ostmt1蛋白质在调控植物产量和/或耐逆性中的应用
技术领域
1.本发明属于生物技术领域,具体涉及ostmt1蛋白质在调控植物产量和/或耐逆性中的应用。
背景技术:
2.绿色植物通过光合作用能够将空气中的二氧化碳吸收,通过一系列固碳反应使其中的c原子能够以碳水化合物的形式储存应用,人类依赖植物作为有机碳的最终来源,因此了解植物如何控制碳的同化和长距离运输十分重要。在高等植物中,糖是在成熟叶片的叶肉细胞中产生的,叶肉细胞被称为源器官。异养细胞,如根和种子,是库器官,它们的营养依赖于糖的供应。碳水化合物分配(cp)是植物从源器官向库器官吸收、运输和分配糖类的过程,是植物生长发育的基础,并且这一过程对植物在非生物胁迫和生物胁迫下的耐受性也至关重要,因此了解碳水化合物分配及其基因调控对突破作物产量和抗逆性十分关键。
3.tmt的全称为液泡膜单糖转运体tonoplast monosaccharide transporter,水稻中有四个tmt基因,分别是:ostmt1 os10g0539900,ostmt2 os02g0229400,ostmt3os03g0128932,ostmt4 os11g0620400。水稻的ostmt1编码一个由740个氨基酸组成的蛋白产物,含有12个跨膜螺旋,第6个和第7个中间有一个较大的环,这个环的长度为320个氨基酸残基,几乎是原核生物和真核生物中所有已知单糖转运子的相应结构的4到5倍,在同核细胞同源物中不存在。
4.水稻作为重要的经济作物,提高水稻产量对我国粮食供应十分关键,同时这也是当下的研究热点。由于现在对ostmt蛋白在经济作物上的研究还十分欠缺,因此,水稻的液泡膜单糖转运蛋白ostmt在水稻生长发育过程中起什么作用?对水稻籽粒有什么影响?在面对非生物及生物胁迫时,ostmt蛋白会起什么作用?这些问题的研究对培育优良水稻品种具有重要的应用价值。
技术实现要素:
5.本发明的第一个目的是提供ostmt1蛋白质与ostmt1蛋白质相关的生物材料的新用途。
6.本发明提供了ostmt1蛋白质与ostmt1蛋白质相关的生物材料在调控植物产量和/或耐逆性中的应用;
7.所述ostmt1蛋白质是如下a)或b)或c)或d)所示的蛋白质:
8.a)由序列表中序列3所示的氨基酸序列组成的蛋白质;
9.b)在序列表中序列3所示的蛋白质的n端和/或c端连接标签得到的融合蛋白;
10.c)将序列表中序列3所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且功能相同的蛋白质;
11.d)与a)-c)中任一所限定的氨基酸序列具有99%以上、95%以上、90%以上、85%以上或者80%以上同源性且具有相同功能的蛋白质;
12.所述生物材料为下述a1)至a8)中的任一种:
13.a1)编码ostmt1蛋白质的核酸分子;
14.a2)含有a1)所述核酸分子的表达盒;
15.a3)含有a1)所述核酸分子的重组载体;
16.a4)含有a2)所述表达盒的重组载体;
17.a5)含有a1)所述核酸分子的重组微生物;
18.a6)含有a2)所述表达盒的重组微生物;
19.a7)含有a3)所述重组载体的重组微生物;
20.a8)含有a4)所述重组载体的重组微生物。
21.上述应用中,a1)所述核酸分子为如下1)或2)或3)所示的基因:
22.1)其编码序列是序列1所示的基因组dna分子或序列2所示的cdna分子;
23.2)与1)限定的核苷酸序列具有75%或75%以上同一性,且编码所述ostmt1蛋白质的cdna分子或基因组dna分子;
24.3)在严格条件下与1)或2)限定的核苷酸序列杂交,且编码所述ostmt1蛋白质的cdna分子或基因组dna分子。
25.本发明的第二个目的是提供b1或b2所示的物质在降低植物产量和/或耐逆性中的应用:
26.b1、抑制或降低植物中ostmt1蛋白质活性或者含量的物质;
27.b2、抑制或降低植物中ostmt1蛋白质编码核酸表达的物质或敲除植物中ostmt1蛋白质编码核酸的物质。
