,当包括与内源或异源启动子和/或增强子元件可操作地连接的报告基因序列的构建体被引入到含有(或者可以使其含有)激活启动子和/或增强子元件所必需的因子的细胞中时,所述核酸易于且可定量地进行测定。报告基因的实例包含但不限于对β-半乳糖苷酶(lacz)进行编码的基因、细菌氯霉素乙酰转移酶(cat)基因、萤火虫荧光素酶基因、对β-葡萄糖醛酸酶(gus)进行编码的基因和对荧光蛋白进行编码的基因。“报告蛋白”是指由报告基因编码的蛋白质。
[0087]
如本文所使用的,术语“荧光报告蛋白”意指基于荧光可检测的报告蛋白,其中荧光可以直接来自报告蛋白、报告蛋白在荧光底物上的活性,或对与荧光标记的化合物结合具有亲和力的蛋白质。荧光蛋白的实例包含绿色荧光蛋白(例如,gfp、gfp-2、taggfp、turbogfp、egfp、祖母绿(emerald)、azami绿、单体azami绿、copgfp、acegfp和zsgreenl)、黄
色荧光蛋白(例如,yfp、eyfp、柠檬黄、venus、ypet、phiyfp和zsyellowl)、蓝色荧光蛋白(例如,bfp、ebfp、ebfp2、石青、mkalamal、gfpuv、天蓝色和t-天蓝色(t-sapphire))、青色荧光蛋白(例如cfp、ecfp、蔚蓝色(cerulean)、cypet、amcyanl和midoriishi-青色)、红色荧光蛋白(例如,rfp、mkate、mkate2、mplum、dsred单体、mcherry、mrfp1、dsred-表达、dsred2、dsred-单体、hcred-tandem、hcredl、asred2、eqfp611、mraspberry、mstrawberry和jred)、橙色荧光蛋白(例如,morange、mko、kusabira-橙色、单体kusabira-橙色、mtangerine和tdtomato),以及可以通过流式细胞术方法检测到细胞中存在的任何其它合适的荧光蛋白。
[0088]“包括(comprising)”或“包含(including)”一个或多个所列举的元件的组合物或方法可以包含其它未具体列举的元件。例如,“包括”或“包含”蛋白质的组合物可以单独含有蛋白质或与其它成分组合的蛋白质。过渡短语“基本上由
……
组成”意指权利要求的范围应被解释为涵盖权利要求中所列举的指定要素以及对要求保护的发明的基本和新颖特性没有实质性影响的那些要素。因此,当在本发明的权利要求中使用时,术语“基本上由
……
组成”不应被解释为等效于“包括”。
[0089]“任选的(optional)”或“任选地(optionally)”是指随后描述的事件或情况可能发生或可能不发生并且此描述包含其中所述事件或情况发生的实例以及其中所述事件或情况不发生的实例。
[0090]
数值范围的指定包含所述范围内或定义所述范围的所有整数以及由所述范围内的整数定义的所有子范围。
[0091]
除非从上下文中明显看出,否则术语“约”涵盖规定值的标准测量误差范围(例如,sem)内的值。
[0092]
术语“和/或”是指并且涵盖关联的所列项中的一个或多个所列项的任何和所有可能组合以及在以替代性方案(“或”)解释时组合的缺少。
[0093]
术语“或”是指特定列表中的任何一个成员,并且还包含所述列表成员的任何组合。
[0094]
除非上下文另外明确指明,否则本文中的单数形式“一个(a)”、“一种(an)”和“所述(the)”包含复数个提及物。例如,术语“蛋白质”或“至少一种蛋白质”可以包含多种蛋白质,包含其混合物。
[0095]
除非另外指示,否则统计学上显著意指p≤0.05。
具体实施方式
[0096]
i.概述
[0097]
提供了在同一脂质纳米颗粒中一起包含crispr/cas协同激活介体系统组分的脂质纳米颗粒,以及使用此类脂质纳米颗粒来增加体内和离体靶基因表达和评估crispr/cas协同激活介体系统增加体内和离体靶基因表达的能力的方法。
[0098]
crispr/cas9,一种rna导向的dna核酸内切酶,催化dna在其向导rna的结合位点处形成双链断裂。在cas9中已经鉴别出了两个重要的催化域:ruvc和hnh域。ruvc域启动与向导rna不互补的dna链的切割,并且hnh域切割与向导rna互补的dna链。任一域均可以失活以使cas9成为切口酶,或者两个域均可以突变以形成催化上死亡的cas9(dcas9)。尽管dcas9不会引起链断裂,但催化上死亡的蛋白可用于将其它蛋白运送到特定的基因组区域。这是
crispr/cas9系统激活和抑制变体的基础。
[0099]
在dcas9协同激活介体(sam)系统中,数个激活域相互作用以引起比任一个因素单独诱导的更大的基因响应。在该系统的初始迭代中,需要引入三种慢病毒。第一慢病毒将含有直接融合到vp64域的dcas9,vp64域是由单纯疱疹病毒蛋白16的四个串联拷贝组成的转录激活子。当vp64与在转录起始位点附近结合的蛋白融合时,它会充当强转录激活子。第二慢病毒将引进与两种附加激活转录因子融合的ms2外壳蛋白(mcp),这两种激活转录因子为:热休克因子1(hsf1)和转录因子65(p65)。mcp自然地与ms2茎环结合。在该系统中,mcp与工程化到crispr相关联sgrna中的ms2茎环相互作用,并且从而将结合的转录因子运送到适当的基因组位置。第三慢病毒将引入含有ms2环的sgrna。虽然三组分系统在细胞培养中具有一定的灵活性,但这种设置在动物模型中不太理想。
[0100]
腺相关联病毒(aav)通常被认为对基因治疗是安全的,因为它们具有低免疫原性并且具有高度可预测的整合位点(人染色体19上的aavs1)。然而,为了提高它们作为基因治疗载体的安全性,wt aav的整合能力已被消除,使得这些载体作为附加体保留在宿主细胞核中。在引入宿主中后,针对aav的免疫响应通常仅限于中和抗体,而没有明确定义的细胞毒性响应。在分裂细胞中,aav dna通过细胞分裂被稀释,使得需要施用更多的病毒来持续治疗响应。这些随后的暴露可能会导致病毒的快速中和,并且因此降低宿主反应。为了解决这个问题,研究人员将使用替代血清型进行循序感染,尽管这受到血清型特异性的阻碍。基于aav的疗法的另一个问题是相对较小的克隆容量:两个反向末端重复序列之间的4.6kb。由于dcas9 sam的完整编码序列为~5.8kb(无启动子),并非所有sam组分都可以从单个aav中表达。
[0101]
解决这个问题的一种方法是在两个或更多个aav中表达所述元件,并希望它们都感染同一细胞。然而,这对于治疗解决方案来说不太理想。考虑到这一点,我们着手优化该系统,使得其可以进行临床翻译。
[0102]
脂质纳米颗粒(lnp)是aav的一种有吸引力的替代品,因为它们通过利用内源性内吞机制经由ldl受体引进分子,从而安全并且有效地将核酸递送至细胞。一旦被引入生物体,调配物的变化可能会影响颗粒的稳定性和趋向性。此外,各种配体的缀合还可以提高lnp的靶特异性。这种递送方法的一个警告是对宿主细胞的瞬时作用,因为递送至细胞的mrna在一些情况下可以在细胞摄入的48小时内被清除。然而,对lnp递送没有免疫响应,这允许良好耐受的循序给药。此外,在具有催化活性的cas9的情况下,具有催化活性的cas9和sgrna的递送将使靶序列产生永久性的变化,其可能在材料已经从细胞中清除后长时间繁殖。然而,用无催化活性的cas9(催化上死亡的cas9或dcas9)进行的转录激活不会导致永久性的遗传变化。此外,该递送系统在递送具有稳定末端修饰的dcas9 sam向导rna方面的应用受到rna合成技术的限制。这些限制已经妨碍了具有稳定末端修饰的sam sgrna的生成,因为这些分子大于110个核苷酸的平台最大值。
[0103]
虽然经由递送lnp配制的sam grna上调靶基因预期会持续显著更短的时间,但令人惊讶的是,我们能够使用lnp介导的递送实现显著的转录激活,其瞬时性远低于预期。sam sgrna与所有其它sam组分的lnp递送是对治疗性dcas9 sam应用的显著增强,因为我们现在可以(1)确保dcas9 sam转录物和sam sgrna着陆在同一细胞中,(2)用调配物/配体掺入介导增加的组织特异性,(3)向生物体重新给药而不必担心免疫响应,以及(4)生成更稳定的
表达水平。总之,核酸递送的这种组合以安全且出乎意料的稳定方式极大地增强了潜在的dcas9应用。
[0104]
ii.增加靶基因的转录或表达和评估crispr/cas增加体内或离体靶基因的转录或表达的能力的方法
[0105]
提供了使用本文所述的脂质纳米颗粒(lnp)来增加或激活靶基因的表达或转录或者评估本文所述的crispr/cas协同激活介体(sam)系统增加/激活体内或离体靶基因的表达或转录的能力的各种方法。这些方法和组合物可以用于增加真核基因组、细胞或生物体中靶基因的转录或表达。此类lnp在同一lnp中一起包含协同激活介体系统的所有组分(一个或多个向导rna或核酸编码、嵌合cas蛋白或核酸编码,以及嵌合衔接蛋白或核酸编码)。例如,此类方法可以包含向细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中引入:(a)编码嵌合规律间隔成簇短回文重复序列(crispr)相关联(cas)蛋白的核酸,该蛋白包含与一个或多个转录激活域融合的无核酸酶活性的cas蛋白;(b)编码嵌合衔接蛋白的核酸,该嵌合衔接蛋白包含与一个或多个转录激活域融合的衔接蛋白;以及(c)一个或多个向导rna或编码该一个或多个向导rna的一个或多个核酸,每个向导rna包含嵌合衔接蛋白可特异性结合的一个或多个衔接结合元件,并且其中一个或多个向导rna中的每一个能够与cas蛋白形成复合物并将其导向至靶基因内的靶序列,从而增加靶基因的表达,其中所有三种组分在同一lnp中一起递送。在一个实例中,引入编码嵌合cas蛋白和嵌合衔接蛋白两者的多顺反子或双顺反子核酸(例如,dna或mrna)(本文称为sam盒或sam mrna)。例如,如本文别处更详细描述的那样,编码嵌合cas蛋白的序列和编码嵌合衔接蛋白的序列可以通过编码2a蛋白的序列链接。引入真核生物体是指用于将组分递送至真核生物体以使它们能进入一个或多个细胞和这些细胞内的靶基因的任何方法。同样地,引入细胞是指用于将组分递送至细胞中以使它们能进入细胞内的靶基因的任何方法。合适的嵌合cas蛋白、嵌合衔接蛋白和向导rna在本文别处更详细地描述。一个或多个向导rna可与嵌合cas蛋白和嵌合衔接蛋白形成复合物并将它们导向至一个或多个靶基因内的靶序列,从而增加一个或多个靶基因的表达。此类方法还可以包含评估一个或多个靶基因的表达或转录。
[0106]
提供用于增加或激活靶基因的表达或转录或者评估crispr/cas sam系统增加/激活体内靶基因的表达或转录的能力的各种方法也可以用于增加或激活靶基因的表达或转录或者评估crispr/cas sam系统在细胞中增加/激活离体靶基因的表达或转录的能力。提供用于增加或激活靶基因的表达或转录或者评估crispr/cas sam系统增加/激活体内靶基因的表达或转录的能力的各种方法也可以用于增加或激活靶基因的表达或转录或者评估crispr/cas sam系统在细胞中增加/激活体外靶基因的表达或转录的能力。
[0107]
在一些方法中,细胞或生物体可以循序用相同的脂质纳米颗粒重新给药两次或更多次。例如,可以循序将脂质纳米颗粒引入细胞或生物体中至少约2、至少约3、至少约4、至少约5、至少约6、至少约7、至少约8、至少约9,或至少约10次。脂质纳米颗粒的给药之间的间隔可以是任何合适的时间量。例如,间隔可以是至少约1天、至少约2天、至少约3天、至少约4天、至少约5天、至少约6天、至少约7天、至少约1周、至少约2周、至少约3周、至少约4周、至少约5周、至少约6周、至少约7周、至少约8周、至少约1个月、至少约2个月、至少约3个月或至少约4个月。例如,脂质纳米颗粒的给药之间的间隔可以是至少约1周(例如,约1周)、至少约2周(例如,约2周)、至少约4周(例如,约4周)、约1周至约5周、约1周至约4周、约1周至约3周、
约1周至约2周、约2周至约5周、约2周至约4周、约2周至约3周、约3周至约5周、约3周至约4周,或约4周至约5周。在一个实例中,脂质纳米颗粒的给药之间的间隔可以是约2周。
[0108]
在一些方法中,在每次循序引入脂质纳米颗粒后,靶基因的表达增加至至少约相同的水平。在一些方法中,靶基因的表达通过循序重新给药维持在约相同的水平。在一些方法中,用脂质纳米颗粒重新给药后靶基因的表达增加至比单次给药脂质纳米颗粒更高的水平(例如,重新给药脂质纳米颗粒时的靶基因表达的增加高于没有重新给药时的靶基因表达的增加)。
[0109]
任选地,可以引入两个或更多个向导rna,它们各自被设计为靶向靶基因内的不同向导rna靶序列。例如,可以设计两个或更多个、三个或更多个、四个或更多个,或五个或更多个向导rna以靶向单个靶基因(例如,可以使用两个、三个、四个或五个向导rna,每个向导rna靶向相同靶基因内的不同向导rna靶序列)。替代地或附加地,可以引入两个或更多个、三个或更多个、四个或更多个,或五个或更多个向导rna,每个向导rna被设计为靶向不同靶基因(例如,两个或更多个、三个或更多个、四个或更多个,或五个或更多个不同靶基因)中的不同向导rna靶序列(即,多路复用)。例如,可以使用两个、三个、四个或五个向导rna,每个向导rna靶向不同的靶基因。
[0110]
嵌合cas蛋白、嵌合衔接蛋白和向导rna可以以任何形式(向导rna的dna或rna;嵌合cas蛋白和嵌合衔接蛋白的dna、rna或蛋白)经由本文别处公开的任何施用途径引入细胞或真核生物体。在一些方法中向导rna、嵌合cas蛋白和嵌合衔接蛋白可以以组织特异性方式引入(例如,以肝脏特异性方式引入)。
[0111]
编码嵌合cas蛋白和嵌合衔接蛋白的向导rna和mrna(例如,sam mrna)可以在5'末端和/或3'末端包含一个或多个稳定末端修饰,如本文别处更详细描述的那样。作为一个实例,rna的5'末端和/或3'末端可以包含一个或多个硫代磷酸酯键。例如,向导rna可以包含向导rna的5'或3'末端处的2、3或4个末端核苷酸之间的硫代磷酸酯键。作为另一个实例,rna的5'末端和/或3'末端可以包含一个或多个2'-o-甲基修饰。例如,rna可以在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。例如,向导rna可以在rna的5'和/或3'末端(例如,5'末端)处的2个、3个或4个末端核苷酸处包括2'-o-甲基修饰。参见例如wo 2017/173054 a1和finn等人(2018),《细胞报告(cell rep.)》22(9):2227-2235,所述文献中的每个文献出于所有目的通过引用整体并入本文。作为另一个实例,rna(例如,mrna)可以在5'末端被加帽(例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构),可以被聚腺苷酸化,并且还可以任选地被修饰以被假尿苷完全取代(即,所有标准尿嘧啶残基均被假尿苷替换,假尿苷是一种尿嘧啶与碳-碳键而不是氮-碳键相连的尿苷异构体)。对向导rna和mrna的其它可能的修饰在本文别处更详细地描述。在具体实例中,rna在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。此类化学修饰可以例如为rna提供更大的稳定性和免受核酸外切酶,允许rna在细胞内比未经修饰的rna持续更长时间。例如,此类化学修饰还可以防止先天的细胞内免疫应答,所述先天的细胞内免疫应答可以主动降解rna或触发导致细胞死亡的免疫级联反应。
[0112]
向导rna可以靶向靶基因中适合增加靶基因转录的任何位置。例如,向导rna的靶序列可以包含靶基因内的调控序列,如启动子或增强子。同样地,靶序列可以与基因的转录起始位点邻近。例如,靶序列可以在转录起始位点1000、900、800、700、600、500、400、300、
200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内、在转录起始位点上游1000、900、800、700、600、500、400、300、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内,或者在转录起始位点下游1000、900、800、700、600、500、400、300、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内。作为具体实例,靶序列可以在靶基因的转录起始位点约200个碱基对内,或者可以在转录起始位点上游约200个碱基对内和转录起始位点下游1个碱基对内。
[0113]
本文公开的方法还可以包含评估靶基因的表达。测量表达或活性的方法将取决于被修饰的靶基因。评估靶基因的增加的转录或表达的方法是众所周知的。
[0114]
例如,如果靶基因包含编码rna或蛋白的基因,则评估表达的方法可以包含测量经编码的rna和/或蛋白的表达或活性。例如,如果经编码的蛋白是释放到血清中的蛋白,则可以测量经编码的蛋白的血清水平。用于测量rna和蛋白的水平和活性的测定法是众所周知的。
[0115]
评估靶基因在真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中的表达可以在来自任何组织或器官的任何细胞类型中进行。例如,可以在来自相同组织或器官的多个细胞类型中或在来自组织或器官内多个位置的细胞中评估靶基因的表达。这可以提供关于靶向靶组织或器官内的哪些细胞类型或者crispr/cas到达组织或器官的哪些部分并修饰它们的信息。作为另一个实例,可以在多个类型的组织或多个器官中评估靶基因的表达。在靶向特定组织或器官的方法中,这可以提供关于靶向该组织或器官的有效性以及其它组织或器官中是否存在脱靶效应的信息。
[0116]
在一些方法中,例如通过评估肝细胞中由靶基因组基因座表达的分泌蛋白的血清水平来评估靶基因在肝细胞中的表达。如果靶基因编码具有特定酶活性的蛋白,则评估可以包含测量靶基因的表达和/或由靶基因编码的蛋白的活性。替代地或附加地,评估可以包含评估从真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)分离的一个或多个细胞中的表达。评估可以包含从真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中分离靶器官或组织,并评估靶基因在靶器官或组织中的表达。评估还可以包含评估靶基因在靶器官或组织内的两种或更多种不同细胞类型中的表达。类似地,评估可以包含从真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)分离非靶器官或组织(例如,两个或更多个非靶器官或组织),并评估靶基因在非靶器官或组织中的表达。
[0117]
在一些方法中,靶基因可以是本文别处所述的疾病相关联基因。作为一个实例,疾病相关联基因可以是与未患病对照的组织或细胞相比,在来源于受疾病影响的组织的细胞中以异常水平或异常形式产生转录或翻译产物的任何基因。它可能是以异常高水平表达的基因,其中改变的表达与疾病的发生和/或发展相关。它可能是以异常低水平表达的基因,其中改变的表达与疾病的发生和/或发展相关。疾病相关联基因也指具有导致疾病病因的突变或遗传变异的基因。转录或翻译的产物可能是已知的或未知的,并且可能处于正常或异常水平。例如,靶基因可以是与蛋白聚集疾病或病症相关联的基因。作为具体的实例,靶基因可以是与蛋白聚集疾病或病症相关联的基因(例如,ttr),并且该方法可以包含增加该靶基因的表达以模拟蛋白聚集疾病或病症。在一些具体的方法中,靶基因可以是ttr。任选地,ttr基因可以包含致病突变(例如,引起淀粉样变性的突变)或致病突变的组合。此类突
变的实例例如在wo 2018/007871中提供,该文献以全文引用的方式并入本文用于所有目的。
[0118]
在一些方法中,靶基因可以是在受试者中该基因产生的增加将是有益的任何基因(例如,疾病相关联基因)。例如,此类靶基因的转录减少、来自此类靶基因的基因产物量减少,或来自此类靶基因的基因产物的活性降低可能与疾病相关联、可能使疾病恶化,或者可能引起疾病,使得靶基因的转录或表达增加将是有益的。此类基因的一个实例是otc(entrez gene id5009)。otc缺乏(鸟氨酸转氨甲酰酶缺乏)的特征是血液中的氨升高,其被认为是一种神经毒素并且可以由高蛋白饮食引起。这是一种x连锁疾病,并且主要影响男性,但由于随机x失活,女性可以发展出较温和的形式。有许多突变会导致疾病的一系列严重程度,因为一些突变仍然允许产生一些野生型otc。作为一个实例,与野生型受试者相比,otc中的突变剪接位点可导致受试者具有约5%的otc酶活性。正是这些患者以及有症状的女性将受益于通过递送sam mrna和靶向otc启动子的向导rna来增加otc的表达,因为野生型otc的表达增加将允许清除血液中过量的氨。增加基因的产生对将受试者有益的其它基因实例包括用于增加胎儿血红蛋白表达的hbg1(entrez gene id 3047)和hbg2(entrez gene id 3048)。增加基因的产生对将受试者有益的其它基因实例包括单倍体不足基因。单倍体不足是当基因的一个拷贝失活或缺失,并且该基因的剩余功能拷贝不足以产生所需的基因产物以保持正常功能时发生的情况。换句话说,对于一些基因,二倍体基因组中一个功能拷贝的缺失或失活会将生物体的表型改变为异常或疾病状态。这些基因被称为单倍体不足,因为这些基因的一个正常拷贝不足以产生正常或野生型表型。单倍体不足基因的一个功能拷贝的丢失与包括神经障碍和智力迟钝在内的疾病有关,并且单倍体不足基因还可以影响一个人对疾病和/或药物副作用的易感性。表2和3中提供了单倍体不足基因和与一个功能拷贝的丢失相关联的相关联疾病/病症/综合征的实例。另见dang等人(2008)《欧洲人类遗传学杂志(eur.j.hum.genet.)》,16(11):1350-1357,该文献以全文引用的方式并入本文用于所有目的。
[0119]
表2:表3中单倍体不足基因表达的实例的子组。
[0120][0121]
表3:单倍体不足基因表达的实例。
[0122]
[0123]
[0124]
[0125]
[0126]
[0127]
[0128]
[0129]
[0130][0131]
可以实现靶基因表达的任何统计学上显著的增加。例如,相对于对照真核基因组、细胞或生物体(例如,在rna水平或蛋白水平上测量),靶基因表达的增加可以为至少约0.5倍、至少约1倍、至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍或至少约20倍。同样地,靶基因表达增加的持续时间可以是任何合适的时间。例如,靶基因表达增加的持续时间可以为至少约1天、至少约2天、至少约3天、至少约4天、至少约5天、至少约6天、至少约1周、至少约2周、至少约3周、至少约4周、至少约1个月或至少约2个月。在具体实例中,相对于对照真核基因组、细胞或生物体,在引入
crispr/cas协同激活介体(sam)系统之后的1天、2天、3天、4天、5天、6天、1周、2周、3周、4周或1个月时靶基因表达的增加可以为至少约0.5倍、至少约1倍、至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍或至少约20倍。在一个实例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约2倍。在另一个实例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约3倍。在另一个实例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约4倍。在另一个实例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约5倍。在另一个实例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约6倍。在另一个实施例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约7倍。在另一个实施例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约8倍。在另一个实施例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约9倍。在另一个实施例中,在使用0.5mg/kg lnp或1mg/kg lnp或2mg/kg lnp的剂量1、2或3周后增加为至少约10倍。靶基因表达的增加可以以接近恒定的水平(即,不显示随时间降低的模式)持续至少约1天、至少约2天、至少约3天、至少约4天、至少约5天、至少约6天、至少约1周、至少约2周、至少约3周、至少约4周或更长。
[0132]
该方法可以用于增加任何真核基因组、细胞或生物体中靶基因的转录或表达。基因组、细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)可以是雄性或雌性。在一些方法中,靶基因的转录或表达在有需要的受试者(例如,生物体或动物或哺乳动物,如人)中增加。例如,有需要的受试者可以是患有与以下因素相关联、被以下因素恶化,或由以下因素引起的疾病、病症或综合征的受试者:靶基因的转录或表达减少、靶基因的基因产物量减少,或靶基因的基因产物的活性降低;使得增加靶基因的转录或表达,增加靶基因的基因产物量,或增加靶基因的基因产物的活性将是有益的。相对于没有疾病、病症或综合征的对照受试者,靶基因在受试者中可能低表达或低水平表达。例如,增加靶基因的转录或表达、增加靶基因的基因产物量或增加靶基因的基因产物的活性可以治疗受试者的疾病、病症或综合征。此类疾病、病症或综合征的实例包括与单倍体不足相关联的疾病、病症或综合征。增加转录或表达将对其有益的单倍体不足基因和其它基因的实例在表2和3中以及本文别处提供。
[0133]
本文提供的真核基因组、细胞或生物体可以是例如多细胞真核生物体、非人真核生物体、动物、非人动物、哺乳动物、非人哺乳动物、人、非人、啮齿动物、小鼠或大鼠基因组、细胞或生物体。真核细胞包括例如真菌细胞(例如,酵母)、植物细胞、动物细胞、哺乳动物细胞、非人哺乳动物细胞和人细胞。术语“动物”包含哺乳动物、鱼类和鸟类。哺乳动物包含例如人、非人灵长类动物、猴子、猿、猫、狗、马、公牛、鹿、野牛、绵羊、兔、啮齿动物(例如,小鼠、大鼠、仓鼠和豚鼠)和家畜(例如,牛种,如奶牛和公牛;绵羊种,如绵羊和山羊;以及猪种,如猪和野猪)。鸟类包含例如鸡、火鸡、鸵鸟、鹅和鸭。还包含家养动物和农业动物。术语“非人动物”不包含人。
[0134]
细胞也可以呈任何类型的未分化或分化状态。例如,细胞可以是全能细胞、多能细胞(例如,人多能细胞或非人多能细胞,如小鼠胚胎干(es)细胞或大鼠es细胞)或非多能细胞。全能细胞包含可以产生任何细胞类型的未分化细胞,并且多能性细胞包含具有发育成
