具有用于模板多核苷酸附接的单个位点的纳米颗粒
1.相关申请的交叉引用
2.本技术是根据专利合作条约提交的国际申请,其要求2019年12月23日提交的美国临时专利申请号62/952,799和2019年12月23日提交的美国临时专利申请号62/952,866的优先权权益,这些临时专利申请的全部内容以引用方式并入本文。
3.序列表
4.本技术包含2020年12月16日创建的序列表;该文件为ascii格式,被命名为h1912656.txt,大小为6.8kb。该文件据此全文以引用方式并入本技术中。
背景技术:
5.许多当前测序平台使用“边合成边测序”(sbs)技术和基于荧光的检测方法。在一些示例中,在称为接种的过程中,将从文库分离的待测序的许多靶多核苷酸或者模板多核苷酸附接到基底的表面。然后,在称为聚类的过程中,可合成模板多核苷酸的多个拷贝,这些拷贝附接到表面上靠近作为拷贝的模板多核苷酸接种的位置。随后,在以下条件下合成聚类多核苷酸的新生拷贝:当每个核苷酸附接到新生链时,聚类多核苷酸的新生拷贝发射识别每个核苷酸的信号。接种的模板多核苷酸的多个拷贝靠近其最初接种的位置进行聚类使得在可视化聚合期间产生的信号扩增,从而改善检测。
6.当模板多核苷酸接种尽可能多的可用基底表面时,sbs的接种和聚类效果很好,这可最大化可在测序运行期间获得的测序信息量。相比之下,一般来说,用于接种和聚类的基底的可用表面积越小,sbs过程的效率可能越低,从而导致时间、反应物、费用增加并且获得给定文库的给定量的测序信息的数据处理变得复杂。
7.当来自文库的具有彼此不同的序列的模板多核苷酸以彼此相距足够远的距离接种或附接到表面的各个位置,使得聚类产生各自来源于单个模板多核苷酸的接种的复制多核苷酸的在空间上不同的簇(这种情况通常称为单克隆性)时,接种和聚类效果也很好。也就是说,模板多核苷酸的文库通常可包括大量的模板多核苷酸分子,这些模板多核苷酸分子的核苷酸序列彼此不同。如果两个这样的模板多核苷酸在基底的表面上接种得太近,则聚类可能会产生复制多核苷酸的在空间上混合的群体,其中一些复制多核苷酸具有接种在附近的模板多核苷酸中的一个的序列,而其他复制多核苷酸具有在表面上也接种在附近的另一个模板多核苷酸的序列。或者,由彼此靠太近接种的两个不同模板多核苷酸形成的两个簇可能彼此太相邻或彼此邻接,使得在sbs过程中使用的成像系统可能无法将它们区分为单独的簇,即使这些簇之间可能没有或极少有基底附接的序列的空间混合。这种不利条件通常被称为多克隆性。从多克隆簇(如果存在)中获取明确的序列信息可能更困难、更耗时、更昂贵且效率更低,并且需要更复杂的数据分析。
技术实现要素:
8.因此,期望在以下条件下进行sbs:基底表面的尽可能多的可用表面积用于接种和聚类,同时还促进接种的模板多核苷酸的分离,以便尽可能最大化簇的单克隆性,并且尽可
能最小化多克隆簇。本文公开了可用于有利地增加sbs中的接种密度和单克隆聚类的组合物和方法。
9.在一个方面,提供了一种纳米颗粒,其包括支架、用于将模板多核苷酸结合到支架的单个模板位点以及用于将辅助寡核苷酸结合到支架的多个辅助位点,其中支架选自一个或多个支架dna分子和一个或多个支架多肽,用于将模板多核苷酸结合到支架的单个模板位点选自共价模板结合位点和非共价模板结合位点,并且用于将辅助寡核苷酸结合到支架的多个辅助位点选自共价辅助寡核苷酸结合位点和非共价辅助寡核苷酸结合位点。
10.在一个示例中,支架包括一个或多个支架dna分子。在另一个示例中,支架包括多个支架dna分子,其中多个支架dna分子包含dna树枝状大分子。在又一个示例中,dna树枝状大分子包括多代分叉组成重复单元,其中代数为2至100。在再一个示例中,分叉组成重复单元各自包括三个组成重复单元寡聚脱氧核苷酸,三个组成重复单元寡聚脱氧核苷酸彼此杂交而形成包括一个上游突出端和两个下游突出端的衔接子,其中第2代及更高代中每个衔接子的上游突出端与紧邻的上游组成重复单元的下游突出端互补,并且第1代中衔接子的下游突出端包括单个模板位点。在另一个示例中,支架包括单链dna。
11.在另一个示例中,支架包括一个或多个支架多肽。在另一个示例中,支架多肽包括绿色荧光蛋白。
12.在另一个示例中,单个模板位点包括共价模板结合位点。在又一个示例中,共价模板结合位点选自胺-nhs酯结合位点、胺-亚氨酸酯结合位点、胺-戊氟苯基酯结合位点、胺-羟甲基膦结合位点、羧基-碳化二亚胺结合位点、硫醇-马来酰亚胺结合位点、硫醇-卤代乙酰基结合位点、硫醇-吡啶基二硫化物结合位点、硫醇-硫代磺酸酯结合位点、硫醇-乙烯基砜结合位点、醛-酰肼结合位点、醛-烷氧基胺结合位点、羟基-异氰酸酯结合位点、叠氮化物-炔烃结合位点、叠氮化物-膦结合位点、反式环辛烯-四嗪结合位点、降冰片烯-四嗪结合位点、叠氮化物-环辛炔结合位点、叠氮化物-降冰片烯结合位点、肟结合位点、spytag-spycatcher结合位点、snap-标签-o
6-苄基鸟嘌呤结合位点、clip-标签-o2-苄基胞嘧啶结合位点和分选酶偶联结合位点。
13.在另一个示例中,单个模板位点包括非共价模板结合位点。在又一个示例中,非共价模板结合位点包括多核苷酸杂交位点。在又一个示例中,非共价模板结合位点包括非共价肽结合位点,并且非共价肽结合位点选自卷曲螺旋结合位点和抗生物素蛋白-生物素结合位点。
14.在另一个示例中,用于将辅助寡核苷酸结合到支架的多个辅助位点包括共价辅助寡核苷酸结合位点。在又一个示例中,共价辅助寡核苷酸结合位点选自胺-nhs酯结合位点、胺-亚氨酸酯结合位点、胺-戊氟苯基酯结合位点、胺-羟甲基膦结合位点、羧基-碳化二亚胺结合位点、硫醇-马来酰亚胺结合位点、硫醇-卤代乙酰基结合位点、硫醇-吡啶基二硫化物结合位点、硫醇-硫代磺酸酯结合位点、硫醇-乙烯基砜结合位点、醛-酰肼结合位点、醛-烷氧基胺结合位点、羟基-异氰酸酯结合位点、叠氮化物-炔烃结合位点、叠氮化物-膦结合位点、反式环辛烯-四嗪结合位点、降冰片烯-四嗪结合位点、叠氮化物-环辛炔结合位点、叠氮化物-降冰片烯结合位点、肟结合位点、spytag-spycatcher结合位点、snap-标签-o
6-苄基鸟嘌呤结合位点、clip-标签-o2-苄基胞嘧啶结合位点、分选酶偶联结合位点以及上述中两种或多种的任意组合。
15.在另一个示例中,辅助寡核苷酸结合位点包括非共价辅助寡核苷酸结合位点。在又一个示例中,非共价辅助寡核苷酸结合位点包括多核苷酸杂交位点。在再一个示例中,非共价辅助寡核苷酸结合位点包括非共价肽结合位点,并且非共价肽结合位点选自卷曲螺旋结合位点和抗生物素蛋白-生物素结合位点中的一者或两者。
16.在另一个示例中,纳米颗粒还包括与单个模板位点结合的单个模板多核苷酸。在又一个示例中,纳米颗粒还包括与多个辅助位点结合的多个辅助寡核苷酸。
17.在另一个示例中,纳米颗粒的直径为至少约10nm、直径为至少约20nm、直径为至少约30nm、直径为至少约40nm、直径为至少约50nm、直径为至少约60nm、直径为至少约70nm、直径为至少约80nm、直径为至少约90nm、直径为至少约100nm、直径为至少约125nm、直径为至少约150nm、直径为至少约175nm、直径为至少约200nm、直径为至少约225nm、直径为至少约250nm、直径为至少约275nm、直径为至少约300nm、直径为至少约325nm、直径为至少约350nm、直径为至少约375nm、直径为至少约400nm、直径为至少约425nm、直径为至少约450nm、直径为至少约475nm、直径为至少约500nm、直径为至少约550nm、直径为至少约600nm、直径为至少约650nm、直径为至少约700nm、直径为至少约750nm、直径为至少约800nm、直径为至少约850nm、直径为至少约900nm或直径为至少约950nm。
18.在另一方面,提供了一种方法,其包括将单个模板多核苷酸结合到纳米颗粒的单个模板位点。
19.在另一方面,提供了一种方法,其包括将多个辅助寡核苷酸结合到纳米颗粒的多个辅助位点。
20.在另一方面,提供了一种方法,其包括将单个模板多核苷酸结合到纳米颗粒的单个模板位点以及将多个辅助寡核苷酸结合到纳米颗粒的多个辅助位点中的至少一项,其还包括合成选自模板多核苷酸的拷贝、与模板多核苷酸互补的多核苷酸的拷贝以及这两者的拷贝的一个或多个支架附接的拷贝,其中支架附接的拷贝从辅助寡核苷酸延伸。
21.在另一个示例中,该方法还包括将支架附接到基底,其中附接包括使辅助寡核苷酸与附接到基底的寡核苷酸杂交。
22.在该方法的又一个示例中,基底包括多个纳米孔,并且附接到基底的寡核苷酸附接在多个纳米孔内。在又一个示例中,不超过一个支架结合在纳米孔中的任一个内。在再一个示例中,该方法还包括合成选自模板多核苷酸的拷贝、与模板多核苷酸互补的多核苷酸的拷贝以及这两者的拷贝的一个或多个基底附接的拷贝,其中基底附接的拷贝从附接到基底的寡核苷酸延伸。在再一个示例中,该方法还包括对支架附接的拷贝和基底附接的拷贝中的至少一者进行测序,其中测序包括边合成边测序。
附图说明
23.当参考附图阅读以下详细描述时将更好地理解本公开的这些和其他特征、方面和优点,其中:
24.图l示出了根据本公开的各方面的纳米颗粒的示例。
25.图2a-图2f示出了根据本公开的各方面的包括dna树枝状大分子的纳米颗粒的支架的示例的各部分。
26.图3示出了根据本公开的各方面的包括单链dna支架的纳米颗粒的示例。
27.图4示出了根据本公开的各方面的纳米颗粒的多肽支架的示例。
28.图5示出了根据本公开的各方面的模板附接到纳米颗粒的示例。
29.图6示出了根据本公开的各方面的合成模板多核苷酸的支架附接的拷贝的示例。
30.图7示出了根据本公开的各方面的共价附接多核苷酸的示例。
31.图8示出了根据本公开的各方面的共价附接到支架的氨基酸的示例。
32.图9示出了根据本公开的各方面的通过杂交将模板多核苷酸非共价附接到纳米颗粒的示例。
33.图l0示出了根据本公开的各方面的通过卷曲螺旋肽结合位点将模板多核苷酸非共价附接到纳米颗粒的示例。
34.图l1示出了根据本公开的各方面的用于共价附接的纳米颗粒的支架的多个辅助位点的示例。
35.图l2示出了根据本公开的各方面的纳米孔中的纳米颗粒的示例。
36.图l3是根据本发明的各方面的示出根据纳米孔表面积的每个纳米孔的纳米颗粒的数量的图。
37.图14a-图14d示出了根据本公开的各方面的使用dna支架用模板多核苷酸接种基底的示例。
具体实施方式
38.本说明书通篇提及的“一个示例”、“另一个示例”、“一种示例”等意指结合该示例描述的特定元素(例如,特征、结构和/或特性)包括在本文所述的至少一个示例中,并且可存在于或不存在于其他示例中。此外,应当理解,用于任何示例的所述元素可以任何合适的方式组合在各种示例中,除非上下文另有明确说明。
39.本公开涉及用于在sbs期间增加单克隆聚类的组合物和方法。在一个示例中,尺寸排除原理用于防止单独的模板多核苷酸接种,因此防止促进彼此太近聚类。通过将每个单独的模板多核苷酸与给定的足够空间维度的纳米颗粒相关联,可诱导模板多核苷酸以彼此相距足够远的距离附接到基底表面,以减少多克隆簇的形成并增加单克隆簇的形成。纳米颗粒可包括模板多核苷酸的结合位点。纳米颗粒可具有仅一个用于附接模板多核苷酸的单个位点。因此,一个且仅一个模板多核苷酸可以能够附接到纳米颗粒,使得模板多核苷酸附接到支架防止第二模板多核苷酸附接到同一纳米颗粒,所附接的模板多核苷酸已占据其单个模板多核苷酸结合位点。每个纳米颗粒仅附接单个模板多核苷酸,并且直接或间接由于所附接的纳米颗粒的尺寸产生彼此附接到此类颗粒的模板多核苷酸的空间分布减少了多克隆簇的形成。
40.除了模板多核苷酸(在本文中称为辅助结合位点)之外,纳米颗粒还可包括其他类型的用于将纳米颗粒附接到组合物或表面的一个或多个结合位点。例如,除了单个模板多核苷酸结合位点之外,纳米颗粒还可包括允许纳米颗粒附接到用于sbs过程中的基底表面的辅助结合位点。在另一个示例中,纳米颗粒可具有用于将一种或多种表面聚合物附接到纳米颗粒的一个或多个辅助结合位点。在另一个示例中,纳米颗粒可包括用于将辅助寡核苷酸附接到纳米颗粒的一个或多个辅助结合位点,其中作为聚类过程的一部分,寡核苷酸可与模板多核苷酸或其拷贝的末端结合,如下文更充分地描述。在另一个示例中,此类辅助
寡核苷酸可与附接到用于sbs过程中的基底表面的寡核苷酸杂交,使得其上附接有单个模板多核苷酸的纳米颗粒可附接到此类基底表面。
41.鉴于支架可包括用于模板多核苷酸的单个结合位点和用于附接例如辅助寡核苷酸的一个或多个辅助位点,单个模板多核苷酸结合位点可具有与辅助结合位点不同的化学或结构。在所有结合位点中,单个模板多核苷酸结合位点可以是具有以下化学和结构的唯一一个模板多核苷酸结合位点:被设计用于附接到具有用于与其附接的对应化学或结构的模板多核苷酸。相比之下,一个或多个辅助结合位点可具有不同的化学或结构,其与结合或附接到模板多核苷酸不相容。相反,一个或多个辅助结合位点可具有与结合或附接到辅助结合位点旨在结合的其他组合物或结构(诸如辅助寡核苷酸、聚合物等)相容并且与结合或附接到模板多核苷酸不相容的化学或结构。因此,模板多核苷酸将无法结合或附接到一个或多个辅助结合位点,从而导致在纳米颗粒的单个模板多核苷酸结合位点处每个纳米颗粒仅附接一个模板多核苷酸。