28.上述应用中,所述调控植物产量体现为调控植物百粒重和/或50粒重和/或粒长和/或粒宽和/或粒厚;
29.所述耐逆性为耐冷性;
30.所述降低植物产量体现为降低或减小植物百粒重和/或50粒重和/或粒长和/或粒宽和/或粒厚。
31.本发明还提供了ostmt1蛋白质或生物材料在培育产量提高和/或耐逆性提高的转基因植物中的应用。
32.本发明还提供了上述b1或b2所示的物质在培育产量降低和/或耐逆性降低的转基因植物中的应用。
33.本发明还有一个目的是提供一种培育产量降低和/或耐逆性降低的转基因植物的方法。
34.本发明提供的培育产量降低和/或耐逆性降低的转基因植物的方法包括降低受体植物中ostmt1蛋白质的含量和/或活性,得到转基因植物的步骤;所述转基因植物的产量和/或耐逆性低于所述受体植物。
35.进一步的,所述转基因植物的产量低于所述受体植物体现为所述转基因植物的百粒重和/或50粒重和/或粒长和/或粒宽和/或粒厚小于所述受体植物。
36.所述耐逆性为耐冷性;所述转基因植物的耐冷性低于所述受体植物体现为在冷处理恢复后,所述转基因植物中叶片恢复正常的植株数少于所述受体植物。所述冷处理恢复具体条件为4℃冷胁迫处理3天后恢复2天。
37.所述降低受体植物中ostmt1蛋白质的含量和/或活性的方法通过对所述受体植物中ostmt1蛋白质的编码基因进行敲除或抑制或沉默来实现。
38.所述ostmt1蛋白质的编码基因的核苷酸序列是序列1所示的dna分子。
39.更进一步的,敲除所述受体植物中的ostmt1蛋白质的编码基因的物质为crispr/cas9系统;
40.所述crispr/cas9系统中sgrna的靶序列为序列表中的序列4。
41.上述任一所述应用或方法中,所述植物为单子叶植物或双子叶植物;所述单子叶植物为禾本科植物;所述禾本科植物为水稻。
42.本发明最后一个目的是提供一种特异sgrna或含有所述sgrna编码基因的表达盒、载体、宿主细胞、工程菌或转基因植物细胞系;所述sgrna的靶序列为序列4。
43.本发明通过构建ostmt1敲除载体,获得ostmt1突变体,并对ostmt1突变体的产量和耐逆性进行了分析和研究。实验结果表明:ostmt1蛋白质可调控植物百粒重、50粒重、粒长、粒宽、粒厚和耐冷性。本发明对改良植物产量、培育耐逆植物品种尤其是耐冷植物品种具有重要价值。
附图说明
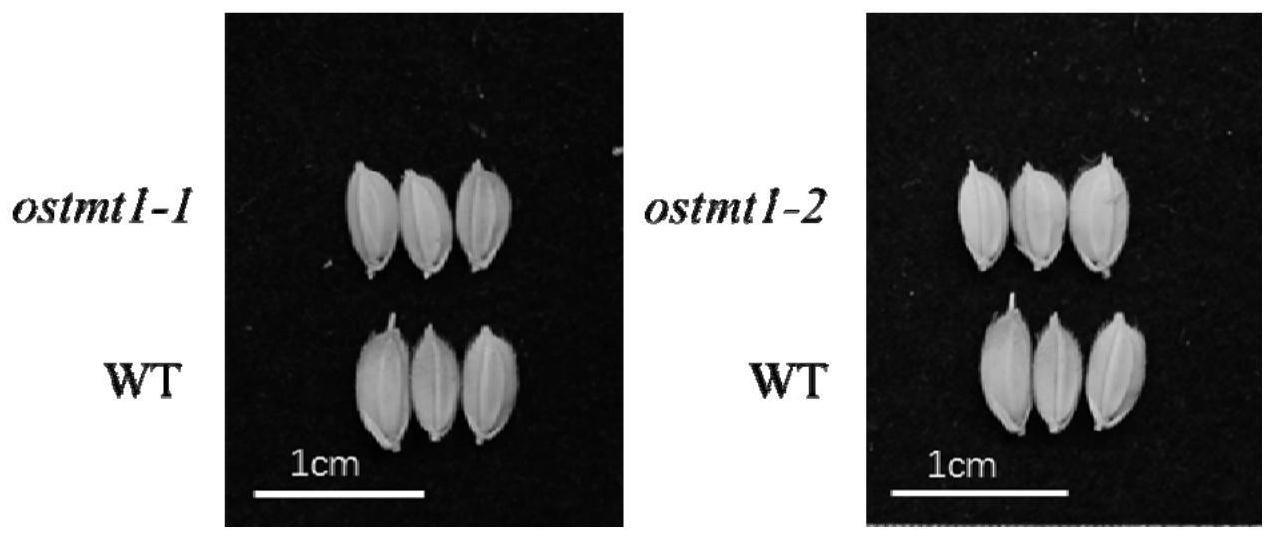
44.图1为水稻ostmt1敲除突变体ostmt1-1和ostmt1-2的种子表型。
45.图2为水稻ostmt1-1和ostmt1-2突变体t2代种子(100粒)的粒重(a)、粒长(b)、粒宽(c)、粒厚(d)的表型统计结果。
46.图3为水稻ostmt1-2和ostmt1-5突变体t3代种子(50粒)的粒重(a)、粒长(b)、粒宽(c)、粒厚(d)的表型统计结果。
47.图4为低温处理前(a),恢复2天(b),恢复9天(c)后的野生型wt和ostmt1-1,ostmt1-2,ostmt1-3的表型。
48.图5为低温胁迫处理wt后ostmt1的rna表达量变化。
49.图6为盐胁迫处理wt后ostmt1的rna表达量变化。
具体实施方式
50.以下的实施例便于更好地理解本发明,但并不限定本发明。下述实施例中的试验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂商店购买得到的。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。
51.下述实施例中的pos-sgrna载体和ph-ubi-cas9-7载体均记载于文献“miao j,guo d,zhang j,huang q,qin g,zhang x,wan j,gu h,qu lj.targeted mutagenesis in rice using crispr-cas system.cell res.2013oct;23(10):1233-6.doi:10.1038/cr.2013.123.epub 2013 sep 3.pmid:23999856;pmcid:pmc3790239.”中。