超过一种分化细胞类型的能力的未分化细胞。此类多能和/或全能细胞可以是例如es细胞或es样细胞,如诱导多能干(ips)细胞。es细胞包含在引入到胚胎中时能够对发育胚胎的任何组织做出贡献的胚胎源性全能或多能性细胞。es细胞可以源自囊胚的内细胞团,并且能够分化成三种脊椎动物胚层(内胚层、外胚层和中胚层)中的任何层的细胞。
[0135]
人多能细胞的实例包括人es细胞、人成体干细胞、发育受限的人祖细胞和人诱导的多能干(ips)细胞,如致敏人ips细胞和幼稚人ips细胞。诱导的多能干细胞包括可以直接衍生自分化的成体细胞的多能干细胞。人ips细胞可以通过将特定组的重编程因子引入细胞中产生,这些重编程因子可以包括例如oct3/4、sox家族转录因子(例如,sox1、sox2、sox3、sox15)、myc家族转录因子(例如,c-myc、l-myc、n-myc)、kr
ü
ppel样家族(klf)转录因子(例如,klf1、klf2、klf4、klf5)和/或相关转录因子,如nanog、lin28和/或glis1。人ips细胞也可以例如通过使用mirna、模拟转录因子作用的小分子或细胞谱系特异性分子产生。人ips细胞的特征在于它们分化成三种脊椎动物胚层(例如内胚层、外胚层或中胚层)的任何细胞的能力。人ips细胞的特征还在于它们在合适的体外培养条件下无限增殖的能力。参见例如takahashi和yamanaka(2006)《细胞(cell)》126:663-676,该文献以全文引用的方式并入本文用于所有目的。致敏人es细胞和致敏人ips细胞包括表达与植入后外胚层细胞的特征类似的特征并用于谱系特异性和分化的细胞。幼稚人es细胞和幼稚人ips细胞包括表达与植入前胚胎的内细胞团的es细胞的特征类似的特征并且不用于谱系特异性的细胞。参见例如nichols和smith(2009)《细胞干细胞(cell stem cell)》4:487-492,该文献以全文引用的方式并入本文用于所有目的。
[0136]
本文所提供的细胞还可以是生殖细胞(例如,精子或卵母细胞)。细胞可以是有丝分裂感受态细胞或有丝分裂非活性细胞、减数分裂感受态细胞或减数分裂非活性细胞。相似地,细胞还可以是初生体细胞或不是初生体细胞的细胞。体细胞包含任何不是配子、生殖细胞、配子母细胞或未分化干细胞的细胞。例如,细胞可以是肝细胞、肾细胞、造血细胞、内皮细胞、上皮细胞、成纤维细胞、间充质细胞、角质形成细胞、血细胞、黑素细胞、单核细胞、单个核细胞、单核细胞前体、b细胞、红细胞-巨核细胞、嗜酸性粒细胞、巨噬细胞、t细胞、胰岛β细胞、外分泌细胞、胰腺祖细胞、内分泌祖细胞、脂肪细胞、前脂肪细胞、神经元、神经胶质细胞、神经干细胞、神经元、成肝细胞、肝细胞、心肌细胞、骨骼肌细胞、平滑肌细胞、导管细胞、腺泡细胞、α细胞、β细胞、δ细胞、pp细胞、胆管细胞、白色或棕色脂肪细胞或眼细胞(例如,小梁网细胞、视网膜色素上皮细胞、视网膜微血管内皮细胞、视网膜周细胞、结膜上皮细胞、结膜成纤维细胞、虹膜色素上皮细胞、角膜细胞、晶状体上皮细胞、非色素睫状上皮细胞、眼脉络膜成纤维细胞、感光细胞、神经节细胞、双极细胞、水平细胞或无长突细胞)。例如,细胞可以是肝细胞(liver cell),如成肝细胞或肝细胞(hepatocyte)。
[0137]
本文所提供的合适的细胞还包含原代细胞。原代细胞包含直接从生物体、器官或组织中分离的细胞或细胞培养物。原代细胞包含既不转化也不永生的细胞。所述原代细胞包含从生物体、器官或组织中获得的任何细胞,所述细胞先前未在组织培养物中进行传代,或者先前已经在组织培养物中进行传代但不能无限期地在组织培养中进行传代。此类细胞可以通过常规技术分离并且包含例如体细胞、造血细胞、内皮细胞、上皮细胞、成纤维细胞、间充质细胞、角质形成细胞、黑素细胞、单核细胞、单一核细胞、脂肪细胞、前脂肪细胞、神经元、神经胶质细胞、肝细胞、骨骼肌成肌细胞和平滑肌细胞。例如,原代细胞可以来源于结缔
组织、肌肉组织、神经系统组织或上皮组织。此类细胞可以通过常规技术分离并且包含例如肝细胞。
[0138]
本文所提供的其它合适的细胞包含永生化细胞。永生化细胞包含来自多细胞生物体的细胞,其通常不会无限增殖但由于突变或改变而逃避正常细胞衰老并且替代地可以继续进行分裂。此类突变或改变可以天然存在或有意诱导。永生化细胞的实例包括中国仓鼠卵巢(cho)细胞、人胚胎肾细胞(例如,hek 293细胞或293t细胞)和小鼠胚胎成纤维细胞(例如,3t3细胞)。永生化细胞系的具体实例是hepg2人肝癌细胞系。多种类型的永生化细胞是众所周知的。永生化或原代细胞包含通常用于培养或表达重组基因或蛋白质的细胞。
[0139]
本文所提供的细胞还包含单细胞期胚胎(即,受精的卵母细胞或受精卵)。此类单细胞期胚胎(例如,啮齿动物单细胞期胚胎)可以来自任何遗传背景(例如,对于小鼠,balb/c、c57bl/6、129或其组合),可以是新鲜的或冷冻的,并且可以源自自然育种或体外受精。
[0140]
本文所提供的细胞可以是正常的、健康的细胞,或者可以是患病或携带突变体的细胞。
[0141]
真核基因组、细胞或生物体可以来自任何遗传背景。例如,合适的小鼠可以来自129品系、c57bl/6品系、129和c57bl/6的混合、balb/c品系或swiss webster品系。129品系的实例包含129p1、129p2、129p3、129x1、129s1(例如,129s1/sv,129s1/svlm)、129s2、129s4、129s5、129s9/svevh、129s6(129/svevtac)、129s7、129s8、129t1和129t2。参见例如festing等人(1999)《哺乳动物基因组(mamm.genome)》10(8):836,所述文献出于所有目的通过引用整体并入本文。c57bl品系的实例包含c57bl/a、c57bl/an、c57bl/grfa、c57bl/kal_wn、c57bl/6、c57bl/6j、c57bl/6byj、c57bl/6nj、c57bl/10、c57bl/10scsn、c57bl/10cr和c57bl/ola。合适的小鼠还可以来自上述129品系和上述c57bl/6品系的混合(例如,50%129和50%c57bl/6)。同样地,合适的小鼠可以来自上述129品系的混合或上述bl/6品系的混合(例如,129s6(129/svevtac)品系)。
[0142]
相似地,大鼠可以来自任何大鼠品系,包含例如aci大鼠品系、黑刺鼠(da)大鼠品系、威斯塔(wistar)大鼠品系、lea大鼠品系、斯泼累格多雷(sprague dawley,sd)大鼠品系或费舍尔(fischer)大鼠品系,如费舍尔f344或费舍尔f6。大鼠还可以从源自上述两种或更多种品系的混合品系中获得。例如,合适的大鼠可以来自da品系或aci品系。aci大鼠品系的特征在于具有腹部和足部呈白色的黑刺鼠以及rt1
av1
单倍型。此类品系可从多种来源获得,包含哈兰实验室(harlan laboratories)。黑刺鼠(da)大鼠品系的特征在于具有刺鼠皮毛和rt1
av1
单倍型。此类大鼠可从多种来源获得,包含查尔斯河和哈兰实验室(charles river and harlan laboratories)。在一些情况下,合适的大鼠可以来自近交大鼠品系。参见例如us 2014/0235933,所述文献出于所有目的通过引用整体并入本文。
[0143]
还提供了用于优化crispr/cas sam系统向细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)的递送或优化体内或离体crispr/cas转录激活活性的多种方法。此类方法可以包含例如:(a)在第一真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)或第一细胞中首次如上所述执行测试crispr/cas sam系统增加靶基因转录或表达的能力的方法;(b)改变变量并在具有改变的变量的第二真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物;即,相同物种)或第二细胞中第二次执行该方法;以及(c)比较步骤(a)中靶基因的表达/转录与步骤(b)中靶基因的表达/转录,并选择导致靶基因最
高表达/转录的方法。
[0144]
替代地或附加地,可以选择导致最高功效、最高一致性或最高特异性的方法。更高功效是指靶基因的更高水平的表达/转录(例如,在特定靶细胞类型内、在特定靶组织内,或在特定靶器官内靶向更高百分比的细胞)。更高一致性是指在靶向多于一种类型的细胞、组织或器官的情况下,不同类型的靶向细胞、组织或器官中靶基因的表达/转录的更一致的增加(例如,靶器官内更多数量的细胞类型的表达/转录增加)。如果靶向特定器官,则更高的一致性也可以指在器官内的所有位置中的表达/转录的更一致的增加。更高的特异性可以指对于所靶向的一个或多个靶基因的更高的特异性、对于所靶向的细胞类型的更高的特异性、对于所靶向的组织类型的更高的特异性或者对于所靶向的器官的更高的特异性。例如,增加的靶特异性是指对其它基因的较少脱靶效应(例如,在非预期的脱靶基因组基因座(例如,相邻基因组基因座)处具有增加的转录的较低百分比的靶向细胞,而不是具有增加的靶基因的转录或除了具有增加的靶基因的转录之外)。同样地,增加的细胞类型、组织或器官类型特异性是指在靶向特定的细胞类型、组织类型或器官类型的情况下,脱靶细胞类型、组织类型或器官类型中的较少效应(即,增加的表达/转录)(例如,当靶向特定器官(例如,肝脏)时,器官或组织中不是预期靶标的细胞中的效应较少(即,增加的表达/转录))。
[0145]
被改变的变量可以是任何参数。作为一个实例,改变的变量可以是用于将sam组分(嵌合cas蛋白、嵌合衔接蛋白和向导rna)引入细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)的施用途径。如静脉内、玻璃体内、实质内和鼻内滴注等施用途径的实例在本文其它地方公开。
[0146]
作为另一个实例,改变的变量可以是所引入的sam组分的浓度或量。作为另一个实例,改变的变量可以是引入sam组分的次数或频率(即,引入lnp的次数或频率)。作为另一个实例,改变的变量可以是引入sam组分的形式。例如,向导rna可以以dna或rna的形式引入,并且嵌合cas蛋白和嵌合衔接蛋白可以以dna、rna或蛋白的形式引入。类似地,向导rna或嵌合cas蛋白或嵌合衔接蛋白(或编码此类组分的核酸)可以包含用于稳定性、减少脱靶效应、促进递送等的各种修饰组合。作为另一个实例,改变的变量可以是所引入的向导rna的序列(例如,引入具有不同序列的不同向导rna或靶向不同的向导rna靶序列)。
[0147]
还提供了使用由本文公开的方法生成的真核细胞或生物体来增加或激活靶基因(特别是其过表达与疾病相关联或者成为疾病的病因的靶基因)的表达或转录的方法。此类具有增加的靶基因表达的真核细胞或生物体(靶基因的过表达与疾病相关联或者成为疾病的病因)可用于例如筛选针对疾病的治疗或预防效果或降低靶基因表达的功效的化合物。此类方法可以包含例如,如本文别处所述,增加或激活真核细胞或生物体中靶基因的转录,将试剂或化合物引入真核细胞或生物体,并且然后评估试剂或化合物的活性(例如,与未用试剂或化合物处理的对照真核细胞或生物体相比,在用试剂或化合物处理的真核细胞或生物体中)。评估可以包含例如评估靶基因的表达(例如,在mrna水平或在蛋白水平),其中靶基因表达的降低可以指示治疗或预防效果。替代地或附加地,评估可以包含评估与靶基因过表达相关联或由靶基因过表达引起的疾病的一个或多个体征或症状,其中体征或症状的存在的减少或改善可以指示治疗或预防效果.如果筛选的试剂或化合物显示出治疗或预防效果,则可以选择其作为候选治疗或预防试剂或化合物。
[0148]
还提供了用于在有需要的受试者中增加或激活靶基因的表达或转录的方法,其中
靶基因的表达或活性降低与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。例如,此类方法可以用于增加或激活靶基因(特别是其低表达与疾病或病症相关联或成为疾病或病症的病因,或者与对疾病或病症或药物的副作用的易感性相关联或成为对疾病或病症或药物的副作用的易感性的病因的靶基因)的表达或转录。例如,靶基因可以是在受试者中低表达或低水平表达的基因,并且低表达或低水平表达与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因。此类靶基因的转录减少、来自此类靶基因的基因产物量减少,或来自此类靶基因的基因产物的活性降低可能与疾病相关联、可能使疾病恶化,或者可能引起疾病,使得靶基因的转录或表达增加将是有益的。此类基因的一个实例是otc(entrez gene id 5009)。此类基因的其它实例是hbg1(entrez gene id 3047)和hbg2(entrez gene id 3048)。此类基因的其它实例包括单倍体不足基因,如表2和3中的那些。受试者可以是例如靶基因表达或活性降低的受试者,如患有与单倍体不足相关联的疾病、病症或综合征的受试者。
[0149]
iii.crispr/cas协同激活介体(sam)系统
[0150]
本文公开的方法和组合物(例如脂质纳米颗粒)利用规律间隔成簇短回文重复序列(crispr)/基于crispr相关联(cas)的协同激活介体(sam)系统,用于在体内或离体激活靶基因转录的方法,和评估sam系统或此类系统的组分(例如,向导rna)在体内或离体激活靶基因组基因座转录的能力。本文所述的sam系统组分全部在同一脂质纳米颗粒中一起递送并且包含如本文别处所述的嵌合cas蛋白、嵌合衔接蛋白和向导rna以激活靶基因的转录。嵌合cas蛋白(例如,嵌合cas蛋白,如嵌合cas9蛋白,如嵌合酿脓链球菌cas9蛋白、嵌合空肠弯曲杆菌cas9蛋白或嵌合金黄色葡萄球菌cas9蛋白(例如,衍生自酿脓链球菌cas9蛋白、空肠弯曲杆菌cas9蛋白,或金黄色葡萄球菌cas9蛋白的嵌合cas9蛋白)和嵌合衔接蛋白(例如,包含与向导rna内的衔接结合元件特异性结合的衔接蛋白和一个或多个异源转录激活域)在本文别处更详细地描述。在一个实例中,嵌合cas蛋白和嵌合衔接蛋白在单个多顺反子或双顺反子核酸(例如,dna或mrna)(称为sam盒或sam mrna)中递送。例如,如本文别处更详细描述的那样,编码嵌合cas蛋白的序列和编码嵌合衔接蛋白的序列可以通过编码2a蛋白的序列链接。在具体实例中,嵌合cas蛋白(例如,nls-cas9-nls-vp64,其中,例如,5'nls是单部分的并且3'nls是双部分的)可以作为多顺反子或双顺反子mrna(例如,体外转录的mrna)提供,其也编码嵌合衔接蛋白(例如,ms2(mcp)-nls-p65-hsf1)。编码嵌合cas蛋白和嵌合衔接蛋白的核酸可以通过编码2a蛋白的核酸链接。作为一个实例,mrna从5'到3'可以包含:nls-cas9-nls-vp64-2a-ms2(mcp)-nls-p65-hsf1。mrna可以在5'末端被加帽(例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构),可以被聚腺苷酸化(聚(a)尾),并且还可以任选地被修饰为被假尿苷完全取代。
[0151]
crispr/cas系统包含转录本和涉及cas基因表达或引导其活性的其它元件。crispr/cas系统可以是例如i型、ii型、iii型系统或v型系统(例如,v-a亚型或v-b亚型)。在本文所公开的组合物和方法中使用的crispr/cas系统可以是非天然存在的。“非天然存在的”系统包含表明涉及人工的任何事物,如系统的一种或多种组分从其自然存在的状态改变或突变,至少基本上不含所述组分在自然界中与其天然相关的至少一种其它组分或与所述组分不与其天然相关的至少一种其它组分相关。例如,一些crispr/cas系统采用包括非天然一起存在的grna和cas蛋白的非天然存在的crispr复合物,采用非天然存在的cas蛋
白,或者采用非天然存在的grna。
[0152]
本文公开的方法和组合物通过使用或测试crispr复合物(包括与嵌合cas蛋白和嵌合衔接蛋白复合的向导rna(grna))在体内诱导靶基因组基因座的转录激活的能力来使用crispr/cas系统。
[0153]
a.嵌合cas蛋白
[0154]
提供了可以结合本文别处公开的向导rna以激活靶基因的转录的嵌合cas蛋白。此类嵌合cas蛋白可以包含:(a)dna结合域,其是能够与向导rna形成复合物并与靶序列结合的规律间隔成簇短回文重复序列(crispr)相关联(cas)蛋白或其功能片段或变体;和(b)一个或多个转录激活域或其功能片段或变体。例如,此类融合蛋白可以包含1、2、3、4、5或更多个转录激活域(例如,两个或更多个异源转录激活域或三个或更多个异源转录激活域)。在一个实例中,嵌合cas蛋白可以包含无催化活性的cas蛋白(例如,dcas9)和vp64转录激活域或其功能片段或变体。例如,此类嵌合cas蛋白可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:1所示的dcas9-vp64嵌合cas蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。然而,还提供了其中转录激活域包含其它转录激活域或其功能片段或变体和/或其中cas蛋白包含其它cas蛋白(例如,无催化活性的cas蛋白)的嵌合cas蛋白。其它合适的转录激活域的实例在本文别处提供。
[0155]
转录激活结构域可以位于n端处、c端处或cas蛋白内的任何位置处。例如,当与酿脓链球菌cas9蛋白最佳比对时,转录激活域可以连接至酿脓链球菌cas9蛋白的rec1域、rec2域、hnh域或pi域,或直系同源cas9蛋白或同源或直系同源cas蛋白的任何对应区域。例如,转录激活域可以连接到位置553的rec1域、位置575的rec1域、位置175-306内的任何位置的rec2域或替换位置175-306内的部分或整个区域、位置715-901内的任何位置的hnh域或替换位置715-901内的部分或整个区域,或酿脓链球菌cas9蛋白的位置1153的pi域。参见例如wo 2016/049258,该文献以全文引用的方式并入本文用于所有目的。如本文别处所述,转录激活域可以在一侧或两侧侧接一个或多个接头。
[0156]
嵌合cas蛋白也可以与附加的异源多肽可操作地链接或融合。融合或链接的异源多肽可以位于n端、c端或嵌合cas蛋白内部的任何位置。例如,嵌合cas蛋白还可以包含核定位信号。合适的核定位信号和对cas蛋白的其它修饰的实例在本文别处更详细地描述。
[0157]
嵌合cas蛋白可以以任何形式提供。例如,嵌合cas蛋白可以以蛋白的形式提供,如与grna复合的嵌合cas蛋白。替代地,可以以编码嵌合cas蛋白的核酸的形式提供嵌合cas蛋白,如rna(例如,信使rna(mrna))或dna。在具体实例中,嵌合cas蛋白可以作为mrna(例如,体外转录的mrna)提供,如也编码嵌合衔接蛋白的多顺反子或双顺反子mrna。任选地,可以对编码嵌合cas蛋白的核酸进行密码子优化以在特定细胞或生物体中有效翻译成蛋白。例如,与天然存在的多核苷酸序列相比,可以修饰编码嵌合cas蛋白的核酸以取代在真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、大鼠细胞或任何其它感兴趣的宿主细胞中具有更高使用频率的密码子。当编码嵌合cas蛋白的核酸被引入到细胞中时,嵌合cas蛋白可以在细胞中进行瞬时、条件性或组成性表达。
[0158]
可以修饰作为mrna提供的嵌合cas蛋白以提高稳定性和/或免疫原性特性。可以对
mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合cas蛋白的mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统中的模式识别受体的激活)。编码嵌合cas蛋白的mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合cas蛋白的mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合cas mrna。同样地,嵌合cas mrna可以通过使用同义密码子消耗尿苷来修饰。其它可能的修饰在本文别处更详细地描述。
[0159]
可以修饰作为mrna提供的嵌合cas蛋白以提高稳定性和/或免疫原性特性。可以对mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合cas蛋白的mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统中的模式识别受体的激活)。编码嵌合cas蛋白的mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合cas蛋白的mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合cas mrna。同样地,嵌合cas mrna可以通过使用同义密码子消耗尿苷来修饰。
[0160]
嵌合cas mrna可以在至少一个、多个或所有尿苷位置包含经修饰的尿苷。经修饰的尿苷可以是在5位置被修饰(例如,用卤素、甲基或乙基)的尿苷。经修饰的尿苷可以是在1位置被修饰(例如,用卤素、甲基或乙基)的假尿苷。经修饰的尿苷可以是例如假尿苷、n1-甲基-假尿苷、5-甲氧基尿苷、5-碘尿苷或其组合。在一些实例中,经修饰的尿苷是5-甲氧基尿苷。在一些实例中,经修饰的尿苷是5-碘尿苷。在一些实例中,经修饰的尿苷是假尿苷。在一些实例中,经修饰的尿苷是n1-甲基-假尿苷。在一些实例中,经修饰的尿苷是假尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是n1-甲基假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-碘尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和5-甲氧基尿苷的组合。
[0161]
本文公开的嵌合cas mrna还可以包含5'帽,如cap0、cap1或cap2。5'帽通常是通过5'-三磷酸酯链接到mrna的5'至3'链的第一核苷酸(即,第一近帽核苷酸)的5'位置的7-甲基鸟嘌呤核糖核苷酸(其可以被进一步修饰,例如,关于arca)。在cap0中,mrna的第一和第二近帽核苷酸的核糖均包含2'-羟基。在cap1中,mrna的第一和第二转录核苷酸的核糖分别包含2'-甲氧基和2'-羟基。在cap2中,mrna的第一和第二近帽核苷酸的核糖均包含2'-甲氧基。参见例如katibah等人(2014)《美国科学院院报(proc.natl.acad.sci.u.s.a.)》111(33):12025-30和abbas等人(2017)《美国科学院院报》114(11):e2106-e2115,它们中的每一个以全文引用的方式并入本文用于所有目的。大多数内源性高等真核mrna,包括哺乳动物mrna,如人mrna,均包含cap1或cap2。cap0和其它不同于cap1和cap2的帽结构可能在哺乳动物(如人类)中具有免疫原性,因为先天免疫系统的组分(如ifit-1和ifit-5)将其识别为非自身,这可能导致细胞因子升高水平(包括i型干扰素)。先天免疫系统的组分如ifit-1和
ifit-5也可能与eif4e竞争mrna与cap1或cap2以外的帽的结合,从而潜在地抑制mrna的翻译。
[0162]
可以共转录地包括帽。例如,arca(抗反向帽类似物;赛默飞世尔科技公司(thermo fisher scientific)商品目录号第am8045号)是包含与鸟嘌呤核糖核苷酸的5'位置链接的7-甲基鸟嘌呤3'-甲氧基-5'-三磷酸酯的帽类似物,其可在体外在起始时掺入转录物中。arca产生cap0帽,其中第一近帽核苷酸的2'位置是羟基。参见例如stepinski等人,(2001)《rna》7:1486-1495,该文献以全文引用的方式并入本文用于所有目的。
[0163]
cleancap
tm ag(m7g(5')ppp(5')(2'omea)pg;trilink生物技术公司商品目录号第n-7113号)或cleancap
tm gg(m7g(5')ppp(5')(2'omeg)pg;trilink生物技术公司商品目录号第n-7133号)可用于共转录地提供cap1结构。3'-o-甲基化版本的cleancap
tm ag和cleancap
tm gg也可从trilink生物技术公司获得,商品目录号分别为第n-7413和第n-7433号。
[0164]
替代地,可以在转录后将帽添加到rna。例如,牛痘加帽酶是可商购的(新英格兰生物实验室(new england biolabs)商品目录号第m2080s号)并具有由其d1亚基提供的rna三磷酸酶和鸟苷酸转移酶活性,以及由其d12亚基提供的鸟嘌呤甲基转移酶活性。因此,在s-腺苷甲硫氨酸和gtp的存在下,它可以将7-甲基鸟嘌呤添加到rna中,从而产生cap0。参见例如guo和moss(1990)《美国科学院院报》87:4023-4027,和mao和shuman(1994)《生物化学杂志》269:24472-24479,它们中的每一个以全文引用的方式并入本文用于所有目的。
[0165]
嵌合cas mrna还可以包含聚腺苷酸化(聚a)尾。例如,聚a尾可以包含至少20个、至少30个、至少40个、至少50个、至少60个、至少70个、至少80个、至少90个或至少100个腺嘌呤,以及任选地多达300个腺嘌呤。例如,聚a尾可以包含95、96、97、98、99或100个腺嘌呤核苷酸。
[0166]
编码嵌合cas蛋白的核酸可用于稳定整合到细胞的基因组中并可操作地链接到细胞中有活性的启动子。替代地,编码嵌合cas蛋白的核酸可以与表达构建体中的启动子可操作地链接。表达构建体包括能够引导基因或其它感兴趣的核酸序列(例如,嵌合cas基因)的表达并且可以将此类感兴趣的核酸序列转移到靶细胞的任何核酸构建体。例如,编码嵌合cas蛋白的核酸可以在包含编码grna的dna的载体中。可替代地,其可以在与包括对grna进行编码的dna的载体分离的载体或质粒中。可以用于表达构建体的启动子包括在例如真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、多能细胞、胚胎干(es)细胞、成体干细胞、发育受限的祖细胞、诱导的多能干(ips)细胞或单细胞期胚胎中的一个或多个中具有活性的启动子。此类启动子可以是例如条件启动子、诱导型启动子、组成型启动子或组织特异性启动子。任选地,启动子可以是双向启动子,其在一个方向上驱动嵌合cas蛋白的表达并且在另一个方向上驱动向导rna的表达。此类双向启动子可以由以下各项组成:(1)含有3个外部控制元件:远侧序列元件(dse)、近侧序列元件(pse)和tata框的完整的、常规的、单向的pol iii启动子;(2)包括在相反定向上与dse的5'末端融合的pse和tata盒的第二基本pol iii启动子。例如,在h1启动子中,dse邻近pse和tata框,并且可以通过产生杂合启动子使启动子双向化,其中通过源自u6启动子的附加pse和tata盒来控制反向转录。参见例如us 2016/0074535,所述文献出于所有目的通过引用整体并入本文。使用双向启动子同时表达编码嵌