42.模板多核苷酸可以是从样品中获得的多核苷酸,诸如从样品中分离的聚脱氧核糖核酸或由从样品中获得的mrna分子复制的cdna分子。例如,可执行sbs过程以确定模板多核苷酸的核苷酸序列,或与参考序列相比,鉴定模板多核苷酸的遗传序列中的一个或多个多态性或改变。可由一个或多个样品制备文库,该文库包括从一个或多个样品中获得的多个模板多核苷酸。可通过获得作为存在于样品中或由样品复制的序列的部分的多核苷酸序列来获得模板多核苷酸。通过在sbs过程中对多个模板多核苷酸进行测序,可确定关于模板多核苷酸的序列、基因型或其他序列相关信息,并且当收集和分析关于文库中的多个模板多核苷酸的序列信息时,可确定关于获得文库的样品的序列、基因型或其他序列相关信息。
43.可将模板多核苷酸作为从样品中获得模板多核苷酸的过程的一部分来处理。处理部分可包括将多核苷酸序列添加到诸如模板的5-末端、3-末端或两端以帮助进行亚序列sbs处理。如本文进一步公开的,模板多核苷酸可通过添加促进或允许与纳米颗粒上的位点形成键的特征来进一步修饰。
44.模板多核苷酸可具有适于在sbs过程中获得测序信息的任何给定长度。例如,模板多核苷酸的长度可为约50个核苷酸、长度为约75个核苷酸、长度为约100个核苷酸、长度为约125个核苷酸、长度为约150个核苷酸、长度为约175个核苷酸、长度为约200个核苷酸、长度为约225个核苷酸、长度为约250个核苷酸、长度为约275个核苷酸、长度为约300个核苷酸、长度为约325个核苷酸、长度为约350个核苷酸、长度为约375个核苷酸、长度为约400个核苷酸、长度为约425个核苷酸、长度为约450个核苷酸、长度为约475个核苷酸、长度为约500个核苷酸、长度为约525个核苷酸、长度为约550个核苷酸、长度为约575个核苷酸、长度为约600个核苷酸、长度为约625个核苷酸、长度为约650个核苷酸、长度为约675个核苷酸、长度为约700个核苷酸、长度为约725个核苷酸、长度为约750个核苷酸、长度为约775个核苷酸、长度为约800个核苷酸、长度为约825个核苷酸、长度为约850个核苷酸、长度为约875个核苷酸、长度为约900个核苷酸、长度为约925个核苷酸、长度为约950个核苷酸、长度为约975个核苷酸、长度为约1,000个核苷酸、长度为约1,025个核苷酸、长度为约1,050个核苷酸、长度为约1,075个核苷酸、长度为约1,100个核苷酸、长度为约1,125个核苷酸、长度为约1,150个核苷酸、长度为约1,175个核苷酸、长度为约1,200个核苷酸、长度为约1,225个核苷酸、长度为约1,250个核苷酸、长度为约1,275个核苷酸、长度为约1,300个核苷酸、长度为约
1,325个核苷酸、长度为约1,350个核苷酸、长度为约1,375个核苷酸、长度为约1,400个核苷酸、长度为约1,425个核苷酸、长度为约1,450个核苷酸、长度为约1,475个核苷酸、长度为约1,500个核苷酸或更长。
45.在一些示例中,在纳米颗粒上可能存在两个或更多个不同的辅助结合位点群体,一些具有与结合或附接到一个组合物或结构群体相容的一种类型的化学或结构,而其他具有与结合或附接到另一个组合物或结构群体相容的第二种类型的化学或结构。例如,一个辅助位点群体可具有与结合到辅助寡核苷酸(这些辅助寡核苷酸可与参与例如模板多核苷酸在纳米颗粒上聚类的模板多核苷酸的拷贝结合,如下文更充分地描述)相容的化学或结构,而其他辅助位点可具有与结合或附接到基底的表面以执行sbs相容的不同化学或结构。
46.纳米颗粒可包括支架。支架是纳米颗粒的根据模板纳米颗粒之间期望的最小距离量或附接到纳米颗粒的模板纳米颗粒的最大密度(如对于给定应用可能期望的)占据体积的结构组件。支架可包括前述结合位点,如单个模板多核苷酸结合位点和一个或多个辅助结合位点。支架与结合位点一起可构成纳米颗粒。可合成支架以包括并且其一旦合成就可包括多于一种类型的用于附接的化学或结构。也就是说,可合成支架以包括或将其修饰成包括附接到模板多核苷酸的单个位点,加上与辅助结合位点对应的具有与单个模板多核苷酸结合位点不同的化学或结构的一个或多个附加结合位点。
47.支架可由若干不同的取代组分合成。在一个示例中,支架可由一个或多个支架脱氧核糖核酸(dna)分子合成。可如本文进一步公开那样设计和结构化dna分子,以便允许包括不同的结合位点(即,对于模板多核苷酸以及辅助结合位点),并且还提供一旦附接到多核苷酸就使模板多核苷酸彼此远离的尺寸排阻特性。在一些示例中,支架可包括杂交在一起以形成树枝状大分子的多个dna分子。例如,衔接子可被形成为包括多个(诸如三个)dna链或寡聚脱氧核苷酸(寡核苷酸-dna)分子,这些分子可以通过沃森-克里克碱基配对彼此杂交而形成y形,其中每个y形的一个末端与另外两个末端中的一个杂交,并且每个y形的另一个末端与另外两个末端中的另一个杂交。
48.此类衔接子可形成树枝状大分子的组成重复单元。例如,y形衔接子的每个末端可具有dna的突出端,其中寡核苷酸-dna中的一个的末端延伸超出其与任何其他寡核苷酸-dna杂交的部分。此类树枝状大分子的一代衔接子可在y形的一端上具有突出端(此处称为上游末端),该突出端可以与构成紧邻的前一代树枝状大分子的组成重复单元的另一个y衔接子的末端的突出端杂交。并且衔接子的另连个末端(称为下游末端)可各自具有突出端,该突出端可以与构成紧邻的后续代树枝状大分子的组成重复单元的y衔接子的上游末端的突出端杂交。因此,一代的衔接子可附接到下一代的两个衔接子,这两个衔接子可附接到后续代的四个衔接子,后续代的四个衔接子可附接到后续代的八个衔接子,以此类推。末端y衔接子中的一个的任何一个末端(无论是任一代的下游突出端(诸如最后一代,其不与另一个衔接子的上游突出端杂交)还是第一代的上游突出端)可包括或附接到单个模板多核苷酸结合位点。在一个示例中,包括第一代衔接子的上游突出端的dna寡核苷酸本身可以是模板多核苷酸的在样品制备期间添加到其上的延伸部。其他末端或突出端可包括或附接到辅助位点。
49.在其他示例中,支架可包括一个或多个单链dna(ssdna)分子,这些单链dna分子被修饰或结构化成允许单个模板多核苷酸和一种或多种辅助组合物或结构附接到一个或多
个辅助结合位点。可使用用于产生基于ssdna的支架的各种方法。在一个示例中,在滚环扩增过程中,双链闭环或质粒可用作ssdna支架分子的编码序列。通过链置换dna聚合酶(例如,phi29)复制其链可产生ssdna分子,该分子包括环状编码链的复制链的串联拷贝。可采用反应条件以合成所需大小的ssdna支架。5-末端或3-末端可被进一步修饰以作为单个模板位点包括或附接到单个模板多核苷酸分子。辅助位点可包括ssdna支架分子的另一个末端,或者对链的单独核苷酸的修饰,如下文进一步描述。
50.在另一个示例中,可通过使用模板独立的聚合酶(例如,末端脱氧核苷酸转移酶或tdt)来合成ssdna支架。tdt在单链dna链的3-末端-羟基末端处掺入脱氧核苷酸,而不需要或复制模板。5-末端或3-末端可被进一步修饰以作为单个模板位点包括或附接到单个模板多核苷酸分子。辅助位点可包括ssdna支架分子的另一个末端,或者对链的单独核苷酸的修饰,如下文进一步描述。如本文所用,“核苷酸”包括含氮杂环碱基、糖以及一个或多个磷酸基团。核苷酸是核酸序列的单体单元。在rna中,糖是核糖,并且在dna中,糖是脱氧核糖,即在核糖中缺少存在于2'位置处的羟基基团的糖。含氮杂环碱基(即,核碱基)可为嘌呤碱基或嘧啶碱基。嘌呤碱基包括腺嘌呤(a)和鸟嘌呤(g)以及它们的修饰衍生物或类似物。嘧啶碱基包括胞嘧啶(c)、胸腺嘧啶(t)和尿嘧啶(u)以及它们的修饰衍生物或类似物。脱氧核糖的c-1原子与嘧啶的n-1或嘌呤的n-9结合。
51.在另一个示例中,可通过任何适用的方法产生多个单链dna分子并将它们连接在一起以形成单个ssdna分子作为支架来合成ssdna支架。例如,聚合酶可通过在流失聚合反应(即,聚合酶在达到编码链的5-末端时停止延伸新生链的情况)中复制线性化dna编码链来聚合dna的新生链的形成。可合成多种ssdna产物,然后将它们端到端连接以形成单个ssdna支架。在一个示例中,一种ssdna产物与另一种ssdna产物的连接可借助于夹板来实现。例如,可设计短寡核苷酸-dna,其3-末端与一种ssdna产物的5-末端互补,并且其5-末端与另一种ssdna产物的3-末端互补,使得dna-寡核苷酸与这两种ssdna产物的杂交将一种产物的5-末端与另一种产物的3-末端一起放在带切口的双链结构中,在该带切口的双链结构中它们与dna-寡核苷酸杂交。然后可使用dna连接酶(例如,t4)来将两端酶促连接在一起,以由这两种产物形成单个ssdna分子。dna-寡核苷酸可包括另外的反应,用于夹板辅助将这种第一反应的产物的一个或两个末端连接到另一种ssdna产物,诸如此类,以构建可能需要的ssdna支架。
52.用于附接到支架的模板多核苷酸可具有任何合适的长度,包括在sbs过程中进行测序。例如,模板多核苷酸的长度可为约50个核苷酸、长度为约75个核苷酸、长度为约100个核苷酸、长度为约125个核苷酸、长度为约150个核苷酸、长度为约175个核苷酸、长度为约200个核苷酸、长度为约225个核苷酸、长度为约250个核苷酸、长度为约275个核苷酸、长度为约300个核苷酸、长度为约325个核苷酸、长度为约350个核苷酸、长度为约375个核苷酸、长度为约400个核苷酸、长度为约425个核苷酸、长度为约450个核苷酸、长度为约475个核苷酸、长度为约500个核苷酸、长度为约525个核苷酸、长度为约550个核苷酸、长度为约575个核苷酸、长度为约600个核苷酸、长度为约625个核苷酸、长度为约650个核苷酸、长度为约675个核苷酸、长度为约700个核苷酸、长度为约725个核苷酸、长度为约750个核苷酸、长度为约775个核苷酸、长度为约800个核苷酸、长度为约825个核苷酸、长度为约850个核苷酸、长度为约875个核苷酸、长度为约900个核苷酸、长度为约925个核苷酸、长度为约950个核苷
酸、长度为约975个核苷酸、长度为约1,000个核苷酸、长度为约1,100个核苷酸、长度为约1,200个核苷酸、长度为约1,300个核苷酸、长度为约1,40个核苷酸、长度为约1,500个核苷酸、长度为约1,600个核苷酸、长度为约1,700个核苷酸、长度为约1,800个核苷酸、长度为约1,900个核苷酸、长度为约2000个核苷酸或更长。
53.单个模板多核苷酸或辅助物(例如,辅助寡核苷酸、辅助组合物或辅助结构)与dna支架的附接可通过在支架和模板多核苷酸或辅助物上包括彼此互补的部分或结构来实现,这意味着它们被构造成彼此共价或非共价结合以在其间形成附接。它们可互补用于共价结合或互补用于非共价结合。dna支架可包括具有与附接到模板多核苷酸的部分或结构(单个模板位点)互补的单个模板位点。dna支架还可包括或附接到与附接到辅助物的部分或结构(辅助位点)互补的其他部分或结构。应当避免附接到模板多核苷酸的部分或结构与辅助位点的部分或结构之间的交叉反应性,以防止多于一个模板多核苷酸附接到dna支架。还应当避免连接到辅助物的部分或结构与单个模板位点的部分或结构之间的交叉反应性,以防止通过防止单个模板多核苷酸与其连接的辅助物对单个模板位点进行占用。在一些示例中,可通过分别在辅助物结合到辅助位点或单个模板多聚核苷酸附接到单个模板位点时,以化学方式阻断单个模板位点或辅助位点,然后不阻断未占用位点以允许单个模板多核苷酸或辅助物附接到其上,来避免此类交叉反应性。
54.表1中呈现了互补结合配偶体的非排他性列表:
55.56.57.[0058][0059]
可以将前述中的任何一种添加到或包括在如本文所公开的支架中以用于附接到模板多核苷酸或辅助物(诸如辅助寡核苷酸),该模板多核苷酸或辅助物可包括或被修饰成包括前述对的互补部分或结构以用于与支架结合。
[0060]
可使用用于在此类互补部分或结构对之间添加或形成键的任何合适的生物缀合方法。经修饰的核苷酸可商购获得,其具有此类互补部分或结构对的示例中的一个或其他的示例,并且用于包括此类部分或结构示例中的一个或多个或者将它们附接或包括到聚合物、核苷酸或多核苷酸的方法也是已知的。还可商购获得的是双功能接头分子,其具有来自表1中列出的一对互补结合配偶体的部分或结构以及来自表1中列出的另一对互补结合配偶体的部分或结构。支架、模板多核苷酸或者辅助物或寡核苷酸或多肽的部分或结构(附接到前述特征中的任一个以便提供用于在此类前述特征中的任一个之间结合的部分或结构)可与这种接头的一端结合,从而导致初始部分或结构被另一个部分或结构(即存在于接头另一端的部分或结构)有效替换。
[0061]
