1.本发明属于种质创新技术领域,具体涉及一种雄性毛白杨母株与后代关系的鉴定方法及应用。
背景技术:
2.林木种质资源是林木育种的基础,但种业发展基础仍不牢固,保障种质自主可控比过去任何时候都更加紧迫。因此,打牢种质资源基础,做好资源普查收集、鉴定评价工作,坚持保护好、高效利用好种质资源十分重要。毛白杨是我国北方地区重要的绿化和用材乡土树种,北京地区也是其主要的适生区之一,目前乡土资源研发不足、种质资源不清、保护利用不全面、雌株飞絮等问题仍有待解决。
技术实现要素:
3.有鉴于此,本发明的目的在于提供一种雄性毛白杨母株与后代关系的鉴定方法及应用,通过相关分子技术手段,对已选出的北京地区雄性毛白杨古树优株进行遗传多样性分析,研发配套的分子标记技术,有效确定该种源与其子代的亲缘关系,实现毛白杨苗木种源可追溯性,为良种保护及推广提供技术保障。
4.为了实现上述发明目的,本发明提供以下技术方案:本发明提供了一种雄性毛白杨母株与后代关系的鉴定方法,包括以下步骤:创建雄性毛白杨古树优良单株的叶绿体基因组图谱,与已知的毛白杨叶绿体基因组序列进行比对,筛选特异性差异基因位点或基因片段作为分子标记位点,利用若干种分子标记方法确定不同种源古树的优良单株与其子代的亲缘关系。
5.优选的,所述雄性毛白杨古树优良单株的性状包括:长寿雄性、不飞絮、不早衰、健康状态良好、树干通直和造林绿化效果优。
6.优选的,所述雄性毛白杨古树优良单株的树龄超过100年。
7.优选的,所述分子标记方法包括:全长序列标记法、特异性引物序列标记法和/或酶切位点标记法。
8.优选的,对一个或多个分子标记位点进行所述分子标记。
9.优选的,当利用特异性引物序列标记法进行分子标记时,针对昌平区沙岭村古树设计的特异性引物包括sl-2f和sl-2r,所述sl-2f的核苷酸序列如seq id no.1所示,所述sl-2r的核苷酸序列如seq id no.2所示;针对房山区沙窝古树设计的特异性引物包括sw-2f和sw-2r,所述sw-2f的核苷酸序列如seq id no.3所示,所述sw-2r的核苷酸序列如seq id no.4所示;针对大兴前安定古树设计的特异性引物包括ad-2f和ad-2r,所述ad-2f的核苷酸序列如seq id no.5所示,所述ad-2r的核苷酸序列如seq id no.6所示;针对大兴区大狼垡村古树设计的特异性引物包括lf-2f和lf-2r,所述lf-2f的核苷酸序列如seq id no.7所示,所述lf-2r的核苷酸序列如seq id no.8所示;
针对大兴辛房古树设计的特异性引物包括xf-2f和lf-2r,所述xf-2f的核苷酸序列如seq id no.9所示,所述xf-2r的核苷酸序列如seq id no.10所示;针对海淀东北义园古树设计的特异性引物包括dbyy-1f和dbyy-1r,所述dbyy-1f的核苷酸序列如seq id no.11所示,所述dbyy-1r的核苷酸序列如seq id no.12所示。
10.优选的,当利用酶切位点标记法进行分子标记时,昌平区沙岭村古树的特异性双酶切位点包括afei和haeii,大兴辛房古树的特异性双酶切位点包括pcii和afliii。
11.本发明还提供了上述的鉴定方法在追溯毛白杨苗木种源中的应用。
12.有益效果:本发明提供了一种雄性毛白杨母株与后代关系的鉴定方法,通过创建雄性毛白杨古树优良单株的叶绿体基因组图谱,与已发表毛白杨叶绿体基因组序列进行比对,筛选出特异性差异基因位点或片段,分别运用不同的分子标记技术标记不同种源古树的雄性毛白杨遗传多样性,从而确定每个种源与其子代的亲缘关系,实现毛白杨苗木种源可追溯性,为良种推广提供技术保障。