合cas蛋白和向导rna的基因允许生成紧凑表达盒以促进递送。
[0167]
(1)cas蛋白
[0168]
cas蛋白通常包括可以与向导rna相互作用的至少一个rna识别或结合结构域。cas蛋白的功能片段或功能变体是保留与向导rna形成复合物并结合靶基因中的靶序列(并且例如激活靶基因的转录)的能力的功能片段或功能变体。
[0169]
除了如本文别处所述的转录激活结构域之外,cas蛋白还可以包含核酸酶域(例如,dnase域或rnase域)、dna结合域、解旋酶域、蛋白-蛋白相互作用域、二聚化域和其它域。一些此类结构域(例如,dnase结构域)可以来自天然cas蛋白。可以添加其它此类结构域以制备经修饰的cas蛋白。核酸酶结构域对核酸切割具有催化活性,所述核酸切割包含核酸分子的共价键的断裂。切割可以产生平末端或交错末端,并且其可以是单链或双链的。例如,野生型cas9蛋白通常将产生钝性切割产物。替代地,野生型cpf1蛋白(例如,fncpf1)可以产生具有5-核苷酸5'突出端的切割产物,其中切割发生在非靶向链上的pam序列的第18个碱基对之后和靶向链上的第23个碱基对之后。cas蛋白可以具有完整的切割活性以在靶基因组基因座处产生双链断裂(例如,具有平末端的双链断裂),或者其可以是在靶基因组基因座处产生单链断裂的切口酶。在一个实例中,本文公开的嵌合cas蛋白的cas蛋白部分已被修饰以具有降低的核酸酶活性(例如,与野生型cas蛋白相比,核酸酶活性减少至少约70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%)或基本上缺乏所有核酸酶活性(即,与野生型cas蛋白相比,核酸酶活性减少至少90%、95%、97%,98%、99%或100%,或具有不超过约0%、1%、2%、3%、5%或10%的野生型cas蛋白的核酸酶活性)。无核酸酶活性的cas蛋白是在其催化(即,核酸酶)域中具有已知为失活性突变的突变(例如,在cpf1蛋白中的ruvc样内切核酸酶域中的失活性突变,或在cas9中的hnh内切核酸酶域和ruvc样内切核酸酶域两者中的失活性突变)的cas蛋白,或与野生型cas蛋白相比具有减少至少约97%、98%、99%或100%的核酸酶活性的cas蛋白。下文公开了降低或基本上消除核酸酶活性的不同cas蛋白突变的实例。
[0170]
cas蛋白的实例包含cas1、cas1b、cas2、cas3、cas4、cas5、cas5e(casd)、cas6、cas6e、cas6f、cas7、cas8a1、cas8a2、cas8b、cas8c、cas9(csn1或csx12)、cas10、cas10d、casf、casg、cash、csy1、csy2、csy3、cse1(casa)、cse2(casb)、cse3(case)、cse4(casc)、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4和cu1966以及其同源物或经修饰的版本。
[0171]
示范性cas蛋白是cas9蛋白或衍生自cas9蛋白的蛋白。cas9蛋白来自ii型crispr/cas系统,并且通常共享具有保守结构的四个关键基序。基序1、2和4是类似ruvc的基序,并且基序3是hnh基序。示例性cas9蛋白来自酿脓链球菌(streptococcus pyogenes)、嗜热链球菌(streptococcus thermophilus)、链球菌(streptococcus sp.)、金黄色葡萄球菌(staphylococcus aureus)、达松维尔拟诺卡氏菌(nocardiopsisdassonvillei)、始旋链霉菌(streptomyces pristinaespiralis)、绿产色链霉菌(streptomyces viridochromogenes)、绿产色链霉菌(streptomyces viridochromogenes)、链孢囊菌(streptosporangiumroseum)、链孢囊菌(streptosporangiumroseum)、酸热脂环酸杆菌(alicyclobacillus acidocaldarius)、假蕈状芽孢杆菌(bacillus pseudomycoides)、硒
化芽孢杆菌(bacillus selenitireducens)、西伯利亚微小杆菌(exiguobacteriumsibiricum)、德氏乳酸杆菌(lactobacillus delbrueckii)、唾液乳杆菌(lactobacillus salivarius)、海洋微颤菌(microscilla marina)、伯克氏菌目细菌(burkholderiales bacterium)、食萘极地单胞菌(polaromonasnaphthalenivorans)、极地单胞菌(polaromonas sp.)、瓦氏鳄球藻(crocosphaerawatsonii)、蓝丝菌(cyanothece sp.)、铜绿微囊藻(microcystis aeruginosa)、聚球藻(synechococcus sp.)、阿拉伯糖醋酸杆菌(acetohalobiumarabaticum)、德根斯产氨菌(ammonifexdegensii)、热解纤维素菌(caldicelulosiruptorbecscii)、候选金矿菌(candidatusdesulforudis)、肉毒杆菌(clostridium botulinum)、艰难梭菌(clostridium difficile)、大芬戈尔德菌(finegoldia magna)、嗜热盐碱厌氧菌(natranaerobius thermophilus)、丙酸降解菌(pelotomaculumthermopropionicum)、喜温嗜酸硫杆菌(acidithiobacilluscaldus)、嗜酸氧化亚铁硫杆菌(acidithiobacillusferrooxidans)、异色变色菌(allochromatiumvinosum)、海杆菌(marinobactersp.)、嗜盐亚硝化球菌(nitrosococcushalophilus)、沃森亚硝化球菌(nitrosococcuswatsoni)、嗜盐假交替单胞菌(pseudoalteromonashaloplanktis)、雷氏纤线杆菌(ktedonobacterracemifer)、伊芙氏甲烷盐菌(methanohalobiumevestigatum)、鱼腥藻(anabaenavariabilis)、泡沫节球藻(nodulariaspumigena)、念珠藻(nostoc sp.)、节旋藻(arthrospira maxima)、盘状节旋藻(arthrospira platensis)、节旋藻(arthrospira sp.)、鞘丝藻(lyngbya sp.)、原型微鞘藻(microcoleuschthonoplastes)、颤蓝细菌(oscillatoria sp.)、运动石袍菌(petrotogamobilis)、非洲栖热腔菌(thermosipho africanus)、深海阿卡罗虎尾草(acaryochloris marina)、脑膜炎奈瑟氏菌(neisseria meningitidis)或空肠弯曲杆菌(campylobacter jejuni)。cas9家族成员的另外实例在wo 2014/131833中进行描述,所述文献出于所有目的通过引用整体并入本文。来自酿脓链球菌的cas9(spcas9)(指定swissprot登录号q99zw2)是示例性cas9蛋白。来自金黄色葡萄球菌的cas9(sacas9)(指定uniprot登录号j7rua5)是另一示例性cas9蛋白。来自空肠弯曲杆菌的cas9(cjcas9)(指定uniprot登录号q0p897)是另一种示例性cas9蛋白。参见例如,kim等人(2017),《自然通讯(nat.commun.)》8:14500,所述文献出于所有目的通过引用整体并入本文。sacas9小于spcas9,并且cjcas9小于sacas9和spcas9两者。来自脑膜炎奈瑟氏球菌的cas9(nme2cas9)是另一种示例性cas9蛋白。参见例如edraki等人,(2019)《分子细胞(mol.cell)》73(4):714-726,该文献以全文引用的方式并入本文用于所有目的。来自嗜热链球菌的cas9蛋白(例如,由crispr1基因座编码的嗜热链球菌lmd-9cas9(st1cas9)或来自crispr3基因座的嗜热链球菌cas9(st3cas9))是其它示例性cas9蛋白。来自新凶手弗朗西丝菌(francisellanovicida)(fncas9)的cas9或识别置换性pam(e1369r/e1449h/r1556a取代)的rha新凶手弗朗西斯菌cas9变体是其它示例性cas9蛋白。这些和其它示例性cas9蛋白例如在cebrian-serrano和davies(2017),《哺乳动物基因组(mamm.genome)》28(7):247-261中综述,所述文献出于所有目的通过引用整体并入本文。在wo 2013/176772、wo 2014/065596、wo 2016/106121和wo 2019/067910中提供了cas9编码序列、cas9 mrna和cas9蛋白序列的实例,它们中的每一个以全文引用的方式并入本文用于所有目的。orf和cas9氨基酸序列的具体实例在wo 2019/067910第段的表30中提供,并且cas9 mrna和orf的具体实例在
wo 2019/067910的第至段中提供。
[0172]
cas蛋白的另一个实例是cpf1(来自普雷沃氏菌(prevotella)和弗朗西斯氏菌(francisella)1的crispr)蛋白。cpf1是含有与cas9的对应结构域同源的ruvc样核酸酶结构域以及与特征性富含精氨酸的cas9簇的对应物的大蛋白质(约1300个氨基酸)。然而,cpf1缺乏cas9蛋白中存在的hnh核酸酶结构域,并且ruvc样结构域在cpf1序列中是连续的,而cas9则相反,其含有包含hnh结构域的长插入物。参见例如,zetsche等人(2015),《细胞》163(3):759-771,所述文献出于所有目的通过引用整体并入本文。示例性cpf1蛋白来自土拉弗朗西斯菌(francisellatularensis)1、土拉弗朗西丝菌新凶手亚种(francisellatularensis subsp.novicida)、易北河普雷沃氏菌(prevotellaalbensis)、毛螺菌科细菌(lachnospiraceae bacterium)mc20171、解蛋白丁酸弧菌(butyrivibrioproteoclasticus)、异域菌门细菌(peregrinibacteria bacterium)gw2011_gwa2_33_10、俭菌超门细菌(parcubacteria bacterium)gw2011_gwc2_44_17、史密斯氏菌属(smithella sp.)scadc、氨基酸球菌属(acidaminococcussp.)bv3l6、毛螺菌科细菌(lachnospiraceae bacterium)ma2020、候选白蚁甲烷支原体(candidatusmethanoplasmatermitum)、挑剔真杆菌(eubacterium eligens)、牛眼莫拉氏菌(moraxella bovoculi)237、稻田氏钩端螺旋体(leptospira inadai)、毛螺菌科细菌(lachnospiraceae bacterium)nd2006、狗口腔卟啉单胞菌(porphyromonascrevioricanis)3、解糖胨普雷沃氏菌(prevotelladisiens)和猕猴卟啉单胞菌(porphyromonasmacacae)。来自新凶手弗朗西丝氏菌(francisellanovicida)u112的cpf1(fncpf1;指定uniprot登录号a0q7q2)是示例性cpf1蛋白。
[0173]
cas蛋白可以是野生型蛋白(即,自然界中存在的那些蛋白)、经修饰的cas蛋白(即,cas蛋白变体)或野生型或经修饰的cas蛋白的片段。就野生型或经修饰的cas蛋白的催化活性而言,cas蛋白也可以是活性变体或片段。就催化活性而言,活性变体或片段可以包括与野生型或经修饰的cas蛋白或其部分具有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更多的序列同一性,其中活性变体保留在期望的切割位点处切割的能力,并因此保留切口诱导或双链断裂诱导活性。对切口诱导或双链断裂诱导活性的测定是已知的,并且通常测量cas蛋白对含有切割位点的dna底物的总体活性和特异性。
[0174]
经修饰的cas蛋白的一个实例是经修饰的spcas9-hf1蛋白,其是具有设计成减少非特异性dna接触的改变(n497a/r661a/q695a/q926a)的酿脓链球菌cas9的高保真变体。参见例如,kleinstiver等人(2016),《自然(nature)》529(7587):490-495,所述文献出于所有目的通过引用整体并入本文。经修饰cas蛋白的另一个实例是被设计成减少脱靶效应的经修饰的espcas9变体(k848a/k1003a/r1060a)。参见例如,slaymaker等人(2016),《科学(science)》351(6268):84-88,所述文献出于所有目的通过引用整体并入本文。其它spcas9变体包含k855a和k810a/k1003a/r1060a。这些和其它经修饰的cas蛋白例如在cebrian-serrano和davies(2017),《哺乳动物genome)》28(7):247-261中综述,所述文献出于所有目的通过引用整体并入本文。另一种经修饰cas9蛋白的实例是xcas9,其是可以识别扩大范围的pam序列的spcas9变体。参见例如hu等人(2018),《自然(nature)》556:57-63,所述文献出于所有目的通过引用整体并入本文。
[0175]
可以修饰cas蛋白以增加或减少核酸结合亲和力、核酸结合特异性和酶活性中的一种或多种。也可以修饰cas蛋白以改变蛋白质的任何其它活性或性质,如稳定性。例如,cas蛋白的一个或多个核酸酶结构域可以是经修饰的、缺失的或失活的,或者可以截断cas蛋白以去除对蛋白质功能不必要的结构域或优化(例如,增强或减少)cas蛋白的活性或性质。
[0176]
cas蛋白可以包括至少一个核酸酶结构域,如dnase结构域。例如,野生型cpf1蛋白通常包括切割靶dna的两条链的ruvc样结构域,其可能呈二聚体构型。cas蛋白还可以包括至少两个核酸酶结构域,如dnase结构域。例如,野生型cas9蛋白通常包括ruvc样核酸酶结构域和hnh样核酸酶结构域。ruvc和hnh结构域可以各自切割不同的双链dna链,以在dna中形成双链断裂。参见例如jinek等人(2012)《科学(science)》337(6096):816-821,所述文献出于所有目的通过引用整体并入本文。
[0177]
核酸酶结构域中的一个或多个或所有核酸酶结构域可以缺失或突变,使得其不再具有功能或具有降低的核酸酶活性。例如,如果cas9蛋白中的核酸酶结构域之一进行缺失或突变,则所得cas9蛋白可以被称为切口酶,并且可以在双链靶dna内产生单链断裂,但不会产生双链断裂(即,其可以切割互补链或非互补链,但不能同时切割两者)。如果两个核酸酶结构域都缺失或突变,则所得cas蛋白(例如,cas9)切割双链dna(例如,核酸酶无效或无核酸酶活性的cas蛋白,或催化上死亡的cas蛋白(dcas))的两条链的能力将降低。将cas9转化为切口酶的突变的实例是来自酿脓链球菌的cas9的ruvc结构域中的d10a(在cas9的位置10处天冬氨酸转化为丙氨酸)突变。同样地,来自酿脓链球菌的cas9的hnh结构域中的h939a(在氨基酸位置839处组氨酸到丙氨酸)、h840a(在氨基酸位置840处组氨酸到丙氨酸)或n863a(在氨基酸位置n863处天冬酰胺到丙氨酸)可以将cas9转化为切口酶。将cas9转化为切口酶的突变的其它实例包含来自嗜热链球菌的cas9的对应突变。参见例如sapranauskas等人(2011)《核酸研究》39(21):9275-9282和wo 2013/141680,它们中的每一个以全文引用的方式并入本文用于所有目的。可以使用如定点诱变、pcr介导的诱变或总基因合成等方法产生此类突变。产生切口酶的其它突变实例可以在例如wo 2013/176772和wo2013/142578中找到,所述文献中的每个文献出于所有目的通过引用整体并入本文。如果cas蛋白中的所有核酸酶结构域都缺失或突变(例如,cas9蛋白中的两个核酸酶结构域都缺失或突变),则所得cas蛋白(例如,cas9)切割双链dna(例如,核酸酶无效或无核酸酶活性的cas蛋白)的两条链的能力将降低。一个具体实例是d10a/h840a酿脓链球菌cas9双突变体或者当与酿脓链球菌cas9最佳比对时来自另一物种的cas9中的对应双突变体。另一个具体实例是d10a/n863a酿脓链球菌cas9双突变体或者当与酿脓链球菌cas9最佳比对时来自另一物种的cas9中的对应双突变体。无催化活性的cas9蛋白(dcas9)的一个实例包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:2所示的dcas9蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。
[0178]
xcas9的催化结构域中的失活突变的实例与上述针对spcas9的突变相同。金黄色葡萄球菌cas9蛋白催化结构域中的失活突变的实例也是已知的。例如,金黄色葡萄球菌cas9酶(sacas9)可以包括用于产生无核酸酶活性的cas蛋白的位置n580处的取代(例如,n580a取代)和位置d10处的取代(例如,d10a取代)。参见例如wo 2016/106236,所述文献出
于所有目的通过引用整体并入本文。nme2cas9的催化结构域中的失活突变的实例也是已知的(例如,d16a和h588a的组合)。st1cas9的催化结构域中失活突变的实例也是已知的(例如,d9a、d598a、h599a和n622a的组合)。st3cas9的催化结构域中的失活突变的实例也是已知的(例如,d10a和n870a的组合)。cjcas9催化结构域中的失活突变的实例也是已知的(例如,d8a和h559a的组合)。fncas9和rha fncas9的催化结构域中的失活突变的实例也是已知的(例如,n995a)。
[0179]
cpf1蛋白的催化结构域中的失活突变的实例也是已知的。参考来自新凶手弗朗西丝氏菌u112(fncpf1)、氨基酸球菌bv3l6(ascpf1)、毛螺科菌nd2006(lbcpf1)和牛眼莫拉氏菌237(mbcpf1 cpf1)的cpf1蛋白,此类突变可以包含ascpf1的位置908、993或1263处或cpf1直系同源物中的对应位置处,或lbcpf1的位置832、925、947或1180或cpf1直系同源物中的对应位置处的突变。此类突变可以包含例如ascpf1的突变d908a、e993a和d1263a或cpf1直系同源物中的对应突变或lbcpf1的d832a、e925a、d947a和d1180a或cpf1直系同源物中的对应突变中的一种或多种突变。参见例如us 2016/0208243,所述文献出于所有目的通过引用整体并入本文。
[0180]
cas蛋白也可以作为融合蛋白与异源多肽可操作地连接。例如,除了转录激活域之外,cas蛋白也可以与切割域或表观遗传修饰域融合。参见wo 2014/089290,所述文献出于所有目的通过引用整体并入本文。cas蛋白也可以与异源多肽融合,提供增加或减少的稳定性。融合结构域或异源多肽可以定位于n端、c端或cas蛋白内部。
[0181]
举例来说,cas蛋白可以与提供亚细胞定位的一种或多种异源多肽融合。此类异源多肽可以包含例如一种或多种核定位信号(nls),如用于靶向细胞核的单组分sv40 nls和/或双组分α-输入蛋白nls、用于靶向线粒体的线粒体定位信号、er保留信号等。参见例如lange等人(2007)《生物化学杂志(j.biol.chem.)》282(8):5101-5105,所述文献出于所有目的通过引用整体并入本文。此类亚细胞定位信号可以定位于n端、c端或cas蛋白内的任何位置。nls可以包含碱性氨基酸的段,并且可以是单部分序列或双部分序列。任选地,cas蛋白可以包含两个或更多个nls,包括n端处的nls(例如,α-输入蛋白nls或单部分nls)和c端处的nls(例如,sv40 nls或双部分nls)。cas蛋白还可以包括n端处的两个或更多个nls和/或c端处的两个或更多个nls。
[0182]
在一个实例中,cas蛋白可以与1至10个nls、1至5个nls或一个nls融合。在使用一个nls的情况下,nls可以链接在cas序列的n端或c端处。它也可以插入到cas序列的内部。在其它实例中,cas蛋白可以与多于一个nls融合。例如,cas蛋白可以与2、3、4或5个nls融合或可以与两个nls融合。在某些情况下,两个nls可以相同(例如,两个sv40 nls)或不同。例如,cas蛋白可以融合到链接在羧基端的两个sv40 nls序列。在另一个实例中,cas蛋白可以与两个nls融合,一个链接在n端并且一个链接在c端。在另一个实例中,cas蛋白可以与3个nls融合。在另一个实例中,cas蛋白可以不与nls融合。在一些实例中,nls可以是单部分序列,例如,sv40 nls、pkkkrkv(seq id no:58)或pkkkrrv(seq id no:59)。在一些实例中,nls可以是双部分序列,如核质蛋白的nls,krpaatkkagqakkkk(seq id no:60)。在具体实例中,单个pkkkrkv(seq id no:58)nls可以链接在rna导向的dna结合剂的c端。一个或多个接头任选地包括在融合位点处。
[0183]
cas蛋白也可以与细胞穿透性结构域或蛋白质转导结构域可操作地连接。例如,细
胞穿透性结构域可以源自hiv-1tat蛋白、来自人乙型肝炎病毒的tlm细胞穿透基序、mpg、pep-1、vp22、来自单纯疱疹病毒的细胞穿透性肽或聚精氨酸肽序列。参见例如wo2014/089290和wo 2013/176772,所述文献中的每个文献出于所有目的通过引用整体并入本文。细胞穿透性结构域可以定位于n端、c端或cas蛋白内的任何位置。
[0184]
cas蛋白也可以与异源多肽可操作地连接以便于进行追踪或纯化,如荧光蛋白、纯化标签或表位标签。荧光蛋白的实例包含绿色荧光蛋白(例如,gfp、gfp-2、taggfp、turbogfp、egfp、祖母绿、azami绿、单体azami绿、copgfp、acegfp、zsgreenl)、黄色荧光蛋白(例如,yfp、eyfp、柠檬黄、venus、ypet、phiyfp、zsyellowl)、蓝色荧光蛋白(例如,ebfp、ebfp2、石青、mkalamal、gfpuv、天蓝色、t-天蓝色(t-sapphire))、青色荧光蛋白(例如,ecfp、蔚蓝色(cerulean)、cypet、amcyanl、midoriishi-青色)、红色荧光蛋白(例如,mkate、mkate2、mplum、dsred单体、mcherry、mrfp1、dsred-表达、dsred2、dsred-单体、hcred-tandem、hcredl、asred2、eqfp611、mraspberry、mstrawberry、jred)、橙色荧光蛋白(例如,morange、mko、kusabira-橙色、单体kusabira-橙色、mtangerine、tdtomato)和任何其它合适的荧光蛋白。标签的实例包含谷胱甘肽-s-转移酶(gst)、几丁质结合蛋白(cbp)、麦芽糖结合蛋白、硫氧还蛋白(trx)、多(nanp)、串联亲和纯化(tap)标签、myc、acv5、au1、au5、e、ecs、e2、flag、血球凝集素(ha)、nus、softag1、softag3、strep、sbp、glu-glu、hsv、kt3、s、s1、t7、v5、vsv-g、组氨酸(his)、生物素羧基载体蛋白(bccp)和钙调蛋白。
[0185]
cas蛋白还可以与经标记的核酸栓系。此类拴系(即,物理链接)可以通过共价相互作用或非共价相互作用来实现,并且拴系可以是直接的(例如,通过直接融合或化学缀合,这可以通过蛋白上的半胱氨酸或赖氨酸残基的修饰或内含子修饰来实现),或者可以通过如链霉亲和素或适体等一个或多个中间接头或衔接子分子来实现。参见例如,pierce等人(2005),《药物化学短评(mini rev.med.chem.)》5(1):41-55;duckworth等人(2007),《德国应用化学会刊(angew.chem.int.ed.engl.)》46(46):8819-8822;schaeffer和dixon(2009),《澳大利亚化学杂志(australian j.chem.)》62(10):1328-1332;goodman等人(2009),《生物化学(chembiochem.)》10(9):1551-1557;以及khatwani等人(2012)《生物有机化学与医药化学(bioorg.med.chem.)》20(14):4532-4539,所述文献中的每个文献出于所有目的通过引用整体并入本文。用于合成蛋白质-核酸缀合物的非共价策略包含生物素-链霉亲和素和镍-组氨酸方法。可以通过使用多种化学反应连接适当功能化的核酸和蛋白质来合成共价蛋白质-核酸缀合物。这些化学反应中的一些化学反应涉及将寡核苷酸直接附着到蛋白质表面上的氨基酸残基(例如,赖氨酸胺或半胱氨酸硫醇),而其它更复杂的方案需要蛋白质的翻译后修饰或者催化或反应蛋白结构域的参与。用于蛋白质与核酸共价附着的方法可以包含例如寡核苷酸与蛋白质赖氨酸或半胱氨酸残基的化学交联、表达的蛋白质连接、化学酶法和光适体的使用。经标记的核酸可以与cas蛋白内的c端、n端或内部区域栓系。在一个实例中,经标记的核酸与cas蛋白的c端或n端栓系。同样地,cas蛋白可以与经标记的核酸内的5'末端、3'末端或内部区域栓系。也就是说,经标记的核酸可以以任何取向和极性拴系。例如,cas蛋白可以与经标记的核酸的5'末端或3'末端栓系。
[0186]
(2)转录激活域
[0187]
本文公开的嵌合cas蛋白可以包含一个或多个转录激活域。转录激活域包括天然存在的转录因子区域,这些区域与dna结合域(例如,与向导rna复合的无催化活性的cas蛋