52.下述实施例中的水稻粳稻品种kitaake记载于文献“miao j,guo d,zhang j,huang q,qin g,zhang x,wan j,gu h,qu lj.targeted mutagenesis in rice using crispr-cas system.cell res.2013oct;23(10):1233-6.doi:10.1038/cr.2013.123.epub 2013 sep 3.pmid:23999856;pmcid:pmc3790239.”中。
53.下述实施例中的农杆菌eha105记载于文献“wu c.,sui y.(2019)efficient and fast production of transgenic rice plants by agrobacterium-mediated transformation.in:kumar s.,barone p.,smith m.(eds)transgenic plants.methods in molecular biology,vol1864.humana press,new york,ny.https://doi.org/10.1007/978-1-4939-8778-8_7”中。
54.实施例1、ostmt1敲除突变体的构建及其表型分析
55.一、ostmt1敲除突变体的构建
56.1、ostmt1敲除载体的构建
57.为了阐明ostmt1蛋白在水稻的生长发育过程中可能的作用,利用crispr技术,通过crispr-p在线网站(http://cbi.hzau.edu.cn/cgi-bin/crispr),筛选水稻ostmt1(loc_os10g0539900)的crispr靶点,并设计引物进行载体构建。具体步骤如下:
58.1)crispr靶点的设计
59.筛选的crispr靶点序列如下:gcagtcctctgatgtccgtc(序列4)。
60.2)oligo二聚体的合成
61.在10ul体系中各加入1p的39440-crispr-1-f引物及39440-crispr-1-r引物,94℃10min,0.1℃/s退火至15℃,15℃保持10min,完成退火,得到退火产物。引物序列如下:
62.39440-crispr-1-f:
63.agatgatccgtggcagcagtcctctgatgtccgtcgttttagagctatgc;
64.39440-crispr-1-r:
65.gcatagctctaaaacgacggacatcagaggactgctgccacggatcatct。
66.3)敲除载体的构建
67.取1ul退火产物与经asci酶切后的pos-sgrna载体infusion连接,得到连接产物,然后转入dh5α,涂于kana固体培养基上。经过电泳确定好质粒大小(15kb左右)后再送测序,测序正确的质粒命名为ostmt1-sgrna。
68.将ostmt1-sgrna与ph-ubi-cas9-7进行gateway lr反应(两种质粒各0.5ul,lr酶1ul,补充至5ul体系,25℃反应1h),再次转入dh5α,涂于spec固体培养基上。测序正确的质粒命名为ostmt1敲除质粒。
69.敲除质粒的详细构建步骤可参考文献“miao j,guo d,zhang j,huang q,qin g,zhang x,wan j,gu h,qu lj.targeted mutagenesis in rice using crispr-cas system.cell res.2013 oct;23(10):1233-6.doi:10.1038/cr.2013.123.epub 2013 sep 3.pmid:23999856;pmcid:pmc3790239.”中的方法。
70.2、农杆菌转化及ostmt1敲除突变体的获得