13.本发明实施例中,通过对6株长寿雄性毛白杨古树:大兴区大狼垡村古树(lf)、大兴辛房古树(xf)、大兴前安定古树(ad)、房山区沙窝古树(sw)、昌平区沙岭村古树(sl)和海淀东北义园古树(dbyy)分别构建其叶绿体基因组图谱,通过与已知的杨树叶绿体基因组序列进行比对分析,最终6个单株均找到了多个特异性位点或片段;进一步通过片段克隆、凝胶电泳、片段的凝胶回收、克隆测序等实验,验证了这些特异性位点或片段准确无误。因此,可将这些特异性位点或片段作为分子标记,确定每个优良古树与其子代的亲缘关系,实现毛白杨苗木种源可追溯性,为这些良种推广提供技术保障。同时采用北京地区其他种源的14个毛白杨无性系苗木(图29)作为对照,验证了本发明所述鉴定方法的准确度。对照样品采集于北京市黄垡苗圃,对照毛白杨种源代号分别为0018、0027、0061、0078、0076、0035、0085、0004、0057、0024、0060、0023、0034、0041。
附图说明
14.图1为昌平沙岭村古树冬季(左)和房山沙窝古树(右);图2为大兴前安定古树,左:冬季,右:春季;图3为大兴前辛房古树(左)和大兴大狼垡古树(右);图4为海淀东北义园三株古毛白杨树;图5为东北义园三株白杨二级古树认证挂牌;图6为populus tomentosa dbyy 叶绿体基因组图谱;图7为雄性毛白杨古树种属间所构建的系统发育树;图8为毛白杨古树sl特异性位点标记图;图9为毛白杨古树sl特异性位点酶切位点标记图;图10为毛白杨古树sw特异性位点标记图;图11为毛白杨古树sw特异性位点酶切位点标记图;图12为毛白杨古树ad特异性位点标记图;图13为ad古树单株特异性核苷酸位点分子标记胶图;图14为毛白杨古树lf特异性位点标记图;图15为毛白杨古树lf特异性位点酶切位点标记图;
图16为毛白杨古树xf特异性位点标记图;图17为毛白杨古树xf特异性位点酶切位点标记图;图18为毛白杨古树dbyy特异性位点标记图;图19为毛白杨古树dbyy特异性位点酶切位点标记图;图20为9个毛白杨叶绿体基因组对比图;图21为所选雄性毛白杨古树单株初代培养;图22为增殖及生根培养;图23为所选雄性毛白杨古树单株大田移栽;图24为sl酶切位点标记法确定子代亲缘关系;图25为dbyy与53个杨树叶绿体基因组序列特异性位点或片段对比;图26为sl与53个杨树叶绿体基因组序列特异性位点或片段对比图;图27为sw与53个杨树叶绿体基因组序列特异性位点或片段对比图;图28为xf与53个杨树叶绿体基因组序列特异性位点或片段对比图;图29为14个北京地区其他种源的毛白杨无性系苗木苗圃地照片;图30为sl酶切位点标记法确定子代亲缘关系(以14个不同种源非亲本子代毛白杨为对照)。
具体实施方式
15.本发明提供了一种雄性毛白杨母株与后代关系的鉴定方法,包括以下步骤:创建雄性毛白杨古树优良单株的叶绿体基因组图谱,与已知的毛白杨叶绿体基因组序列进行比对,筛选特异性差异基因位点或片段作为分子标记位点,利用若干种分子标记方法标记不同种源古树在所述分子标记位点的遗传多样性,并确定每个优良古树与其子代的亲缘关系。
16.本发明所述雄性毛白杨古树优良单株的性状优选包括:长寿雄性、不飞絮、不早衰、健康状态良好、树干通直和造林绿化效果优;并且所述古树的树龄优选超过100年。具体的,在本发明实施例中,选用的雄性毛白杨古树优良单株优选如表1所示:大兴区大狼垡村古树(100~300年,图3中右图)、大兴辛房古树(100~300年,图3中左图)、大兴前安定古树(100~300年,图2)、昌平区沙岭村古树(300~500年,图1中左图)、房山区沙窝古树(300~500年,图1中右图)、海淀东北义园古树(300~500年,图4和图5)。
17.表1 北京地区雄性毛白杨收集信息表
本发明对所述叶绿体基因组图谱的构建方法并没有特殊限定,优选包括提取所选雄性毛白杨古树单株的dna,进行叶绿体基因组测序,数据过滤,数据质量评估,基因组组装及基因组注释与分析。本发明优选使用geseq程序(https://chlorobox.mpimp-golm.mpg.de/geseq.html)对叶绿体基因组序列进行注释。在本发明实施例中,具体的方案优选包括:将拼接的全叶绿体序列加入在线程序,运用blatx将其比对到geseq提供的mpi-mp叶绿体基因库(包括:核编码基因,65%相似度和rrna,85%相似度);为了确保编码蛋白和rna基因的预测准确性,分别使用第三方程序hmmer对蛋白和aragorn v1.2.38 对trna基因预测,最终注释结果人工进行校正。根据注释结果使用ogdraw(https://chlorobox.mpimp-golm.mpg.de/ogdraw.html),绘制叶绿体基因组图谱。