白)结合,可以通过直接接触转录机器或通过其它蛋白(如共激活子)来激活来自启动子的转录。转录激活域还包括转录因子的此类区域的功能片段或变体,以及衍生自天然的、天然存在的转录激活域或人工产生或合成以激活靶基因转录的工程化转录激活域。功能片段是当可操作地链接到合适的dna结合域时能够激活靶基因转录的片段。功能变体是当可操作地链接到合适的dna结合域时能够激活靶基因转录的变体。
[0188]
用于本文公开的嵌合cas蛋白的特定转录激活域包含vp64转录激活域或其功能片段或变体。vp64是来自单纯疱疹vp16激活域的最小激活域的四聚体重复。例如,转录激活域可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:3所示的vp64转录激活域蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。
[0189]
转录激活域的其它实例包括单纯疱疹病毒vp16反式激活域、vp64(单纯疱疹病毒vp16的四重串联重复)、nf-κb p65(nf-κb反式激活亚基p65)激活域、myod1反式激活域、hsf1反式激活域(来自人热休克因子1的反式激活域)、rta(epstein barr病毒r反式激活子激活域)、set7/9反式激活域、p53激活域1、p53激活域2、creb(camp响应元件结合蛋白)激活域、e2a激活域、nfat(激活的t细胞的核因子)激活域,及其功能片段和变体。参见例如us 2016/0298125、us 2016/0281072和wo 2016/049258,它们中的每一个以全文引用的方式并入本文用于所有目的。转录激活域的其它实例包括gcn4、mll、rtg3、gln3、oaf1、pip2、pdr1、pdr3、pho4、leu3及其功能片段和变体。参见例如us 2016/0298125,该文献以全文引用的方式并入本文用于所有目的。转录激活域的其它实例包括spl、vax、gata4及其功能片段和变体。参见例如wo 2016/149484,该文献以全文引用的方式并入本文用于所有目的。其它实例包括来自oct1、oct-2a、ap-2、ctf1、p300、cbp、pcaf、src1、pvalf、erf-2、osgai、half-1、c1、ap1、arf-5、arf-6、arf-7、arf-8、cprf1、cprf4、myc-rp/gp和trab1pc4的激活域及其功能片段和变体。参见例如us 2016/0237456、ep3045537和wo 2011/146121,所述文献中的每个文献出于所有目的通过引用整体并入。附加的合适转录激活域也是已知的。参见例如wo 2011/146121,该文献以全文引用的方式并入本文用于所有目的。
[0190]
b.嵌合衔接蛋白
[0191]
还提供了可以结合到本文别处公开的向导rna的嵌合衔接蛋白。本文公开的嵌合衔接蛋白可用于dcas协同激活介体(sam)样系统,以增加针对靶基因内的靶序列的转录激活域的数量和多样性,从而激活靶基因的转录。
[0192]
此类嵌合衔接蛋白包括:(a)与向导rna内的衔接结合元件特异性结合的衔接子(即,衔接结构域或衔接蛋白);以及(b)一个或多个异源转录激活结构域。例如,此类融合蛋白可以包括1、2、3、4、5或更多个转录激活结构域(例如,两个或更多个异源转录激活结构域或三个或更多个异源转录激活结构域)。在一个实例中,此类嵌合衔接蛋白可以包括:(a)与向导rna中的衔接结合元件特异性结合的衔接子(即,衔接结构域或衔接蛋白);以及(b)两个或更多个转录激活结构域。例如,嵌合衔接蛋白可以包括:(a)与向导rna中的一个或多个ms2适体(例如,向导rna中的单独定位中的两个ms2适体)特异性结合的ms2外壳蛋白衔接子;以及(b)一个或多个(例如,两个或更多个转录激活结构域)。例如,两个转录激活结构域可以是p65和hsf1转录激活结构域或其功能片段或变体。然而,还提供了其中转录激活结构域包括其它转录激活结构域或其功能片段或变体的嵌合衔接蛋白。
[0193]
一个或多个转录激活结构域可以与衔接子直接融合。可替代地,一个或多个转录激活结构域可以通过接头或接头组合或通过一个或多个另外的结构域连接到衔接子。同样,如果存在两个或更多个转录激活结构域,则所述结构域可以直接彼此融合或可以通过接头或接头组合或通过一个或多个另外的结构域彼此连接。可以用于这些融合蛋白的接头可以包含不干扰融合蛋白的功能的任何序列。示例性接头是短的(例如,2-20个氨基酸)并且通常是柔性的(例如,包括具有高自由度的氨基酸,如甘氨酸、丙氨酸和丝氨酸)。接头的一些具体实例包含由gggs(seq id no:4)或ggggs(seq id no:5)(如gggs(seq id no:4)或ggggs(seq id no:5)的两个、三个、四个或更多个重复的任何组合)组成的一个或多个单元。也可以使用其它接头序列。
[0194]
一个或多个转录激活结构域和衔接子可以在嵌合衔接蛋白内呈任何顺序。作为一种选择,一个或多个转录激活结构域可以在衔接子的c端并且衔接子可以在一个或多个转录激活结构域的n端。例如,一个或多个转录激活结构域可以在嵌合衔接蛋白的c端,并且衔接子可以在嵌合衔接蛋白的n端。然而,一个或多个转录激活结构域可以在衔接子的c端而不在嵌合衔接蛋白的c端(例如,如果核定位信号处于嵌合衔接蛋白的c端处)。同样,衔接子可以在一个或多个转录激活结构域的n端而不在嵌合衔接蛋白的n端处(例如,如果核定位信号处于嵌合衔接蛋白的n端处)。作为另一种选择,一个或多个转录激活结构域可以在衔接子的n端并且衔接子可以在一个或多个转录激活结构域的c端。例如,一个或多个转录激活结构域可以在嵌合衔接蛋白的n端,并且衔接子可以在嵌合衔接蛋白的c端。作为另一种选择,如果嵌合衔接蛋白包括两个或更多个转录激活结构域,则两个或更多个转录激活结构域可以位于衔接子的侧翼。
[0195]
嵌合衔接蛋白也可以与另外的异源多肽可操作地连接或融合。融合或连接的异源多肽可以定位于n端、c端或嵌合衔接蛋白内部的任何位置。例如,嵌合衔接蛋白可以进一步包括核定位信号。此类蛋白质的具体实例包括与ms2外壳蛋白(mcp)的p65转录激活结构域c端和p65转录激活结构域的hsf1转录激活结构域c端(直接或通过nls)连接的ms2外壳蛋白(衔接子)。此类蛋白质可以从n端到c端包括:mcp;核定位信号;p65转录激活结构域;以及hsf1转录激活结构域。例如,嵌合衔接蛋白可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:6所示的mcp-p65-hsf1嵌合衔接蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。
[0196]
嵌合衔接蛋白也可以与提供亚细胞定位的一个或多个异源多肽融合或链接。此类异源多肽可以包括例如一个或多个核定位信号(nls),如用于靶向细胞核的sv40 nls和/或α-输入蛋白nls、用于靶向线粒体的线粒体定位信号、er保留信号等等。参见例如lange等人(2007)《生物化学杂志(j.biol.chem.)》282(8):5101-5105,所述文献出于所有目的通过引用整体并入本文。此类亚细胞定位信号可以位于嵌合衔接蛋白内的n端、c端或任何位置(例如,在嵌合衔接蛋白的衔接蛋白组分的c端或n端或在嵌合衔接蛋白的转录激活子域组分的c端或n端)。nls可以包含例如碱性氨基酸的段,并且可以是单部分序列或双部分序列。任选地,嵌合衔接蛋白包含两个或更多个nls,包括n端处的nls(例如,α-输入蛋白nls)和/或c端处的nls(例如,sv40 nls)。嵌合衔接蛋白还可以包含n端处的两个或更多个nls和/或c端处的两个或更多个nls。
链霉亲和素和镍-组氨酸方法。可以通过使用多种化学反应连接适当功能化的核酸和蛋白质来合成共价蛋白质-核酸缀合物。这些化学反应中的一些化学反应涉及将寡核苷酸直接附着到蛋白质表面上的氨基酸残基(例如,赖氨酸胺或半胱氨酸硫醇),而其它更复杂的方案需要蛋白质的翻译后修饰或者催化或反应蛋白结构域的参与。用于蛋白质与核酸共价附着的方法可以包含例如寡核苷酸与蛋白质赖氨酸或半胱氨酸残基的化学交联、表达的蛋白质连接、化学酶法和光适体的使用。经标记的核酸可以与嵌合衔接蛋白内的c端、n端或内部区域拴系。同样地,嵌合衔接蛋白可以与经标记的核酸内的5'末端、3'末端或内部区域拴系。也就是说,经标记的核酸可以以任何取向和极性拴系。
[0201]
嵌合衔接蛋白可以以任何形式提供。例如,嵌合衔接蛋白可以以蛋白的形式提供,如与grna复合的嵌合衔接蛋白。替代地,可以以编码嵌合衔接蛋白的核酸的形式提供嵌合衔接蛋白,如rna(例如,信使rna(mrna))或dna。在具体实例中,嵌合衔接蛋白可以作为mrna(例如,体外转录的mrna)提供,如也编码嵌合cas蛋白的多顺反子或双顺反子mrna。任选地,可以对编码嵌合衔接蛋白的核酸进行密码子优化以在特定细胞或生物体中有效翻译成蛋白。例如,与天然存在的多核苷酸序列相比,可以修饰编码嵌合衔接蛋白的核酸以取代在真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、大鼠细胞或任何其它感兴趣的宿主细胞中具有更高使用频率的密码子。当编码嵌合衔接蛋白的核酸被引入到细胞中时,嵌合衔接蛋白可以在细胞中进行瞬时、条件性或组成性表达。
[0202]
可以修饰作为mrna提供的嵌合衔接蛋白以提高稳定性和/或免疫原性特性。可以对mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合衔接蛋白的mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统中的模式识别受体的激活)。编码嵌合衔接蛋白的mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合衔接蛋白的mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合衔接mrna。同样地,嵌合衔接mrna可以通过使用同义密码子消耗尿苷来修饰。其它可能的修饰在本文别处更详细地描述。
[0203]
可以修饰作为mrna提供的嵌合衔接蛋白以提高稳定性和/或免疫原性特性。可以对mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合衔接蛋白的mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统中的模式识别受体的激活)。编码嵌合衔接蛋白的mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合衔接蛋白的mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合衔接mrna。同样地,嵌合衔接mrna可以通过使用同义密码子消耗尿苷来修饰。
[0204]
嵌合衔接mrna可以在至少一个、多个或所有尿苷位置包含经修饰的尿苷。经修饰的尿苷可以是在5位置被修饰(例如,用卤素、甲基或乙基)的尿苷。经修饰的尿苷可以是在1
位置被修饰(例如,用卤素、甲基或乙基)的假尿苷。经修饰的尿苷可以是例如假尿苷、n1-甲基-假尿苷、5-甲氧基尿苷、5-碘尿苷或其组合。在一些实例中,经修饰的尿苷是5-甲氧基尿苷。在一些实例中,经修饰的尿苷是5-碘尿苷。在一些实例中,经修饰的尿苷是假尿苷。在一些实例中,经修饰的尿苷是n1-甲基-假尿苷。在一些实例中,经修饰的尿苷是假尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是n1-甲基假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-碘尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和5-甲氧基尿苷的组合。
[0205]
本文公开的嵌合衔接mrna还可以包含5'帽,如cap0、cap1或cap2。5'帽通常是通过5'-三磷酸酯链接到mrna的5'至3'链的第一核苷酸(即,第一近帽核苷酸)的5'位置的7-甲基鸟嘌呤核糖核苷酸(其可以被进一步修饰,例如,关于arca)。在cap0中,mrna的第一和第二近帽核苷酸的核糖均包含2'-羟基。在cap1中,mrna的第一和第二转录核苷酸的核糖分别包含2'-甲氧基和2'-羟基。在cap2中,mrna的第一和第二近帽核苷酸的核糖均包含2'-甲氧基。参见例如katibah等人(2014)《美国科学院院报(proc.natl.acad.sci.u.s.a.)》111(33):12025-30和abbas等人(2017)《美国科学院院报》114(11):e2106-e2115,它们中的每一个以全文引用的方式并入本文用于所有目的。大多数内源性高等真核mrna,包括哺乳动物mrna,如人mrna,均包含cap1或cap2。cap0和其它不同于cap1和cap2的帽结构可能在哺乳动物(如人类)中具有免疫原性,因为先天免疫系统的组分(如ifit-1和ifit-5)将其识别为非自身,这可能导致细胞因子升高水平(包括i型干扰素)。先天免疫系统的组分如ifit-1和ifit-5也可能与eif4e竞争mrna与cap1或cap2以外的帽的结合,从而潜在地抑制mrna的翻译。
[0206]
可以共转录地包括帽。例如,arca(抗反向帽类似物;赛默飞世尔科技公司(thermo fisher scientific)商品目录号第am8045号)是包含与鸟嘌呤核糖核苷酸的5'位置链接的7-甲基鸟嘌呤3'-甲氧基-5'-三磷酸酯的帽类似物,其可在体外在起始时掺入转录物中。arca产生cap0帽,其中第一近帽核苷酸的2'位置是羟基。参见例如stepinski等人,(2001)《rna》7:1486-1495,该文献以全文引用的方式并入本文用于所有目的。
[0207]
cleancap
tm ag(m7g(5')ppp(5')(2'omea)pg;trilink生物技术公司商品目录号第n-7113号)或cleancap
tm gg(m7g(5')ppp(5')(2'omeg)pg;trilink生物技术公司商品目录号第n-7133号)可用于共转录地提供cap1结构。3'-o-甲基化版本的cleancap
tm ag和cleancap
tm gg也可从trilink生物技术公司获得,商品目录号分别为第n-7413和第n-7433号。
[0208]
替代地,可以在转录后将帽添加到rna。例如,牛痘加帽酶是可商购的(新英格兰生物实验室(new england biolabs)商品目录号第m2080s号)并具有由其d1亚基提供的rna三磷酸酶和鸟苷酸转移酶活性,以及由其d12亚基提供的鸟嘌呤甲基转移酶活性。因此,在s-腺苷甲硫氨酸和gtp的存在下,它可以将7-甲基鸟嘌呤添加到rna中,从而产生cap0。参见例如guo和moss(1990)《美国科学院院报》87:4023-4027,和mao和shuman(1994)《生物化学杂志》269:24472-24479,它们中的每一个以全文引用的方式并入本文用于所有目的。
[0209]
嵌合衔接mrna还可以包含聚腺苷酸化(聚a)尾。例如,聚a尾可以包含至少20个、至少30个、至少40个、至少50个、至少60个、至少70个、至少80个、至少90个或至少100个腺嘌
呤,以及任选地多达300个腺嘌呤。例如,聚a尾可以包含95、96、97、98、99或100个腺嘌呤核苷酸。
[0210]
编码嵌合衔接蛋白的核酸可以稳定地整合在细胞的基因组中,并可操作地链接到细胞中有活性的启动子。替代地,编码嵌合衔接蛋白的核酸可以与表达构建体中的启动子可操作地链接。表达构建体包括能够引导基因或其它感兴趣的核酸序列(例如,嵌合衔接基因)的表达并且可以将此类感兴趣的核酸序列转移到靶细胞的任何核酸构建体。例如,编码嵌合衔接蛋白的核酸可以在包含编码grna和/或嵌合cas蛋白的dna的载体中。替代地,其可以在与包含编码grna的dna或编码嵌合cas蛋白的dna的载体分离的载体或质粒中。可以用于表达构建体的启动子包括在例如真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、多能细胞、胚胎干(es)细胞、成体干细胞、发育受限的祖细胞、诱导的多能干(ips)细胞或单细胞期胚胎中的一个或多个中具有活性的启动子。此类启动子可以是例如条件启动子、诱导型启动子、组成型启动子或组织特异性启动子。任选地,启动子可以是双向启动子。此类双向启动子可以由以下各项组成:(1)含有3个外部控制元件:远侧序列元件(dse)、近侧序列元件(pse)和tata框的完整的、常规的、单向的pol iii启动子;(2)包括在相反定向上与dse的5'末端融合的pse和tata盒的第二基本pol iii启动子。例如,在h1启动子中,dse邻近pse和tata框,并且可以通过产生杂合启动子使启动子双向化,其中通过源自u6启动子的附加pse和tata盒来控制反向转录。参见例如us 2016/0074535,所述文献出于所有目的通过引用整体并入本文。
[0211]
(1)衔接子
[0212]
衔接子(即,衔接结构域或衔接蛋白)是特异性识别并与不同序列结合(例如,与不同dna和/或rna序列,如序列特异性方式的适体结合)的核酸结合结构域(例如,dna结合结构域和/或rna结合结构域)。适体包含通过其采用特异性三维构象的能力可以以高亲和力和特异性与靶分子结合的核酸。例如,此类衔接子可以与特异性rna序列和二级结构结合。这些序列(即,衔接结合元件)可以被工程化成向导rna。例如,ms2适体可以被工程化成向导rna,以特异性结合ms2外壳蛋白(mcp)。例如,衔接子可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:7所示的mcp序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。
[0213]
衔接子和靶标的一些具体实例包括存在于噬菌体外壳蛋白多样性内的rna结合蛋白/适体组合。例如,可以使用以下衔接蛋白或其功能片段或变体:ms2外壳蛋白(mcp)、pp7、qβ、f2、ga、fr、jp501、m12、r17、bz13、jp34、jp500、ku1、m1l、mx1、tw18、vk、sp、fi、id2、nl95、tw19、ap205、φcb8r、φcb12r、φcb23r、7s和prr1。参见例如wo 2016/049258,该文献以全文引用的方式并入本文用于所有目的。衔接蛋白的功能片段或功能变体是保留与特定衔接结合元件结合的能力(例如,以序列特异性方式与特定衔接结合序列结合的能力)的功能片段或功能变体。例如,可以使用pp7假单胞菌噬菌体外壳蛋白变体,其中氨基酸68-69突变为sg并且氨基酸70-75从野生型蛋白中缺失。参见例如wu等人(2012)《生物物理学杂志(biophys.j.)》102(12):2936-2944,和chao等人(2007)《自然结构与分子生物学(nat.struct.mol.biol.)》15(1):103-105,它们中的每一个以全文引用的方式并入本文用于所有目的。同样地,可以使用mcp变体,如n55k突变体。参见例如spingola和peabody
(1994)《生物化学杂志》269(12):9006-9010,该文献以全文引用的方式并入本文用于所有目的。
[0214]
可以使用的衔接蛋白的其它实例包括全部或部分内切核糖核酸酶csy4或λn蛋白(例如,它的dna结合)。参见例如wo 2016/0312198,该文献以全文引用的方式并入本文用于所有目的。
[0215]
(2)转录激活域
[0216]
本文公开的嵌合衔接蛋白包括一个或多个转录激活结构域。此类转录激活结构域可以是天然存在的转录激活结构域,可以是天然存在的转录激活结构域的功能片段或功能变体,或者可以是工程化或合成的转录激活结构域。可以使用的转录激活域包括在本文别处描述的用于嵌合cas蛋白的那些。
[0217]
用于本文公开的嵌合衔接蛋白的特定转录激活域包含p65和/或hsf1转录激活域或其功能片段或变体。hsf1转录激活域可以是人热休克因子1(hsf1)的转录激活域。p65转录激活域可以是转录因子p65的转录激活域,也称为核因子nf-κ-b p65亚基,由rela基因编码。作为一个实例,转录激活域可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:8所示的p65转录激活域蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。作为另一个实例,转录激活域可以包含以下氨基酸序列、基本上由以下氨基酸序列组成,或由以下氨基酸序列组成:与seq id no:9所示的hsf1转录激活域蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列。
[0218]
c.sam向导rna
[0219]
还提供了可以结合本文别处公开的嵌合cas蛋白和嵌合衔接蛋白以激活靶基因转录的向导rna。
[0220]
一个或多个向导rna可用于本文公开的方法或组合物。例如,可以使用两个或更多个、三个或更多个、四个或更多个,或五个或更多个向导rna。两个或更多个向导rna可以靶向单个靶基因中的不同靶序列。例如,两个或更多个、三个或更多个、四个或更多个,或五个或更多个向导rna可以各自靶向单个靶基因中的不同靶序列。类似地,向导rna可以靶向多个靶基因(例如,两个或更多个、三个或更多个、四个或更多个,或五个或更多个靶基因)。向导rna靶序列的实例在本文别处公开。
[0221]
(1)向导rna
[0222]“向导rna”或“grna”是与cas蛋白(例如,cas9蛋白)结合并将cas蛋白靶向靶dna内的特定位置的rna分子。向导rna可以包含两个区段:“dna靶向区段”(也称为“向导序列”)和“蛋白结合区段”。“区段”包括分子的一部分或区域,如rna中的核苷酸的连续段。一些grna,如cas9的那些,可以包括两个单独的rna分子:“激活因子rna”(例如,tracrrna)和“靶向因子rna”(例如,crispr rna或crrna)。其它grna是单个rna分子(单个rna多核苷酸),其也可以被称为“单个分子grna”、“单个向导rna”或“sgrna”。参见例如wo 2013/176772、wo 2014/065596、wo 2014/089290、wo2014/093622、wo 2014/099750、wo 2013/142578和wo 2014/131833,它们中的每一个以全文引用的方式并入本文用于所有目的。向导rna可以指crispr rna(crrna)或crrna和反式激活crispr rna(tracrrna)的组合。crrna和tracrrna可以结合为单个rna分子(单个向导rna或sgrna)或在两个单独的rna分子(双向导rna或dgrna)中结
合。例如,对于cas9,单向导rna可以包括(例如,通过连接子)与tracrrna融合的crrna。例如,对于cpf1,只需要crrna就可以实现与靶序列的结合。术语“向导rna”和“grna”包含双分子(即,模块化)grna和单分子grna两者。在本文公开的一些方法和组合物中,c5grna是酿脓链球菌cas9 grna或其等价物。在本文公开的一些方法和组合物中,c5grna是金黄色葡萄球菌cas9 grna或其等价物。
[0223]
示范性双分子grna包含crrna样(“crispr rna”或“靶向子rna”或“crrna”或“crrna重复序列”)分子和对应的tracrrna样(“反式激活crispr rna”或“激活子rna”)或“tracrrna”)分子。crrna包括grna的dna靶向片段(单链)和核苷酸段,所述核苷酸段形成grna的蛋白质结合片段的dsrna双链体的一半。定位在dna靶向区段下游(3')的crrna尾的实例包含guuuuagagcuaugcu(seq id no:10)、基本上由其组成或由其组成。本文公开的dna靶向区段中的任一个都可以与seq id no:10的5'末端连接以形成crrna。
[0224]
对应的tracrrna(激活因子rna)包括形成grna的蛋白质结合区段的dsrna双链体的另一半的核苷酸段。crrna的核苷酸段与tracrrna的核苷酸段互补并且与其杂交,以形成grna的蛋白质结合结构域的dsrna双链体。因此,每个crrna可以被视为具有对应的tracrrna。tracrrna序列的实例包含agcauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuu(seq id no:11)、aaacagcauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu(seq id no:50)、guuggaaccauucaaaacagcauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugc(seq id no:51)中的任一个、基本上由其组成或由其组成。
[0225]
在需要crrna和tracrrna两者的系统中,crrna和对应的tracrrna杂交以形成grna。在仅需要crrna的系统中,crrna可以是grna。crrna另外提供与靶dna的互补链杂交的单链dna靶向区段。如果用于细胞内的修饰,则给定的crrna或tracrrna分子的确切序列可以被设计成对将使用rna分子的物种具有特异性。参见例如mali等人,(2013)《科学》339(6121):823-826;jinek等人,(2012)《科学》337(6096):816-821;hwang等人,(2013)《自然生物技术》31(3):227-229;jiang等人,(2013)《自然生物技术》31(3):233-239;以及cong等人,(2013)《科学》339(6121):819-823,它们中的每一个以全文引用的方式并入本文用于所有目的。
[0226]
给定grna的dna靶向区段(crrna)包括与靶dna的互补链上的序列互补的核苷酸序列,如下文更详细地描述的。grna的dna靶向区段通过杂交(即,碱基配对)以序列特异性方式与靶dna相互作用。因此,dna靶向区段的核苷酸序列可以不同,并且测定grna和靶dna将与其相互作用的靶dna内的定位。可以修饰主题grna的dna靶向区段以与靶dna内的任何期望序列杂交。天然存在的crrna因crispr/cas系统和生物体而不同,但通常含有长度为21到72个核苷酸的侧接有长度为21到46个核苷酸的两个直接重复序列(dr)的靶向区段(参见例如,wo 2014/131833,所述文献出于所有目的通过引用整体并入本文)。在酿脓链球菌的情况下,dr的长度为36个核苷酸,并且靶向区段的长度为30个核苷酸。定位于3'的dr与对应的tracrrna互补并杂交,后者进而与cas蛋白结合。
[0227]