71.将ostmt1敲除质粒转入农杆菌eha105进行水稻转基因试验,以水稻粳稻品种kitaake(oryza sativa l.cv.kitaake)为受体材料,对其单糖转运蛋白ostmt1进行编辑,得到t0代转基因植株。对t0代转基因植株进行dna检测(dna检测的引物序列如下:39440-crispr-identify-1-f:atcatcacgacgttctctggag;39440-crispr-identify-1-r:tgctgggtaagacatacaaacct),确定转基因植物编辑方式及纯杂合,获得t1代种子,在温室进行播种,对t1代水稻再次进行dna检测并测序,检测编辑方式是否稳定遗传及植株编辑位点的纯杂合,对于纯合编辑的水稻在收获t2代种子后开始进行表型观察。
72.通过测序鉴定,最终获得了t1代ostmt1纯合突变体ostmt1-crispr#1(简称ostmt1-1)、ostmt1-crispr#2(简称ostmt1-2)、ostmt1-crispr#3(简称ostmt1-3)和ostmt1-crispr#5(简称ostmt1-5)。
73.与野生型水稻kitaake基因组dna相比,ostmt1-1的差异仅在于对应于序列2所示的ostmt1基因序列发生了5个碱基“ggaca”的缺失(即序列2自5’末端起第381至385位缺失),从而造成移码并提前终止,ostmt1蛋白功能缺失。
74.与野生型水稻kitaake基因组dna相比,ostmt1-2的差异仅在于对应于序列2所示的ostmt1基因序列发生了1个碱基“g”的缺失(即序列2自5’末端起第381位缺失),从而造成移码并提前终止,ostmt1蛋白功能缺失。
75.与野生型水稻kitaake基因组dna相比,ostmt1-3的差异仅在于对应于序列2所示的ostmt1基因序列发生了1个碱基“g”的插入(即序列2自5’末端起第382位和第383位之间插入了一个碱基g),从而造成移码并提前终止,ostmt1蛋白功能缺失。
76.与野生型水稻kitaake基因组dna相比,ostmt1-5的差异仅在于对应于序列2所示的ostmt1基因序列第379位发生了一个碱基突变(即序列2自5’末端起第379位由碱基a突变为碱基c),且发生了6个碱基“cggaca”的缺失(即序列2自5’末端起第380-385位缺失),从而导致2个氨基酸的缺失,ostmt1蛋白功能缺失。
77.二、ostmt1敲除突变体的表型分析
78.1、ostmt1的缺失会造成种子减产,籽粒减小
79.将t1代ostmt1纯合突变体ostmt1-crispr#1(ostmt1-1)和ostmt1-crispr#2(ostmt1-2)和ostmt1-crispr#5(ostmt1-5)后代进行实验。检测t2代和t3代ostmt1纯合突变体种子的农艺性状,表型结果见图1。同时以野生型水稻粳稻品种kitaake作为对照。t2代统计了ostmt1-1和ostmt1-2的百粒重,结果如图2所示。t3代统计了ostmt1-5的50粒重,结果如图3所示。
80.在百粒重方面,ostmt1-1和ostmt1-2与野生型wt的平均百粒重分别为1.79888(ostmt1-1)、1.82615(ostmt1-2)和1.91349(wt),ostmt1-1和ostmt1-2与野生型wt都有极显著差异,明显减小。
81.在50粒重方面,ostmt1-5与野生型wt的平均50粒重分别为0.75587(ostmt1-5)和0.957(wt),ostmt1-5的50粒重与野生型wt差异极显著,明显减少。
82.2、ostmt1对水稻籽粒的粒长影响较大,其缺失会造成粒长显著变短
83.t2代统计了ostmt1-1和ostmt1-2的粒长、粒宽和粒厚。结果如图2所示。
84.在粒长方面,ostmt1-1和ostmt1-2与野生型wt的平均粒长分别为6.345(ostmt1-1)、6.506(ostmt1-2)和6.77(wt),ostmt1-1和ostmt1-2与wt都有极显著差异,明显减小。