18.本发明采用叶绿体基因组测序,叶绿体基因组属于母系遗传,母系遗传对开花植物是稳定的,核基因遗传只有纯合表达才具有稳定遗传。叶绿体基因组非常保守,主要表现在基因组结构、基因排列顺序和基因种类等方面。具有单亲遗传、小分子量、多拷贝、分子进化速率慢等特点,所以本发明所述叶绿体基因组遗传稳定性高。
19.得到叶绿体基因组图谱,本发明将所述叶绿体基因组图谱与已知的毛白杨叶绿体基因组序列进行比对,已知的毛白杨叶绿体基因组信息数据来源于ncbi(national center for biotechnology information (nih.gov)),ncbi号分别为:mk192135、mk251149和mk267301,9个毛白杨叶绿体基因组对比结果如图20所示,可筛选特异性差异基因位点或片段作为分子标记位点。
20.筛选出分子标记位点后,本发明进一步通过片段克隆、凝胶电泳、片段的凝胶回收、克隆测序等实验,验证了这些特异性位点或片段准确无误。因此,可将这些特异性位点或片段作为分子标记,确定每个优良古树与其子代的亲缘关系,实现毛白杨苗木种源可追溯性,为这些良种推广提供技术保障。本发明所述分子标记方法优选包括:全长序列标记法、特异性引物序列标记法和/或酶切位点标记法。本发明可对一个分子标记位点进行所述分子标记,也可同时对多个分子标记位点进行所述分子标记。
21.在本发明中,当利用特异性引物序列标记法进行分子标记时,针对不同的古树设计如表2所示的特异性引物:表2 针对不同古树设计的特异性引物信息
本发明在进行比对时,优选还包括与ad的后代(ddl)进行比对以验证准确度,且在进行比对时,以 mk251149作为consensus。在本发明中,7株毛白杨古树叶绿体基因组比对,sl的特异性位点位置为:45803位点,其他毛白杨该位点为碱基t,sl该位点为碱基g,同时完成标记agcgct位点标记,酶切位点为:afei(agc/gct)、haeii(/gcgc),两个酶切位点均可用,可有效切开片段;sw的特异性位点位置为:53494位点,其他毛白杨该位点为碱基a,sw该位点为碱基g,无可用酶切位点;ad的特异性位点位置为:289位点,其他毛白杨该位点为碱基t,ad该位点为空缺,无可用酶切位点;lf的特异性位点位置为:81838位点,其他毛白杨该位点为碱基a,lf该位点为空缺,无可用酶切位点;xf的特异性位点位置为:64826位点,其他毛白杨该位点为碱基c,xf该位点为碱基t,且完成标记acatgt位点标记,有两个可用酶切位点:pcii(acatgt/tgtaca)和afliii(acrygt/tgyrca),可有效切开片段;dbyy的特异性位点位置为:7309-7314,其他毛白杨该位点为空缺,dbyy该位点为碱基片段attttt,无可用酶切位点。
22.在本发明中,当利用酶切位点标记法进行分子标记时,昌平区沙岭村古树的特异性双酶切位点优选包括afei和haeii,大兴辛房古树的特异性双酶切位点优选包括pcii和afliii。
23.本发明所述鉴定方法,品种及序列比对参照多,精度高,比对共包括72个叶绿体基因组序列(图7),其中68个杨柳科属内、4个杨柳科属外,68个科属内又包括53个杨树、15个柳树,53个杨树内又包括9个毛白杨单株。确定6株古树毛白杨与已有ncbi毛白杨叶绿体基因组数据聚在一起,同时确定其与响叶杨亲缘关系更近,通过与已知的杨树叶绿体基因组序列比对(53个杨树,包括9个毛白杨对比,发现特异性位点或片段,图25~图28),获取每个单株的特异性基因位点和片段,采用全长序列标记法、特异性引物序列标记法和/或酶切位点标记法等标记手段,成功完成所选6株雄性毛白杨古树单株叶绿体基因组分子标记。通过长期对所选雄性毛白杨古树单株进行调查,筛选、雌雄判定及组织培养技术研发,成功获得