dna靶向区段的长度可以为例如至少约12、15、17、18、19、20、25、30、35或40个核苷酸。此类dna靶向区段的长度可以为例如约12到约100、约12到约80、约12到约50、约12到约40、约12到约30、约12到约25或约12到约20个核苷酸。例如,dna靶向区段可以为约15到约25
个核苷酸(例如,约17到约20个核苷酸或约17、18、19或20个核苷酸)。参见例如us 2016/0024523,所述文献出于所有目的通过引用整体并入本文。对于来自酿脓链球菌的cas9,典型的dna靶向区段的长度介于16与20个核苷酸之间或长度介于17与20个核苷酸之间。对于来自金黄色葡萄球菌的cas9,典型的dna靶向区段的长度介于21与23个核苷酸之间。对于cpf1,典型的dna靶向区段的长度为至少16个核苷酸或至少18个核苷酸。
[0228]
在一个实例中,dna靶向区段的长度可以为约20个核苷酸。然而,更短和更长的序列也可用于靶向区段(例如,长度为15至25个核苷酸,如15、16、17、18、19、20、21、22、23、24或25个核苷酸)。dna靶向区段和对应的向导rna靶序列之间的相同程度(或dna靶向区段和向导rna靶序列的另一链之间的互补程度)可以是例如约75%、约80%、约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%。dna靶向区段和对应的向导rna靶序列可以含有一个或多个错配。例如,向导rna的dna靶向区段和对应的向导rna靶序列可以含有1至4、1至3、1至2、1、2、3或4个错配(例如,其中向导rna靶序列的总长度为至少17个、至少18个、至少19个或至少20个或更多个核苷酸)。例如,向导rna的dna靶向区段和对应的向导rna靶序列可以含有1至4、1至3、1至2、1、2、3或4个错配,其中向导rna靶序列的总长度为20个核苷酸。
[0229]
tracrrna可以呈任何形式(例如,全长tracrrna或活性部分tracrrna)并具有不同长度。tracrrna可以包含初级转录本或经处理的形式。例如,tracrrna(作为单向导rna的一部分或作为作为双分子grna的一部分的单独分子)可以包括野生型tracrrna序列(例如,野生型tracrrna序列的约或超过约20、26、32、45、48、54、63、67、85或更多个核苷酸)、基本上由其组成或由其组成。来自酿脓链球菌的野生型tracrrna序列的实例包含171-核苷酸、89-核苷酸、75-核苷酸和65-核苷酸版本。参见例如deltcheva等人(2011)《自然》471(7340):602-607;wo 2014/093661,所述文献中的每个文献出于所有目的通过引用整体并入本文。单向导rna(sgrna)内的tracrrna的实例包含在 48、 54、 67和 85版本的sgrna中发现的tracrrna区段,其中“ n”指示野生型tracrrna的至多 n个核苷酸包含在sgrna中。参见us 8,697,359,所述文献出于所有目的通过引用整体并入本文。
[0230]
向导rna的dna靶向区段与靶dna的互补链之间的互补性百分比可以为至少60%(例如,至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少98%、至少99%或100%)。dna靶向片段与靶dna的互补链之间的互补性百分比在约20个连续核苷酸上可以为至少60%。举例来说,dna靶向区段与靶dna的互补链之间的互补性百分比在靶dna的互补链的5'末端处的14个连续核苷酸上可以为100%,并且在剩余部分上低至0%。在此类情况下,dna靶向片段可以被视为长度为14个核苷酸。作为另一个实例,dna靶向区段与靶dna的互补链之间的互补性百分比在靶dna的互补链的5'末端处的七个连续核苷酸上可以为100%,并且在剩余部分上低至0%。在此类情况下,dna靶向区段可以被视为长度为7个核苷酸。在一些向导rna中,dna靶向区段内的至少17个核苷酸与靶dna的互补链互补。例如,dna靶向区段的长度可以是20个核苷酸,并且可以包括与靶dna的互补链的1、2或3个错配。在一个实例中,错配不与对应于原型间隔子相邻基序(pam)序列的互补链的区域(即,pam序列的反向补体)相邻(例如,错配处于向导rna的dna靶向区段的5'末端中,或者错配距离对应于pam序列的互补链的区域至少2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18或19个碱基对)。
[0231]
grna的蛋白质结合区段可以包括彼此互补的两个核苷酸段。蛋白质结合区段的互补核苷酸杂交形成双链rna双链体(dsrna)。主题grna的蛋白质结合片段与cas蛋白相互作用,并且grna通过dna靶向片段将结合的cas蛋白引导到靶dna内的特异性核苷酸序列。
[0232]
单向导rna可以包括dna靶向区段和支架序列(即,向导rna的蛋白质结合或cas结合序列)。例如,此类向导rna可以具有与3'支架序列连接的5'dna靶向区段。示范性支架序列包含以下各项、基本上由以下各项组成或由以下各项组成:guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcu(版本1;seq id no:12);guuggaaccauucaaaacagcauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugc(版本2;seq id no:13);guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugc(版本3;seq id no:14);guuuaagagcuaugcuggaaacagcauagcaaguuuaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugc(版本4;seq id no:15);guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuuuuu(版本5;seq id no:52);guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu(版本6;seq id no:53);或guuuaagagcuaugcuggaaacagcauagcaaguuuaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuuuu(版本7;seq id no:54)。靶向本文公开的向导rna靶序列中的任一个的向导rna可以包括例如与向导rna的3'末端上的示范性向导rna支架序列中的任一个融合的向导rna的5'末端上的dna靶向区段。即,本文所公开的dna靶向区段中的任何dna靶向区段可以与上述支架序列中的任何支架序列的5'末端连接以形成单向导rna(嵌合向导rna)。
[0233]
向导rna可以包含提供另外的期望特征的修饰或序列(例如,经修饰或经调控的稳定性;亚细胞靶向;用荧光标记追踪;蛋白质或蛋白质复合物的结合位点;等等)。也就是说,向导rna可以包括一个或多个经修饰核苷或核苷酸,或一个或多个非天然和/或天然存在的组分或构型,其用于代替或补充标准的a、g、c和u残基.此类修饰的实例包括例如5'帽(例如,7-甲基鸟苷酸帽(m7g));3'聚腺苷酸化尾(即,3'聚(a)尾);核糖开关序列(例如,以允许调控稳定性和/或调控蛋白和/或蛋白复合物的可及性);稳定性控制序列;形成dsrna双链体(即,发夹)的序列;将rna靶向亚细胞位置(例如,细胞核、线粒体、叶绿体等等)的修饰或序列;提供追踪的修饰或序列(例如,与荧光分子直接缀合、与促进荧光检测的部分缀合、允许荧光检测的序列等);为蛋白(例如,作用于dna的蛋白,如转录激活子)提供结合位点的修饰或序列;及其组合。修饰的其它实例包含工程化茎环双链体结构、工程化凸起区、茎环双链体结构的工程化发夹3'或其任何组合。参见例如us 2015/0376586,所述文献出于所有目的通过引用整体并入本文。凸起可以是由crrna样区域和最小tracrrna样区域构成的双链体内的核苷酸的未配对区域。凸起在双链体的一侧可以包括未配对的5'-xxxy-3',其中x是任何嘌呤,并且y可以包括可以与相对链上的核苷酸形成摇摆对的核苷酸;并且在双链体的另一侧包括未配对核苷酸区。
[0234]
未经修饰的核酸可能易于降解。外源性核酸也可以诱导先天免疫应答。修饰可以有助于引入稳定性并降低免疫原性。向导rna可以包含经修饰的核苷和经修饰的核苷酸,包括例如以下各项中的一个或多个:(1)磷酸二酯主链键中的非连接磷酸氧中的一个或两个和/或连接磷酸氧中的一个或多个的改变或替换(示范性主链修饰);(2)核糖的组成的改变或替换,如核糖上的2'羟基的改变或替换(示范性糖修饰);(3)用脱磷接头替换(例如,批量
替换)磷酸酯部分(示范性主链修饰);(4)天然存在的核碱基的修饰或替换,包括用非标准核碱基(示范性碱基修饰);(5)核糖-磷酸酯主链的替换或修饰(示范性主链修饰);(6)寡核苷酸的3'末端或5'末端的修饰(例如,末端磷酸基团的去除、修饰或替换,或部分、帽或接头的缀合(此类3'或5'帽修饰可以包含糖和/或主链修饰));以及(7)糖的修饰或替换(示范性糖修饰)。其它可能的向导rna修饰包含尿嘧啶或聚尿嘧啶束的修饰或替代。参见例如wo2015/048577和us 2016/0237455,所述文献中的每个文献出于所有目的通过引用整体并入本文。可以对如cas mrna等cas编码核酸进行类似的修饰。例如,可以通过使用同义密码子耗尽尿苷来修饰cas mrna。
[0235]
可以组合如上所列出的化学修饰以提供经修饰的grna和/或mrna,其包含可具有两个、三个、四个或更多个修饰的残基(核苷和核苷酸)。例如,经修饰残基可以具有经修饰的糖和经修饰的核碱基。在一个实例中,grna的每个碱基均被修饰(例如,所有碱基均具有经修饰的磷酸基团,如硫代磷酸酯基团)。例如,grna的所有或基本上所有磷酸基团均可以被硫代磷酸酯基团替换。替代地或附加地,经修饰的grna可以在5'末端处或其附近包含至少一个经修饰残基。替代地或附加地,经修饰的grna可以在3'末端处或其附近包含至少一个经修饰残基。
[0236]
一些grna包含一个、两个、三个或更多个经修饰残基。例如,经修饰的grna中至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%或100%的位置可以是经修饰的核苷或核苷酸。
[0237]
未经修饰的核酸可能易于降解。外源性核酸也可以诱导先天免疫应答。修饰可以有助于引入稳定性并降低免疫原性。本文所述的一些grna可以含有一个或多个经修饰的核苷或核苷酸以向细胞内或基于血清的核酸酶引入稳定性。当引入细胞群时,本文所述的一些经修饰的grna可以表现出降低的先天免疫响应。
[0238]
本文公开的grna可以包含主链修饰,其中经修饰残基的磷酸基团可以通过用不同的取代基替换氧中的一个或多个来修饰。修饰可以包括用如本文所述的经修饰的磷酸基团批量替换未经修饰的磷酸酯部分。磷酸酯主链的主链修饰还可以包括导致不带电接头或具有不对称电荷分布的带电接头的改变。
[0239]
经修饰磷酸基团的实例包括硫代磷酸酯、硒代磷酸酯、硼酸磷酸酯、硼酸磷酸酯、膦酸氢酯、亚氨代磷酸酯、烷基或芳基膦酸酯和磷酸三酯。未经修饰的磷酸基团中的磷原子是非手性的。然而,用上述原子或原子团中的一个替换非桥接氧中的一个可以使磷原子手性。立体异构磷原子可以具有“r”构型(rp)或“s”构型(sp)。主链也可以通过用氮(桥接的亚氨代磷酸酯)、硫(桥接的硫代磷酸酯)和碳(桥接的亚甲基膦酸酯)替换桥接氧(即,将磷酸酯与核苷链接的氧)来修饰。替换可以发生在链接氧处或两个链接氧处。
[0240]
在某些主链修饰中,磷酸基团可以被不含磷的连接头替换。在一些实施例中,带电荷的磷酸基团可以被中性部分替换。可以替换磷酸基团的部分的实例可以包括但不限于例如膦酸甲酯、羟氨基、硅氧烷、碳酸酯、羧甲基、氨基甲酸酯、酰胺、硫醚、环氧乙烷接头、磺酸酯、磺酰胺、硫代甲缩醛、甲缩醛、肟、亚甲基亚氨基、亚甲基甲基亚氨基、亚甲基肼基、亚甲基二甲基肼基和亚甲基氧基甲基亚氨基。
[0241]
也可以构建可以模拟核酸的支架,其中磷酸酯接头和核糖被核酸酶抗性核苷或核
苷酸替代物替换。此类修饰可以包含主链和糖修饰。在一些实施例中,核碱基可以由替代主链拴系。实例可以包括但不限于吗啉代、环丁基、吡咯烷和肽核酸(pna)核苷替代物。
[0242]
经修饰的核苷和经修饰的核苷酸可以包括对糖基团的一个或多个修饰(糖修饰)。例如,2'羟基(oh)可以被修饰(例如,被多个不同的氧或脱氧取代基替换。对2'羟基的修饰可以增强核酸的稳定性,因为羟基不能再被去质子化以形成2'-醇盐离子。
[0243]
2'羟基修饰的实例可以包括烷氧基或芳氧基(or,其中“r”可以是例如烷基、环烷基、芳基、芳烷基、杂芳基或糖);聚乙二醇(peg)、o(ch2ch2o)nch2ch2or,其中r可以是例如h或任选取代的烷基,并且n可以是0至20的整数(例如0至4、0至8、0至10、0至16、1至4、1至8、1至10、1至16、1至20、2至4、2至8、2至10、2至16、2至20、4至8、4至10、4至16和4至20)。2'羟基修饰可以是2'-o-me。同样地,2'羟基修饰可以是2'-氟修饰,其用氟化物替换2'羟基。2'羟基修饰可以包括锁定核酸(lna),其中2'羟基可以例如通过c
1-6
亚烷基或c
1-6
杂亚烷基桥接到相同核糖的4'碳,其中示范性桥可以包括亚甲基、丙烯、醚或氨基桥;o-氨基(其中氨基可以是例如nh2;烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基或二杂芳基氨基、乙二胺或聚氨基)和氨基烷氧基、o(ch2)
n-氨基(其中氨基可以是例如nh2;烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基或二杂芳基氨基、乙二胺或聚氨基)。2'羟基修饰可以包括未锁定的核酸(una),其中核糖环缺少c2'-c3'键。2'羟基修饰可以包括甲氧基乙基(moe),(och2ch2och3,例如,peg衍生物)。
[0244]
脱氧2'修饰可以包括氢(即,脱氧核糖,例如,在部分dsrna的突出部分);卤素(例如溴、氯、氟或碘);氨基(其中氨基可以是例如nh2;烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基、二杂芳基氨基或氨基酸);nh(ch2ch2nh)nch2ch
2-氨基(其中氨基可以是,例如,如本文所述)、-nhc(o)r(其中r可以是,例如,烷基、环烷基、芳基、芳烷基,杂芳基或糖)、氰基;巯基;烷基硫代烷基;硫代烷氧基;以及烷基、环烷基、芳基、烯基和炔基,它们可以任选地被例如本文所述的氨基取代。
[0245]
糖修饰可以包含糖基团,该糖基团还可以含有具有与核糖中对应碳的立体化学构型相反的立体化学构型的一个或多个碳。因此,经修饰的核酸可以包括含有例如阿拉伯糖作为糖的核苷酸。经修饰的核酸还可以包括脱碱基糖。这些脱碱基糖也可以在组成糖原子中的一个或多个处被进一步修饰。经修饰的核酸还可以包括呈l形式的一个或多个糖(例如l-核苷)。
[0246]
本文所述的经修饰核苷和经修饰核苷酸(其可掺入经修饰核酸中)可以包括经修饰的碱基,也称为核碱基。核碱基的实例包括但不限于腺嘌呤(a)、鸟嘌呤(g)、胞嘧啶(c)和尿嘧啶(u)。这些核碱基可以被修饰或完全替换以提供可以掺入经修饰核酸中的经修饰残基。核苷酸的核碱基可以独立地选自嘌呤、嘧啶、嘌呤类似物或嘧啶类似物。在一些实施例中,核碱基可以包括例如碱基的天然存在和合成衍生物。
[0247]
在双向导rna中,crrna和tracrrna中的每一个均可以含有修饰。此类修饰可以在crrna和/或tracrrna的一端或两端。在sgrna中,sgrna的一端或两端的一个或多个残基可以被化学修饰,和/或内部核苷可以被修饰,和/或整个sgrna可以被化学修饰。一些grna包含5'末端修饰。一些grna包含3'末端修饰。
[0248]
本文公开的向导rna可以包含wo 2018/107028 a1中公开的修饰模式中的一个,该文献以全文引用的方式并入本文用于所有目的。本文公开的向导rna还可以包含us2017/
0114334中公开的结构/修饰模式中的一个,该文献以全文引用的方式并入本文用于所有目的。本文公开的向导rna还可以包含在wo 2017/136794、wo 2017/004279、us2018/0187186或us 2019/0048338中公开的结构/修饰模式中的一个,它们中的每一个以全文引用的方式并入本文用于所有目的。
[0249]
举例来说,向导rna的5'末端或3'末端的核苷酸可以包含硫代磷酸酯键(例如,碱基可以具有经修饰的磷酸酯基,所述经修饰的磷酸酯基是硫代磷酸酯基)。例如,向导rna可以包含向导rna的5'或3'末端处的2、3或4个末端核苷酸之间的硫代磷酸酯键。作为另一个实例,向导rna的5'和/或3'末端的核苷酸可以具有2'-o-甲基修饰。例如,向导rna可以包含向导rna的5'和/或3'末端(例如,5'末端)的2、3或4个端核苷酸处的2'-o-甲基修饰。参见例如wo 2017/173054 a1和finn等人(2018)《细胞报告(cell rep.)》22(9):2227-2235,它们中的每一个以全文引用的方式并入本文用于所有目的。其它可能的修饰在本文别处更详细地描述。在具体实例中,向导rna包含前三个5'和3'末端rna残基处的2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。此类化学修饰可以例如提供更大的稳定性和防止核酸外切酶向导rna,允许所述向导rna比未经修饰的向导rna在细胞内持续更长时间。例如,此类化学修饰还可以防止先天的细胞内免疫应答,所述先天的细胞内免疫应答可以主动降解rna或触发导致细胞死亡的免疫级联反应。
[0250]
作为一个实例,本文所述的向导rna中的任一个可以包含至少一个修饰。在一个实例中,该至少一个修饰包含2'-o-甲基(2'-o-me)修饰的核苷酸、核苷酸之间的硫代磷酸酯(ps)键、2'-氟(2'-f)修饰的核苷酸,或其组合。例如,该至少一个修饰可以包含2'-o-甲基(2'-o-me)修饰的核苷酸。替代地或附加地,该至少一个修饰可以包含核苷酸之间的硫代磷酸酯(ps)键。替代地或附加地,该至少一个修饰可以包含2'-氟(2'-f)修饰的核苷酸。在一个实例中,本文所述的向导rna包含一个或多个2'-o-甲基(2'-o-me)修饰的核苷酸和核苷酸之间的一个或多个硫代磷酸酯(ps)键。
[0251]
修饰可以发生在向导rna中的任何地方。作为一个实例,向导rna包含在向导rna的5'末端的前五个核苷酸中的一个或多个处的修饰,向导rna包含在向导rna的3'末端的最后五个核苷酸中的一个或多个处的修饰,或其组合。例如,向导rna可以包含向导rna的前四个核苷酸之间的硫代磷酸酯键、向导rna的最后四个核苷酸之间的硫代磷酸酯键,或其组合。替代地或附加地,向导rna可以包含在向导rna的5'末端的前三个核苷酸处的2'-o-me修饰的核苷酸,可以包含在向导rna的3'末端的最后三个核苷酸处的2'-o-me修饰的核苷酸,或其组合。
[0252]
已显示影响核苷酸糖环的另一种化学修饰是卤素取代。例如,核苷酸糖环上的2'-氟(2'-f)取代可以增加寡核苷酸结合亲和力和核酸酶稳定性。脱碱基核苷酸是指那些缺乏含氮碱基的核苷酸。反向碱基是指具有从正常5'至3'键反向的键(即,5'至5'键或3'至3'键)的碱基。
[0253]
脱碱基核苷酸可以通过反向键连接。例如,脱碱基核苷酸可经由5'至5'键连接至末端5'核苷酸,或脱碱基核苷酸可经由3'至3'键连接至末端3'核苷酸。在末端5'或3'核苷酸处的反向脱碱基核苷酸也可称为反向脱碱基端帽。
[0254]
在一个实例中,5'端处的前三个、四个或五个核苷酸中的一个或多个,以及3'端处的最后三个、四个或五个核苷酸中的一个或多个被修饰。修饰可以是,例如,2'-o-me、2'-f、
反向脱碱基核苷酸、硫代磷酸酯键或其它众所周知的提高稳定性和/或性能的核苷酸修饰。
[0255]
在另一个实例中,5'端处的前四个核苷酸和3'端处的最后四个核苷酸可以用硫代磷酸酯键链接。
[0256]
在另一个实例中,5'端处的前三个核苷酸和3'端处的最后三个核苷酸可以包含2'-o-甲基(2'-o-me)修饰的核苷酸。在另一个实例中,5'端处的前三个核苷酸和3'端处的最后三个核苷酸包含2'-氟(2'-f)修饰的核苷酸。在另一个实例中,5'端处的前三个核苷酸和3'端处的最后三个核苷酸包含反向脱碱基核苷酸。
[0257]
在一些向导rna(例如,单向导rna)中,向导rna的至少一个环(例如,两个环)通过插入与一个或多个衔接子(即,衔接蛋白或结构域)结合的不同rna序列而被修饰.此类衔接蛋白可以用于进一步募集一个或多个异源功能域,如转录激活域。包括此类衔接蛋白(即,嵌合衔接蛋白)的融合蛋白的实例在本文别处公开。例如,ms2结合环ggccaacaugaggaucacccaugucugcagggcc(seq id no:16)可以替换seq id no:12、14、52或53所示的sgrna支架(主链)或wo 2016/049258和konermann等人(2015)《自然》517(7536):583-588中所描述的酿脓链球菌crispr/cas9系统的sgrna主链的核苷酸 13至 16和核苷酸 53至 56,这些文献中的每一个以全文引用的方式并入本文用于所有目的。参见例如图3。本文所使用的向导rna编号是指向导rna支架序列(即,向导rna的dna靶向区段下游的序列)中的核苷酸编号。例如,向导rna支架的第一个核苷酸是 1,支架的第二个核苷酸是 2,等等。与seq id no:12、14、52或53中的核苷酸 13至 16相对应的残基是跨越seq id no:12、14、52或53中的核苷酸 9至 21的区域(本文中被称为四环的区域)中的环序列。与seq id no:12、14、52或53中的核苷酸 53至 56相对应的残基是跨越seq id no:12、14、52或53中的核苷酸 48至 61的区域(本文中被称为茎环2的区域)中的环序列。seq id no:12、14、52或53中的其它茎环序列包含茎环1(核苷酸 33至 41)和茎环3(核苷酸 63至 75)。所得结构是其中四环和茎环2序列中的每一个都被ms2结合环替代的sgrna支架。四环和茎环2以使得添加ms2结合环不应干扰任何cas9残基的方式从cas9蛋白中突出。另外地,四环和茎环2位点与dna的接近表明定位到这些定位可能导致dna与任何募集的蛋白质如转录激活因子之间的高度相互作用。因此,在一些sgrna中,对应于seq id no:12、14、52或53所示的向导rna支架的 13至 16的核苷酸和/或对应于 53至 56的核苷酸或对应残基在与这些支架/主链中的任一个支架/主链最佳比对时被能够与一个或多个衔接蛋白或域结合的不同rna序列替换。可替代地或另外地,可以将衔接结合序列添加到向导rna的5'端或3'端。在四环和茎环2区域中包含ms2结合环的示范性向导rna支架可以包含seq id no:40或56所示的序列、基本上由其组成或由其组成。在四环和茎环2区域中包含ms2结合环的示范性通用单个向导rna可以包含seq id no:45或57所示的序列、基本上由其组成或由其组成。
[0258]
可以以任何形式提供向导rna。例如,grna可以以rna的形式提供,作为两个分子(单独的crrna和tracrrna)或作为一个分子(sgrna),并且任选地以与cas蛋白的复合物的形式提供。grna也可以以对grna进行编码的dna的形式提供。对grna进行编码的dna可以编码单个rna分子(sgrna)或单独的rna分子(例如,单独的crrna和tracrrna)。在后一种情况下,对grna进行编码的dna可以作为一个dna分子或作为分别对crrna和tracrrna进行编码的单独dna分子提供。
[0259]
当grna以dna的形式提供时,grna可以在细胞中瞬时、有条件或组成型表达。对
grna进行编码的dna可以稳定地整合到细胞的基因组中,并且与在细胞中具有活性的启动子可操作地连接。可替代地,对grna进行编码的dna可以与表达构建体中的启动子可操作地连接。例如,编码grna的dna可以处于包含异源核酸的载体中。可以用于此类表达构建体的启动子包括在例如真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、多能细胞、胚胎干(es)细胞、成体干细胞、发育受限的祖细胞、诱导的多能干(ips)细胞或单细胞期胚胎中的一个或多个中具有活性的启动子。此类启动子可以是例如条件启动子、诱导型启动子、组成型启动子或组织特异性启动子。此类启动子也可以是例如双向启动子。合适启动子的具体实例包含rna聚合酶iii启动子,如人u6启动子、大鼠u6聚合酶iii启动子或小鼠u6聚合酶iii启动子。
[0260]
可替代地,可以通过各种其它方法制备grna。例如,可以使用例如t7 rna聚合酶通过体外转录制备grna(参见例如,wo 2014/089290和wo 2014/065596,所述文献中的每个文献出于所有目的通过引用整体并入本文)。向导rna也可以是通过化学合成制备的合成产生的分子。例如,可以化学合成向导rna以在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。
[0261]
向导rna(或对向导rna进行编码的核酸)可以在包括一个或多个向导rna(例如,1个、2个、3个、4个或更多个向导rna)和增加向导rna稳定性(例如,延长在给定储存条件(例如,-20℃、4℃或环境温度)下降解产物保持在阈值以下的时间,如低于起始核酸或蛋白质重量的0.5%;或增加体内稳定性)的载体的组合物中。此类载体的非限制性实例包含聚乳酸(pla)微球体、聚(d,l-乳酸-乙醇酸共聚物)(plga)微球体、脂质体、胶束、反胶束、脂质螺旋体和脂质微管。此类组合物可以进一步包括cas蛋白,如cas9蛋白,或对cas蛋白进行编码的核酸。
[0262]
(2)向导rna靶序列
[0263]
用于向导rna的靶dna包含存在于dna中的核酸序列,grna的dna靶向区段将与其结合,前提是存在足够的结合条件。合适的dna/rna结合条件包含细胞中通常存在的生理条件。其它合适的dna/rna结合条件(例如,无细胞系统中的条件)是本领域已知的(参见例如,《分子克隆:实验室手册》,第3版(sambrook等人,海港实验室出版社(harbor laboratory press)2001),所述文献出于所有目的通过引用整体并入本文)。与grna互补并杂交的靶dna的链可以被称为“互补链”,并且与“互补链”互补(并且因此不与cas蛋白或grna互补)的靶dna的链可以称为“非互补链”或“模板链”。