例如,双功能接头可在一端具有来自表1中列出的那些部分的部分,诸如nhs-酯基团。其可在另一端具有另一个基团,诸如叠氮化物基团。末端可通过接头(诸如连接序列中的一个或多个peg基团、烷基链、它们的组合等)彼此连接。如果结合位点(诸如支架或者模板多核苷酸或辅助物的结合位点)具有用于结合的胺基团,则双功能接头的nhs-酯末端可以与胺基团结合,从而使游离叠氮化物末端可与携带叠氮化物基团的结合配偶体(例如,炔烃、膦、环辛炔或降冰片烯)的组合物(例如,模板多核苷酸或辅助物或支架)结合。或者,如果结合位点(诸如支架或者模板多核苷酸或辅助物的结合位点)具有叠氮化物基团的结合配偶体(例如,炔烃、膦、环辛炔或降冰片烯),则双功能接头的叠氮化物末端可与胺基团结合,从而使游离nhs-酯末端可用于与携带胺基团的组合物(例如,模板多核苷酸或辅助物或
支架)结合。双功能接头的许多其他示例可商购获得,其在一端包括在表1中鉴定的用于形成一种类型的结合位点的部分,并且在另一端包括在表1中鉴定的用于形成另一类型的结合位点的不同部分。
[0062]
经修饰的氨基酸可商购获得,其具有此类互补部分或结构对的示例中的一个或其他的示例,并且用于包括此类部分或结构示例中的一个或多个或者将它们附接到氨基酸或多肽的方法也是已知的。用于在此类互补部分或结构对的成员之间形成键的方法是已知的。因此,可以将此类互补部分或结构添加到或包括在支架和模板多核苷酸或支架和辅助物中,以形成结合位点并允许在它们之间进行附接。
[0063]
在一个示例中,支架的单个模板多核苷酸结合位点可包括来自表1的第一部分或结构,并且一个或多个辅助位点可包括来自表1的一个或多个其他部分或结构。在其中第一结合配偶体不会与其他一个或多个结合配偶体反应以形成结合位点并且其他部分或结构不会与第一结合配偶体反应以形成结合位点的条件下,第一部分或结构可以能够与第一结合配偶体形成结合位点,并且其他部分或结构可以能够与其他一个或多个结合配偶体形成结合位点。在另一个示例中,选择第一部分或结构以及其他部分或结构,使得它们不会彼此形成结合配偶体。
[0064]
如本文所用,术语“多肽”旨在表示通过肽键结合在一起的氨基酸的链。术语“蛋白质”和“多肽”可互换使用。多肽可包括通过肽键彼此结合的多个氨基酸的序列,并且氨基酸的数量可为约2或更多、约5或更多、约10或更多、约15或更多、约20或更多、约25或更多、约30或更多、约35或更多、约40或更多、约45或更多、约50或更多、约55或更多、约60或更多、约65或更多、约70或更多、约75或更多、约80或更多、约85或更多、约90或更多、约95或更多、约100或更多、约110或更多、约120或更多、约130或更多、约140或更多、约150或更多、约160或更多、约170或更多、约180或更多、约190或更多、约200或更多、约225或更多、约250或更多、约275或更多、约300或更多、约325或更多、约350或更多、约375或更多、约400或更多、约425或更多、约450或更多、约475或更多、约500或更多、约550或更多、约600或更多、约650或更多、约700或更多、约750或更多、约800或更多、约850或更多、约900或更多、约950或更多、约1000或更高。
[0065]
在一些情况下,多肽或蛋白质可采用结构或三维构象以促进或允许与另一种结合配偶体(诸如也采用有利于这种结合的三维结构的另一种多肽,或其他非蛋白质结合配偶体)结合。多肽也可采用有利于对其他底物多肽或其他分子执行酶促反应或者用作另一种酶促反应或其他反应的底物的三维构象。多肽也可采用三维构象,使得一个或多个位点(诸如氨基末端、羧基末端、氨基酸的侧基或对氨基酸的修饰)可接近以与另一个分子结合。
[0066]
可以使用各种生物缀合化学将模板多核苷酸附接到dna支架的核苷酸或者附接到5-末端或3-末端(例如,包括在树枝状大分子dna支架的未杂交突出端或其他核苷酸中的核苷酸,或ssdna支架的3-末端或5-末端或它们之间的核苷酸)。此外,可实施对包括在dna支架中的核苷酸的修饰(诸如在磷酸基团、碱基或糖上),以提供用于附接的单个模板位点。可将化学部分包括在或添加到具有与互补化学部分形成共价缀合的能力的此类位点中,该互补部分可附接到或包括在模板多核苷酸中。然后可将模板多核苷酸缀合到dna支架,诸如通过互补化学部分之间的共价附接。在一个示例中,可在聚合反应期间包括经修饰以包括附接部分(能够通过聚合酶掺入多核苷酸链中,但也包括互补部分可与其反应以在它们之间
形成共价键的化学部分)的核苷酸,以形成dna支架或其部分。
[0067]
在另一个示例中,dna支架可包括或附接到作为单个模板位点的多肽序列,该多肽序列能够与另一个多肽序列或其他化学部分形成共价附接。然后,此类其他多肽或其他化学部分可包括在或附接到模板多核苷酸中,使得支架和模板多核苷酸的单个模板位点可彼此共价结合。另选地,模板多核苷酸可具有第一此类多肽序列,并且支架的单个模板位点可具有能够与模板多核苷酸的多肽序列共价结合的此类其他多肽序列或其他化学部分。此类对的非限制性示例包括spytag/spycatcher系统、snap-标签/o
6-苄基鸟嘌呤系统和clip-标签/o
2-苄基胞嘧啶系统。
[0068]
可获得互补spytag/spycatcher系统对和编码它们的多核苷酸的氨基酸序列。这些序列的示例在表1中提供。可获得spytag序列和spycatcher序列的若干氨基酸位点突变以包括在重组多肽中。snap标签是功能性o-6-甲基鸟嘌呤-dna甲基转移酶,并且clip标签是snap标签的修饰版本。可商购获得编码snap标签、clip标签、spycatcher的核苷酸序列以用于亚克隆和包括在工程化多肽序列中。
[0069]
另选地,用于在支架的单个模板位点和模板多核苷酸上进行共价附接的互补对可经由共价键的酶促催化形成而彼此共价附接。例如,支架的单个模板位点和模板多核苷酸可包括能够通过分选酶介导的偶联彼此共价附接的基序,例如在一者上为lpxtg氨基酸序列,而在另一者上为低聚甘氨酸亲核序列(具有例如3至5个甘氨酸的重复)。然后可进行分选酶介导的转肽作用,以使支架和模板多核苷酸在单个模板位点处共价附接。
[0070]
在另一个示例中,dna支架可包括用于单个模板多核苷酸在单个模板位点处非共价附接的区域。例如,树枝状大分子dna支架的未杂交突出端可通过沃森-克里克碱基配对与模板多核苷酸的末端杂交。在一个示例中,第一代树枝状大分子的衔接子的上游突出端可包括与模板多核苷酸的末端中包括的核苷酸序列互补的核苷酸序列。或者,ssdna支架的3-末端或5-末端可具有与模板多核苷酸的末端中包括的核苷酸序列互补的核苷酸序列。因此,此类互补核苷酸序列通过沃森-克里克碱基配对彼此杂交可允许dna支架的单个模板位点非共价附接到模板多核苷酸。
[0071]
在另一个示例中,dna支架和模板多核苷酸可包括或附接到互补肽结合位点。例如,dna支架和模板多核苷酸可包括或附接到可作为互补卷曲螺旋基序对彼此结合的肽序列。卷曲螺旋基序是一些多肽的结构特征,其中两个或更多个多肽链各自形成α-螺旋二级结构,并且α-螺旋卷曲在一起以形成紧密的非共价键。卷曲螺旋序列可包括七肽重复序列,即七个氨基酸hpphcpc的重复模式(其中h表示疏水性氨基酸,c通常表示带电氨基酸,并且p表示极性的亲水性氨基酸)。七肽重复序列的示例在亮氨酸拉链卷曲螺旋中发现,其中七肽的第四个氨基酸通常为亮氨酸。
[0072]
dna支架可包括或附接到形成卷曲螺旋结合对的一部分的一个氨基酸序列,并且模板多核苷酸可附接到形成卷曲螺旋结合对的另一部分的另一个氨基酸序列,该另一部分与作为或附接到dna支架的部分互补,使得这两个部分彼此附接。例如,dna支架可共价附接到形成卷曲螺旋结合对的一部分的一个氨基酸序列,并且模板多核苷酸可附接到形成卷曲螺旋结合对的另一部分的另一个氨基酸序列,该另一部分与作为或附接到dna支架的部分互补,使得这两个部分彼此附接。
[0073]
在另一个示例中,dna支架和模板多核苷酸可各自包括或附接到非共价结合在一
起的肽对的其他互补配偶体。示例包括生物素-抗生物素蛋白结合对。生物素和抗生物素蛋白肽(诸如抗生物素蛋白、链霉抗生物素蛋白和中性抗生物素蛋白,除非另外特别说明,否则所有这些均统称为“抗生物素蛋白”)彼此形成强非共价键。此类对的一个组分(无论是生物素还是抗生物素蛋白的结合部分)可以是dna支架或模板多核苷酸的一部分或者附接到dna支架或模板多核苷酸,其中互补组分相应地是dna支架或模板多核苷酸的一部分或者附接到dna支架或模板多核苷酸,从而允许在它们之间进行非共价附接。
[0074]
许多方法可用于包括一种或多种生物素部分或者将一种或多种生物素部分添加到dna分子、模板多核苷酸、dna支架、寡核苷酸-dna、多肽支架、其他多肽或如本文所公开的用于将分子结合在一起(诸如将模板多核苷酸结合到支架或将辅助物结合到支架)的其他组合物中。例如,可商购获得生物素化的核苷酸以用于通过聚合酶掺入dna分子中,并且可商购获得试剂盒以用于将生物素部分添加到多核苷酸或多肽中。也可以将生物素残基添加到氨基酸或修饰的氨基酸或者核苷酸或修饰的核苷酸中。表1中所示的连接化学还可以用于将生物素基团添加到蛋白质的诸如羧酸基团、胺基团或硫醇基团上。若干生物素连接酶也可用于酶促靶向生物素化,诸如多肽(例如,多肽中包括的avitag氨基酸序列glndifeaqkiewhe(seq id no:3)的赖氨酸残基)的酶促靶向生物素化。基因工程化的抗坏血酸盐过氧化物酶(apex)也可用于修饰生物素,以允许富含电子的氨基酸(诸如酪氨酸以及可能的色氨酸、半胱氨酸或组氨酸)的生物素化。
[0075]
在另一个示例中,包括氨基酸序列dslefiaskla(seq id no:4)的多肽可被生物素化(在序列中存在的两个s残基的更靠n末端),其是sfp磷酸泛酰巯基乙胺基转移酶催化的利用辅酶a(coa)缀合的小分子共价附接到其上的底物。例如,包括该序列的多肽可以通过利用coa-生物素缀合物共价附接到其上而被生物素化。该系统还可以用于附接表1中鉴定的许多其他类型的结合部分或结构,以用于形成如本文所公开的使支架结合到dna分子或多肽或其他分子的结合位点。例如,与表1中鉴定的反应对部分中的任一种缀合的coa可通过sfp磷酸泛酰巯基乙胺基转移酶共价附接到含有上述序列的多肽,从而允许包括互补结合配偶体的另一种组合物与其结合。
[0076]
其他酶可用于将结合部分添加到多肽。例如,硫辛酸连接酶可以添加硫辛酸分子或者包括表1中鉴定的结合部分(诸如炔烃或叠氮化物基团)的修饰的硫辛酸分子可以共价连接到多肽中包括的氨基酸序列devlveietdkavlevpggeee(seq id no:5)或gfeidkvwydlda(seq id no:6)中的赖氨酸残基的侧基的胺。在另一个示例中,其中包括的或旨在与其结合的支架、模板多核苷酸或其他多肽或dna分子可包括或附接到活性丝氨酸水解酶。氟代膦酸盐分子共价连接到丝氨酸水解酶的活性位点中的丝氨酸残基。包括表1中鉴定的结合部分(诸如叠氮化物基团或脱硫生物素基团)的氟代膦酸盐分子的可商购获得的类似物是可以与抗生物素蛋白结合的生物素的类似物。因此,此类基团可以共价附接到包括在或附接到如本文所公开的支架中所用或附接的多肽或dna分子中的丝氨酸水解酶,并且此类结合部分或结构可以通过附接合适的修饰的氟代膦酸盐分子来共价添加到其上,以在此类蛋白质上为来自表1的互补结合配偶体(诸如叠氮化物-炔烃、叠氮化物-膦、叠氮化物-环辛炔、叠氮化物-降冰片烯或脱硫生物素-抗生物素蛋白结合)产生结合位点。
[0077]
前述生物素化组合物(以促进与包括抗生物素蛋白序列的多肽(诸如包括在或附接到另一种组合物中的抗生物素蛋白多肽)结合或者以其他方式将官能团添加到多肽作为
支架的一部分、附接到支架、作为辅助物的一部分或附接到辅助物或模板多核苷酸,以用于在支架与模板多核苷酸之间或在支架与辅助物之间进行结合)方法中的任何一种可用于允许或促进在如本文所公开的此类组分之间进行结合。
[0078]
在另一个示例中,支架可由氨基酸(诸如多肽或蛋白质分子)合成。在一个示例中,用于附接模板多核苷酸的单个模板位点可以是或附接到此类多肽支架的n末端或c末端。在另一个示例中,用于附接模板多核苷酸的单个模板位点可以是或附接到多肽支架的内部氨基酸。各种生物缀合化学可以用于将模板多核苷酸附接到例如多肽的c末端与n末端之间的氨基酸的侧基,或附接到末端中的一个。此外,可实施对多肽支架的氨基酸(诸如对氨基酸中的一个的侧链)的修饰,以提供用于附接的单个模板位点。可将化学部分包括在或添加到具有与互补化学部分形成共价缀合的能力的此类位点中,该互补部分可附接到或包括在模板多核苷酸中。然后可将模板多核苷酸缀合到多肽支架,诸如通过互补化学部分之间的共价附接。
[0079]
在另一个示例中,多肽支架可包括或附接到作为单个模板位点的多肽序列,该多肽序列能够与另一个多肽序列或其他化学部分形成共价附接。然后,此类其他多肽或其他化学部分可包括在或附接到模板多核苷酸中,使得支架和模板多核苷酸的单个模板位点可彼此共价结合。另选地,模板多核苷酸可具有第一此类多肽序列,并且支架的单个模板位点可具有能够与模板多核苷酸的多肽序列共价结合的此类其他多肽序列或其他化学部分。此类对的非限制性示例包括spytag/spycatcher系统、snap-标签/o