其子代(图21~23),取子代嫩叶提取dna,通过本发明的3种及以上方法,可鉴定子代基因是否具备该特异性分子标记位点,若具备则为该种源子代,若不具备则不是其子代。本发明所述鉴定方法分子标记全面且特异性高,采用dna snapgene4.2.4软件进行dna多态性的比对,可直观看出叶绿体基因组上的多态性变异趋势,多态性越高,则基因序列差异较大,反之则基因序列差异很小,标记该种源个体的叶绿体基因组,通过遗传多样性分析疑似为该种源的苗木,若与之一致,则可判定为其子代,通过分析表明毛白杨叶绿体基因组序列稳定性高,差异小,可有效实现分子特异性标记。
24.本发明还提供了上述的鉴定方法在追溯毛白杨苗木种源中的应用。
25.白杨派树种种质资源丰富,但杂而不清、保护利用不全面,种是苗木质量保证的关键,本发明所述毛白杨叶绿体基因组分子标记法,可有效标记所选种源及其无性繁殖子代,在良种推广过程中,可用于确定毛白杨苗木的亲缘关系,追溯苗木种源,避免鱼龙混杂,为雄性古树毛白杨野生乡土资源保护、利用及推广提供有效技术支撑。同时,本发明也对种质资源保护、利用及推广起到重要作用。
26.下面结合实施例对本发明提供的一种雄性毛白杨母株与后代关系的鉴定方法及应用进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
27.实施例11、对表1所示的古树进行叶绿体基因组测序,使用植物dna提取试剂盒提取所选雄性毛白杨古树单株的dna,进行叶绿体基因组测序,数据过滤,数据质量评估,基因组组装及基因组注释与分析。根据注释结果使用ogdraw(https://chlorobox.mpimp-golm.mpg.de/ogdraw.html),绘制叶绿体基因组图谱,以东北义园(dbyy)为例(表3和图6)。完成所选6株长寿雄性毛白杨古树叶绿体基因组测序(表3-8)。
28.表3
ꢀꢀ
populus tomentosa dbyy 叶绿体基因组基本特征统计结果length(bp)156497gccontent(%)36.71atcontent(%)63.28lsclength(bp)84676ssclength(bp)16551irlength(bp)27635genenumber130protein-codinggenenumber85protein-codinggene(%)65.38rrnagenenumber8rrna(%)6.15trnagenenumber37trna(%)28.46genenumberinirregions342、安定(ad)叶绿体基因组测序结果与其子代(ddl)测序结果比对通过古树安定(ad)及其子代(图21~23,下同)叶绿体基因组测定,测序结果一致,确定叶绿体基因组稳定性高,也说明了此次叶绿体基因组测序结果的准确性高。测序结果
如下表4所示。
29.表4 populus tomentosa ad与子代ddl叶绿体基因组基本特征比对表基本特征adddllength(bp)156513156513gccontent(%)36.7236.72atcontent(%)63.2863.28lsclength(bp)8469284692ssclength(bp)1655116551irlength(bp)2763527635genenumber132132protein-codinggenenumber8585protein-codinggene(%)64.3964.39rrnagenenumber88rrna(%)6.066.06trnagenenumber3737trna(%)28.0328.03genenumberinirregions36363、古树sl分子标记3.1古树sl特异性位点比对通过毛白杨具体叶绿体基因组序列比对(图8),sl特异性位点位置为:45803位点(seq id no.13所示序列的545bp),其他毛白杨该位点为碱基t,sl该位点为碱基g,设计引物sl-2f和sl-2r,结果显示扩增产物与seq id no.13所示序列的82~903bp的位点完全符合,序列匹配度100%,特异性位点存在,可进行标记。
