1.本发明涉及微生物基因工程领域,具体涉及含有烯醇激酶基因和异戊烯基磷酸激酶基因的重组菌株及其在萜类化合物生产中的应用。
背景技术:
2.萜类化合物是一类种类繁多、功能多样的化合物,广泛应用于医药、农业、生物燃料、香料和保健品等各个领域,具有重要的生理活性和极高的经济与医药战略价值。一些萜类化合物已经是经过批准的临床用药,还有很多正在进行临床测试。研究者们利用微生物生产萜类取得了很大进步,是目前公认的低成本、高效率、可持续生产的最有效方式。虽然在工程菌中已经实现了多种萜类的异源合成,但产量较低,无法满足巨大的市场需求。因此,如何对工程菌进行改造,进一步提高萜类化合物的产量,将为解决利用微生物工业化生产高价值萜类的技术瓶颈奠定基础。
3.萜类生物合成中面临的主要瓶颈之一是前体分子生成量不足。萜类的共同前体为异戊烯焦磷酸(ipp)和二甲基丙烯基焦磷酸(dmapp),二者缩合成萜类骨架,是整个生物合成途径的关键节点,起到承上启下的作用。ipp和 dmapp的自然合成路径为甲羟戊酸途径(mva)和甲基赤藓糖醇4-磷酸途径 (mep),二者与细胞的中心碳代谢密切相关并受其动态调控,不可避免地受牵制于碳源、低能效和生物体内其他代谢途径,因此以自然合成途径为切入点,增强前体的代谢通量,不能从根本上解决问题。
4.为了突破这一瓶颈,本发明构建了全新的异戊烯醇途径(iap),该途径仅由两种激酶组成,能将异戊烯醇催化为异戊烯磷酸(ip、dmap)和萜类化合物前体分子(ipp、dmapp)。而且在酵母工程菌中引入iap途径合成前体,能够脱离于中心碳代谢;整个合成路径简单,只需两种酶和一个辅因子参与,可解决原代谢途径(mva途径和mep途径)冗长及中间产物对酶毒性的问题,构建酵母平台菌株,为产业化合成萜类化合物提供基础。
技术实现要素:
5.针对现有技术的不足,本发明提供含有烯醇激酶基因和异戊烯基磷酸激酶基因的重组菌株及其在萜类化合物生产中的应用。
6.本发明的目的通过下述技术方案实现:
7.本发明一方面,提供一种重组质粒,所述重组质粒包括烯醇激酶基因和异戊烯基磷酸激酶基因。
8.作为优选地实施方式,所述烯醇激酶基因包括下述的至少一个基因:
9.(1)scck基因,其核苷酸序列如seq id no:1所示,其编码的氨基酸序列如seq id no:2所示,来源于酿酒酵母菌(saccharomyces cerevisiae);
10.(2)seck基因,其核苷酸序列如seq id no:3所示,其编码的氨基酸序列如seq id no:4所示,来源于真贝酵母(saccharomyces eubayanus)。
11.作为优选地实施方式,所述异戊烯基磷酸激酶基因包括下述的至少一个基因:
12.(1)atipk基因,其核苷酸序列如seq id no:5所示,其编码的氨基酸序列如seq id no:6所示,来源于拟南芥(arabidopsis thaliana);
13.(2)csipk基因,其核苷酸序列如seq id no:7所示,其编码的氨基酸序列如seq id no:8所示,来源于大麻(cannabis sativa)。
14.本发明又一方面,提供上述重组质粒的构建方法,将所述烯醇激酶基因和所述异戊烯基磷酸激酶基因分别通过融合pcr的方式与启动子、终止子组合成表达框,再利用gibson组装的方式与载体骨架连接,即得到重组质粒;
15.优选地,所述烯醇激酶基因的启动子为gal1启动子;
16.优选地,所述异戊烯基磷酸激酶基因的启动子为gal10启动子;
17.在本发明的一种实施方式中,所述重组质粒为酵母菌-大肠杆菌之间的穿梭载体,在大肠杆菌中的抗性标记为氨苄青霉素(ampr),在酵母菌中的筛选标记为亮氨酸(leu)缺陷型互补。
18.本发明又一方面,提供一种重组菌株,所述重组菌株包含烯醇激酶基因和异戊烯基磷酸激酶基因;
19.优选地,所述重组菌株选自酵母菌或大肠杆菌。