[0264]
靶dna包含与向导rna杂交的互补链上的序列和非互补链上的对应序列(例如,与前间区序列邻近基序(pam)邻近)。如本文所使用的,术语“向导rna靶序列”具体指非互补链上的与向导rna在互补链上与其杂交的序列相对应(即,其反向补体)的序列。即,向导rna靶序列是指非互补链上的与pam相邻(例如,在cas9的情况下,pam的上游或5')的序列。向导rna靶序列等同于向导rna的dna靶向区段,但具有胸腺嘧啶而不是尿嘧啶。举例来说,spcas9酶的向导rna靶序列可以指非互补链上的5'-ngg-3'pam上游的序列。向导rna被设计成与靶dna的互补链互补,其中向导rna的dna靶向区段与靶dna的互补链之间的杂交促进了crispr复合物的形成。不一定需要完全互补,条件是存在足够的互补性来引起杂交并促进crispr复合物的形成。如果向导rna在本文中被称为靶向向导rna靶序列,则意味着向导rna与靶dna的互补链序列杂交,所述互补链序列是非互补链上的向导rna靶序列的反向补体。
[0265]
靶dna或向导rna靶序列可以包括任何多核苷酸,并且可以定位于例如细胞的细胞核或细胞质中或细胞的细胞器如线粒体或叶绿体内。靶dna或向导rna靶序列可以是细胞内源性或外源性的任何核酸序列。向导rna靶序列可以是编码基因产物(例如,蛋白质)的序列或非编码序列(例如,调节序列)或者可以包含两者。
[0266]
优选的可以是靶序列与基因的转录起始位点邻近。例如,靶序列可以在转录起始位点1000、900、800、700、600、500、400、300、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内、在转录起始位点上游1000、900、800、700、600、500、400、300、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内,或者在转录起始位点下游1000、900、800、700、600、500、400、300、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、10、5或1个碱基对内。任选地,靶序列处于转录起始位点上游200个碱基对和转录起始位点下游1个碱基对的区域内(-200到 1)。
[0267]
靶序列可以在希望被靶向以用于转录激活的任何基因内。在一些情况下,靶基因可以是非表达基因或弱表达基因(例如,仅在背景之上最低限度地表达,如1.1倍、1.2倍、1.3倍、1.4倍、1.5倍、1.6倍、1.7倍、1.8倍、1.9倍或2倍)。与对照基因相比,靶基因也可以是低水平表达的基因。靶基因也可以是表观遗传沉默的基因。术语“表观遗传沉默”是指由于遗传变化以外的机制(如突变),没有被转录或被转录的水平相对于对照样品(例如,对应的对照细胞,如正常细胞)中的基因的转录水平降低的基因。基因沉默的表观遗传机制是众所周知的,并且包括例如基因的5'调控区域的cpg岛中cpg二核苷酸的高甲基化和由于例如组蛋白乙酰化引起的染色质中的结构变化,使得基因转录被减少或抑制。
[0268]
靶基因可以包括在特定器官或组织(如肝脏)中表达的基因。靶基因可以包括疾病相关联基因。疾病相关联基因是指与未患病对照的组织或细胞相比,在来源于受疾病影响的组织的细胞中以异常水平或异常形式产生转录或翻译产物的任何基因。它可能是以异常高水平表达的基因,其中改变的表达与疾病的发生和/或发展相关。疾病相关联基因也指具有导致疾病病因的突变或遗传变异的基因。转录或翻译的产物可能是已知的或未知的,并且可能处于正常或异常水平。例如,靶基因可以是与蛋白聚集疾病和病症相关联的基因,如阿尔茨海默病、帕金森病、亨廷顿病、肌萎缩侧索硬化症、朊病毒病和淀粉样变性如转甲状腺素蛋白淀粉样变性(例如,ttr)。靶基因还可以是涉及与疾病或病症(如高胆固醇血症或动脉粥样硬化)相关的途径的基因,或者当过表达时可以模拟此类疾病或病症的基因。靶基因还可以是在一个或多个类型的癌症中表达或过表达的基因。参见例如santarius等人(2010)《癌症自然评论(nat.rev.cancer)》10(1):59-64,该文献以全文引用的方式并入本文用于所有目的。
[0269]
此类靶基因的一个具体实例是ttr基因。任选地,ttr基因可以包含致病突变(例如,引起淀粉样变性的突变)。此类突变的实例例如在wo 2018/007871中提供,该文献以全文引用的方式并入本文用于所有目的。示范性人ttr蛋白和示范性人ttr基因分别由uniprot id p02766和entrez gene id 7276鉴别。示范性小鼠ttr蛋白和示范性小鼠ttr基因分别由uniprot id p07309和entrez gene id 22139鉴别。转甲状腺素蛋白(ttr)是一种在血清和脑脊液中发现的蛋白质,其携带甲状腺激素和视黄醇结合蛋白至视黄醇。肝脏将ttr分泌到血液中,而脉络丛将其分泌到脑脊液中。ttr也在视网膜色素上皮中产生并且分
泌到玻璃体中。错误折叠和聚集的ttr在淀粉样蛋白疾病老年系统性淀粉样变性(ssa)、家族性淀粉样蛋白多发性神经病变(fap)和家族性淀粉样蛋白心肌病(fac)中的多个组织和器官中积聚。转甲状腺素蛋白(ttr)是一种127-氨基酸、55kda血清和脑脊液转运蛋白,主要由肝脏合成,但也由脉络丛产生。其还被称为前白蛋白、甲状腺素结合前白蛋白、attr、tbpa、cts、cts1、hel111、hst2651和palb。在其天然状态下,ttr以四聚体的形式存在。在纯合子中,同源四聚体包括相同的富含127-氨基酸β片层亚基。在杂合子中,ttr四聚体可以由变体和/或野生型亚基组成,通常以统计学方式组合。ttr负责在血清和脑脊液中携带甲状腺素(t4)和视黄醇结合的rbp(视黄醇结合蛋白)。小鼠ttr基因中的向导rna靶序列(不包括pam)的实例分别在seq id no:34、35和36中示出。seq id no:34位于ttr转录起始位点的-63(基因组坐标:build mm10,chr18, 链,20665187
–
20665209),seq id no:35位于ttr转录起始位点的-134(基因组坐标:build mm10,chr18, 链,20665116
–
20665138),并且seq id no:36位于ttr转录起始位点的-112(基因组坐标:build mm10,chr18, 链,20665138
–
20665160)。对应于分别在seq id no:34、35和36中示出的向导rna靶序列的向导rna dna靶向区段分别在seq id no:41、42和43中示出。包含这些dna靶向区段的单个向导rna的实例分别在seq id no:37、38和39或55中示出。
[0270]
疾病相关联基因还可以包括在受试者中该基因产生的增加将是有益的任何基因(例如,以治疗或预防疾病)。此类基因可以是其低表达或低水平表达与疾病、病症或综合征相关联或者成为疾病、病症或综合征的病因的基因。例如,此类靶基因的转录减少、来自此类靶基因的基因产物量减少,或来自此类靶基因的基因产物的活性降低可能与疾病相关联、可能使疾病恶化,或者可能引起疾病,使得靶基因的转录或表达增加将是有益的。此类基因的一个实例是otc(entrez gene id 5009)。此类基因的其它实例是hbg1(entrez gene id 3047)和hbg2(entrez gene id 3048)。此类基因的其它实例包括单倍体不足基因,如表2和3中的那些。
[0271]
cas蛋白对靶dna的位点特异性结合和切割可以发生在由(i)向导rna与靶dna的互补链之间的碱基配对互补性和(ii)靶dna的非互补链中的短基序(被称为原型间隔子相邻基序(pam))两者测定的定位处。pam可以侧接向导rna靶序列。任选地,向导rna靶序列可以在3'末端上侧接有pam(例如,对于cas9)。可替代地,向导rna靶序列可以在5'末端上侧接有pam(例如,对于cpf1)。例如,cas蛋白的切割位点可以是pam序列上游或下游(例如,向导rna靶序列内)的约1到约10或约2到约5个碱基对(例如,3个碱基对)。在spcas9的情况下,pam序列(即,非互补链上)可以是5'-n1gg-3',其中n1是任何dna核苷酸,并且pam紧接着靶dna的非互补链上的向导rna靶序列的3'。因此,对应于互补链上的pam的序列(即,反向补体)将是5'-ccn
2-3',其中n2是任何dna核苷酸并且紧接着向导rna的dna靶向片段在靶dna的互补链上与其杂交的序列的5'。在一些此类情况下,n1和n2可以是互补的,并且n
1-n2碱基对可以是任何碱基对(例如,n1=c并且n2=g;n1=g并且n2=c;n1=a并且n2=t;或n1=t并且n2=a)。在来自金黄色葡萄球菌的cas9的情况下,pam可以是nngrrt或nngrr,其中n可以是a、g、c或t,并且r可以是g或a。在来自空肠弯曲杆菌的cas9的情况下,pam可以是例如nnnnacac或nnnnryac,其中n可以是a、g、c或t,并且r可以是g或a。在一些情况下(例如,对于fncpf1),pam序列可以是5'末端上游并具有序列5'-ttn-3'。
[0272]
向导rna靶序列的实例是紧接在spcas9蛋白识别的ngg基序之前的20个核苷酸的
dna序列。例如,向导rna靶序列加pam的两个实例是gn
19
ngg(seq id no:17)或n
20
ngg(seq id no:18)。参见例如,wo 2014/165825,所述文献出于所有目的通过引用整体并入本文。5'末端处的鸟嘌呤可以促进细胞中rna聚合酶的转录。向导rna靶序列加pam的其它实例可以包括5'末端处的两个鸟嘌呤核苷酸(例如,ggn
20
ngg;seq id no:19)以促进体外t7聚合酶的高效转录。参见例如,wo 2014/065596,所述文献出于所有目的通过引用整体并入本文。其它向导rna靶序列加pam可以具有长度介于4至22个核苷酸之间的seq id no:17至19,包括5'g或gg和3'gg或ngg。又其它向导rna靶序列加pam可以具有长度介于14至20个核苷酸之间的seq id no:17至19。
[0273]
与靶dna杂交的crispr复合物的形成可以导致在对应于向导rna靶序列(即,靶dna的非互补链上的向导rna靶序列和互补链上的向导rna与其杂交的反向补体)的区域内或附近的靶dna的一条或两条链的切割。例如,切割位点可以处于向导rna靶序列内(例如,处于相对于pam序列的所定义的定位处)。“切割位点”包含cas蛋白产生单链断裂或双链断裂的靶dna的位置。切割位点可以仅在双链dna的一条链上(例如,当使用切口酶时)或在两条链上。切割位点可以在两条链上的相同位置处(产生平端;例如,cas9))或可以在每条链上的不同位点处(产生交错末端(即,突出端);例如,cpf1)。例如,可以通过使用两种cas蛋白产生交错末端,其中每一种在不同链的不同切割位点处产生单链断裂,从而产生双链断裂。例如,第一切口酶可以在双链dna(dsdna)的第一条链上产生单链断裂,并且第二切口酶可以在dsdna的第二条链上产生单链断裂,使得产生突出序列。在一些情况下,第一链上的切口酶的向导rna靶序列或切割位点与第二链上的切口酶的向导rna靶序列或切割位点相隔至少2个、3个、4个、5个、6个、7个、8个、9个、10个、15个、20个、25个、30个、40个、50个、75个、100个、250个、500个或1,000个碱基对。
[0274]
d.编码嵌合cas蛋白、嵌合衔接蛋白、向导rna或协同激活介体的核酸
[0275]
本文别处详细描述的嵌合cas蛋白、嵌合衔接蛋白和向导rna可以在本文公开的方法和组合物中以核酸(例如,dna或rna)的形式提供。例如,核酸可以是嵌合cas蛋白表达盒、嵌合衔接蛋白表达盒、协同激活介体(sam)表达盒(其包含编码嵌合cas蛋白和嵌合衔接蛋白两者的核酸)、向导rna表达盒,或其任何组合。此类核酸可以是rna(例如,信使rna(mrna))或dna,可以是单链或双链的,并且可以是线性的或环状的。例如,核酸可以是嵌合cas蛋白mrna、嵌合衔接蛋白mrna、协同激活介体(sam)mrna(其包含编码嵌合cas蛋白和嵌合衔接蛋白两者的核酸)、向导rna,或其任何组合。dna可以是如表达载体或靶向载体等载体的一部分。载体还可以是病毒载体,如腺病毒、腺关联病毒、慢病毒和逆转录病毒载体。当本文公开的核酸中的任一个被引入细胞中时,经编码的嵌合dna靶向蛋白、嵌合衔接蛋白或向导rna可以在细胞中进行瞬时、条件性或组成性表达。
[0276]
任选地,可以对核酸进行密码子优化以在特定细胞或生物体中有效翻译成蛋白。例如,与天然存在的多核苷酸序列相比,可以修饰核酸以取代在真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、大鼠细胞或任何其它感兴趣的宿主细胞中具有更高使用频率的密码子。
[0277]
在一些组合物和方法中,cas蛋白、嵌合衔接蛋白和向导rna可以以rna的形式提供。此类rna可以是经修饰的rna。参见例如wo 2017/173054、us 2019/0136231和wo2018/107028,它们中的每一个以全文引用的方式并入本文用于所有目的。例如,rna中的一个或
多个可以被修饰成包含5'末端和/或3'末端处的一个或多个稳定末端修饰。此类修饰可以包括例如5'末端和/或3'末端处的一个或多个硫代磷酸酯键或5'末端和/或3'末端(例如,5'端或3'端)处的一个或多个2'-o-甲基修饰。作为一个实例,至少5'末端的前1、2、3或4个核苷酸可以被修饰,并且至少3'末端的最后1、2、3或4个核苷酸可以被修饰。例如,此类修饰可以包括在5'末端的前1、2、3或4个核苷酸处的2'-o-甲基修饰的核苷酸和/或在3'末端的最后1、2、3或4个核苷酸处的2'-o-甲基修饰的核苷酸。附加地或替代地,此类修饰可以包括例如在5'末端的前四个核苷酸中的一个或多个之间或在3'末端的最后四个核苷酸中的一个或多个之间的硫代、磷酸酯键。例如,5'末端的前四个核苷酸可以通过硫代磷酸酯键链接,和/或3'末端的最后四个核苷酸可以通过硫代磷酸酯键链接。在具体实例中,rna(例如,向导rna,如化学合成的向导rna)在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。此类化学修饰可以例如提供更大的稳定性和防止核酸外切酶向导rna,允许所述向导rna比未经修饰的向导rna在细胞内持续更长时间。例如,此类化学修饰还可以防止先天的细胞内免疫应答,所述先天的细胞内免疫应答可以主动降解rna或触发导致细胞死亡的免疫级联反应。
[0278]
经修饰的核苷或核苷酸可以存在于向导rna或mrna中。包含一个或多个经修饰核苷或核苷酸的向导rna或mrna称为经修饰rna,以描述一个或多个非天然和/或天然存在的组分或构型的存在,这些组分或构型用于替代或补充标准a、g、c和u残基。经修饰的核苷和核苷酸可以包括以下各项中的一个或多个:(1)磷酸二酯主链键中的非连接磷酸氧中的一个或两个和/或连接磷酸氧中的一个或多个的改变或替换(主链修饰);(2)核糖的组成的改变或替换(例如,核糖上的2'羟基的改变或替换)(糖修饰);(3)用脱磷接头批量替换磷酸酯部分(主链修饰);(4)天然存在的核碱基的修饰或替换(例如,用非标准核碱基)(碱基修饰);(5)核糖-磷酸酯主链的替换或修饰(主链修饰);(6)寡核苷酸的3'末端或5'末端的修饰(例如,末端磷酸基团的去除、修饰或替换,或部分、帽或接头的缀合(此类3'或5'帽修饰可以包含糖和/或主链修饰));以及(7)糖的修饰或替换(糖修饰)。
[0279]
可以组合修饰以提供包含核苷和核苷酸(残基)的经修饰rna,其可以具有两个、三个、四个或更多个修饰。例如,经修饰残基可以具有经修饰的糖和经修饰的核碱基。在一些实例中,grna或mrna的每个碱基均被修饰(例如,所有碱基均具有经修饰的磷酸基团,如硫代磷酸酯基团)。例如,grna或mrna分子的所有或基本上所有磷酸基团均可以被硫代磷酸酯基团替换。在其它实例中,经修饰的rna在rna的5'末端处或其附近和/或在rna的3'末端处或其附近包含至少一个经修饰残基。
[0280]
在一些实例中,经修饰的grna或mrna包含一个、两个、三个或更多个经修饰残基。在一些实例中,grna或mrna在grna或mrna的5'和3'末端中的每一个处包含一个、两个、三个或更多个经修饰残基。在一些实例中,经修饰的mrna包含5、10、15、50、100、200、300、400、500、600、700、800、900、1000或更多个经修饰残基。在一些实例中,经修饰的grna或mrna中至少5%(例如,至少约5%、至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%或约100%)的的位置是经修饰的核苷或核苷酸。
[0281]
未经修饰的核酸可能易于被例如细胞核酸酶降解。例如,核酸酶可以水解核酸磷
酸二酯键。为了提供稳定性,本文所述的rna(例如,向导rna或嵌合cas蛋白mrna或嵌合衔接蛋白mrna)可以含有一个或多个经修饰的核苷或核苷酸。在一些实例中,本文所述的经修饰的rna分子在体内和离体引入细胞群时可以表现出降低的先天免疫响应。术语先天性免疫响应包括对外源核酸(包括单链核酸)的细胞响应,其涉及诱导细胞因子表达和释放(特别是干扰素),以及细胞死亡。
[0282]
在主链修饰的一些实例中,经修饰残基的磷酸基团可以通过用不同的取代基替换氧中的一个或多个来修饰。此外,经修饰残基(例如,经修饰的核酸中存在的经修饰残基)可以包括用如本文所述的经修饰的磷酸基团批量替换未经修饰的磷酸酯部分。在一些实例中,磷酸酯主链的主链修饰可以包括导致不带电接头或具有不对称电荷分布的带电接头的改变。
[0283]
经修饰磷酸基团的实例包括硫代磷酸酯、硒代磷酸酯、硼酸磷酸酯、硼酸磷酸酯、膦酸氢酯、亚氨代磷酸酯、烷基或芳基膦酸酯和磷酸三酯。未经修饰的磷酸基团中的磷原子是非手性的。然而,用上述原子或原子团中的一个替换非桥接氧中的一个可以使磷原子手性。立体异构磷原子可以具有“r”构型或“s”构型。主链也可以通过用氮(桥接的亚氨代磷酸酯)、硫(桥接的硫代磷酸酯)和碳(桥接的亚甲基膦酸酯)替换桥接氧(即,将磷酸酯与核苷链接的氧)来修饰。替换可以发生在链接氧处或两个链接氧处。
[0284]
在某些主链修饰中,磷酸基团可以被不含磷的连接头替换。例如,带电荷的磷酸基团可以被中性部分替换。可以替换磷酸基团的部分的实例包括例如膦酸甲酯、羟氨基、硅氧烷、碳酸酯、羧甲基、氨基甲酸酯、酰胺、硫醚、环氧乙烷接头、磺酸酯、磺酰胺、硫代甲缩醛、甲缩醛、肟、亚甲基亚氨基、亚甲基甲基亚氨基、亚甲基肼基、亚甲基二甲基肼基和亚甲基氧基甲基亚氨基。
[0285]
也可以构建可以模拟核酸的支架,其中磷酸酯接头和核糖被核酸酶抗性核苷或核苷酸替代物替换。此类修饰可以包含主链和糖修饰。例如,核碱基可以由替代主链拴系。实例包括吗啉代、环丁基、吡咯烷和肽核酸(pna)核苷替代物。
[0286]
经修饰的核苷和经修饰的核苷酸还可以包括对糖基团的一个或多个修饰。例如,2'羟基(oh)可以被多个不同的氧或脱氧取代基修饰或替换。对2'羟基的此类修饰可以增强核酸的稳定性,因为羟基不能再被去质子化以形成2'-醇盐离子。
[0287]
2'羟基修饰的实例包括烷氧基或芳氧基(or,其中r可以是例如烷基、环烷基、芳基、芳烷基、杂芳基或糖)或聚乙二醇(peg)、o(ch2ch2o)nch2ch2or,其中r可以是例如h或任选取代的烷基,并且n可以是0至20的整数(例如0至4、0至8、0至10、0至16、1至4、1至8、1至10、1至16、1至20、2至4、2至8、2至10、2至16、2至20、4至8、4至10、4至16和4至20)。在一个实例中,2'羟基修饰可以是2'-o-me。在另一个实例中,2'羟基修饰可以是2'-氟修饰,其用氟化物替换2'羟基。在其它实例中,2'羟基修饰可以包括锁定核酸(lna),其中2'羟基可以例如通过c
1-6
亚烷基或c
1-6
杂亚烷基桥接到相同核糖的4'碳。示范性桥可以包括亚甲基、丙烯、醚或氨基桥;o-氨基(其中氨基可以是例如nh2;烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基或二杂芳基氨基、乙二胺或聚氨基)和氨基烷氧基、o(ch2)
n-氨基(其中氨基可以是例如nh2、烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基或二杂芳基氨基、乙二胺或聚氨基)。在一些实例中,2'羟基修饰可以包括未锁定的核酸(una),其中核糖环缺少c2'-c3'键。在一些实例中,2'羟基修饰可以包括甲氧基乙基(moe),
(och2ch2och3,例如,peg衍生物)。
[0288]
脱氧2'修饰可以包括氢(即,脱氧核糖,例如,在部分dsrna的突出部分);卤素(例如,溴、氯、氟或碘);氨基(其中氨基可以是例如-nh2、烷基氨基、二烷基氨基、杂环基、芳基氨基、二芳基氨基、杂芳基氨基、二杂芳基氨基或氨基酸);nh(ch2ch2nh)nch2ch
2-氨基(其中氨基可以是,例如,如本文所述)、-nhc(o)r(其中r可以是,例如,烷基、环烷基、芳基、芳烷基,杂芳基或糖)、氰基;巯基;烷基硫代烷基;硫代烷氧基;以及烷基、环烷基、芳基、烯基和炔基,它们可以任选地被例如本文所述的氨基取代。
[0289]
糖修饰可以包含糖基团,该糖基团还可以含有具有与核糖中对应碳的立体化学构型相反的立体化学构型的一个或多个碳。因此,经修饰的核酸可以包括含有例如阿拉伯糖作为糖的核苷酸。经修饰的核酸还可以包括脱碱基糖。这些脱碱基糖也可以在组成糖原子中的一个或多个处被进一步修饰。经修饰的核酸还可以包括呈l形式的一个或多个糖(例如,l-核苷)。
[0290]
本文所述的经修饰核苷和经修饰核苷酸(其可掺入经修饰核酸中)可以包括经修饰的碱基,也称为核碱基。核碱基的实例包括但不限于腺嘌呤(a)、鸟嘌呤(g)、胞嘧啶(c)和尿嘧啶(u)。这些核碱基可以被修饰或完全替换以提供可以掺入经修饰核酸中的经修饰残基。核苷酸的核碱基可以独立地选自嘌呤、嘧啶、嘌呤类似物或嘧啶类似物。在一些实例中,核碱基可以包括例如碱基的天然存在和合成衍生物。
[0291]
grna或mrna的一端或两端处的一个或多个残基可以被化学修饰,或者整个grna或mrna可以被化学修饰。一些实例包含5'末端修饰。一些示例实施例包含3'末端修饰。在某些grna中,grna分子单链突出端中的一个或多个或所有核苷酸均为脱氧核苷酸。在某些经修饰的mrna中,mrna可以含有5'末端和/或3'末端修饰。
[0292]
可以修饰作为mrna提供的嵌合cas蛋白、嵌合衔接蛋白或这两者以提高稳定性和/或免疫原性特性。可以对mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统中的模式识别受体的激活)。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合cas mrna、嵌合衔接蛋白mrna或sam mrna。同样地,嵌合cas mrna、嵌合衔接蛋白mrna或sam mrna可以通过使用同义密码子消耗尿苷来修饰。其它可能的修饰在本文别处更详细地描述。
[0293]
可以修饰作为mrna提供的嵌合cas蛋白和/或嵌合衔接蛋白以提高稳定性和/或免疫原性特性。可以对mrna内的一种或多种核苷进行修饰。对mrna核碱基的化学修饰的实例包括假尿苷、1-甲基-假尿苷和5-甲基-胞苷。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被加帽。帽可以是,例如,其中 1核糖核苷酸在核糖的2'o位置被甲基化的帽1结构。例如,加帽可以在体内产生优异的活性(例如,通过模拟天然帽),可以产生减少对宿主先天免疫系统的刺激的天然结构(例如,可以减少先天免疫系统
中的模式识别受体的激活)。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被聚腺苷酸化(以包含聚(a)尾)。编码嵌合cas蛋白的mrna、编码嵌合衔接蛋白的mrna或编码两者的sam mrna也可以被修饰以包括假尿苷(例如,可以被假尿苷完全取代)。例如,可以使用含有n1-甲基假尿苷的加帽和聚腺苷酸化的嵌合cas mrna、嵌合衔接蛋白或sam mrna。同样地,嵌合cas mrna、嵌合衔接蛋白mrna或sam mrna可以通过使用同义密码子消耗尿苷来修饰。
[0294]
嵌合cas mrna、嵌合衔接mrna或sam mrna可以在至少一个、多个或所有尿苷位置包含经修饰的尿苷。经修饰的尿苷可以是在5位置被修饰(例如,用卤素、甲基或乙基)的尿苷。经修饰的尿苷可以是在1位置被修饰(例如,用卤素、甲基或乙基)的假尿苷。经修饰的尿苷可以是例如假尿苷、n1-甲基-假尿苷、5-甲氧基尿苷、5-碘尿苷或其组合。在一些实例中,经修饰的尿苷是5-甲氧基尿苷。在一些实例中,经修饰的尿苷是5-碘尿苷。在一些实例中,经修饰的尿苷是假尿苷。在一些实例中,经修饰的尿苷是n1-甲基-假尿苷。在一些实例中,经修饰的尿苷是假尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是n1-甲基假尿苷和5-甲氧基尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和n1-甲基-假尿苷的组合。在一些实例中,经修饰的尿苷是假尿苷和5-碘尿苷的组合。在一些实例中,经修饰的尿苷是5-碘尿苷和5-甲氧基尿苷的组合。
[0295]
本文公开的嵌合cas mrna、嵌合衔接蛋白mrna或sam mrna还可以包含5'帽,如cap0、cap1或cap2。5'帽通常是通过5'-三磷酸酯链接到mrna的5'至3'链的第一核苷酸(即,第一近帽核苷酸)的5'位置的7-甲基鸟嘌呤核糖核苷酸(其可以被进一步修饰,例如,关于arca)。在cap0中,mrna的第一和第二近帽核苷酸的核糖均包含2'-羟基。在cap1中,mrna的第一和第二转录核苷酸的核糖分别包含2'-甲氧基和2'-羟基。在cap2中,mrna的第一和第二近帽核苷酸的核糖均包含2'-甲氧基。参见例如katibah等人(2014)《美国科学院院报(proc.natl.acad.sci.u.s.a.)》111(33):12025-30和abbas等人(2017)《美国科学院院报》114(11):e2106-e2115,它们中的每一个以全文引用的方式并入本文用于所有目的。大多数内源性高等真核mrna,包括哺乳动物mrna,如人mrna,均包含cap1或cap2。cap0和其它不同于cap1和cap2的帽结构可能在哺乳动物(如人类)中具有免疫原性,因为先天免疫系统的组分(如ifit-1和ifit-5)将其识别为非自身,这可能导致细胞因子升高水平(包括i型干扰素)。先天免疫系统的组分如ifit-1和ifit-5也可能与eif4e竞争mrna与cap1或cap2以外的帽的结合,从而潜在地抑制mrna的翻译。