30.扩增体系为kod plus-酶(高保真)25μl体系(下同):kod buffer 2.5μl、dntps 2.5μl、mgso41μl、引物各0.5μl、dna 0.4μl、kod酶0.5μl和余量的ddh2o。
31.pcr程序(下同):94℃预变性2min;94℃变性20s,58℃退火30s,68℃延伸1min,35循环;68℃再延伸5min,pcr产物跑胶,胶回收,t克隆(peasy-blunt cloning),阳性送测,测序位点与标记位点一致,位点准确有效。
32.3.2古树sl特异性位点标记1)酶切位点标记法毛白杨属于无性繁殖,无性繁殖不改变原有基因型,且叶绿体基因组稳定性强进化慢,可用于亲缘关系鉴定,标记该种源个体的叶绿体基因组特异性,通过遗传多样性分析疑似为该种源的苗木,若与之一致,则可判定为其子代。
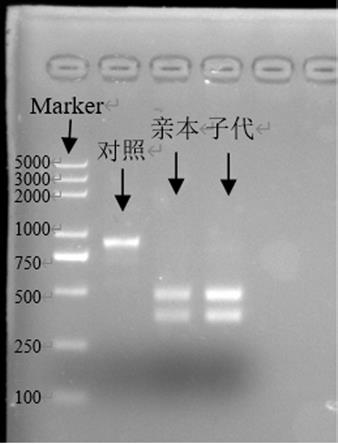
33.标记位点位置:45803(g),完成标记agcgct位点标记,酶切位点为(图9)afei(agc/gct)、haeii(/gcgc),可切除seq id no.13所示序列的82~903bp,结果:两个酶切位点均可用,可有效切开片段。以haeii(/gcgc)酶为例,酶切体系为15μl 体系,dna 5μl、10xne buffer 1.5μl、restriction enzyme 0.5μl和余量的ddh2o,37℃温育30-60 min,产物跑胶,看序。酶切对应结果由图24可看出,对照为非亲本子代,不具备相应的酶切位点,不能被切开,而亲本和子代具备相同且特异的酶切位点,所以可以被切开,原条带为822bp,切开后
为两条300~500bp的条带,可确定sl亲本与子代的关系。并随机挑选古树毛白杨子代苗3株,以来源于北京的14个其他种源的毛白杨苗木为对照,酶切结果如图30,鉴定结果准确,可有效确定sl亲本与子代的关系,更加确定了本发明所述鉴定方法的准确度高,以及应用中的有效性好。2)特异性引物序列标记法利用sl-2f和sl-2r对基因组dna进行pcr,扩增产物822bp,与seq id no.13所示序列的82~903bp序列一致,且结果显示位点准确,该位点可作为标记位点进行标记。(扩增体系同上)3)sl种源全长序列标记法毛白杨古树sl叶绿体基因组全长序列156515 bp,全长包含了36.72%的gc,63.28%的at,lsc长度为84692 bp,ssc长度为16552 bp,ir长度为27635,所含基因数目为132,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表5)。
34.表5 populus tomentosa sl叶绿体基因组基本特征统计结果length(bp)156515gccontent(%)36.72atcontent(%)63.28lsclength(bp)84692ssclength(bp)16552irlength(bp)27635genenumber132protein-codinggenenumber85protein-codinggene(%)64.39rrnagenenumber8rrna(%)6.06trnagenenumber37trna(%)28.03genenumberinirregions364、古树sw分子标记4.1古树sw特异性位点比对通过毛白杨具体叶绿体基因组序列比对,特异性位点位置为:53494位点,其他毛白杨该位点为碱基a,sw该位点为碱基g(图10),设计引物sw-2f和sw-2r进行特异性位点验证,结果显示与seq id no.14所示序列的83~865bp完全符合,序列匹配度100%,特异性位点存在,可进行标记。
35.4.2古树sw特异性位点标记标记位点位置:53494 (g)(图11),sali、acci及hincii等酶切位点均位于特异性位点临近位置,但未包含,均不可用,无可用酶切位点。