6-苄基鸟嘌呤系统和clip-标签/o
2-苄基胞嘧啶系统。另选地,用于在支架的单个模板位点和模板多核苷酸上进行共价附接的互补对可经由共价键的酶促催化形成而彼此共价附接。例如,支架的单个模板位点和模板多核苷酸可包括能够通过分选酶介导的偶联彼此共价附接的基序,例如在一者上为lpxtg氨基酸序列,而在另一者上为低聚甘氨酸亲核序列(具有例如3至5个甘氨酸的重复)。然后可进行分选酶介导的转肽作用,以使支架和模板多核苷酸在单个模板位点处共价附接。
[0080]
在另一个示例中,多肽支架可包括用于单个模板多核苷酸在单个模板位点处非共价附接的区域。例如,寡核苷酸-dna可共价附接到多肽支架上的单个位点。例如,多肽支架和寡核苷酸-dna上的互补化学部分可允许在它们之间很像上文针对模板多核苷酸和多肽支架的直接共价附接那样进行共价附接。寡核苷酸-dna可具有与模板多核苷酸的一部分(诸如模板多核苷酸的3-末端或5-末端)互补的核苷酸序列。此类寡核苷酸-dna与模板多核苷酸之间的互补性可允许通过沃森-克里克碱基配对在模板寡核苷酸的该部分与附接到多肽支架的寡核苷酸-dna之间进行杂交。
[0081]
在另一个示例中,多肽支架和模板多核苷酸可包括或附接到互补肽结合位点。例如,肽支架和模板多核苷酸可包括或附接到可作为互补卷曲螺旋基序对彼此结合的肽序列。多肽支架可包括或附接到形成卷曲螺旋结合对的一部分的一个氨基酸序列,并且模板多核苷酸可附接到形成卷曲螺旋结合对的另一部分的另一个氨基酸序列,该另一部分与作为或附接到多肽支架的部分互补,使得这两个部分彼此附接。
[0082]
在另一个示例中,多肽支架和模板多核苷酸可各自包括或附接到非共价结合在一起的肽对的其他互补配偶体。示例包括生物素-抗生物素蛋白结合对。生物素和抗生物素蛋白肽彼此形成强非共价键。此类对的一个组分(无论是生物素还是抗生物素蛋白的结合部
分)可以是多肽支架或模板多核苷酸的一部分或者附接到多肽支架或模板多核苷酸,其中互补组分相应地是多肽支架或模板多核苷酸的一部分或者附接到多肽支架或模板多核苷酸,从而允许在它们之间进行非共价附接。
[0083]
为了附接到dna支架或多肽支架的单个模板位点,模板多核苷酸可具有添加到其上的互补附接部分或结构。在一个示例中,在制备文库样品期间,可制备多个模板多核苷酸以用于测序。通常,在此类样品制备期间,除了已经包括在其中作为待测序的文库的一部分的序列之外,文库样品的模板多核苷酸经修饰以包括特定核苷酸序列。此类添加的核苷酸序列可提供各种功能中的任一种,包括用于随后鉴定模板多核苷酸或作为接种过程的一部分附接到sbs基底的表面。根据本公开,模板多核苷酸的此类制备还可包括附接到其上或包括在其中的互补附接部分或结构。
[0084]
例如,对于树枝状大分子dna支架,模板多核苷酸的制备可包括在模板多核苷酸中添加或包括寡核苷酸,该寡核苷酸具有与第一代树枝状大分子的衔接子的上游末端的核苷酸序列对应的核苷酸序列。然后,第一代衔接子可包括构成其的三个多核苷酸序列中的一个,将此类序列添加到模板多核苷酸中。在另一个示例中,可将核苷酸序列添加到与树枝状大分子dna支架的衔接子的突出端(诸如树枝状大分子dna支架的第一代衔接子的上游突出端)互补的模板多核苷酸中。
[0085]
在另一个示例中,模板多核苷酸的制备可包括在模板多核苷酸中附接核苷酸序列,诸如从其末端中的一个延伸,并且该序列与其他序列包括在或附接到支架的单个模板位点中的另一个序列互补。由于沃森-克里克碱基配对引起的杂交使得在这两者之间进行结合。在另一个示例中,辅助物(诸如辅助寡核苷酸)可经修饰以允许与其互补的部分或结构共价附接到其上。例如,可包括对辅助物中包括的核苷酸(诸如辅助寡核苷酸)的修饰(诸如在磷酸基团、碱基或糖上)以提供与支架的辅助位点共价附接的位点。支架的辅助位点又可包括允许附接到辅助物(诸如寡核苷酸-dna辅助物)的互补部分或结构。在一个示例中,核苷酸经修饰以包括附接部分,在样品制备期间支架的辅助结合位点的互补部分通过该附接部分包括在添加到模板多核苷酸的多核苷酸序列中。可商购获得携带此类化学部分的许多修饰的核苷酸以用于将组合物共价附接到其中已掺入此类修饰的核苷酸的dna分子上。
[0086]
在另一个示例中,可诸如在样品制备期间通过附接到多肽来修饰模板多核苷酸。此类多肽可具有氨基酸序列和结构,以便与支架的单个模板位点的氨基酸结构互补,使得模板多核苷酸可经由其附接的多肽附接到支架的单个模板位点。上文提供了用于在支架的单个模板位点与模板多核苷酸之间进行共价或非共价结合的多肽对的示例,并且这些示例作为非限制性示例包括具有用于彼此形成卷曲螺旋附接的七肽重复序列的α螺旋氨基酸序列、生物素-抗生物素蛋白结合对、spytag/spycatcher系统、用于分选酶介导的转肽结合的lpxtg/低聚甘氨酸亲核对。在另一个示例中,可在样品制备期间修饰模板多核苷酸以包括snap标签序列或o
6-苄基鸟嘌呤中的一者,并且支架的单个模板位点可包括这两者中的另一者,以允许根据snap-标签/o
6-苄基鸟嘌呤系统在这两者之间进行共价结合。在另一个示例中,可在样品制备期间修饰模板多核苷酸以包括clip-标签序列或o
2-苄基胞嘧啶中的一者,并且支架的单个模板位点可包括这两者中的另一者,以允许根据clip-标签/o
2-苄基胞嘧啶系统和clip-标签/o
2-苄基胞嘧啶系统在这两者之间进行共价结合。
[0087]
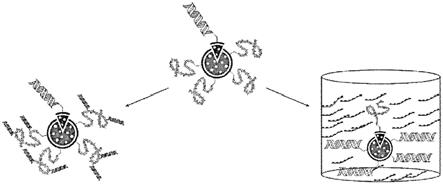
前述示例中的任一个同样可用于将一个或多个辅助物附接到支架上的一个或多
个辅助位点。为了附接到dna支架或多肽支架的辅助位点,辅助物(诸如辅助寡核苷酸-dna)可具有添加到其上的互补附接部分或结构。在一个示例中,核苷酸序列可包括在辅助物中或附接到辅助物,并且可包括附接到其上或包括在其中的互补附接部分或结构。
[0088]
例如,对于树枝状大分子dna支架,核苷酸序列可包括在辅助物中或附接到辅助物,并且该序列可与树枝状大分子dna支架的衔接子(诸如与树枝状大分子dna支架的最后一代衔接子的下游突出端,或与树枝状大分子的其他代衔接子的以其他方式未杂交的下游突出端)互补。
[0089]
在另一个示例中,辅助物(诸如辅助寡核苷酸-dna)可包括或附接到核苷酸序列,诸如在辅助寡核苷酸-dna的情况下从其末端中的一个延伸,并且该序列与其他序列包括在或附接到支架的辅助位点中的另一个序列互补。互补序列之间的沃森-克里克碱基配对使得这两者之间进行杂交和结合,因此将辅助物附接到辅助结合位点。在另一个示例中,辅助物可包括其共价修饰以允许与其互补的部分或结构共价附接到其上。例如,可包括对包括在模板多核苷酸中的核苷酸的修饰(诸如在磷酸基团、碱基或糖上)以提供与支架的辅助位点共价附接的位点。支架的辅助位点又可包括允许附接到辅助物(诸如寡核苷酸-dna辅助物)的互补部分或结构。在一个示例中,核苷酸经修饰以包括附接部分,支架的辅助结合位点的互补部分可通过该附接部分包括在添加到或包括在辅助物(诸如辅助寡核苷酸-dna)中以允许它们之间进行结合的多核苷酸序列中。可商购获得携带此类化学部分的许多修饰的核苷酸以用于将组合物共价附接到其中已掺入此类修饰的核苷酸的dna分子上。
[0090]
在另一个示例中,可通过附接到多肽来修饰辅助物。此类多肽可具有氨基酸序列和/或结构,以便与支架的辅助位点的氨基酸结构互补,使得辅助物可经由其附接的多肽附接到支架的辅助位点。上文提供了用于在辅助位点与辅助物之间进行共价或非共价结合的多肽对的示例,并且这些示例作为非限制性示例包括具有用于彼此形成卷曲螺旋附接的七肽重复序列的α螺旋氨基酸序列、生物素-抗生物素蛋白结合对、spytag/spycatcher系统、用于分选酶介导的转肽结合的lpxtg/低聚甘氨酸亲核对。在另一个示例中,可修饰辅助物(诸如辅助寡核苷酸-dna)以包括snap-标签序列或o
6-苄基鸟嘌呤中的一者,并且支架的辅助位点可包括这两者中的另一者,以允许根据snap-标签/o
6-苄基鸟嘌呤系统在这两者之间进行共价结合。在另一个示例中,辅助物可包括clip-标签序列或o
2-苄基胞嘧啶中的一者,并且支架的辅助位点可包括这两者中的另一者,以允许根据clip-标签/o
2-苄基胞嘧啶系统在这两者之间进行共价结合。
[0091]
模板多核苷酸与支架的模板位点的附接或者辅助物(诸如辅助寡核苷酸-dna)与支架的辅助位点的附接可通过它们之间的直接结合进行。在其他示例中,可以包括将dna支架的核苷酸或多肽支架的氨基酸连接到单个模板位点或辅助位点或两者的间隔物、聚合物或其他化学组合物。在一个示例中,可将用于在模板多核苷酸或辅助物之间进行结合的部分或结构掺入多肽支架的修饰的氨基酸或dna支架的修饰的核苷酸中,并且可将其直接结合到直接附接到模板多核苷酸或辅助物的互补部分或结构。在另一个示例中,间隔物、聚合物或其他化学组合物可从dna支架的核苷酸或多肽支架的氨基酸或两者延伸,并且用于结合模板多核苷酸或辅助物的部分或结构可存在于间隔物、聚合物或其他化学部分上,与所述间隔物、聚合物或其他化学部分与支架的附接有一定距离。在另一个示例中,间隔物、聚合物或其他化学组合物可从模板多核苷酸或辅助物或两者延伸,并且用于结合支架的部分
20,000范围内的整数,并且m为在1-100,000范围内的整数。
[0097]
在一些示例中,该丙烯酰胺单体可包括叠氮基乙酰胺基戊基丙烯酰胺单体:在一些示例中,丙烯酰胺单体可包括n-异丙基丙烯酰胺
[0098]
在一些方面,该杂聚物可包括以下结构:
[0099]
其中x为在1-20,000范围内的整数,并且y为在1-100,000范围内的整数,或其中y为1-20,000范围内的整数,并且x和z为整数,其中x和z的总和可在1至100,000的范围内,其中每个rz独立地为h或c
1-4
烷基,并且分别地,x:y的比率可为大约10:90至大约1:99,或可为大约5:95,或者(x:y):z的比率可为大约85:15至大约95:5,或可为大约90:10(其中x:(y:z)的比率可为大约1:(99)to至大约10:(90),或可为大约5:(95))。在这些示例中,约是指一个的相对量可与列出的比率中所述的量相差最多至5%。
[0100]“杂聚物”是至少两种不同的重复亚基(单体)组成的大分子。“丙烯酰胺单体”是具有结构或其取代的类似物(例如,甲基丙烯酰胺或n-异丙基丙烯酰胺)的单体。包括丙烯酰胺基团和叠氮基团的单体的示例是上文所示的叠氮基乙酰胺基戊基丙烯酰胺。“烷基”是指完全饱和(即,不包含双键和三键)的直链或支链烃链。示例性烷基基团包括甲
基、乙基、丙基、异丙基、丁基、异丁基和叔丁基。作为示例,名称“c
1-4
烷基”指示烷基链中存在一个至四个碳原子,即,该烷基链选自甲基、乙基、丙基、异丙基、正丁基、异丁基、仲丁基和叔丁基。
[0101]
前述聚合物中的任何一种或多种中的一种或多种可作为一个或多个辅助物附接到dna支架或多肽支架,或者用于将一个或多个辅助物附接到支架,诸如辅助寡核苷酸-dna分子。例如,支架可含有一个或多个炔基基团或者可与叠氮化物基团(诸如降冰片烯基团)反应和结合的一个或多个其他基团,并且聚合物的叠氮化物可经由环加成点击化学反应与炔烃、降冰片烯或支架的其他一个或多个基团共价结合。在另一个示例中,其他组合物或另外的辅助物(诸如其他组合物或结构,作为一个示例,包括寡核苷酸-dna分子)还可含有或被修饰成含有一个或多个炔基基团或者可与叠氮化物基团(诸如降冰片烯基团)反应和结合的一个或多个其他基团,并且聚合物的叠氮化物可经由环加成点击化学反应与炔烃、降冰片烯或此类其他化合物或另外的辅助物的其他一个或多个基团共价结合。因此,一种或多种聚合物可附接到支架,并且一种或多种此类附接的聚合物可进一步附接到一种或多种其他组合物,诸如另外的辅助物,诸如寡核苷酸-dna分子。在其他示例中,反应性化学可用于将聚合物附接到支架和辅助物,诸如将寡核苷酸-dna分子附接到聚合物。
[0102]
在一个示例中,单个模板多核苷酸可与支架的单个模板位点结合,并且多个辅助核苷酸(诸如辅助寡核苷酸-dna分子)可与支架的辅助位点结合(无论是直接结合还是经由如上文所公开的聚合物或者其他聚合物或间隔物或其他组合物)。此类寡核苷酸-dna分子的示例可以是用于在支架上进行聚类的引物。作为常规聚类过程的一部分,模板多核苷酸或其补体的拷贝在基底的表面上制成。如上所述,在一些情况下,此类表面上聚类可能会不利地导致形成一个或多个多克隆簇。如本文所公开的,聚类可诸如在溶液中的支架上进行,而无需将支架预先附接到表面。在其他示例中,附接的具有单个模板多核苷酸的支架可附接到基底的表面,然后可在基底的表面上、在支架上或在支架上和在基底的表面上进行聚类。
[0103]
对于聚类程序,可诸如在样品制备期间对模板多核苷酸进行修饰,以在其3-末端和5-末端中的一者或两者处包括一个或多个核苷酸序列。然后可在如本文所公开的支架上合成模板核苷酸和与模板核苷酸互补的核苷酸序列的一个或多个拷贝,从而形成簇。此类支架上聚类可导致形成单克隆簇。
[0104]