85.在粒宽方面,ostmt1-1和ostmt1-2与野生型wt的平均粒宽分别为3.1477(ostmt1-1)、3.187(ostmt1-2)和3.464(wt),ostmt1-1与wt有极显著差异,ostmt1-2与wt有显著差异,趋势为缩短。
86.在粒厚方面,ostmt1-1和ostmt1-2与野生型wt的平均粒厚分别为2.119(ostmt1-1)、2.101(ostmt1-2)和2.19(wt),ostmt1-1与wt差异不显著,ostmt1-2与wt有显著差异,趋势为缩短。
87.t3代统计了ostmt1-2和ostmt1-5的粒长、粒宽和粒厚。结果如图3所示。
88.在粒长方面,ostmt1-2和ostmt1-5与野生型wt的平均粒长分别为6.929(ostmt1-2)、6.921(ostmt1-5)和7.299(wt),ostmt1-2和ostmt1-5都明显小于wt,差异极显著。
89.在粒宽方面,ostmt1-2和ostmt1-5与野生型wt的平均粒宽分别为3.017(ostmt1-2)、3.173(ostmt1-5)和3.221(wt),ostmt1-2和ostmt1-5都小于wt,ostmt1-2与wt的差异极显著,ostmt1-5与wt的差异不显著。
90.在粒厚方面,ostmt1-2和ostmt1-5与野生型wt的平均粒厚分别为1.7189(ostmt1-2)、1.949(ostmt1-5)和2.093(wt),ostmt1-2和ostmt1-5都明显小于wt,差异极显著。
91.通过两代种子表型观察,发现水稻ostmt1敲除突变体的种子与野生型在重量上有极显著差别,在种子的大小方面,粒长上的差别更显著且稳定遗传,而粒宽和粒厚方面的差异不够显著且不稳定,但趋势都是小于野生型。因此水稻中ostmt1的缺失会影响水稻种子重量下降造成种子减产的表型,ostmt1对于水稻产量大小起关键作用。
92.三、ostmt1敲除突变体的耐逆性分析
93.1、ostmt1与水稻冷胁迫相关
94.将野生型与t2代ostmt1纯合突变体ostmt1-1,ostmt1-2,ostmt1-3在28℃光照培养箱中用木村b营养液培养至16天,然后在恒温的4℃低温水浴锅中冷胁迫处理3天,处理后再放回光照培养箱恢复2天及9天后观察表型。同时以野生型水稻粳稻品种kitaake作为对照。每个材料统计32个植株,共计128株。
95.结果发现,处理前wt与ostmt1纯合突变体植株无明显差别。处理并恢复2天后,ostmt1纯合突变体ostmt1-1和ostmt1-2植株全部卷叶,ostmt1-3植株约2株叶片恢复正常,wt有少量(约4株)叶片恢复正常,但也是大量卷叶。处理并恢复9天后,所有卷叶并未恢复,其余正常生长(图4)。
96.通过q-pcr技术,检测低温处理后的野生型水稻粳稻品种kitaake的ostmt1 rna水平表达量,发现4℃处理3h后的表达量的下降明显,且变化的趋势和幅度有差别,但在处理时间为0-3h时ostmt1在rna水平的表达均上升,3h后下降明显(图5),然后随着处理时间增加,ostmt1虽然存在几次增加然后降低的变化,但都没有恢复到处理前的水平。通过以上结果推测,在短时间的冷胁迫下,ostmt1通过增加表达量,响应植物抗寒机制。
97.2、ostmt1能响应水稻盐胁迫
98.对在木村b营养液中生长至两周的野生型水稻粳稻品种kitaake进行浓度为250mm nacl的盐胁迫处理,并通过q-pcr技术进行了ostmt1的rna表达量的检测,检测其是否与植物抗盐反应相关。结果发现在盐胁迫下ostmt1的rna表达量从处理后1h后开始发生明显上升,在处理3h时表达量最高约为0h时的2倍,6h和24h时有所下降,但仍高于未处理时(图6)。由此推测ostmt与水稻的盐胁迫下的应激反应有关。
99.以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明技术原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。