20.作为优选地实施方式,所述烯醇激酶基因包括下述的至少一个基因:
21.(1)scck基因,其核苷酸序列如seq id no:1所示,其编码的氨基酸序列如seq id no:2所示,来源于酿酒酵母菌(saccharomyces cerevisiae);
22.(2)seck基因,其核苷酸序列如seq id no:3所示,其编码的氨基酸序列如seq id no:4所示,来源于真贝酵母(saccharomyces eubayanus)。
23.在本发明的技术方案中,所述烯醇激酶基因的编码产物即烯醇激酶能够将烯醇底物磷酸化。
24.作为优选地实施方式,所述烯醇激酶作用的底物包括3-甲基-3-丁烯-1-醇 (isoprenol)和3-甲基-2-丁烯醇(prenol);
25.在本发明的技术方案中,所述烯醇激酶将3-甲基-3-丁烯-1-醇(isoprenol) 磷酸化为异戊烯基单磷酸(ip);
26.在本发明的技术方案中,所述烯醇激酶将3-甲基-2-丁烯醇(prenol)磷酸化为二甲基丙烯基单磷酸(dmap)。
27.作为优选地实施方式,所述异戊烯基磷酸激酶基因包括下述的至少一个基因:
28.(1)atipk基因,其核苷酸序列如seq id no:5所示,其编码的氨基酸序列如seq id no:6所示,来源于拟南芥(arabidopsis thaliana);
29.(2)csipk基因,其核苷酸序列如seq id no:7所示,其编码的氨基酸序列如seq id no:8所示,来源于大麻(cannabis sativa)。
30.在本发明的技术方案中,所述异戊烯基磷酸激酶基因的编码产物即异戊烯基磷酸激酶能够将异戊烯基磷酸底物磷酸化为萜类前体分子。
31.作为优选地实施方式,所述异戊烯基磷酸激酶作用的底物包括异戊烯基单磷酸(ip)和二甲基丙烯基单磷酸(dmap);
32.在本发明的技术方案中,所述异戊烯基磷酸激酶将异戊烯基单磷酸(ip) 磷酸化
为萜类前体分子异戊烯焦磷酸(ipp);
33.在本发明的技术方案中,所述异戊烯基磷酸激酶将二甲基丙烯基单磷酸 (dmap)磷酸化为萜类前体分子二甲基丙烯基焦磷酸(dmapp)。
34.在本发明的一种实施方式中,利用融合pcr的方式,将合成番茄红素 (lycopene)相关的三个基因crte(香叶基香叶基焦磷酸合酶,ggpp合成酶)、 crtb(八氢番茄红素合成酶)、crti(八氢番茄红素脱氢酶)分别与gal10启动子、gal1启动子、gal7融合并组装成一个表达框;再利用crispr-cas9技术将该表达框敲入酵母基因组gal80的位置,获得产lycopene的重组菌 gly80。
35.在本发明的一种实施方式中,采用peg/liac法将上述重组质粒转入酵母细胞中,并在sc-leu的平板上进行筛选;平板上生长的粉红色单克隆即为引入iap途径的产番茄红素(lycopene)的酵母工程菌。
36.本发明又一方面,提供上述重组质粒或上述重组菌株在萜类化合物生产中的应用。
37.本发明又一方面,提供一种利用重组菌株生产萜类化合物的方法,包括如下步骤:
38.在培养基中培养上述重组菌株;加入烯醇底物;从所述培养基中分离所述萜类化合物。
39.作为优选地实施方式,所述烯醇底物选自3-甲基-3-丁烯-1-醇(isoprenol) 和3-甲基-2-丁烯醇(prenol)中的一种或两种;
40.优选地,所述底物添加的时间为培养基中重组菌的od600为4~5。
41.优选地,所述烯醇底物为3-甲基-3-丁烯-1-醇(isoprenol)和3-甲基-2-丁烯醇(prenol)时,其摩尔比为3-甲基-3-丁烯-1-醇(isoprenol):3-甲基-2
‑ꢀ
丁烯醇(prenol)=3:1。