36.1)特异性引物序列标记法利用sw-2f和sw-2r进行pcr扩增,片段长度:783bp,与seq id no.14所示序列的83~865bp的序列匹配度100%,结果显示位点准确,可进行标记。
37.2)全长序列标记法毛白杨古树sw叶绿体基因组全长序列156514 bp,全长包含了36.72%的gc,63.28%的at,lsc长度为84691 bp,ssc长度为16553 bp,ir长度为27635,所含基因数目为132,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表6)。
38.表6
ꢀꢀ
populus tomentosa sw叶绿体基因组基本特征统计结果length(bp)156514gccontent(%)36.72atcontent(%)63.28lsclength(bp)84691ssclength(bp)16553irlength(bp)27635genenumber132protein-codinggenenumber85protein-codinggene(%)64.39rrnagenenumber8rrna(%)6.06trnagenenumber37trna(%)28.03genenumberinirregions365、古树ad分子标记5.1古树ad特异性位点比对通过毛白杨具体叶绿体基因组序列比对(图12),特异性位点位置为:289位点,其他毛白杨该位点为碱基t,ad该位点为空缺,设计引物ad-2f和ad-2r进行pcr扩增,扩增体系同上,结果显示与seq id no.15所示序列的45~568bp一致,位点准确,可进行标记。
39.5.2古树ad特异性位点标记标记位点位置:289(空缺t),无可用酶切位点。
40.因此,目前选择特异性引物标记和叶绿体基因组全长序列标记。相关的酶标法仍旧处于设计过程中。
41.1)特异性引物序列标记法针对毛白杨古树ad特异性位点位置289位点,特异性:其他毛白杨该位点为碱基t,ad该位点为空缺,即少一个t,设计特异性引物ad-2f和ad-2r进行特异性位点所属区域片段序列标记,片段长度:524bp(图13),结果显示位点准确,可进行标记。
42.2)ad种源全长序列标记法毛白杨古树ad叶绿体基因组全长序列156513 bp,全长包含了36.72%的gc,63.28%的at,lsc长度为84692 bp,ssc长度为16551 bp,ir长度为27635,所含基因数目为132,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表4)。
43.6、古树lf分子标记6.1古树lf特异性位点比对通过毛白杨具体叶绿体基因组序列比对,特异性位点位置为:81838位点(图14),
其他毛白杨该位点为碱基a,lf该位点为空缺,设计引物lf-2f和lf-2r进行特异性位点验证,扩增体系同上,结果显示与seq id no.16所示序列的185~1066bp序列匹配度99.9%,特异性位点存在,可进行标记。
44.6.2古树lf特异性位点标记标记位点位置:81838(空缺t),该位点无可用酶切位点(图15)。
45.1)特异性引物序列标记法针对毛白杨古树lf特异性位点位置81838 (空缺)位点,其他毛白杨该位点为碱基a,lf位点为空缺,利用特异性引物lf-2f和lf-2r进行特异性位点所属区域片段序列标记,片段长度:881bp,结果显示位点准确,可进行标记。
46.2)lf种源全长序列标记法毛白杨古树lf叶绿体基因组全长序列156511 bp,全长包含了36.72%的gc,63.28%的at,lsc长度为84690 bp,ssc长度为16551 bp,ir长度为27635,所含基因数目为132,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表7)。