例如,模板多核苷酸可与单个模板附接位点结合,其5-末端朝向支架取向,并且其3-末端远离与支架结合的位点取向。3-末端可包括与第一引物中包含的核苷酸序列互补的核苷酸序列。“引物”被定义为充当dna或rna合成的起点的单链核酸序列(例如,单链dna或单链rna)。引物可以是任何数量的碱基长度,并且可以包括多种非天然核苷酸。在一个示例中,引物为短链,范围为20至40个碱基或10至20个碱基。与模板多核苷酸的3-末端互补的引物的拷贝可直接或通过附接到聚合物(作为非限制性示例,诸如pazam或本文所公开的相关聚合物)、间隔物或如本文所公开的其他化学组合物进一步附接到支架的辅助位点。
[0105]
然后可进行聚合反应,其中模板多核苷酸的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第一引物杂交。聚合反应中的聚合酶可产生附接到支架的模板多核苷酸的新生链补体,从与模板多核苷酸的3-末端杂交的支架附接的引物开始。然后可使模板多核苷酸及其补体去杂交。
[0106]
模板多核苷酸的补体在补体的3-末端可包括与第二引物序列互补的核苷酸序列。与模板多核苷酸的补体的3-末端互补的第二引物的拷贝可进一步附接到支架的辅助位点。然后可进行第二聚合反应,其中模板多核苷酸的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第一引物杂交,并且模板多核苷酸的补体的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第二引物杂交。第二聚合反应中的聚合酶可产生附接到支架的模板多核苷酸的另一新生链补体,从与模板多核苷酸的3-末端杂交的支架附接的第一引物开始。并且第二聚合反应中的聚合酶可进一步产生附接到支架的模板多核苷酸的新生链拷贝,从与在先前的聚合反应中聚合的模板的补体的3-末端杂交的支架附接的第二引物开始。然后可使模板多核苷酸和其拷贝及其补体去杂交。
[0107]
然后可在迭代过程中进行后续聚合反应。支架结合的模板多核苷酸和其拷贝的3-末端与和其互补的支架结合的第一引物杂交,并且模板多核苷酸的支架结合的补体的3-末端与和其互补的支架结合的第二引物杂交。新生链通过聚合酶聚合,在与支架结合的模板多核苷酸和其补体及其拷贝杂交的支架结合的第一引物和第二引物处开始。在聚合后使链去杂交后,进行连续的聚合反应,从而使模板多核苷酸的拷贝和其附接到支架的补体的数量成倍增加。以这种方式,模板多核苷酸的拷贝和补体得以扩增,其中扩增的拷贝与支架结合,从而形成簇。如本文所公开的,这种聚类过程可诸如在溶液中的支架上执行,这与常规聚类完全不同,常规聚类在常规sbs过程中在基底的表面上进行。因为只有聚集在支架上的单个模板多核苷酸的拷贝和补体,所以支架上存在单克隆簇。
[0108]
在此类示例中,当模板多核苷酸的5-末端处或附接到该处的序列与单个模板位点结合,使模板多核苷酸的3-末端远离支架取向时,模板多核苷酸可通过与附接到单个模板位点或其一部分的引物序列(称为模板位点引物)杂交而与支架的单个模板位点结合。在一个示例中,如通过样品制备过程制备的模板多核苷酸可在其5-末端具有与模板位点引物互补的核苷酸序列或使该核苷酸序列附接到该末端。对于与模板位点引物互补的此类核苷酸序列的3-末端,模板多核苷酸可包括与上述第二引物的核苷酸序列相对应的核苷酸序列(第二引物是可通过互补沃森-克里克碱基配对与模板多核苷酸的补体的3-末端杂交的支架附接的引物)。在模板多核苷酸中包括此类序列意味着在聚合步骤期间合成的模板多核苷酸的补体将朝向其3-末端具有与此类第二引物的序列互补的多核苷酸序列。朝向模板多核苷酸的补体的3-末端具有此类序列使得能够在聚类期间在后续聚合反应期间使补体的3-末端与此类第二引物杂交。
[0109]
在模板多核苷酸的3-末端,远离与单个模板位点结合的模板多核苷酸5-末端取向,模板多核苷酸可包括与如上所述的第一引物互补的序列。在第一聚合步骤期间,如上所述,模板多核苷酸3-末端处的此类核苷酸序列可与第一引物杂交,然后将新生补体聚合到模板多核苷酸。在模板多核苷酸的与模板位点引物杂交的部分与模板多核苷酸中包括第二引物的序列的位于其3-末端的核苷酸序列之间终止将补体的聚合到模板多核苷酸可能是有利的。也就是说,模板多核苷酸的补体在其3-末端具有与第二引物互补的序列可能是有利的。然而,如果在向模板多核苷酸的新生补体添加与对应于第二引物的序列互补的核苷酸序列之后没有终止聚合,那么模板多核苷酸的补体的3-末端将不会在那里结束。
[0110]
例如,如果与模板位点引物互补的核苷酸序列是与第二引物互补的序列的5-末端并与该序列连续,则模板多核苷酸的补体的3-末端可包括模板位点引物中包括的核苷酸序
列。例如,在将补体聚合到模板多核苷酸时,dna聚合酶可置换模板位点引物使其与模板多核苷酸的5-末端杂交,并将与其对应的核苷酸序列的添加聚合到模板多核苷酸的补体的3-末端。如果结果削弱了模板多核苷酸的补体的3-末端与上述第二引物在辅助位点处杂交的能力,则这种结果可能是不希望的。
[0111]
因此,在一个示例中,可能期望掺入终止将模板多核苷酸的3-末端聚合到5-末端,其中模板多核苷酸的此类5-末端通过与模板位点引物杂交与单个模板位点结合。例如,可以包括接头(诸如peg接头、烷基接头或其他化学部分)以将与模板位点引物杂交的核苷酸序列连接到模板多核苷酸的5-末端。此类接头而不是连续核苷酸序列连接部的存在将阻止聚合酶将与模板位点引物对应的核苷酸序列添加到模板多核苷酸的补体的3-末端,这将替代地如可能期望的以与第二引物的核苷酸序列互补的核苷酸序列结束。
[0112]
在其他示例中,模板多核苷酸可具有或附接到模板多核苷酸3-末端处的多核苷酸序列,该多核苷酸序列与作为支架的单个模板位点的一部分或附接到该单个模板位点的引物(称为模板位点引物)互补。在模板多核苷酸的3-末端的或附接到该末端的此类序列与模板位点引物杂交之后,可进行聚合过程,其中dna聚合酶聚合与模板多核苷酸互补的新生多核苷酸的形成,从模板位点引物开始。然后进行模板多核苷酸与模板多核苷酸的支架附接的补体去杂交。模板多核苷酸的支架附接的补体的3-末端(远离支架的附接位点取向)可包括与上述第二引物序列互补的核苷酸序列(第二引物是可通过互补沃森-克里克碱基配对与模板多核苷酸的补体的3-末端杂交的支架附接的引物)。与模板多核苷酸的补体的3-末端互补的第二引物的拷贝可进一步附接到支架的辅助位点。然后可进行第二聚合反应,其中模板多核苷酸的补体的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第二引物杂交。第二聚合反应中的聚合酶可产生模板多核苷酸的新生链拷贝(即模板多核苷酸的支架结合的补体的补体),从与在先前的聚合反应中聚合的模板的补体的3-末端杂交的支架附接的第二引物开始。然后可进行去杂交步骤,以使模板多核苷酸的支架结合的补体和模板多核苷酸的拷贝彼此去杂交。
[0113]
模板多核苷酸的拷贝在拷贝的3-末端可包括与上述第一引物序列互补的核苷酸序列。与上述模板多核苷酸的拷贝的3-末端互补的第一引物的拷贝可进一步附接到支架的辅助位点。然后可进行第三聚合反应,其中模板多核苷酸的拷贝的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第一引物杂交,并且模板多核苷酸的补体的3-末端经由沃森-克里克碱基配对与和其互补的支架结合的第二引物杂交。第三聚合反应中的聚合酶可产生附接到支架的模板多核苷酸的另一新生链补体,从与模板多核苷酸的拷贝的3-末端杂交的支架附接的第一引物开始。并且第三聚合反应中的聚合酶可进一步产生模板多核苷酸的新生链拷贝,从与在先前的聚合反应中聚合的模板的补体的3-末端杂交的支架附接的第二引物开始。然后可进行去杂交步骤,使模板多核苷酸的拷贝和补体彼此去杂交。
[0114]
然后可在迭代过程中进行后续聚合反应。模板多核苷酸的支架结合的拷贝的3-末端与和其互补的支架结合的第一引物杂交,并且模板多核苷酸的支架结合的补体的3-末端与和其互补的支架结合的第二引物杂交。新生链通过聚合酶聚合,在与支架结合的模板多核苷酸和其补体及其拷贝杂交的支架结合的第一引物和第二引物处开始。在聚合之后进行链的去杂交,然后进行连续聚合反应,随后进行进一步的去杂交。以这种方式,模板多核苷酸的拷贝和补体得以扩增,其中扩增的拷贝和补体与支架结合,从而形成簇。如本文所公开
的,这种聚类过程可诸如在溶液中的支架上执行,这与常规聚类完全不同,常规聚类在常规sbs过程中在基底的表面上进行。因为只有聚集在支架上的单个模板多核苷酸的拷贝和补体,所以支架上存在单克隆簇。
[0115]
在一个示例中,模板多核苷酸的末端包括或附接到与包括在支架的单个模板位点中或附接到支架的单个模板位点的核苷酸序列互补的核苷酸序列,称为第三模板位点引物。在一个示例中,模板多核苷酸的补体可在第三模板位点引物处起始的支架上合成。
[0116]
在如本文所公开的支架聚类的示例中,支架可以是如本文所公开的dna支架或多肽支架。模板多核苷酸可根据本文所公开的各种共价或非共价键中的任一种结合到支架的单个模板位点。例如,模板多核苷酸的任一端可包括诸如表1中所示的来自结合位点对的部分或结构,并且同一对的互补部分或结构可存在于支架的单个模板位点处。接连几轮的聚合随后可很像上面描述那样进行。例如,模板多核苷酸在模板多核苷酸5-末端处或朝向该处结合到支架的单个模板位点的3末端可与结合到支架的辅助位点的寡核苷酸引物和由dna聚合酶合成的其补体杂交。然后可如上所述进行连续几轮聚合,使模板多核苷酸的多个拷贝和从支架的辅助位点产生的其补体聚合。因为仅单个模板多核苷酸与支架结合,支架仅具有单个模板多核苷酸位点,所以此类拷贝将在支架上构成单克隆簇。
[0117]
在另一个示例中,支架可附接到基底的表面,诸如用于sbs程序中的基底的表面。例如,支架的辅助位点可包括或是或附接到与基底的表面或者与基底的表面结合的组合物附接的位点。在一个示例中,基底的表面可与引物(作为非限制性示例,诸如与如上所述的第一引物或第二引物或两者互补的引物的拷贝)结合。此类互补引物可直接附接到基底的表面,或者可附接到经修饰的表面,诸如聚合物分子(例如,pazam或相关聚合物)所附接的表面,其中引物附接到此类聚合物。前述第一引物和第二引物可附接到支架的辅助位点(直接或者经由聚合物(诸如上文作为非限制性示例所公开的pazam或其他pazam样聚合物)或间隔物或其他组合物)。支架或附接到支架的此类第一引物和第二引物可与与其互补的引物杂交,如附接到基底的表面,从而将支架结合到基底的表面。
[0118]
如上所述的第一引物和第二引物的示例可包括用于现有sbs过程的引物。合适的引物的具体示例包括p5和/或p7引物,其用于illumina,inc.销售的商业流通池的表面上,以用于在hiseq
tm
、hiseqx
tm
、miseq
tm
、miseqdx
tm
、miniseq
tm
、nextseq
tm
、nextseqdx
tm
、novaseq
tm
、genome analyzer
tm
、iseq
tm
和其他仪器平台上进行测序。并且模板多核苷酸的包括与如上文所公开的第一引物或第二引物对应或互补的核苷酸序列的部分可具有例如根据如在上述sbs平台或其他平台中使用的此类引物序列的与p5引物对应或互补的序列(包括aatgatacggcgaccaccgagatctacac(seq id no:7)的核苷酸序列)、p7引物(包括caagcagaagacggcatacgagat(seq id no:8)的核苷酸序列)或两者。
[0119]
作为非限制性示例,用于sbs过程的基底可包括在前述sbs平台或其他平台中的任一种中使用的基底。作为非限制性示例,这种基底可以是流通池。如本文所用,术语“流通池”旨在表示具有其中可以进行反应的室(即,流动通道)、用于将试剂递送到室的入口以及用于从室中移除试剂的出口的容器。在一些示例中,室使得能够检测在室中发生的反应或信号。例如,室可以包括允许对室中的阵列、光学标记分子等进行光学检测的一个或多个透明表面。如本文所用,“流动通道”或“流动通道区”可以是限定在两个粘结部件之间的区域,该区域可以选择性地接收液体样品。在一些示例中,流动通道可限定在图案化支撑件与盖
之间,因此可与限定在图案化支撑件中的一个或多个凹入部流体连通。在其他示例中,流动通道可限定在非图案化支撑件与盖之间。
[0120]
如本文所用,术语“凹入部”是指图案化支撑件中的离散凹面特征部,该离散凹面特征部具有完全被图案化支撑件表面的间隙区域包围的表面开口。凹入部可以在其表面中的开口处具有多种形状中的任一种,包括例如圆形、椭圆形、正方形、多边形、星形(具有任何数量的顶点)等。与该表面正交截取的凹入部的横截面可以为弯曲的、正方形、多边形、双曲线形、圆锥形、角形等。作为一个示例,凹入部可以是孔。同样如本文所用,“功能化凹入部”是指其中附接有引物(在一些示例中,通过聚合物(诸如pazam或类似聚合物)附接到凹入部的表面)的离散凹面特征部。
[0121]