42.在本发明的一种实施方式中,所述方法是挑取酵母菌作为重组菌株,将酵母重组菌株单克隆接种至氨基酸缺陷型培养基(sc-leu 2%葡萄糖)中活化,在30℃,800rpm下培养16小时。24小时后转接至新的氨基酸缺陷型培养基 (sc-leu 2%葡萄糖)中扩大培养,起始od为0.2,继续培养24小时后,培养基中酵母菌的od600为4~5,加入25mm的底物(isoprenol和/或prenol) 和10%的十二烷(v/v)。
43.本发明具有以下优点和有益效果:
44.本发明通过以下研究:(1)通过生物信息学的方法,选择多组与iap途径相关的酶基因,并构建其组合表达框。(2)向酿酒酵母中引入新的合成途径iap,该途径用于积累中间体ipp和dmapp以及对iap途径相关的两种酶 (烯醇激酶和磷酸激酶)进行筛选。(3)利用iap途径提高酵母底盘中萜类化合物的产量。(4)提供一种培养发酵条件,使iap途径在酵母底盘中得到高效利用。
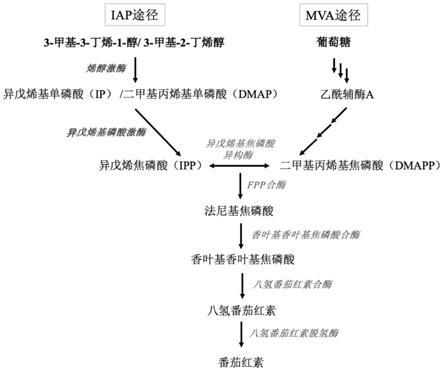
45.与现有技术相比,本发明具有如下优点:
46.本发明通过在酿酒酵母(saccharomyces cerevisiae)细胞中引入一条新的代谢途径:异戊烯醇途径(isoprenoid alcohol pathway,iap),来合成ipp和 dmapp。iap途径只需2步反应和一个辅因子(atp)的参与,并脱离于中心碳代谢;使萜类的生物合成过程具有更强的代谢流,最终帮助提高萜类化合物的产量。
47.(1)该途径脱离中心碳代谢,不与细胞内其它代谢途径竞争,有利于中间体ipp和
dmapp积累。
48.(2)该途径只需要2步反应和一个辅因子(atp)参与,有利于产物快速积累。
附图说明
49.图1是利用异戊烯醇途径(iap)途径和mva途径在酿酒酵母中合成番茄红素的示意图;
50.图2是实施例1中构建的番茄红素基因表达框;
51.图3是实施例1中酿酒酵母敲入工具cas9-sgrna载体;
52.图4是实施例2中构建的重组质粒图谱;
53.图5是实施例5中以3-甲基-3-丁烯-1-醇为底物的含有不同重组质粒的酵母合成番茄红素的产量对比图;
54.图6是实施例5中以3-甲基-3-丁烯-1-醇和3-甲基-2-丁烯醇为底物的含有不同重组质粒的酵母合成番茄红素的产量对比图。
具体实施方式
55.下面结合实施例以及对比例,对本发明作进一步地详细说明,但本发明的实施方式不限于此。具体包括以下实施例:
56.为了使本发明要解决的技术问题、技术方案及有益效果更加清楚明白,以下结合实施例与附图,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅用以解释本发明,并不用于限定本发明。
57.实施例1:异源合成四萜化合物(番茄红素)酵母底盘的构建
58.用高保真酶(max super-fidelity dna polymerase)pcr获得启动子(p
gal1
、p
gal10
、p
gal7
),终止子(t
adh1
、t
cyc1
、t
tdh2
)和异源基因crte (香叶基香叶基焦磷酸合酶,ggpp合成酶)、crtb(八氢番茄红素合酶)和 crti(八氢番茄红素脱氢酶)。通过融合pcr,将上述元件组装 p
gal10-crte-t
adh1
、p
gal1-crtb-t
cyc1
、p
gal7-crti-t
tdh2
并构建成外源基因表达框crte-crtb-crti,结构如图2所示。切胶回收,获得纯化的目的片段。
59.采用crispr/cas9工具将上述外源基因表达框crte-crtb-crti整合到野生型酿酒酵母cen.pk2-1c基因组gal80位置,得到产萜酵母底盘。
60.本实施例中的敲入载体pcut-gal80为本领域常规的重组载体(图3所示),并含有尿嘧啶筛选标记,其能够转化尿嘧啶营养缺陷型酵母。
61.在进行基因组敲入转化时,目的片段为1μg,pcut-g80载体为250ng。转化方法为peg/liac法。为了保证效率,酿酒酵母转化所用的感受态细胞现制现用。具体过程如下:
62.(1)从ypd平板挑取野生型酵母cen.pk2-1c单克隆于10ml 1*ypd 培养基,30℃,200rpm下过夜培养。
63.(2)第二天上午,取一定量(1)中发酵液转接至50ml新鲜的2*ypd 液体培养基中,转接后的初始od600为0.2;30℃,200rpm培养4.5小时左右。
64.(3)测得od600为1左右,低速离心(3000rpm,10min)收集菌体。
65.(4)用等体积的灭菌水将上述菌体清洗一遍,低速离心(3000rpm,10 min)收集菌体。再用1/2体积的0.1m lioac将上述菌体清洗一遍,低速离心收集菌体。
66.(5)冰上分装为50μl/5od感受态细胞。
67.(6)向上述50μl感受态细胞中加入260μl peg3350(50%w/v),36μl 1m lioac,10μl单链鲑鱼精dna(100℃温育5min解链),1μg敲入片段, 250ng pcut-gal80载体。
68.(7)将该混合体系上下颠倒并涡旋震荡均匀后放于42℃水浴锅中温育50 分钟。
69.(8)收集上述混合体系,低速离心(4000rpm,2min)收集菌体;1ml 灭菌水清洗一遍后,再次低速离心收集菌体。
70.(9)100μl灭菌水重悬菌体,涂布于sc-ura平板。待平板吹干后倒置于30℃培养箱中3-4天。
71.上述转化平板所长出的红色单克隆进行脱pcut、pcr鉴定和测序鉴定,经过序列比对,萜类外源基因表达框已成功、正确的敲入野生型酵母基因组 gal80位点。
72.本实施例中所获得的产四萜化合物(番茄红素)酵母底盘命名为gly80。
73.实施例2:iap途径功能元件的组装和表达
74.通过融合pcr将不同来源的烯醇激酶(iak)基因、磷酸激酶(ipk)基因与双向强启动子p
gal10-gal1
组装成iak-ipk表达框。采用gibson组装技术将iak-ipk表达框与低拷贝载体prs415-leu2(cen/ars)重组,得到重组质粒prs415-iak-ipk(结构如图4所示)。将这些重组质粒转入上述产萜酵母底盘gly80中进行表达,转化方法同上述实施例1。
75.本实施例中构建的重组质粒为:prs415-scck-atipk和 prs415-seck-csipk。
76.其中,prs415-scck-atipk质粒组合中具体的基因型为:来源于酿酒酵母菌(saccharomyces cerevisiae)的scck基因(核苷酸序列如seq id no:1 所示,其编码的氨基酸序列如seq id no:2所示)和来源于拟南芥(arabidopsisthaliana)的atipk基因(其核苷酸序列如seq id no:5所示,其编码的氨基酸序列如seq id no:6所示)。
77.prs415-seck-csipk质粒组合中具体的基因型为:来源于真贝酵母菌的 seck基因(核苷酸序列如seq id no:3所示,其编码的氨基酸序列如seq idno:4所示)和来源于大麻的csipk基因(核苷酸序列如seq id no:7所示,其编码的氨基酸序列如seq id no:8所示)。
78.转化时菌体涂布于sc-leu平板,30℃培养箱中3天。
79.实施例3:异戊烯醇途径(iap)在酵母底盘中的应用
80.将上述实施例2中转化的酵母菌涂布平板3天左右长出粉红色单克隆。
81.(1)挑取单克隆于灭菌的24孔板中,24孔板每个孔装载2ml氨基酸缺陷型培养基(sc-leu 2%葡萄糖),30℃,800rpm过夜培养。
82.(2)第二天转接,取一定量上述菌液至新的24孔板中。同上,24孔板每个孔装载2ml氨基酸缺陷型培养基(sc-leu 2%葡萄糖)。转接后的初始od600为0.2;30℃,800rpm培养24小时。
83.(3)24小时后,测得上述菌体生长至od600为4-5左右,向培养基中加入烯醇底物isoprenol和(或)prenol,终浓度为25mm,再加入10%十二烷 (v/v),即200μl。