[0296]
可以共转录地包括帽。例如,arca(抗反向帽类似物;赛默飞世尔科技公司(thermo fisher scientific)商品目录号第am8045号)是包含与鸟嘌呤核糖核苷酸的5'位置链接的7-甲基鸟嘌呤3'-甲氧基-5'-三磷酸酯的帽类似物,其可在体外在起始时掺入转录物中。arca产生cap0帽,其中第一近帽核苷酸的2'位置是羟基。参见例如stepinski等人,(2001)《rna》7:1486-1495,该文献以全文引用的方式并入本文用于所有目的。
[0297]
cleancap
tm ag(m7g(5')ppp(5')(2'omea)pg;trilink生物技术公司商品目录号第n-7113号)或cleancap
tm gg(m7g(5')ppp(5')(2'omeg)pg;trilink生物技术公司商品目录号第n-7133号)可用于共转录地提供cap1结构。3'-o-甲基化版本的cleancap
tm ag和cleancap
tm gg也可从trilink生物技术公司获得,商品目录号分别为第n-7413和第n-7433
号。
[0298]
替代地,可以在转录后将帽添加到rna。例如,牛痘加帽酶是可商购的(新英格兰生物实验室(new england biolabs)商品目录号第m2080s号)并具有由其d1亚基提供的rna三磷酸酶和鸟苷酸转移酶活性,以及由其d12亚基提供的鸟嘌呤甲基转移酶活性。因此,在s-腺苷甲硫氨酸和gtp的存在下,它可以将7-甲基鸟嘌呤添加到rna中,从而产生cap0。参见例如guo和moss(1990)《美国科学院院报》87:4023-4027,和mao和shuman(1994)《生物化学杂志》269:24472-24479,它们中的每一个以全文引用的方式并入本文用于所有目的。
[0299]
嵌合cas mrna、嵌合衔接蛋白mrna或sam mrna还可以包含聚腺苷酸化(聚a)尾。例如,聚a尾可以包含至少20个、至少30个、至少40个、至少50个、至少60个、至少70个、至少80个、至少90个或至少100个腺嘌呤,以及任选地多达300个腺嘌呤。例如,聚a尾可以包含95、96、97、98、99或100个腺嘌呤核苷酸。
[0300]
替代地,cas蛋白、嵌合衔接蛋白和向导rna可以以dna的形式提供。dna或表达盒可以用于稳定整合到细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)的基因组(即,染色体)中,或者它可以用于在染色体外部表达(例如,染色体外复制dna)。稳定整合的表达盒或核酸可以随机整合到真核生物体(例如动物、非人动物、哺乳动物或非人哺乳动物)的基因组中(即,转基因),或者它们可以整合到真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)的基因组的预定区域中(即,敲入)。
[0301]
本文所述的核酸或表达盒可以可操作地链接到任何合适的启动子以在真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)体内表达或离体在细胞内表达。真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)可以是如本文别处所述的任何合适的真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)。作为一个实例,核酸或表达盒(例如,嵌合cas蛋白表达盒、嵌合衔接蛋白表达盒或包含编码嵌合cas蛋白和嵌合衔接蛋白两者的核酸的sam盒)可以用于可操作地链接到基因组基因座处的内源启动子。替代地,盒核酸或表达盒可以可操作地链接到外源启动子,如组成型活性启动子(例如,cag启动子或u6启动子)、条件型启动子、诱导型启动子、时间受限启动子(例如,发育调控启动子)或空间受限启动子(例如,细胞特异性或组织特异性启动子)。此类启动子是众所周知的并且在本文别处讨论。可以在表达构建体中使用的启动子包括例如在真核细胞、非人真核细胞、动物细胞、非人动物细胞、哺乳动物细胞、非人哺乳动物细胞、人细胞、非人细胞、啮齿动物细胞、小鼠细胞、大鼠细胞、仓鼠细胞、兔细胞、多能细胞、胚胎干(es)细胞或合子中的一个或多个中具有活性的启动子。此类启动子可以是例如条件启动子、诱导型启动子、组成型启动子或组织特异性启动子。
[0302]
例如,编码向导rna的核酸可以可操作地链接到u6启动子,如人u6启动子或小鼠u6启动子。合适的启动子(例如,用于表达向导rna)的具体实例包括rna聚合酶iii启动子,如人u6启动子、大鼠u6聚合酶iii启动子或小鼠u6聚合酶iii启动子。
[0303]
任选地,启动子可以是驱动一个基因(例如,编码嵌合cas蛋白的基因)和另一个方向上的第二基因(例如,编码向导rna或嵌合衔接蛋白的基因)的表达的双向启动子。此类双向启动子可以由以下各项组成:(1)含有3个外部控制元件:远侧序列元件(dse)、近侧序列元件(pse)和tata框的完整的、常规的、单向的pol iii启动子;(2)包括在相反定向上与dse的5'末端融合的pse和tata盒的第二基本pol iii启动子。例如,在h1启动子中,dse邻近pse
和tata框,并且可以通过产生杂合启动子使启动子双向化,其中通过源自u6启动子的附加pse和tata盒来控制反向转录。参见例如us 2016/0074535,所述文献出于所有目的通过引用整体并入本文。使用双向启动子同时表达两个基因允许生成紧凑的表达盒以促进递送。
[0304]
核酸中的一个或多个可以一起在多顺反子表达构建体或多顺反子信使rna中。例如,编码嵌合cas蛋白的核酸和编码嵌合衔接蛋白的核酸可以一起在双顺反子表达构建体中。多顺反子表达载体同时表达来自相同mrna(即,由相同启动子产生的转录物)的两种或更多种单独的蛋白质。用于蛋白质多顺反子表达的合适策略包含例如使用2a肽和使用内部核糖体进入位点(ires)。例如,此类构建体可以包含:(1)编码一个或多个嵌合cas蛋白和一个或多个嵌合衔接蛋白的核酸;(2)编码两个或更多个嵌合衔接蛋白的核酸;(3)编码两个或更多个嵌合cas蛋白的核酸;(4)编码两个或更多个向导rna的核酸;(5)编码一个或多个嵌合cas蛋白和一个或多个向导rna的核酸;(6)编码一个或多个嵌合衔接蛋白和一个或多个向导rna的核酸;或(7)编码一个或多个嵌合cas蛋白、一个或多个嵌合衔接蛋白和一个或多个向导rna的核酸。作为一个实例,此类多顺反子载体可以使用一个或多个内部核糖体进入位点(ires)以允许从mrna的内部区域引发翻译。作为另一个实例,此类多顺反子载体可以使用一种或多种2a肽。这些肽是长度通常为18-22个氨基酸的小“自切割”肽,并且从相同mrna产生等摩尔水平的多个基因。核糖体在2a肽的c端跳过甘氨酰-脯氨酰肽键的合成,从而导致2a肽与其直接下游肽之间的“切割”。参见例如kim等人(2011)《公共科学图书馆
·
综合》6(4):e18556,所述文献出于所有目的通过引用整体并入本文。“切割”发生在c端上存在的甘氨酸与脯氨酸残基之间,这意味着上游顺反子将在末端添加一些另外的残基,而下游顺反子将从脯氨酸开始。因此,“切断”的下游肽在其n端具有脯氨酸。2a介导的切割是所有真核细胞中的普遍现象。已从小核糖核酸病毒、昆虫病毒和c型轮状病毒中鉴定出2a肽。参见例如szymczak等人(2005)《生物疗法专家意见(expert opin.biol.ther.》5(5):627-638,该文献以全文引用的方式并入本文用于所有目的。可以使用的2a肽的实例包括:thoseaasigna病毒2a(t2a);猪捷申病毒-12a(p2a);马鼻炎a病毒(erav)2a(e2a);以及fmdv 2a(f2a)。示范性t2a、p2a、e2a和f2a序列包括以下各项:t2a(egrgslltcgdveenpgp;seq id no:20);p2a(atnfsllkqagdveenpgp;seq id no:21);e2a(qctnyallklagdvesnpgp;seq id no:22);以及f2a(vkqtlnfdllklagdvesnpgp;seq id no:23)。gsg残基可以添加到这些肽中的任何肽的5'端以提高切割效率。
[0305]
核酸或表达盒中的任一个还可以包含编码序列上游的聚腺苷酸化信号或转录终止子。例如,嵌合cas蛋白表达盒、嵌合衔接蛋白表达盒、sam表达盒或向导rna表达盒可以在表达盒中的编码序列上游包含聚腺苷酸化信号或转录终止子。聚腺苷酸化信号或转录终止子可以侧接有由位点特异性重组酶识别的重组酶识别位点。聚腺苷酸化信号或转录终止子阻止由编码序列(例如,嵌合cas蛋白、嵌合衔接蛋白、向导rna或重组酶)编码的蛋白质或rna的转录和表达。然而,在暴露于位点特异性重组酶后,聚腺苷酸化信号或转录终止子将被切除,并且可以表达蛋白质或rna。
[0306]
如果以组织特异性或发育期特异性方式切除聚腺苷酸化信号或转录终止子,则表达盒(例如,嵌合cas蛋白表达盒或sam表达盒)的此类构型可以在包含表达盒的真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中实现组织特异性表达或发育期特异性表达。例如,在嵌合cas蛋白的情况下,这可以降低由于嵌合cas蛋白在细胞或真核生物体
(例如,动物、非人动物、哺乳动物或非人哺乳动物)中延长表达而导致的毒性,或减少嵌合cas蛋白在真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中在不希望的发育期或在不希望的细胞或组织类型中的表达。参见例如parikh等人(2015)《公共科学图书馆
·
综合》10(1):e0116484,该文献以全文引用的方式并入本文用于所有目的。如果包含表达盒的真核生物(例如,动物、非人动物、哺乳动物或非人哺乳动物)还包含可操作地链接到组织特异性或发育期特异性启动子的位点特异性重组酶的编码序列,则可以实现以组织特异性或发育期特异性方式切除聚腺苷酸化信号或转录终止子。然后,聚腺苷酸化信号或转录终止子将仅在那些组织中或在那些发育阶段被切除,从而实现组织特异性表达或发育阶段特异性表达。在一个实例中,嵌合cas蛋白、嵌合衔接蛋白、嵌合cas蛋白和嵌合衔接蛋白,或向导rna可以以肝脏特异性方式表达。
[0307]
可以使用任何转录终止子或聚腺苷酸化信号。如本文所使用的,“转录终止子”是指引起转录终止的dna序列。在真核生物中,转录终止子被蛋白质因子识别,并且聚腺苷酸化在终止之后,所述聚腺苷酸化是在poly(a)聚合酶存在的情况下向mrna转录物添加poly(a)尾部的过程。哺乳动物poly(a)信号通常由约45个核苷酸长的核心序列组成,所述核心序列可以侧接有用于增强切割和聚腺苷酸化效率的不同辅助序列。核心序列由以下组成:mrna中高度保守的上游元件(aataaa或aauaaa),所述上游元件被称为poly a识别基序或poly a识别序列,由切割和聚腺苷酸化特异性因子(cpsf)识别;以及受切割刺激因子(cstf)约束的定义不清的下游区域(在us或gs和us中富集)。可以使用的转录终止子的实例包含例如人生长激素(hgh)聚腺苷酸化信号、猿猴病毒40(sv40)晚期聚腺苷酸化信号、兔β-珠蛋白聚腺苷酸化信号、牛生长激素(bgh)聚腺苷酸化信号、磷酸甘油酸激酶(pgk)聚腺苷酸化信号、aox1转录终止序列、cyc1转录终止序列或已知适合于调节真核细胞中基因表达的任何转录终止序列。
[0308]
位点特异性重组酶包含可以促进重组酶识别位点之间重组的酶,其中两个重组位点在单个核酸内或在单独的核酸上物理分离。重组酶的实例包含cre、flp和dre重组酶。cre重组酶基因的一个实例是crei,其中两个对cre重组酶进行编码的外显子被内含子隔开,以防止其在原核细胞中表达。此类重组酶可以进一步包括用于促进定位到核(例如,nls-crei)的核定位信号。重组酶识别位点包含由位点特异性重组酶识别并且可以用作重组事件的底物的核苷酸序列。重组酶识别位点的实例包含frt、frt11、frt71、attp、att、rox和lox位点,如loxp、lox511、lox2272、lox66、lox71、loxm2和lox5171。
[0309]
本文公开的表达盒也可以包含其它组分。此类表达盒(例如,嵌合cas蛋白表达盒、嵌合衔接蛋白表达盒、sam表达盒、向导rna表达盒或重组酶表达盒)还可以包含在表达盒5'末端的3'剪接序列和/或编码序列之后的第二聚腺苷酸化信号(例如,编码嵌合cas蛋白、嵌合衔接蛋白或向导rna)。术语3'剪接序列是指3'内含子/外显子边界处可以被识别并通过剪接机制结合的核酸序列。表达盒还可以包含选择盒,其包含例如抗药性蛋白的编码序列。合适的选择标志物的实例包括新霉素磷酸转移酶(neor)、潮霉素b磷酸转移酶(hygr)、嘌呤霉素-n-乙酰转移酶(puror)、杀稻瘟素s脱氨酶(bsrr)、黄嘌呤/鸟嘌呤磷酸核糖转移酶(gpt)和单纯疱疹病毒胸苷激酶(hsv-k)。任选地,选择盒可以侧接有位点特异性重组酶的重组酶识别位点。如果表达盒还包含侧接如上所述编码序列上游的聚腺苷酸化信号的重组酶识别位点,则选择盒可以侧接相同的重组酶识别位点,或者可以侧接被不同的重组酶识
别的一组不同的重组酶识别位点。
[0310]
表达盒还可以包含编码一个或多个报告蛋白,如荧光蛋白(例如,绿色荧光蛋白)的核酸。可以使用任何合适的报告蛋白。例如,可以使用荧光报告蛋白,或者可以使用非荧光报告蛋白。本文别处提供了荧光报告蛋白的实例。非荧光报告蛋白包括例如可用于组织化学或生物发光测定的报告蛋白,如β-半乳糖苷酶、萤光素酶(例如,海肾萤光素酶、萤火虫萤光素酶和nanoluc萤光素酶)和β-葡萄糖醛酸酶。表达盒可以包括可以在流式细胞术测定中检测到的报告蛋白(例如,荧光报告蛋白,如绿色荧光蛋白)和/或可以在组织化学测定中检测到的报告蛋白(例如,β-半乳糖苷酶蛋白)。此类组织化学测定的一个实例是通过通过水解x-gal(5溴-4-氯-3-吲哚基-b-d-半乳糖皮蒽)(其产生蓝色沉淀物),或使用荧光底物,如β-甲基伞形基半乳糖苷(mug)和荧光素二半乳糖苷(fdg)对原位β-半乳糖苷酶表达进行组织化学可视化。
[0311]
本文所述的表达盒可以是任何形式。例如,表达盒可以在载体或质粒中。表达盒可以可操作地链接到能够引导蛋白或rna表达的表达构建体中的启动子(例如,在去除上游聚腺苷酸化信号后)。替代地,表达盒可以在靶向载体中。例如,靶向载体可以包含侧接表达盒的同源臂,其中同源臂适用于引导与所需靶基因组基因座的重组以促进内源序列的基因组整合和/或替换。
[0312]
编码无催化活性的cas蛋白的核酸的具体实例可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:2所示的dcas9蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸。任选地,核酸可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:24所示的序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸(任选地,其中该序列编码与seq id no:2所示的dcas9蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的蛋白)。
[0313]
编码嵌合cas蛋白的核酸的具体实例可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:1所示的嵌合cas蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸。任选地,核酸可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:25所示的序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸(任选地,其中该序列编码与seq id no:1所示的嵌合cas蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的蛋白)。
[0314]
编码衔接子的核酸的具体实例可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:7所示的mcp序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸。任选地,核酸可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:26所示的序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸(任选地,其中该序列编码与seq id no:7所示的mcp序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的蛋白)。
[0315]
编码嵌合衔接蛋白的核酸的具体实例可以包含以下核酸、基本上由以下核酸组
成,或由以下核酸组成:编码与seq id no:6所示的嵌合衔接蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸。任选地,核酸可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:27所示的序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸(任选地,其中该序列编码与seq id no:6所示的嵌合衔接蛋白序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的蛋白)。
[0316]
编码转录激活域的核酸的具体实例可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:3、8或9中分别所示的vp64、p65或hsf1序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸。任选地,核酸可以包含以下核酸、基本上由以下核酸组成,或由以下核酸组成:编码与seq id no:28、29或30中分别所示的序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的氨基酸序列的核酸(任选地,其中该序列编码与seq id no:3、8或9中分别所示的vp64、p65或hsf1序列至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同的蛋白)。
[0317]
协同激活介体(sam)表达盒的一个实例从5'到3'包含:(a)3'剪接序列;(b)第一重组酶识别位点(例如,loxp位点);(c)抗药性基因的编码序列(例如,新霉素磷酸转移酶(neor)编码序列);(d)聚腺苷酸化信号;(e)第二重组酶识别位点(例如,loxp位点);(f)嵌合cas蛋白编码序列(例如,dcas9-nls-vp64融合蛋白);(g)2a蛋白编码序列(例如,t2a编码序列);以及(e)嵌合衔接蛋白编码序列(例如,mcp-nls-p65-hsf1)。参见例如seq id no:31(seq id no:46所示的编码序列和seq id no:44所示的编码蛋白,以及seq id no:61所示的mrna序列)。
[0318]
通用向导rna阵列表达盒的一个实例从5'到3'包含:(a)3'剪接序列;(b)第一重组酶识别位点(例如,rox位点);(c)抗药性基因的编码序列(例如,嘌呤霉素-n-乙酰转移酶(puror)编码序列);(d)聚腺苷酸化信号;(e)第二重组酶识别位点(例如,rox位点);(f)包含一个或多个向导rna基因的向导rna(例如,第一u6启动子后接第一向导rna编码序列,第二u6启动子后接第二向导rna编码序列,以及第三u6启动子后接第三向导rna编码序列)。参见例如seq id no:32。包含启动子和向导rna编码序列的seq id no:32区域在seq id no:47中示出。此类编码靶向小鼠ttr的向导rna的向导rna阵列表达盒在seq id no:33中示出。包含启动子和向导rna编码序列的seq id no:33的区域在seq id no:48中示出。
[0319]
通用向导rna阵列表达盒的另一个实例包含一个或多个向导rna基因(例如,第一u6启动子后接第一向导rna编码序列,第二u6启动子后接第二向导rna编码序列,以及第三u6启动子后接第三向导rna编码序列)。此类通用向导rna阵列表达盒在seq id no:48中示出。用于特定基因的此类向导rna阵列表达盒的实例在例如seq id no:33、48和49中示出。
[0320]
iv.脂质纳米颗粒和将向导rna和其它组分引入细胞和真核生物体
[0321]
本文还公开了用于将同一lnp中的所有sam系统组分递送至细胞或真核生物体以便增加靶基因的转录或表达的脂质纳米颗粒(lnp)。本文公开的方法包含在同一lnp中一起将协同激活介体(sam)系统的所有组分(一个或多个向导rna或核酸编码、嵌合cas蛋白或核酸编码,以及嵌合衔接蛋白或核酸编码)引入细胞或真核生物体(例如,动物、非人动物、哺
乳动物或非人哺乳动物)。例如,此类lnp可以包含负荷物,该负荷物包含:(a)编码嵌合规律间隔成簇短回文重复序列(crispr)相关联(cas)蛋白的核酸,该蛋白包含与一个或多个转录激活域融合的无核酸酶活性的cas蛋白;(b)编码嵌合衔接蛋白的核酸,该嵌合衔接蛋白包含与一个或多个转录激活域融合的衔接子;以及(c)一个或多个向导rna或编码该一个或多个向导rna的一个或多个核酸,每个向导rna包含嵌合衔接蛋白可特异性结合的一个或多个衔接结合元件,并且其中一个或多个向导rna中的每一个能够与cas蛋白形成复合物并将其导向至靶基因内的靶序列,从而增加靶基因的表达。在一个实例中,协同激活介体系统的所有组分以rna的形式在同一lnp中一起引入。“引入”包括以核酸或蛋白进入细胞内部或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)内的细胞内部的方式向细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)呈递核酸或蛋白。
[0322]
可以将向导rna以rna的形式(例如,体外转录的rna)或对向导rna进行编码的dna的形式引入到细胞中。同样地,可以将如嵌合cas蛋白和嵌合衔接蛋白等蛋白组分以dna、rna或蛋白的形式引入到细胞中。当以dna的形式引入时,对向导rna进行编码的dna可以可操作地连接到在细胞中具有活性的启动子。此类dna可以在一种或多种表达构建体中。例如,此类表达构建体可以是单个核酸分子的组分。可替代地,其可以在两个或多个核酸分子之间以任何组合分离(即,对一种或多种crispr rna进行编码的dna和对一种或多种tracrrna进行编码的dna可以是单独的核酸分子的组分)。编码嵌合cas蛋白、嵌合衔接蛋白或向导rna的核酸在本文别处更详细地讨论。
[0323]
在具体实例中,一个或多个向导rna、嵌合cas蛋白和嵌合衔接蛋白各自以rna的形式经由lnp介导的递送在同一lnp中引入。如本文别处更详细讨论的,rna中的一种或多种可以被修饰成包括5'端和/或3'端处的一种或多种稳定端修饰。此类修饰可以包含例如5'端和/或3'端处的一个或多个硫代磷酸酯键或5'端和/或3'端处的一个或多个2'-o-甲基修饰。通过此类方法的递送导致瞬时cas表达和/或向导rna的存在,并且生物可降解脂质提高清除率、提高耐受性并降低免疫原性。脂质调配物可以保护生物分子免于降解,同时改善其细胞摄取。
[0324]
脂质纳米颗粒是包括通过分子间力彼此物理相关的多个脂质分子的颗粒。这些颗粒包含微球体(包含单层和多层囊泡,例如,脂质体)、乳液中的分散相、胶束或悬浮液中的内相。此类脂质纳米颗粒可以用于封装一个或多个核酸或蛋白质以供递送。含有阳离子脂质的调配物可用于递送如核酸等聚阴离子。其它可以包含在内的脂质是中性脂质(即,不带电荷或两性离子脂质)、阴离子脂质、增强转染的辅助脂质和增加纳米颗粒可以在体内存在的时间长度的隐形脂质。合适的阳离子脂质、中性脂质、阴离子脂质、辅助脂质和隐形脂质的实例可以在wo 2016/010840 a1和wo 2017/173054 a1中找到,所述文献出于所有目的通过引用整体并入本文。示例性脂质纳米颗粒可以包括阳离子脂质和一种或多种其它组分。在一个实例中,其它组分可以包括如胆固醇等辅助脂质。在另一个实例中,其它组分可以包括如胆固醇等辅助脂质和如dspc等中性脂质。在另一个实例中,其它组分可以包括如胆固醇等辅助脂质、如dspc等任选的中性脂质以及如s010、s024、s027、s031或s033等隐形脂质。
[0325]
lnp可以含有以下中的一种或多种或全部:(i)用于封装和用于内体逃逸的脂质;(ii)用于稳定的中性脂质;(iii)用于稳定的辅助脂质;(iv)隐形脂质。参见例如,finn等人(2018),《细胞报告》22(9):2227-2235和wo 2017/173054 a1,所述文献中的每个文献出于
所有目的通过引用整体并入本文。在某些lnp中,负荷物可以包括向导rna或编码向导rna的核酸。在某些lnp中,负荷物可以包括sam mrna和向导rna或编码向导rna的核酸。
[0326]
用于包封和内体逃逸的脂质可以是阳离子脂质。脂质还可以是生物可降解脂质,如生物可降解可电离脂质。合适的脂质的一个实例是脂质a或lp01,即(9z,12z)-3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基十八-9,12-二烯酸酯,也被称为3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基(9z,12z)-十八-9,12-二烯酸酯。参见例如,finn等人(2018),《细胞报告》22(9):2227-2235和wo 2017/173054a1,所述文献中的每个文献出于所有目的通过引用整体并入本文。合适的脂质的另一个实例是脂质b,即((5-((二甲氨基)甲基)-1,3-亚苯基)双(氧))双(辛烷-8,1-二基)双(癸酸酯),也被称为((5-((二甲氨基)甲基)-1,3-亚苯基)双(氧基))双(辛烷-8,1-二基)双(癸酸酯)。合适的脂质的另一个实例是脂质c,即2-((4-(((3-(二甲氨基)丙氧基)羰基)氧基)十六酰基)氧基)丙烷-1,3-二基(9z,9'z,12z,12'z)-双(十八-9,12-二烯酸酯)。合适的脂质的另一个实例是脂质d,即3-(((3-(二甲氨基)丙氧基)羰基)氧基)-13-(辛酰氧基)十三烷基3-辛基十一烷酸酯。其它合适的脂质包括三十七-6,9,28,31-四烯-19-基4-(二甲氨基)丁酸酯(也被称为[(6z,9z,28z,31z)-三十七-6,9,28,31-四烯-19-基]4-(二甲氨基)丁酸酯或dlin-mc3-dma(mc3))。