术语流通池“支撑件”或“基底”是指可在其上添加表面化学物质的支撑件或基底。术语“图案化基底”是指凹入部限定在其中或在其上的支撑件。术语“非图案化基底”是指基本上平面的支撑件。基底在本文中也可称为“支撑件”、“图案化支撑件”或“非图案化支撑件”。支撑件可以是晶片、面板、矩形片材、管芯或任何其他合适的构型。支撑件通常为刚性的并且不溶于水性液体。支撑件可对用于修饰凹入部的化学物质呈惰性。例如,支撑件可以对用于形成聚合物涂层、将引物附接到诸如已沉积的聚合物涂层等的化学物质呈惰性。合适的支撑件的示例包括环氧硅氧烷、玻璃及改性或功能化的玻璃、多面体低聚倍半硅氧烷(poss)及其衍生物、塑料(包括丙烯酸、聚苯乙烯以及苯乙烯和其他材料的共聚物、聚丙烯、聚乙烯、聚丁烯、聚氨酯、聚四氟乙烯(诸如得自chemours的)、环烯烃/环烯烃聚合物(cop)(诸如得自zeon的)、聚酰亚胺等)、尼龙、陶瓷/陶瓷氧化物、二氧化硅、熔融的二氧化硅或基于二氧化硅的材料、硅酸铝、硅和改性的硅(例如,硼掺杂的p 硅)、氮化硅(si3n4)、氧化硅(sio2)、五氧化二钽(tao5)或其他氧化钽(tao
x
)、氧化铪(hao2)、碳、金属、无机玻璃等。支撑件还可以是玻璃或硅或硅基聚合物(诸如poss材料),任选地在表面处具有氧化钽或另一种陶瓷氧化物的涂层。poss材料可以是kejagoas等人,microelectronic engineering 86(2009)776-668中所公开的poss,该文献全文以引用方式并入。
[0122]
在一个示例中,凹入部可以是孔,使得图案化基底在其表面上包括孔阵列。孔可以是微孔或纳米孔。每个孔的尺寸可通过其体积、孔开口面积、深度和/或直径来表征。
[0123]
每个孔可以具有能够限制流体的任何体积。可以选择最小或最大体积,例如以适应流通池的下游使用所期望的通量(例如复用度)、分辨率、分析物组成或分析物反应性。例如,体积可以为至少约1
×
10-3
μm3、约1
×
10-2
μm3、约0.1μm3、约1μm3、约10μm3、约100μm3或更大。另选地或除此之外,体积可以为至多约1
×
104μm3、约1
×
103μm3、约100μm3、约10μm3、约1μm3、约0.1μm3或更小。
[0124]
每个孔开口在表面上所占据的面积可以基于与上文针对孔体积所述的标准类似的标准来选择。例如,每个孔开口在表面上的面积可以为至少约1
×
10-3
μm2、约1
×
10-2
μm2、约0.1μm2、约1μm2、约10μm2、约100μm2或更大。另选地或除此之外,面积可以为至多约1
×
103μm2、约100μm2、约10μm2、约1μm2、约0.1μm2、约1
×
10-2
μm2或更小。每个孔开口所占据的面积可以大于、小于上文指定的值或介于它们之间。
[0125]
每个孔的深度可以为至少约0.1μm、约1μm、约10μm、约100μm或更大。另选地或除此之外,深度可以为至多约1
×
103μm、约100μm、约10μm、约1μm、约0.1μm或更小。每个孔14'的
深度可以大于、小于上文指定的值或介于它们之间。
[0126]
在一些情况下,每个孔的直径可以为至少约50nm、约0.1μm、约0.5μm、约1μm、约10μm、约100μm或更大。另选地或除此之外,直径可以为至多约1
×
103μm、约100μm、约10μm、约1μm、约0.5μm、约0.1μm或更小(例如,约50nm)。直径可以为约150nm、约200nm、约250nm、约300nm、约350nm、约400nm、约450nm、约500nm、约550nm、约600nm、约650nm、约700nm、约750nm、约800nm、约900nm、约950nm、约1μm、约1.25μm、约1.5μm、约1.74μm、约2μm、约2.25μm、约2.5μm、约2.75μm、约3μm、约3.25μm、约3.5μm、约3.75μm、约4μm、约4.25μm、约4.5μm、约4.75μm、约5μm、约5.25μm、约5.5μm、约5.75μm、约6μm、约6.25μm、约6.5μm、约6.75μm、约7μm、约7.25μm、约7.5μm、约7.75μm、约8μm、约8.25μm、约8.5μm、约8.75μm、约9μm、约9.25μm、约9.5μm或约9.75μm。每个孔的直径可以大于、小于上文指定的值或介于它们之间。如本文所用的术语的纳米孔旨在表示具有最大直径为约1μm或更小的圆形开口的孔。
[0127]
应当理解,本文提供的范围包括规定范围和规定范围内的任何值或子范围。例如,约100nm至约1μm(1000nm)的范围应被解释为不仅包括明确列举的约100nm至约1μm的限值,而且还包括单个值(诸如约708nm、约945.5nm等)和子范围(诸如约425nm至约825nm、约550nm至约940nm等)。此外,当利用“约”和/或“基本上”来描述值时,它们意在涵盖相对于规定值的微小变化(高达 /-10%)。
[0128]
在一个示例中,纳米颗粒的大小可使得纳米颗粒在孔(诸如纳米孔)中的存在占据如此多的孔体积以致另一个纳米颗粒不能同时占据该孔。可参考基底表面中孔的已知大小来设计或确定纳米颗粒的大小,使得其可进入其中不存在其他纳米颗粒但其进入孔将由于先前进入孔并仍然存在于孔中的另一纳米颗粒的存在而被阻止的孔。大小被设定成一个孔不能容纳两个以上的纳米颗粒可有利于孔内簇的单克隆性。例如,在常规sbs过程中,可将模板多核苷酸以如下浓度引入溶液中被图案化成具有孔的流通池中:经校准以最大化模板多核苷酸将接种的孔的数量,(即,诸如直接或经由与模板核苷酸的部分的核苷酸序列互补的表面附接的聚合物与附接到孔的引物结合),但足够低以便尽可能最小化多克隆簇的形成。
[0129]
在一个示例中,流通池可包括不是凹入部或纳米孔而是模板多核苷酸或支架可结合或接种在其内的以其他方式在空间上隔离的区域(在本文中称为纳米垫)的纳米级区域。在一些示例中,流通池表面包括被表面中模板多核苷酸或支架可能不结合的区域彼此分离的纳米垫。纳米垫可彼此间隔开,以促进单克隆簇的形成。例如,纳米垫可彼此分离,使得在由单个模板多核苷酸接种的一个纳米垫内形成的簇将与仅由一个模板多核苷酸接种的另一个此类纳米垫充分分离。然而,可能难以防止多于一个模板多核苷酸接种纳米垫,从而形成一个或多个多克隆簇。在如本文所公开的一个示例中,纳米颗粒可通过防止多于一个模板多核苷酸接种或附接在给定纳米垫内来促进单克隆簇优于多克隆簇形成。例如,纳米颗粒的大小可使得在纳米垫上没有足够的空间用于结合多于一个纳米颗粒,其中模板多核苷酸结合到支架的单个模板多核苷酸位点。
[0130]
在一些情况下,如果具有彼此不同的核苷酸序列的两个或更多个模板多核苷酸结合或接种在彼此相同的孔内,则可能出现多克隆簇。分子可基于泊松分布在所施加的溶液内基于其浓度在孔之间分布,根据泊松分布,在最小化未占用的孔的数量(用于增加sbs运行的效率)与最小化被多个不同的模板多核苷酸占据的孔的数量之间存在平衡。在最小孔
大小与模板多核苷酸的大小之间的差异(例如,b-dna分子的直径可为大约2nm)可能导致在一方面可用或优选的在不利用那么多基底表面(诸如孔内的表面)的浓度之间选择,并且导致形成不期望或不期望的高数量的多克隆簇。
[0131]
如本文所公开的,模板多核苷酸可结合到纳米颗粒,每个纳米颗粒仅有一个模板多核苷酸结合。纳米颗粒的尺寸可被设定成允许纳米颗粒进入流通池的其中不存在另一纳米颗粒的孔中,但不允许其进入流通池的其中已经存在另一纳米颗粒的孔中。聚类(诸如单克隆聚类)可在纳米颗粒进入孔之前在纳米颗粒上发生,从而使得孔中存在单克隆簇。或者,模板多核苷酸可结合到纳米颗粒的模板位点,并且纳米颗粒可进入孔内并结合在其内(例如,通过辅助位点与表面结合或对孔的表面进行修饰),从而仅用单个纳米颗粒接种孔,然后可在孔内进行聚类,从而使得孔中存在单克隆簇。在一些示例中,在纳米颗粒进入孔之前可能会在纳米颗粒上发生一定程度的聚类,而在纳米颗粒进入孔之后可能会发生进一步的聚类。所有此类示例包括单克隆簇在孔内形成的示例。此外,调节纳米颗粒的大小以便减少、最小化或在一个示例中消除在孔中一次同时存在多于一个纳米颗粒可减少、最小化或在一个示例中消除多克隆簇的形成。
[0132]
纳米颗粒大小可通过改变支架的大小、改变与辅助位点结合的辅助物(诸如附接到其上的聚合物)的大小或这两种方法来调节。纳米颗粒的大小也可通过在纳米颗粒上发生或尚未发生的聚类的量来改变,例如通过改变纳米颗粒上模板多核苷酸的拷贝和补体可在聚类期间在多轮聚合过程中结合在其上的位点的数量,此类位点越少可能导致纳米颗粒大小的上限越低,而此类位点越多可能导致纳米颗粒大小的上限越大。聚类期间聚合的轮数也可改变纳米颗粒大小,轮数越多导致与纳米颗粒结合的模板多核苷酸的拷贝和补体越多,因此可能增加其大小上限,而轮数越少导致与纳米颗粒结合的模板多核苷酸的拷贝和补体越少,因此可能降低其大小上限。纳米颗粒的大小可根据其在支架上发生聚类之前或在支架上发生聚类之后的大小来确定。
[0133]
如本文所用,术语“纳米颗粒”旨在表示最大尺寸的大小最高达到约1,000nm的颗粒。根据几何形状,尺寸可指长度、宽度、高度、直径等。尽管“直径”通常用于描述如本文的一个示例的尺寸,但是本文所述的纳米颗粒不需要是球形的或圆形的。如本文所公开的纳米颗粒的直径可为约2nm、约5nm、约7nm、约10nm、约12nm、约15nm、约17nm、约20nm、约22nm、约25nm、约27nm、约30nm、约32nm、约35nm、约40nm、约42nm、约45nm、约47nm、约50nm、约52nm、约55nm、约57nm、约60nm、约62nm、约65nm、约67nm、约70nm、约72nm、约75nm、约77nm、约80nm、约82nm、约85nm、约87nm、约90nm、约92nm、约95nm、约97nm、约100nm、约125nm、约150nm、约175nm、约200nm、约225nm、约250nm、约275nm、约300nm、约325nm、约350nm、约375nm、约400nm、约425nm、约450nm、约475nm、约500nm、约525nm、约550nm、约575nm、约600nm、约625nm、约650nm、约675nm、约700nm、约725nm、约750nm、约775nm、约800nm、约825nm、约850nm、约875nm、约900nm、约925nm、约950nm、约975nm或约1,000nm。纳米颗粒的直径通过动态光散射(dls)(也称为准弹性光散射)测量,表示为流体动力学半径(rh)的两倍,流体动力学半径可在dls系统或包括dls和其他功能的其他系统(例如,malvern instruments limited)上测定。
[0134]
如本文所公开的纳米颗粒的直径可在约2nm至约10nm、约5nm至约15nm、约7nm至约20nm、约10nm至约25nm、约15nm至约30nm、约20nm至约50nm、约40nm至约60nm、约50nm至约
75nm、约60nm至约100nm、约70nm至约100nm、约75nm至约100nm、约80nm至约110nm、约90nm至约130nm、约100nm至约150nm、约100nm至约200nm、约150nm至约225nm、约200nm至约250nm、约200nm至约300nm、约225nm至约275nm、约250nm至约300nm、约275nm至约325nm、约300nm至约400nm、约300nm至约350nm、约325nm至约375nm、约350nm至约400nm、约375nm至约425nm、约400nm至约500nm、约400nm至约450nm、约425nm至约475nm、约450nm至约500nm、约475nm至约525nm、约500nm至约600nm、约500nm至约550nm、约525nm至约575nm、约550nm至约600nm、约575nm至约625nm、约600nm至约700nm、约600nm至约625nm、约625nm至约675nm、约650nm至约700nm、约675nm至约725nm、约700nm至约800nm、约700nm至约725nm、约725nm至约775nm、约750nm至约800nm、约775nm至约825nm、约800nm至约900nm、约800nm至约850nm、约825nm至约875nm、约850nm至约900nm、约875nm至约925nm、约900nm至约1,000nm、约900nm至约950nm、约925nm至约975nm、约950nm至约1,000nm、约300nm至约450nm、约350nm至约500nm、约400nm至约550nm、约450nm至约600nm、约500nm至约650nm、约550nm至约700nm、约600nm至约750nm、约650nm至约800nm、约700nm至约850nm、约750nm至约900nm、约800nm至约950或约850nm至约1,000nm的范围内。
[0135]
非限制性示例
[0136]