47.表7 populus tomentosa lf叶绿体基因组基本特征统计结果length(bp)156511gccontent(%)36.72atcontent(%)63.28lsclength(bp)84690ssclength(bp)16551irlength(bp)27635genenumber132protein-codinggenenumber85protein-codinggene(%)64.39rrnagenenumber8rrna(%)6.06trnagenenumber37trna(%)28.03genenumberinirregions367、古树xf分子标记7.1古树xf特异性位点比对通过毛白杨具体叶绿体基因组序列比对,特异性位点位置为:64826位点(图16),其他毛白杨该位点为碱基c,xf该位点为碱基t,设计引物xf-1f和xf-1r进行特异性位点验证,扩增体系同上,结果显示扩增产物为seq id no.17所示序列的84~823bp,序列匹配度100%,特异性位点存在,可进行标记。
48.7.2古树xf特异性位点标记1)酶切位点标记法标记位点位置:64826 (t)
ꢀꢀ
完成标记acatgt位点标记,酶切位点(图17):pcii(acatgt/tgtaca);afliii(acrygt/tgyrca),与原有序列检测,结果显示酶切片段与上述扩增产物一致,两个酶切位点均可用,可有效切开片段。
49.2)特异性引物序列标记法针对毛白杨古树xf特异性位点,利用引物xf-1f和xf-1r进行特异性位点所属区域片段序列标记,片段长度:740bp,序列与上述扩增产物一致,结果显示位点准确,可进行标记。
50.3)全长序列标记法毛白杨古树xf叶绿体基因组全长序列156514 bp,全长包含了36.72%的gc,63.28%的at,lsc长度为84693 bp,ssc长度为16551 bp,ir长度为27635,所含基因数目为132,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表8)。
51.表8 populus tomentosa xf叶绿体基因组基本特征统计结果length(bp)156514gccontent(%)36.72atcontent(%)63.28lsclength(bp)84693ssclength(bp)16551irlength(bp)27635genenumber132protein-codinggenenumber85protein-codinggene(%)64.39rrnagenenumber8rrna(%)6.06trnagenenumber37trna(%)28.03genenumberinirregions368、古树dbyy分子标记8.1古树dbyy特异性位点比对通过毛白杨具体叶绿体基因组序列比对,特异性位点位置为:7309-7314(图18),其他毛白杨该位点为空缺,dbyy该位点为碱基片段attttt,设计引物dbyy-1f和dbyy-1r进行特异性位点验证,扩增体系同上,结果显示与seq id no.18所示序列的24~729bp一致,位点准确,可进行标记。
52.8.2古树dbyy特异性位点标记标记位点位置:7309-7314 (attttt),无可用酶切位点(图19)。
53.1)特异性引物序列标记法针对毛白杨古树dbyy特异性位点位置,利用引物dbyy-1f和dbyy-1r进行特异性位点所属区域片段序列标记,片段长度:901bp,结果显示位点准确,可进行标记。
54.2)全长序列标记法毛白杨古树dbyy叶绿体基因组全长序列156497 bp,全长包含了36.71%的gc,63.28%的at,lsc长度为84676 bp,ssc长度为16551 bp,ir长度为27635,所含基因数目为130,编码蛋白基因数目85,rrna基因数目8,trna基因数目37(表3)。
55.以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人
员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。