[0327]
适用于本文所述的lnp的一些此类脂质在体内是生物可降解的。例如,包括此类脂质的lnp包含在8小时、10小时、12小时、24小时或48小时或3天、4天、5天、6天、7天或10天内从血浆中清除脂质的至少75%的那些。作为另一个实例,lnp的至少50%在8小时、10小时、12小时、24小时或48小时或3天、4天、5天、6天、7天或10天内从血浆中清除。
[0328]
根据其所在的介质的ph值,此类脂质可以是可电离的。例如,在微酸性介质中,脂质可以被质子化并且因此带有正电荷。相反,在弱碱性介质中,例如在ph大约为7.35的血液中,脂质可能不会被质子化并且因此不带电荷。在一些实施例中,脂质可以在至少约9、9.5或10的ph下质子化。这种脂质带电荷的能力与其固有pka有关。例如,脂质的pka可以独立地处于约5.8到约6.2的范围内。
[0329]
中性脂质的作用是稳定和改善lnp的处理。合适的中性脂质的实例包含各种中性、不带电荷或两性离子脂质。适用于本公开的中性磷脂的实例包括但不限于5-十七烷基苯-1,3-二醇(间苯二酚)、二棕榈酰磷脂酰胆碱(dppc)、二硬脂酰磷脂酰胆碱或1,2-二硬脂酰-sn-甘油-3-磷酸胆碱(dspc)、磷酸胆碱(dopc)、二肉豆蔻酰磷脂酰胆碱(dmpc)、磷脂酰胆碱(plpc)、1,2-二花生四烯酰-sn-甘油-3-磷酸胆碱(dapc)、磷脂酰乙醇胺(pe)、卵磷脂酰胆碱(epc)、二月桂酰磷脂酰胆碱(dlpc)、二肉豆蔻酰磷脂酰胆碱(dmpc)、1-肉豆蔻酰-2-棕榈酰磷脂酰胆碱(mppc)、1-棕榈酰-2-肉豆蔻酰磷脂酰胆碱(pmpc)、1-棕榈酰-2-硬脂酰磷脂酰胆碱(pspc)、1,2-二花生酰-sn-甘油-3-磷酸胆碱(dbpc)、1-硬脂酰-2-棕榈酰磷脂酰胆碱(sppc)、1,2-二二十碳烯酰-sn-甘油-3-磷酸胆碱(depc)、棕榈酰油酰磷脂酰胆碱(popc)、溶血磷脂酰胆碱、二油酰磷脂酰乙醇胺(dope)、二亚油酰磷脂酰胆碱二硬脂酰磷脂酰乙醇胺(dspe)、二肉豆蔻酰磷脂酰乙醇胺(dmpe)、二棕榈酰磷脂酰乙醇胺(dppe)、棕榈酰油酰磷脂酰乙醇胺(pope)、溶血磷脂酰乙醇胺、1-硬脂酰-2-油酰-sn-甘油-3-磷酸胆碱(sopc)和其组合。例如,中性磷脂可以选自由以下组成的组:二硬脂酰磷脂酰胆碱(dspc)和二肉豆蔻酰磷脂酰乙醇胺(dmpe)。
[0330]
辅助脂质包含增强转染的脂质。辅助脂质增强转染的机制可以包含增强颗粒稳定性。在某些情况下,辅助脂质可以增强膜融合性。辅助脂质包含类固醇、甾醇和烷基间苯二酚。合适的辅助脂质的实例包含胆固醇、5-十七烷基间苯二酚和胆固醇半琥珀酸酯。在一个实例中,辅助脂质可以是胆固醇或胆固醇半琥珀酸酯。
[0331]
隐形脂质包含改变纳米颗粒可以在体内存在的时间长度的脂质。隐形脂质可以通过例如减少颗粒聚集和控制粒度来帮助调配过程。隐形脂质可以调节lnp的药代动力学性质。合适的隐形脂质包含具有连接到脂质部分的亲水性头部基团的脂质。
[0332]
隐形脂质的亲水性头部基团可以包括例如选自基于peg(有时称为聚(环氧乙烷))、聚(噁唑啉)、聚(乙烯醇)、聚(甘油)、聚(n-乙烯基吡咯烷酮)、聚氨基酸和聚n-(2-羟丙基)甲基丙烯酰胺的聚合物的聚合物部分。术语peg意指任何聚乙二醇或其它聚亚烷基醚聚合物。在某些lnp调配物中,peg是peg-2k,也被称为peg 2000,其平均分子量为约2,000道尔顿。参见例如wo 2017/173054 a1,所述文献出于所有目的通过引用整体并入本文。
[0333]
隐形脂质的脂质部分可以衍生自例如二酰基甘油或二烷基甘酰胺,其包含包括二烷基甘油或二烷基甘酰胺基团的那些,所述二烷基甘油或二烷基甘酰胺基团具有独立地包括约c4到约c40个饱和或不饱和碳原子的烷基链长度,其中链可以包括一个或多个官能团,例如酰胺或酯。二酰基甘油或二烷基甘酰胺基团可以进一步包括一个或多个经取代的烷基。
[0334]
作为一个实例,隐形脂质可以选自peg-二月桂酰甘油、peg-二肉豆蔻酰甘油(peg-dmg)、peg-二棕榈酰甘油、peg-二硬脂酰甘油(peg-dspe)、peg-二月桂甘酰胺、peg-二肉豆蔻甘酰胺、peg-二棕榈酰甘酰胺和peg-二硬脂酰甘酰胺、peg-胆固醇(l-[8'-(胆甾-5-en-3[β]-氧基)甲酰胺基-3',6'-二氧杂辛基]氨甲酰基-[ω]-甲基-聚(乙二醇)、peg-dmb(3,4-二十四烷基苄基-[ω]-甲基-聚(乙二醇)醚)、1,2-二肉豆蔻酰-sn-甘油-3-磷酸乙醇胺-n-[甲氧基(聚乙二醇)-2000](peg2k-dmg)、1,2-二硬脂酰-sn-甘油-3-磷酸乙醇胺-n-[甲氧基(聚乙二醇)-2000](peg2k-dspe)、1,2-二硬脂酰-sn-甘油、甲氧基聚乙烯乙二醇(peg2k-dsg)、聚(乙二醇)-2000-二甲基丙烯酸酯(peg2k-dma)和1,2-二硬脂氧基丙基-3-胺-n-[甲氧基(聚乙二醇)-2000](peg2k-dsa)。在一个特定实例中,隐形脂质可以是peg2k-dmg。
[0335]
lnp可以包括调配物中相应摩尔比的组分脂质。ccd脂质的mol-%可以为例如约30mol-%到约60mol-%、约35mol-%到约55mol-%、约40mol-%到约50mol-%、约42mol-%到约47mol-%或约45%。辅助脂质的mol-%可以为例如约30mol-%到约60mol-%、约35mol-%到约55mol-%、约40mol-%到约50mol-%、约41mol-%到约46mol-%或约44mol-%。中性脂质的mol-%可以为例如约1mol-%到约20mol-%、约5mol-%到约15mol-%、约7mol-%到约12mol-%或约9mol-%。隐形脂质的mol-%可以为例如约1mol-%到约10mol-%、约1mol-%到约5mol-%、约1mol-%到约3mol-%、约2mol-%或约1mol-%。
[0336]
lnp在生物可降解脂质(n)的带正电荷的胺基团与待封装的核酸的带负电荷的磷酸基团(p)之间可以具有不同的比率。这可以由等式n/p在数学上表示。例如,n/p比率可以为约0.5至约100、约1至约50、约1至约25、约1至约10、约1至约7、约3至约5、约4至约5、约4、约4.5或约5。n/p比率也可以为约4到约7或约4.5到约6。在具体实例中,n/p比率可以为4.5或者可以为6。
[0337]
在一些lnp中,负荷物可以包含cas mrna或sam mrna(例如,编码例如通过2a编码
序列分开的嵌合cas蛋白和嵌合衔接蛋白两者的双顺反子mrna)和grna。cas mrna/sam mrna和grna的比率可以不同。例如,lnp调配物的cas mrna/sam mrna与grna核酸的比率的范围可以为约25:1至约1:25、约10:1至约1:10、约5:1至约1:5,或为约1:1。替代地,lnp调配物的cas mrna/sam mrna与grna核酸的比率可以为约1:1至约1:5,或为约10:1。替代地,lnp调配物的cas mrna/sam mrna与grna核酸的比率可以为约1:10、25:1、10:1、5:1、3:1、1:1、1:3、1:5、1:10或1:25。替代地,lnp调配物的cas mrna/sam mrna与grna核酸的比率可以为约1:1至约1:2。在具体实例中,cas mrna/sam mrna与grna的比率可以为约1:1或约1:2。
[0338]
lnp的示范性剂量包括约0.1、约0.25、约0.3、约0.5、约1、约2、约3、约4、约5、约6、约8或约10mg/kg体重(mpk)或约0.1至约10、约0.25至约10、约0.3至约10、约0.5至约10、约1至约10、约2至约10、约3至约10、约4至约10、约5至约10、约6至约10、约8至约10、约0.1至约8、约0.1至约6、约0.1至约5、约0.1至约4、约0.1至约3、约0.1至约2、约0.1至约1、约0.1至约0.5、约0.1至约0.3、约0.1至约0.25、约0.25至约8、约0.3至约6、约0.5至约5、约1至约5,或约2至约3mg/kg体重(相对于总rna(cas9 mrna和grna)负荷物含量)。此类lnp可以例如静脉内施用。在一个实例中,可以使用约0.01mg/kg至约10mg/kg、约0.1至约10mg/kg或约0.01至约0.3mg/kg的lnp剂量。例如,可以使用约0.01、约0.03、约0.1、约0.3、约0.5、约1、约2、约3或约10mg/kg的lnp剂量。在一个实例中,可以使用约0.5至约10、约0.5至约5、约0.5至约3、约1至约10、约1至约5、约1至约3,或约1至约2mg/kg的lnp剂量。
[0339]
合适的lnp的具体实例的氮磷(n/p)比率为4.5,并且含有摩尔比为45:44:9:2的可生物降解阳离子脂质、胆固醇、dspc和peg2k-dmg。生物可降解阳离子脂质可以是(9z,12z)-3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基十八-9,12-二烯酸酯,也被称为3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基(9z,12z)-十八-9,12-二烯酸酯。参见例如finn等人(2018)《细胞报告》22(9):2227-2235,所述文献出于所有目的通过引用整体并入本文。cas9 mrna/sam mrna与向导rna的重量比可以为1:1。合适的lnp的另一个具体实例含有摩尔比为50:38.5:10:1.5的dlin-mc3-dma(mc3)、胆固醇、dspc和peg-dmg。
[0340]
合适的lnp的另一个具体实例的氮磷(n/p)比率为6,并且含有摩尔比为50:38:9:3的可生物降解阳离子脂质、胆固醇、dspc和peg2k-dmg。生物可降解阳离子脂质可以是(9z,12z)-3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基十八-9,12-二烯酸酯,也被称为3-((4,4-双(辛氧基)丁酰基)氧基)-2-((((3-(二乙氨基)丙氧基)羰基)氧基)甲基)丙基(9z,12z)-十八-9,12-二烯酸酯。cas9 mrna/sam mrna与向导rna的重量比可以为1:2。
[0341]
合适的lnp的另一个具体实例的氮磷(n/p)比率为3,并且含有阳离子脂质、结构脂质、胆固醇(例如,胆固醇(ovine)(avanti 700000))和peg2k-dmg(例如,peg-dmg 2000(nof美国公司-gm-020(dmg-peg)),比率为50:10:38.5:1.5或47:10:42:1。结构脂质可以是例如dspc(例如dspc(avanti 850365))、sopc、dopc或dope。阳离子/可电离脂质可以是,例如,dlin-mc3-dma(例如,dlin-mc3-dma(biofine国际公司)。
[0342]
合适的lnp的另一个具体实例含有比率为45:9:44:2的dlin-mc3-dma、dspc、胆固醇和peg脂质。合适的lnp的另一个具体实例含有比率为50:10:39:1的dlin-mc3-dma、dope、胆固醇和peg脂质或peg dmg。合适的lnp的另一个具体实例具有比率为55:10:32.5:2.5的
dlin-mc3-dma、dspc、胆固醇和peg2k-dmg。合适的lnp的另一个具体实例具有比率为50:10:38.5:1.5的dlin-mc3-dma、dspc、胆固醇和peg-dmg。合适的lnp的另一个具体实例具有比率为50:10:38.5:1.5的dlin-mc3-dma、dspc、胆固醇和peg-dmg。
[0343]
体内施用可以通过任何合适的途径,包含例如肠胃外、静脉内、口服、皮下、动脉内、颅内、鞘内、腹膜内、局部、鼻内或肌肉内施用。全身施用方式包含例如口服和肠胃外途径。肠胃外途径的实例包含静脉内、动脉内、骨内、肌肉内、皮内、皮下、鼻内和腹膜内途径。具体的实例是静脉输液。鼻滴注和玻璃体内注射是其它具体实例。局部施用模式包括例如鞘内、脑室内、脑实质内(例如,局部脑实质内递送至纹状体(例如,进入尾状核或进入壳核种)、大脑皮层、中央前回、海马体(例如,进入齿状回或ca3区域)、颞叶皮层、杏仁核、额叶皮层、丘脑、小脑、髓质、下丘脑、顶盖、被盖或黑质)、眼内、眶内、结膜下、玻璃体内、视网膜下和经巩膜途径。与全身施用(例如,静脉内)相比,当局部施用(例如,脑实质内或玻璃体内)时,显著更少量的组分(与全身方法相比)可以发挥作用。局部施用方式还可以降低或消除当全身施用治疗有效量的组分时可能发生的潜在毒副作用的发生率。
[0344]
体内施用可以通过任何合适的途径,包含例如肠胃外、静脉内、口服、皮下、动脉内、颅内、鞘内、腹膜内、局部、鼻内或肌肉内施用。具体的实例是静脉输液。鼻滴注和玻璃体内注射是其它具体实例。可以使用一个或多个生理学和药学上可接受的载体、稀释剂、赋形剂或助剂来调配包含向导rna(或编码向导rna的核酸)的组合物。调配物可以取决于所选择的施用途径。术语“药学上可接受的”意指载体、稀释剂、赋形剂或助剂与调配物的其它成分相容并且对其接受者基本上无害。
[0345]
施用频率和给药次数可以取决于例如向导rna或嵌合cas蛋白或嵌合衔接蛋白mrna的半衰期和施用途径以及其它因素。将核酸或蛋白引入到细胞或真核生物体(例如,动物、非人动物、哺乳动物或非人哺乳动物)中可以在一段时间内执行一次或多次。例如,引入可以按以下频率执行:一段时间内仅一次、一段时间内至少两次、一段时间内至少三次、一段时间内至少四次、一段时间内至少五次、一段时间内至少六次、一段时间内至少七次、一段时间内至少八次、一段时间内至少九次、一段时间内至少十次、至少十一次、一段时间内至少十二次、一段时间内至少十三次、一段时间内至少十四次、一段时间内至少十五次、一段时间内至少十六次、一段时间内至少十七次、一段时间内至少十八次、一段时间内至少十九次或一段时间内至少二十次。
[0346]
lnp的示范性剂量包括约0.1、约0.25、约0.3、约0.5、约1、约2、约3、约4、约5、约6、约8或约10mg/kg体重(mpk)或约0.1至约10、约0.25至约10、约0.3至约10、约0.5至约10、约1至约10、约2至约10、约3至约10、约4至约10、约5至约10、约6至约10、约8至约10、约0.1至约8、约0.1至约6、约0.1至约5、约0.1至约4、约0.1至约3、约0.1至约2、约0.1至约1、约0.1至约0.5、约0.1至约0.3、约0.1至约0.25、约0.25至约8、约0.3至约6、约0.5至约5、约1至约5,或约2至约3mg/kg体重(相对于总rna(cas9 mrna和grna)负荷物含量)。此类lnp可以例如静脉内施用。
[0347]
出于所有目的,上文或下文引用的所有专利申请、网站、其它出版物、登录号等都通过引用整体并入,其程度如同每个单独的项目被单独并且具体地指出通过引用的方式并入。如果序列的不同版本与不同时间的登录号相关联,则意指在本技术的有效提交日期与登录号相关联的版本。有效提交日期是指实际提交日期或提及登记号的优先权申请的提交
日期(在适用情况下)中较早的日期。同样地,如果出版物、网站等的不同版本在不同时间发布,除非另有说明,否则指在申请的有效提交日期最近发布的版本。除非另外具体说明,否则本发明的任何特征、步骤、元件、实施例或方面都可以与任何其它特征、步骤、元件、实施例或方面结合使用。尽管为了清楚和理解起见,已通过图解和实例方式详细地对本发明进行了描述,但显而易见的是,可以在所附权利要求的范围内进行某些改变和修改。
[0348]
序列简要说明
[0349]
使用核苷酸碱基的标准字母缩写和氨基酸的三字母代码示出随附序列表中列出的核苷酸和氨基酸序列。核苷酸序列遵循从序列的5'末端开始并且向前(即,在每行中从左到右)到达3'末端的标准惯例。每个核苷酸序列仅示出一条链,但任何提及的显示链均应理解为包含互补链。当提供对氨基酸序列进行编码的核苷酸序列时,应当理解的是还提供了其对相同氨基酸序列进行编码的密码子简并变体。当提供编码氨基酸序列的dna序列时,应当理解,还提供了编码相同氨基酸序列的rna序列(通过用尿嘧啶替换胸腺嘧啶)。氨基酸序列遵循从序列的氨基端开始并且向前(即,在每行中从左到右)到达羧基端的标准惯例。
[0350]
表4:序列描述。
[0351]
[0352][0353]
实例
[0354]
实例1:lnp介导的dcas9 sam递送
[0355]
在该实例中,我们集中于使用dcas9(催化上死亡的cas9)协同激活介体(sam)系统上调基因表达。在该系统中,数个激活域相互作用以引起比任一个因素单独诱导的更大的基因响应。组分包括:(1)直接融合到vp64域的dcas9,vp64域是由单纯疱疹病毒蛋白16的四个串联拷贝组成的转录激活子;(2)融合到两种附加激活转录因子的ms2外壳蛋白(mcp),这两种激活转录因子为:热休克因子1(hsf1)和转录因子65(p65);以及(3)含有ms2环的sgrna(包括ms2结合环使得长度从~97个核苷酸增加到~166个核苷酸)。当vp64与在转录起始位点附近结合的蛋白融合时,它会充当强转录激活子。mcp自然地与ms2茎环结合。在该系统中,mcp与工程化到crispr相关联sgrna中的ms2茎环相互作用,并且从而将结合的转录因子运送到适当的基因组位置。
[0356]
该系统的初始迭代使用三个独立的慢病毒来递送三个独立的组分。虽然三组分系统在细胞培养中具有一定的灵活性,但这种设置在动物模型中不太理想。相反,我们首先选择引入dcas9、vp64、mcp、hsf1和p65作为由鼠科动物rosa26启动子驱动的转录物。然后,我们可以通过将重组腺相关病毒(aav)注射到小鼠尾静脉中来引入向导rna,以进行肝脏特异性上调。wt aav通常被认为对基因治疗是安全的,因为它们具有低免疫原性并且具有高度可预测的整合位点(人19号染色体上的aavs1)。然而,为了提高它们作为基因治疗载体的安全性,wt aav的整合能力已被消除,使得这些载体作为附加体保留在宿主细胞核中。出于本实例的目的,所有aav参考均指示重组变体。在引入宿主中后,针对aav的免疫响应通常仅限于中和抗体,而没有明确定义的细胞毒性响应。在非分裂细胞中,这些aav附加体在宿主细胞的生命中保持完整。在分裂细胞中,aav dna通过细胞分裂被稀释,使得需要施用更多的
134,并且seq id no:36(ttrga3)位于ttr转录起始位点的-112。靶向这些向导rna靶序列的单个向导rna分别在seq id no:37、38和39中示出。这些向导被设计为将dcas9 sam组分引导至ttr转录起始位点(tss)上游的100-200bp区域。参见图2。每个向导rna(包括ms2茎环)的结构的一般示意图示于图3(seq id no:45)中。
[0362]
评估了三组小鼠:(1)rosa26-dcas9-sam(未处理);(2)rosa26-dcas9-sam(aav8-gfp);和(3)rosa26-dcas9-sam(aav8-gttr阵列(三个向导靶向ttr))。这些小鼠在八周龄时注射了aav8-gfp或aav8-gttr阵列,并在注射后跟踪至八个月。在各个早期时间点并且然后每月通过elisa测量ttr的血清量,并观察这些动物的任何病理变化。虽然在注射后8个月在这些动物中未观察到病理变化,但到第19天,它们的循环ttr初始增加11倍,到注射后5个月,发现ttr为升高为约4倍的稳定状态水平。如图4所示,用无关病毒处理的dcas9sam小鼠维持大约1000μg/ml的循环ttr,类似于wt小鼠。同时,到第19天,用表达aav的sam向导阵列给药的dcas9 sam小鼠的循环ttr蛋白水平飙升至11,000μg/ml。参见图4。随着病毒颗粒被中和或自然稳态恢复,该水平随着时间的推移缓慢下降。参见图4。无论哪种方式,预期循环ttr蛋白水平将在一年内下降至接近野生型,而无法向究小鼠重新给药。
[0363]
虽然预期lnp上调会持续显著更短的时间,但重新给药的益处可以克服这一限制。考虑到这一点,我们努力表征单个sam ttr sgrna的单个lnp递送可以维持多长时间的蛋白升高。评估了二组小鼠:(1)rosa26-dcas9-sam(未处理);和(2)rosa26-dcas9-sam(r-lnp277-gttr(一个向导靶向ttr))。由向导rna靶向的小鼠ttr基因中的向导rna靶序列(不包括pam)在seq id no:36(gacaataagtagtcttactc)中示出。靶向该向导rna靶序列的单个向导rna在seq id no:55中示出。单个向导rna被修改以在前三个5'和最后三个3'残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。
[0364]
对于lnp调配物,使用(6z,9z,28z,31z)-三十七-6,9,28,31-四烯-19基4-(二甲氨基)丁酸酯(mc3;biofine)、1,2-二硬脂酰-sn-甘油-3-磷酸胆碱(dspc;avanti)、胆固醇(chol;avanti)和在乙醇中50mm的1,2-二肉豆蔻酰-sn-甘油甲氧基聚乙二醇(peg-dmg(2000);nof)的储液。混合这些脂质以产生50:10:38.5:1.5(mc3:dspc:chol:peg-dmg)的摩尔比。grna在10mm柠檬酸钠(ph 5)中制备成225μg/ml。通过使用benchtopnanoassemblr(precision nanosystems公司)的微流体混合,rna和脂质以12ml/min的流速和3:1的rna:脂质体积比混合。将lnp稀释到pbs(ph 7.4)中以稀释乙醇,并且后续使用离心过滤器(amicon,10kd截留)浓缩。通过改良的ribrogreen测定(life technologies公司)对rna进行定量,并在te和具有2%triton x-100的te中对lnp进行定量。通过测量triton-x100样品(总rna)-te样品(游离rna)中的rna来确定总封装rna。在递送至动物之前,lnp通过0.22μm注射器式滤器过滤并在pbs(ph 7.4)中稀释至适当浓度,用于静脉内注射的总体积为200μl。
[0365]
在组(1)中测试3只小鼠,并且在组(2)中测试2只小鼠。这些小鼠在12周龄时注射了在200μl pbs中的1mpk lnp,并在注射后跟踪至67天。在各个早期时间点并且然后每月通过elisa测量ttr的血清量。令人惊讶的是,4,000μg/ml的增加的ttr蛋白水平持续数周保持恒定水平,表明上调远没有预期的那么短暂。参见图5a和5b。值得注意的是,虽然与aav递送相关联的蛋白的初始峰值要高得多,但我们的lnp递送仅包括三种sam ttrsgrna中的一种。此外,通过lnp递送实现的ttr上调在数周内保持在相当恒定的水平,而通过aav递送诱导实
现的上调高度不稳定(初始在19天内急剧增加,并且然后随着时间的推移不断下降)。aav递送诱导了强烈的初始上调并允许表达超过一年,但随着时间的推移它持续下降。lnp递送允许表达快速增加并且稳定数周,之后如果不提供后续给药,蛋白水平可能恢复正常。
[0366]
我们接下来评估了施用不同剂量的lnp的效果。通过lnp以三种剂量将如以上实验中的单个sam ttr sgrna(ttr ga3)引入雄性rosa26-dcas9-sam小鼠:0.5毫克每千克小鼠体重(mpk)、1mpk和2mpk。经由尾静脉注射lnp,以表征单剂量lnp可以维持多长时间的蛋白升高。这种瞬时递送方法产生了大约三周的剂量依赖性基因激活,并使血清ttr水平升高了超过一个月。此外,通过elisa观察到了剂量依赖性升高。最低剂量增加了7倍,而最高剂量增加了15倍。参见图6。进行了第二项研究以评价循序给药的影响。在研究开始时,所有小鼠均被注射了0.5mpk的用ttr ga2配制的lnp,并且每周抽血一次。这些小鼠的子组在两周或四周时再注射另外的0.5mpk lnp,另外注射幼稚小鼠以确认lnp功能(图7a和7b)。在所有情况下,重新给药均成功地增强了ttr表达。在单个时间点成功向动物重新给药而没有不良影响表明对相同动物进行循序给药可能是可行的。进行了一项附加研究,其中小鼠被给药3次:在第0天用0.5mpk的用ttr ga2配制的lnp,在第2周再次用0.5mpk,以及在第4周用0.5mpk的最终给药。我们观察到ttr持续上调超过2倍,持续7周。参见图8。
[0367]
单个lnp被配制为包括负荷物,其包括编码协同激活介体(dcas9-vp64-t2a-mcp-p65-hsf1)的体外转录的mrna和化学合成的靶向ttr的sam sgrna。mrna被加帽和聚腺苷酸化,并且未经修饰或经假尿苷(psu)修饰(所有标准尿嘧啶残基均被假尿苷替换,假尿苷是一种尿嘧啶与碳-碳键而不是氮-碳键相连的尿苷异构体)。sam grna被修饰以在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。在第0天向野生型小鼠注射2mpk的含有靶向ttr(ttrga2)的经修饰的mrna和sam sgrna的lnp或含有未经修饰mrna和sam sgrna的lnp,并在21天内测量ttr的血清水平。使用未处理的野生型小鼠作为阴性对照。如图9所示,含有靶向ttr的经修饰的mrna和sam sgrna的lnp到第6天成功地将ttr血清水平从低于1000μg/ml提高到超过3000μg/ml,并且含有靶向ttr的未经修饰的mrna和sam sgrna的lnp在较小程度上增加了ttr血清水平。单剂量的这种上调持续至少2周。
[0368]
然后生成如上配制的单个lnp以包括负荷物,其包括编码协同激活介体(dcas9-vp64-t2a-mcp-p65-hsf1)的体外转录的mrna和靶向相同基因的多个化学合成的sam sgrna。mrna被聚腺苷酸化和加帽(trilink),并且sgrna被修饰以在前三个5'和3'端rna残基处包括2'-o-甲基类似物和3'硫代磷酸酯核苷酸间键。向野生型小鼠注射lnp,并评估sam sgrna靶向的靶基因的表达。
[0369]
sam sgrna与所有其它sam组分的lnp递送是对治疗性dcas9 sam应用的显著增强,因为我们现在可以(1)确保dcas9 sam转录物和sam sgrna着陆在同一细胞中,(2)用调配物/配体掺入介导增加的组织特异性,(3)向生物体重新给药而不必担心免疫响应,以及(4)生成更稳定的表达水平。总之,核酸递送的这种组合以安全且出乎意料的稳定方式极大地增强了潜在的dcas9应用。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。