以下示例旨在示出本公开的特定示例,但绝不旨在限制其范围。
[0137]
图1示出了如本文所公开的纳米颗粒的非限制性示例的图示。在该非限制性示例中,单个模板多核苷酸位点被示出为如所公开的纳米颗粒的支架部分的楔形部分。单个模板多核苷酸被示出为与单个模板位点结合。同样在该非限制性示例中,多个辅助物被示出为从支架的辅助位点延伸。在中心图示中,辅助物被示出为聚合物。在左图中,示出了与模板多核苷酸互补的多核苷酸的许多拷贝和模板多核苷酸的拷贝,其中此类拷贝附接到支架并从支架延伸。在该示例中,它们从聚合物延伸,所述聚合物又从支架延伸。
[0138]
在右图中,在基底的孔中示出了在单个模板位点处结合有模板多核苷酸的纳米颗粒。多个辅助寡核苷酸被示出为从支架延伸。尽管在右侧图中未示出,但在该示例中,辅助寡核苷酸从附接到支架的聚合物延伸。在其他示例中,辅助寡核苷酸可直接从支架延伸而它们之间不存在居间聚合物。辅助寡核苷酸的核苷酸序列与附接到孔表面的引物互补。辅助寡核苷酸由此与孔附接的引物杂交并附接到孔的表面。此处,由于纳米颗粒的大小相对于孔的大小,孔中一次只能存在一个纳米颗粒。因此,在孔中从单个模板多核苷酸开始的聚类将导致在孔内形成单克隆簇。
[0139]
图2a示出了dna支架的非限制性工作示例。在该非限制性示例中,dna支架包括多个支架dna分子,其中多个支架dna分子形成dna树枝状大分子。dna树枝状大分子包括多代分叉组成重复单元,也称为衔接子。第一代被示出在第二代的上游。第一代的衔接子被示出在底部,并且第二代的衔接子被示出在上面。每个衔接子包括组成重复单元寡聚脱氧核苷酸,这些组成重复单元寡聚脱氧核苷酸彼此杂交而形成包括一个上游突出端和两个下游突出端的衔接子。对于每代的衔接子,三个寡核苷酸-dna分子如图所示彼此杂交(此处,在50nm nacl中从90c冷却至20c),以形成具有上游突出端和两个下游突出端的衔接子。
[0140]
在该示例中,将第一代衔接子的上游突出端确定为捕获位点,意指用于将单个模板多核苷酸结合到支架的单个模板位点。第一代衔接子1的下游突出端具有与第二代衔接子1'的上游突出端互补并可与其杂交的核苷酸序列。然后,第一代衔接子和第二代衔接子
彼此杂交,使得第二代衔接子1'的上游突出端由于沃森-克里克碱基配对杂交附接到第一代衔接子1的下游突出端。然后在存在t4 dna连接酶、1mm atp和10mm mgcl2的情况下,在室温下将序列连接在一起(在该示例中连接10分钟)。可将亚序列代添加或连接到如图所示的该结构,其中添加的第n'代的衔接子的下游突出端与紧邻的前一代即第n 1代的衔接子的下游突出端互补。在一个示例中,可将2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、25、30、35、40、45、50、60、70、80、90、100或更多代形成为树枝状大分子dna支架的一部分。可部分地通过控制支架中包含的代数来控制dna树枝状大分子支架的大小(诸如直径),其中相对于包括更少代的支架,更多代对应更大的纳米颗粒。
[0141]
在一个非限制性示例中,第一代衔接子(g1)由以下寡核苷酸序列(5-末端到3-末端)合成:
[0142]
g1a:gaatgccgcttacagtacgcctaggtcagt(seq id no:9);
[0143]
g1b:tccgactaagccagtaagcggcattccagt(seq id no:10);以及
[0144]
g1c:acctaggcgtacttggcttagtcggattttttttttgtgtagatctcggtggtcgccgtatcatt(seq id no 11);其中g1a和g1b的下划线部分表示下游突出端,并且g1c的下划线部分表示上游突出端。对于此类示例,g1c的上游突出端可以包括单个模板核苷酸位点。例如,模板多核苷酸可以从其5-末端延伸以下序列(5-末端到3-末端)aatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct(seq id no:12)。该序列的5-末端与g1c的3-末端上游突出端杂交以形成图2b所示的非限制性工作示例中所示的结构。
[0145]
第二代衔接子(g2)的一个非限制性示例由以下寡核苷酸序列(5-末端到3-末端)合成:
[0146]
g2a:gaatgccgcttacagtacgcctaggtactg(seq id no:13);
[0147]
g2b:tccgactaagccagtaagcggcattccgat(seq id no:14);以及
[0148]
g2c:acctaggcgtacttggcttagtcggacgat(seq id no:15);其中g2b和g2c的下划线部分表示下游突出端,并且g2a的下划线部分表示上游突出端。对于此类示例,两个g2a序列中的每一个的actg上游突出端与g1a和g1b的下游catg突出端杂交以形成图2c所示的非限制性工作示例中所示的结构。
[0149]
第三代衔接子(g3)的一个非限制性示例由以下寡核苷酸序列(5-末端到3-末端)合成:
[0150]
g3a:gaatgccgcttacagtacgcctaggtatcg(seq id no:16);
[0151]
g3b:tccgactaagccagtaagcggcattcgcat(seq id no:17);以及
[0152]
g3c:acctaggcgtacttggcttagtcggagcat(seq id no:18);其中g3b和g3c的下划线部分表示下游突出端,并且g3a的下划线部分表示上游突出端。这些寡核苷酸序列杂交在一起以形成图2d所示的非限制性工作示例中所示的结构。两个g3a序列中的每一个的上游atcg突出端与g2b和g2c的下游cgat突出端杂交。
[0153]
第四代衔接子(g4)的一个非限制性示例由以下寡核苷酸序列(5-末端到3-末端)合成:
[0154]
g4a:gaatgccgcttacagtacgcctaggtatgc(seq id no:19);g4b:tccgactaagccagtaagcggcattcttgc(seq id no:20);以及
[0155]
g4c:acctaggcgtacttggcttagtcggattgc(seq id no:21);其中g4b和g4c的下划线
部分表示下游突出端,并且g4a的下划线部分表示上游突出端。这些寡核苷酸序列杂交在一起以形成图2e所示的非限制性工作示例中所示的结构。对于此类示例,两个g4a序列中的每一个的上游atgc突出端与g3b和g3c的下游gcat突出端杂交。
[0156]
第五代衔接子(g5)的一个非限制性示例由以下寡核苷酸序列(5-末端到3-末端)合成:
[0157]
g5a:gaatgccgcttacagtacgcctaggtgcaa(seq id no:22);
[0158]
g5b:tccgactaagccagtaagcggcattcggat(seq id no:23);以及
[0159]
g5c:acctaggcgtacttggcttagtcggaggat(seq id no:24);其中g5b和g5c的下划线部分表示下游突出端,并且g5a的下划线部分表示上游突出端。这些寡核苷酸序列杂交在一起以形成图2f所示的非限制性工作示例中所示的结构。对于此类示例,两个g5a序列中的每一个的上游gcaa突出端与g4b和g4c的下游ttgc突出端杂交。在该非限制性示例中,g5c的3-末端附接到荧光团(alexa fluorthermo fisher scientific)以允许通过荧光成像对纳米颗粒进行可视化。
[0160]
在另一个示例中,可包括如上所示的荧光团与g5c寡核苷酸的3-末端之间的多核苷酸或其他间隔物(例如,具有cctcctcctcctcctcctcctcct(seq id no:25)的非限制性示例核苷酸序列)。
[0161]
对于具有多于五代的树枝化基元,可使用寡核苷酸g5a、g2b和g2c来合成第五代(以及此后相关的每一个第三代)的衔接子,后续两代的衔接子由分别具有第三代和第四代的衔接子的序列的寡核苷酸制成。
[0162]
可通过连续组装给定代的衔接子、使其与前一代杂交(对于第二代或更高代的衔接子)并将寡核苷酸的末端连接在一起(其中它们在一代的上游衔接子与下一代的下游衔接子之间的边界处发生粘性末端杂交时相遇)来从一代到下一代构建树枝状大分子dna支架。
[0163]
组装衔接子和dna树枝状大分子的非限制性示例如下。为了产生衔接子,使用来自上述非限制性示例的每一代衔接子的相关序列,将用于组装衔接子的三种寡核苷酸悬浮在组装缓冲液(10mm tris,ph 8.0;1mm edta;50mm nacl)中,浓度为200μm。然后将10μl每种溶液与20μl组装缓冲液组合。将该组合在95℃下变性2分钟,在65℃下冷却2分钟,然后在60度下退火6分钟。然后进行三十九个退火步骤,每个步骤30秒,温度降低0.1℃(从59.1℃开始)。
[0164]
为了将第二代衔接子附接到第一代,制备150μl溶液,其中包括t4dna连接酶(neb m0202m)和15μl 10x连接酶缓冲液,并用含有0.5μm第1代衔接子和2μm第2代衔接子的组装缓冲液使总体积达到150μl。然后将100μl该反应与450μl组装缓冲液组合,然后转移到50kda mwco过滤器中。然后将样品在15,000
×
g下离心1分钟,重复过滤10次,每个离心步骤添加400μl组装缓冲液。再添加400μl组装缓冲液后,通过将过滤器倒置放置在新管中并在5,000
×
g下离心3分钟来从过滤器中洗脱出树枝状大分子。然后通过添加组装缓冲液使体积达到100μl。
[0165]
然后将第三代树枝状大分子的衔接子以4μm至0.5μm的比例添加到第1代至第2代树枝状大分子的溶液中,其中用组装缓冲液使t4 dna连接酶和连接酶缓冲液的体积达到大约60μl。将第4代衔接子以8μm至0.5μm的比例添加到第1代至第3代树枝状大分子的溶液中,
其中用组装缓冲液使t4 dna连接酶和连接酶缓冲液的体积达到大约63μl。将第5代衔接子以15μm至0.5μm的比例添加到第1代至第4代树枝状大分子的溶液中,其中用组装缓冲液使t4 dna连接酶和连接酶缓冲液的体积达到大约69μl。将第6代衔接子以22.5μm至0.5μm的比例添加到第1代至第5代树枝状大分子的溶液中,其中用组装缓冲液使t4 dna连接酶和连接酶缓冲液的体积达到大约75μl。将第6代衔接子以22.5μm至0.25μm的比例添加到第1代至第5代树枝状大分子的溶液中,其中用组装缓冲液使t4 dna连接酶和连接酶缓冲液的体积达到大约75μl。
[0166]
在一个示例中,dna树枝状大分子支架的大小可根据其包括的衔接子的代数来确定。例如,如上所述合成具有2至9代的树枝化基元dna支架,并且通过dls测量其直径。结果在表2中示出:
[0167]
表2:根据代数确定的树枝化基元dna支架的直径
[0168][0169]
图3示出了ssdna支架的非限制性示例。在该非限制性示例中,示出了合成ssdna支架的不同方式。单个模板位点存在于ssdna支架的末端上。四角星指示支架上的辅助位点。p5和p7表示辅助寡核苷酸,并且五角星表示与辅助位点互补的部分或结构。在左上方所示的示例中,在滚环扩增过程中,将环状dna模板用作通过dna聚合酶合成ssdna分子的模板。通过例如链置换dna聚合酶(例如,phi29)复制链可产生ssdna分子,该分子包括环状编码链的复制链的串联拷贝。可部分地通过控制环状模板的大小和滚环扩增过程的持续时间(在滚环扩增过程中聚合的持续时间越长,产生的ssdna支架越长)来确定ssdna支架的大小。
[0170]
在中上方所示的另一个非限制性示例包括通过使用不依赖模板的聚合酶(例如,末端脱氧核苷酸转移酶或tdt)来合成ssdna模板。不依赖模板的聚合酶(诸如tdt)在单链dna链的3-末端-羟基末端处掺入脱氧核苷酸,而不需要或复制模板。可通过修改合成支架的聚合过程的持续时间来控制通过使用不依赖模板的聚合酶合成的ssdna的大小。
[0171]
用于合成ssdna支架的方法的另一个非限制性示例在右上方示出。在该示例中,通过任何期望的方法合成若干单链dna分子。在一个示例中,在流失聚合过程(其中聚合酶沿着编码链行进,诸如从线性化质粒合成与其互补的新生链,直到它到达线性编码链的末端)中合成ssdna分子。在到达末端后,聚合酶从编码链的末端流失,就完成ssdna分子的任何合成。可合成多种ssdna产物,然后将它们端到端连接以形成包括多种产物中的每一种的单个ssdna支架。在一个示例中,一种ssdna产物与另一种ssdna产物的连接可借助于夹板来实现,如图所示。例如,可设计短寡核苷酸-dna,其3-末端与一种ssdna产物的5-末端互补,并且其5-末端与另一种ssdna产物的3-末端互补,使得dna-寡核苷酸与这两种ssdna产物的杂交将一种产物的5-末端与另一种产物的3-末端一起放在带切口的双链结构中,在该带切口