84.(4)继续培养3-4天。30℃,800rpm。
85.(5)发酵结束后,离心收集菌体。
86.(6)hplc测定番茄红素的产量,并确定在酵母细胞中异戊烯醇途径发挥功能的最佳iak-ipk酶组合。
87.本实施例中,酵母底盘中合成番茄红素的途径,如图1所示,mva途径为酵母底盘合
成萜类的天然途径,其为葡萄糖经由3个乙酰辅酶a通过6步反应生成ipp,ipp在异构酶的作用下进而生成dmapp;ipa途径为以烯醇为底物经过2步反应生成ipp和dmapp。
88.实施例4:萜类产物(番茄红素)提取
89.(1)发酵结束后,将24孔板置于水平落地离心机离心;4000rpm,10min。
90.(2)弃掉上层发酵液,每个孔加入1ml去离子水重旋清洗菌体;4000rpm 10min。
91.(3)重复(2)中操作一次。
92.(4)用1ml 3m盐酸重旋菌体,取出置于1.5ml ep管中。
93.(5)将上述菌体置于100℃金属浴中3min,然后立即置于冰水浴中3min。重复3次。
94.(6)12000rpm离心5min,弃上层液体。
95.(7)加入1ml去离子水重旋清洗菌体,12000rpm离心5min,弃上层液体。重复一次。
96.(8)向收集有菌体的1.5ml ep管中加入500μl丙酮,涡旋震荡10s使细胞彻底分散,萃取其中的番茄红素。然后置于超声清洗仪中超声30min,彻底提取其中的番茄红素。
97.(9)离心收集上层有机相;再加入500μl丙酮,重复(8)一次。
98.(10)将收集的1ml有机相(含目标产物番茄红素)用0.45μm的有机滤头过滤,滤液用于后续的hplc定量分析。
99.实施例5:高效液相色谱(hplc)对萜类产物(番茄红素)定量分析
100.hplc分析条件:色谱柱:ymc carotenoid,4.6*250mm,5.0μm。流动相:a相:90%乙腈,b相:100%甲基叔丁基醚。检测所用机器,安捷伦1260;条件设置:0min,40%b;5min,95%b;10min,95%b;10.01min,40%b; 20min,40%b。上样量,20μl。流速,1ml/min。柱温,30℃。标准品为: 2.5mg/l、10mg/l、25mg/l、50mg/l、100mg/l番茄红素(用丙酮溶解)。
101.根据hplc定量的结果如图5,图6所示,图中,横坐标为酵母底盘类型,纵坐标为番茄红素产量。
102.图5为饲喂25mm 3-甲基-3-丁烯-1-醇底物后的发酵结果。其中底盘g80 和g80(prs415)为对照组;底盘g80(prs415-scck-atipk)和 g80(prs415-seck-csipk)为实验组。图6为饲喂25mm双底物3-甲基-3-丁烯-1-醇 3-甲基-2-丁烯醇(摩尔比3:1)的发酵结果。其中底盘g80和 g80(prs415)为对照组;底盘g80(prs415-scck-atipk)和 g80(prs415-seck-csipk)为实验组。从图5和图6中可以得出结论:将 prs415-scck-atipk和prs415-seck-csipk转入酵母底盘gly80中,可将番茄红素的产量提高1-1.3倍。
103.上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受所述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
104.显然,本领域的技术人员可以对本发明进行各种改动和变型而不脱离本发明的精神和范围。这样,倘若本发明的这些修改和变型属于本发明权利要求及其等同技术的范围之内,则本发明也意图包含这些改动和变型在内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。