的双链结构中它们与dna-寡核苷酸杂交。然后可使用dna连接酶(例如,t4)来将两端酶促连接在一起,以由这两种产物形成单个ssdna分子。dna-寡核苷酸可包括另外的反应,用于夹板辅助将这种第一反应的产物的一个或两个末端连接到另一种ssdna产物,诸如此类,以构建可能需要的ssdna支架。以这种方式制成的ssdna的大小可通过控制连接在一起以形成ssdna支架的ssdna分子的数量和大小来控制。这些示例不是详尽的。它们也不是相互排斥的,因为多于一个或所有三个可一起用于ssdna支架的合成。
[0172]
在该非限制性示例中,辅助寡核苷酸被示出为与ssdna支架结合。辅助寡核苷酸-dna可通过本文所公开的用于这样做的各种方法中的任一种结合到辅助位点。然后,模板的5-末端被示出为通过非共价沃森-克里克碱基配对杂交在支架的互补3-末端处与单个模板位点结合。然后对支架执行聚类过程。模板多核苷酸的未与ssdna支架杂交的部分的末端含有与p5和p7辅助寡核苷酸对应或互补的序列。在多轮聚合之后,可以看到模板多核苷酸的支架结合的补体和模板多核苷酸的支架结合的拷贝已从p5和p7辅助寡核苷酸的5-末端延伸。在该示例中,第一聚合没有置换模板多核苷酸的5-末端使其与支架的3-末端杂交。因此,与模板多核苷酸的5-末端的以下部分互补的序列不包括在合成的模板多核苷酸的支架结合的补体中:与ssdna支架的杂交末端互补。
[0173]
用于将模板多核苷酸或辅助物(诸如辅助寡核苷酸)结合到如本文所公开的dna支架的上文所公开的部分或结构中的任一种可用于将模板多核苷酸或辅助物结合到支架上。在一些示例中,可商购获得的携带此类部分或结构(包括叠氮化物基团、炔烃基团、环辛炔基团、生物素基团或硫醇基团)并且能够掺入新生dna链中的核苷酸可包括在dna支架中。例如,在合成dna支架的聚合酶反应中,可相对于存在非修饰核苷酸的浓度以选定浓度将修饰的核苷酸接种到聚合反应中。根据此类浓度,掺入dna支架中的一定百分比的核苷酸将是修饰的核苷酸。可将多于一种类型的修饰的核苷酸接种到反应中,以包括多于一种类型的用于通过具有与其互补的部分或结构的组合物结合到dna支架的部分或结构。通过将修饰的核苷酸掺入dna支架中,或修饰的核苷酸能够被进一步修饰以向其添加用于在支架、辅助物或模板多核苷酸之间结合的如本文所公开的部分或结构,单个模板位点和辅助位点可包括在dna支架中。
[0174]
在一个示例中,核苷酸可经修饰以便包括与另一个核苷酸(诸如其所连接的多核苷酸的核苷酸)的接头,诸如聚乙二醇或其他接头。在另一个示例中,核苷酸可经修饰以便包括与氨基酸(诸如其所连接的多肽的氨基酸)接头,诸如聚乙二醇或其他接头。此类连接到多核苷酸或连接到多肽可以是模板多核苷酸或辅助物的结合位点,诸如通过本文所公开的非共价结合的示例。
[0175]
图4示出了多肽支架的非限制性工作示例的图示。在该示例中,使用具有图4中所示的氨基酸序列和构象的绿色荧光蛋白(gfp)。gfp含有三个半胱氨酸残基。当gfp采用如图4所示的三维结构时,半胱氨酸中的仅一个(在图4中呈现的序列中表示为c137)从如图4所示的分子面向外暴露。c125和c195的半胱氨酸残基可掩埋在gfp的三维构象内,而不是暴露在如图所示的结构外部,因此可能无法用于键形成。当仅单个半胱氨酸残基(例如,图4中所描绘的结构的面向外的c137)可用于键形成时,硫醇基团可用作gfp蛋白支架的单个模板多核苷酸位点。
[0176]
在两个其他示例中,通过标准重组方法将c125和c195在一个示例中均突变为丙氨
酸,并且在另一个示例中均突变为缬氨酸,仅在c137处留下单个硫醇位点作为支架模板核苷酸结合位点。具有其硫醇基团的此类单个半胱氨酸残基可以是单个模板核苷酸位点,因为gfp蛋白支架缺乏其他硫醇基团,仅具有一个硫醇基团,并且可以与作为与其互补的部分或结构的此类硫醇基团形成键的部分或结构的几种可能性可用于将模板多核苷酸结合到其上。gfp蛋白支架还可包括许多赖氨酸残基(例如,如图4所示的序列中所示的19)。赖氨酸残基包括其侧链的胺基团。因此,gfp支架中的赖氨酸残基的胺基团可用作辅助位点,并且含有可以与此类胺基团结合的部分或结构的辅助物(诸如寡核苷酸-dna分子)或聚合物可存在于此类辅助物上以用于与赖氨酸胺基团辅助位点结合。
[0177]
在另一个示例中,可将多肽支架诸如gfp的胺基团有效地转化为其他附接位点。在一个示例中,在一端具有nhs-酯而在另一端具有叠氮化物基团(被peg
24
序列分开)的双功能接头附接到gfp多肽支架的胺位点。双功能接头的nhs-酯末端与gfp多肽支架的胺基团结合,使叠氮化物基团作为辅助结合位点暴露出来。这些添加使得通过凝胶电泳测量的gfp多肽支架的大小增加大约20kda,这与添加20个双功能接头(每个大小为1157da)一致,一个接头添加到19个赖氨酸残基的胺基团中的每一个,一个接头添加到gfp多蛋白支架的n末端。在其他示例中,不同的双功能接头可以用于有效地替换硫醇位点,或有效地用不同部分或结构替换胺基团或硫醇基团。
[0178]
图5是用于将模板多核苷酸结合到支架的不同方法的图示。在顶部示出了具有单个模板多核苷酸位点的支架。在左侧,模板位点引物包括在单个模板多核苷酸位点中,并且其末端与模板多核苷酸的末端互补,以允许模板多核苷酸通过沃森-克里克碱基对杂交非共价结合到支架。在中间,模板多核苷酸和单个模板多核苷酸位点分别具有互补部分或结构,使得在模板多核苷酸与支架之间形成共价键。在右侧,多肽包括在单个模板位点中,并且与其互补的多肽附接到模板多核苷酸的末端。单个模板位点的多肽与模板多核苷酸之间的非共价结合将模板多核苷酸结合到支架。
[0179]
图6示出了其中模板多核苷酸通过与从支架的单个模板位点延伸的引物杂交而与蛋白质支架结合,然后通过聚合酶形成模板多核苷酸的支架结合的补体来延伸引物的非限制性工作示例。马来酰亚胺部分附接到p5寡核苷酸的5-末端。p5寡核苷酸通过寡核苷酸的马来酰亚胺基团与gfp支架的可接近半胱氨酸残基的硫醇基团之间的硫醇-马来酰亚胺键附接到gfp多肽支架的单个可接近模板位点。左侧的聚丙烯酰胺凝胶电泳(page)印迹中由箭头(标记)指示的迁移条带示出了支架与p5寡核苷酸之间的硫醇-马来酰亚胺结合。3-末端与p5寡核苷酸互补的模板多核苷酸通过与p5寡核苷酸的非共价杂交结合到支架。然后进行聚合反应以形成支架结合,以形成模板多核苷酸的支架结合的补体。
[0180]
右侧的page印迹示出了在三种不同条件下运行的聚合反应的结果。在条件a中,p5寡核苷酸通过硫醇-马来酰亚胺键结合到支架。列a中的箭头(第1链)指示的条带表示在聚合期间在支架上形成模板多核苷酸的支架结合的补体。在列b中,通过在p5寡核苷酸的3-末端核苷酸上的阻断位置中附接cy5荧光团,防止其被聚合酶延伸,来防止p5寡核苷酸的延伸。列b中的箭头表示未形成模板多核苷酸的基于支架的补体,证实了列a中所示的阳性结果。在列c中,cy5经由六胸苷(t6)结合到p5寡核苷酸,没有延伸阻断。列c中的箭头与列a中的箭头相匹配,再次证实形成了模板多核苷酸的支架结合的补体,并且其在列b中不存在并不是仅仅是由于存在cy5而导致的假阴性结果。
[0181]
图7示出了包括(plp)介导的对蛋白质的n-末端特异的转氨作用的生物缀合的非限制性示例。该反应将n-末端胺氧化成酮或醛,然后酮或醛与烷氧基胺形成稳定的肟键。
[0182]
图8示出了可以与天然氨基酸形成键以结合到支架的辅助位点的非限制性示例,如此处所示或也针对单个模板位点。示例包括硫醇-马来酰亚胺与半胱氨酸残基偶联、胺-nhs在赖氨酸残基处结合、铑卡宾体与色氨酸残基结合、α、β-二羰基与精氨酸残基结合以及plp介导的蛋白质n末端的转氨作用然后形成肟键。右侧示出了修饰的非天然存在的氨基酸残基处蛋白质的位点特异性修饰的示例。一个示例示出了与羟胺反应以形成肟的非天然酮氨基酸。第二反应示出了与四嗪反应的非天然降冰片烯(或其他应变烯烃/炔烃)。
[0183]
图9示出了通过与由px(gfp-寡核苷酸缀合物)表示的模板位点引物杂交而结合到蛋白质支架的单个模板位点的模板多核苷酸的示例。示出了文库的模板多核苷酸的两个示例,一个是具有p5和p7序列的标准文库分子,其中在3-末端具有与px模板位点引物(命名为px
′
)互补的区域,或其中px'序列通过peg接头与p5序列分离的修饰版本。醚链结合到支架的单个模板位点。标准文库序列可以用于第一链延伸聚合反应中,px引物用作起始引物以聚合与模板多核苷酸互补的新生链。
[0184]
图10示出了非共价键、特别是卷曲螺旋肽非共价键(在一个示例中,kd在《1
×
10-10
m的皮摩尔范围内)的示例。示出了α螺旋多肽结构的形成卷曲螺旋附接的两个互补结合配偶体的两个氨基酸序列。通过将一个此类序列附接到支架(诸如图10所示的多肽支架(gfp-肽融合物))并将另一个序列附接到模板多核苷酸(肽-寡核苷酸缀合物)或文库模板多核苷酸,模板多核苷酸可以经由α螺旋之间的非共价结合与支架结合。在另一个示例中,α螺旋序列中与附接到支架的序列互补的一个可以附接到辅助物,诸如用于将辅助寡核苷酸附接到辅助位点的辅助寡核苷酸。
[0185]
图11示出了使用活化的nhs-酯的多肽支架的赖氨酸侧链的选择性生物缀合。赖氨酸与二苯并环辛炔(dbco)结合。dbco的炔烃部分随后可以经由应变促进的[3 2]环加成点击反应附加含叠氮化物的分子。作为非限制性示例,随后可以附加到dbco基序的分子包括寡核苷酸和聚合物。gfp多肽支架很容易用dbco标记,从而产生dbco标记的支架的混合物。通过对反应产物进行sds page凝胶分析未检测到未标记的gfp(未示出)。
[0186]
图12示出了具有附接到其的单个模板多核苷酸的支架(附接到流通池的孔的表面)的非限制性工作示例的示例图示。在该非限制性示例中,支架是dna树枝状大分子。单个模板位点从第一代衔接子的上游突出端延伸,并且辅助位点从最后一代衔接子的下游突出端延伸。图13示出了可以存在于给定纳米孔表面积(sa)或直径(d)的纳米孔(树枝状大分子/纳米孔)中的给定大小的支架的数量的测试的非限制性工作示例的图(接种事件与纳米孔表面积)。将直径为约100nm的dna树枝状大分子纳米颗粒接种到直径为185、285或375nm的纳米孔中,并且测量每个纳米孔的树枝状大分子的数量。从结果的最佳拟合曲线(y=3e-0.5x-3.4874,r2=0.9991)外推表明,在该示例中,使用直径为约100nm的纳米颗粒和直径为约100nm的纳米孔将使得纳米孔接种单个纳米颗粒。
[0187]
图14a-图14d示出了根据本公开的各方面的使用dna支架用模板多核苷酸接种基底的示例。图14a示出了根据本公开的包括dna树枝状大分子的支架dna分子的描绘。支架的直径为50nm-150nm。支架包括用于结合模板多核苷酸和多个辅助位点(cpx)的单个模板位点(pa)。图14b示出了模板多核苷酸及其补体,其中引物序列添加到每个末端(p5/cp5和p7/
cp7)。模板多核苷酸的p5-引物末端通过peg接头连接到引物(cpa)。cpa引物与图14a中所描绘的支架dna分子的单个模板位点(pa)互补。图14c是图14a中所描绘的支架dna分子的描绘,该支架dna分子经由其单个模板位点(pa)与模板多核苷酸杂交,并且经由cpa引物与其在图4b中所描绘的补体杂交。支架附接到基底。基底附接到与支架的辅助位点(cpx)互补的引物(px)。基底也附接到引物,以允许模板末端与其杂交,从而允许在基底上聚类
[0188]
图14d描绘了根据前述内容的示例,其展示了使用具有单个模板位点的支架用模板多核苷酸接种基底然后聚类。根据本公开的附接到模板多核苷酸的支架在图14a-图14c中示出。以所示的摩尔比将树枝状大分子支架与模板多核苷酸(文库)组合,用其接种基底(具有用于接种的纳米孔的流通池),然后根据重组酶驱动的簇扩增过程(examp簇扩增)进行聚类。阴性对照包括没有模板的支架和没有支架的模板。作为阳性对照( 对照),在模板分子与未经由支架附接到基底的引物杂交之后,在基底上使用聚类来在没有树枝化基元的情况下在基底上进行聚类。
[0189]
左图是根据上述条件的聚类过程之后的流通池的图像(2个阴性对照、各种支架:模板摩尔比的5个条件和1个阳性对照)。除阴性对照之外的所有条件下的荧光指示具有单个模板结合位点的支架可以用模板多核苷酸接种基底并支持聚类过程。柱形图是8个条件的聚类结果的定量测量。上图c1强度是作为簇大小或产率的间接量度的循环1强度(强度与簇大小或产率成正比)。下图%pf是%通过过滤器,其是通过阈值过滤器的纳米孔的百分比,表示在其中形成(即与具有单克隆簇的纳米孔的数量成正比)的簇的纯度。
[0190]
应当理解,前述概念和本文更详细讨论的附加概念(假设此类概念不相互矛盾)的所有组合都被设想为是本文所公开的发明主题的一部分。具体地,出现在本公开末尾的要求保护的主题的所有组合都被设想为是本文所公开的发明主题的一部分,并且可用于实现本文所述的益处和优点。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。